
Abstract
CD200 is widely distributed in the central nervous system and plays an essential role in the immune response in neurological diseases. However, little is currently known about the effects of CD200 signaling on the blood–brain barrier (BBB) function after intracerebral hemorrhage (ICH). In this study, the role of CD200 during ICH in an autologous blood induced mouse ICH model was investigated. Following ICH, critical protein expression, BBB permeability, and neurological function were measured with or without CD200Fc administration. Our results showed that both the expression of CD200 and CD200R1 decreased after ICH and administration of CD200Fc attenuated BBB leakage and improved neurological functions. In conclusion, our work demonstrated that CD200Fc might be a potential treatment option for ICH by protecting the BBB and improving functional outcomes.
Introduction
Intracerebral hemorrhage (ICH) is associated with high mortality and high morbidity 1 –3 . Currently, no effective treatment option is available to improve functional outcomes in patients with ICH. Beyond the immediate impact of ICH, secondary brain injury is the leading cause of death and long-term disability following ICH 4 . Blood–brain barrier (BBB) disruption, which leads to tissue exudation, neuronal necrosis, and apoptosis, further propagates brain edema 5 . The combination of such pathologies has been noted as an essential aspect of secondary brain injury.
CD200 is a type-I cell surface glycoprotein. It interacts with its receptor CD200 R, which includes CD200R1, CD200R2, CD200R3, and CD200R4 in mice and is expressed on myeloid cells (including microglia). The CD200 R family is responsible for the regulation of microglia function in several neurological diseases, such as Parkinson’s disease 6 , multiple sclerosis 7 , white matter ischemia 8 , and spina bifida 9 . The main isoforms of the CD200 R family which are expressed in the mouse brain are CD200R1 and R4. While the human CD200 R family only contain CD200R1 and CD200R2 10 . Thus, we focused our attention on CD200R1 in this study. CD200Fc, a fusion protein of CD200 and the immunoglobulin (Ig)G1 Fc segment applied in this study was reported to interact specifically with CD200R1 but not CD200R2-R4 11 –13 .
In previous studies which utilized a CD200 knockout mice model, higher BBB permeability and more infiltration of peripheral proinflammatory cells in the nervous system was observed 14 , which indicated that CD200 might be related to BBB function and neuroinflammation.
In this study, we aimed to explore whether CD200Fc treatment can attenuate ICH-induced BBB disruption.
Materials and Methods
Animals
Animal protocols were approved by the Zhejiang University Committee on the Use and Care of Animals. A total of 124 male C57/Bl6 mice (6.5% mortality or asymptomatic) weighing 20–24 g were used in this study.
ICH Models
The homologous blood injection ICH model was adapted from a previously described model in mice 15 . The animals were anesthetized with 1% pentobarbital (80 mg/kg, intraperitoneally), and then positioned in a stereotaxic frame (World Precision Instruments, Sarasota, FL, USA), while core temperature was maintained at 37.5°C with a heating pad (temperature was monitored with a rectal thermometer). A cranial hole (1 mm) was drilled 2.2 mm lateral and 0.2 mm anterior to the bregma, near the right coronal suture. Autologous blood was collected from the caudal artery. A 26-gauge needle was inserted to the right basal ganglia through the hole (3.5 mm ventral). A total of 20 μl of autologous blood was injected at a rate of 2 μl/min using an automated pump (World Precision Instruments). The needle remained fixed for 5 min to prevent reflux and then was gently removed to limit any unintended damage. Animals in the sham groups only had a needle inserted and then removed.
Experimental Design
The experiments were conducted as follows.
Experiment 1
The time course of CD200 and CD200R1 expression after ICH were analyzed using Western blot. The ipsilateral hemispheres of mice from group sham, 6 h, 12 h, 24 h, 72 h were collected for analysis.
Experiment 2
Immunofluorescence was performed to check the localization of CD200R1 at 24 h after ICH. Four mice were used in both the sham group and the ICH group.
Experiment 3
For the Evans blue test, 24 mice were randomly divided into sham, ICH, ICH + CD200Fc (0.08 μg/g, intracerebroventricularly (i.c.v)), and ICH + IgG (0.08 μg/g, i.c.v) groups (n=6). Treatments were administrated at 1 h after surgery. The function of the BBB in each group was assessed at 24 h using Evans blue. For the brain water content test, a total of 24 mice were divided and treated similarly. Brain edema was evaluated by measuring brain water content.
Experiment 4
Mice were randomly divided into four groups with six animals in each group: sham, ICH, ICH + CD200Fc (0.08 μg/g, i.c.v), and ICH + IgG (0.08 μg/g, i.c.v) groups. Neurobehavioral tests including the Garcia test, foot-fault, corner turn, and rota-rod were performed.
Experiment 5
The downstream molecule Dok1 was measured in sham, ICH, ICH + CD200Fc (0.08 μg/g, i.c.v), and ICH + IgG (0.08 μg/g, i.c.v) groups (n=4), using Western blot.
Western Blotting
Mice were transcardially perfused with 50 ml of ice-cold phosphate-buffered saline (PBS). The brain tissue was homogenized and then centrifuged at 12,000 r/min for 30 min at 4°C, and the supernatant was collected. Protein concentration was determined by the bicinchoninic acid protein assay kit (Pierce, Thermo Scientific, Waltham, MA, USA). Protein was then loaded and separated using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a nitrocellulose (NC) membrane. After blocking with 5% nonfat milk, the membranes were incubated overnight at 4°C with primary antibodies: rabbit anti-CD200 (1:300, Santa Cruz Biotechnology, Santa Cruz, CA, USA), goat anti-CD200R1 (1:300, Santa Cruz), mouse anti-Dok1 (1:300, Santa Cruz) and mouse anti-glyceraldehyde 3-phosphate dehydrogenase (GAPDH; 1:3000; KangChen, Shanghai, China). The membranes were then processed with a secondary antibody against goat IgG (1:3000; Multi Sciences, Hangzhou, Zhejiang, China) or mouse IgG (1:3000; Multi Sciences, China) for 2 h at room temperature. Bands were visualized using the chemiluminescence (ECL) detection reagents and then analyzed with ImageJ software.
Immunofluorescence Staining
Mice were perfused with 4% paraformaldehyde in 0.1 mol/l PBS (pH 7.4). The brains were kept in 4% paraformaldehyde for 12 h and then dehydrated in 30% sucrose for 2 to 3 days at 4°C. A series of 10-μm coronal brain sections (LEICA, CM3050 S, Buffalo Grove, IL, USA) were blocked with 5% donkey serum albumin for 2 h at room temperature and then incubated with goat anti-CD200R1 (1:200, R&D Systems, Minneapolis, MN, USA), rabbit anti-Iba-1 (1:500, Wako, Chuo-ku, Osaka, Japan), rabbit anti-GFAP (1:200, Abcam, Cambridge, MA, USA), and rabbit anti-NeuN (1:200, Abcam) at 4°C for 2 nights. After being rinsed three times with PBS, the sections were incubated with appropriate secondary antibodies for 2 h at room temperature. The perihematomal areas of stained sections were examined under a confocal laser scanning microscope (Olympus, FV3000, Tokyo, Japan). Microphotographs were analyzed with FV-10 ASW software.
Evans Blue Assay
Mice received an intraperitoneal injection of 2% Evans blue solution (4 ml/kg of body weight), at 24 h after surgery. Moreover, the stain was allowed to circulate for 3 h. Following circulation, mice were perfused with 50 ml ice-cold PBS. The brains were then removed and divided into left and right hemispheres, frozen in liquid nitrogen, and stored at −80°C. Right hemisphere samples were homogenized in PBS and then centrifuged at 12,000 r/min for 30 min at 4°C, and the supernatant was collected. An equal amount of 50% trichloroacetic acid was added to each 500 μl of the supernatants and then incubated overnight at 4°C. Later it was centrifuged at 12,000 r/min for 30 min at 4°C, and then the quantity of Evans blue stain was detected by spectrophotometer (BioTek, Winooski, VT, USA) at 610 nm and quantified according to a standard curve. These results are presented as milligrams of Evans blue stain per gram of brain tissue.
Brain Water Content
Brain water content was measured using the wet/dry method. Mice were sacrificed under anesthesia without perfusion at 24 h after surgery. The brains were removed quickly and divided into ipsilateral and contralateral cortices (Ipsi-CX and Cont-CX), ipsilateral and contralateral basal ganglia (Ipsi-BG and Cont-BG), and cerebellum (Cerebel). The wet weight of each part was measured immediately, and the dry weight was obtained after the brain tissue had been dried for 24 h.
Short-Term Neurobehavioral Test
Garcia test at 24 h after ICH was carried out, including assessment of spontaneous activity, axial sensation, vibrissae proprioception, symmetry of limb movement, lateral turning, forelimb walking, climbing, and grabbing.
Long-Term Neurobehavioral Test
Foot-fault, corner turn, and rota-rod tests were performed every 24 h in the first 7 days after ICH. Foot-fault: animals were placed on a horizontal 26 × 13 grid floor (wire diameter 1.3 cm) for 5 min. The number of foot faults of each forelimb were recorded using a camera (Logitech Webcam, Lausanne, Switzerland). Corner turn: the mice were placed in a 30-degree corner. We then recorded whether the animals turned left or right. The corner turn test was repeated 10 times for each mouse, and the percentage of right turns was calculated. Rota-rod: animals were placed on an accelerating rod (YLS-4C, Xuzhou, Jiangsu, China), with a max speed of 40 r/min and the time to fall was recorded for each run. All animals were trained twice daily until their results were consistent, before surgery.
Statistical Analysis
All data was given in means ± standard deviation (SD). The data was analyzed using GraphPad Prism version 6.02 with a Student’s t test and one-way analysis of variance (ANOVA). Multiple comparisons between the groups were performed using a Tukey test. Differences were considered significant at p < 0.05.
Results
Expression of CD200 and CD200R1 after ICH
Results of the Western blot showed that the expression of CD200 decreased at 24 h (Fig. 1B). While the expression of CD200R1 decreased since 6 h (p < 0.05, versus sham) and rose again at 72 h (Fig. 1A).

CD200R1 and CD200 expression in brain after ICH. (A) Western blot showed significant decrease of CD200R1 at 6 h, 12 h, and 24 h after ICH (∗p < 0.05 versus sham), and increased at 72 h (#p < 0.05, versus 24 h). (B) CD200 decreased at 24 h after ICH (∗p < 0.05 versus sham). N=4 mice/group. Values are expressed as mean ± SD.
Expression of CD200R1 in Perihematomal Tissue
Double immunofluorescence staining was performed in the basal ganglia area of sham and ICH groups. The staining found that CD200R1 was expressed on microglia in both sham and ICH groups (Fig. 2A and B), while CD200R1 was not expressed on astrocytes or neurons (Fig. 2C and D).
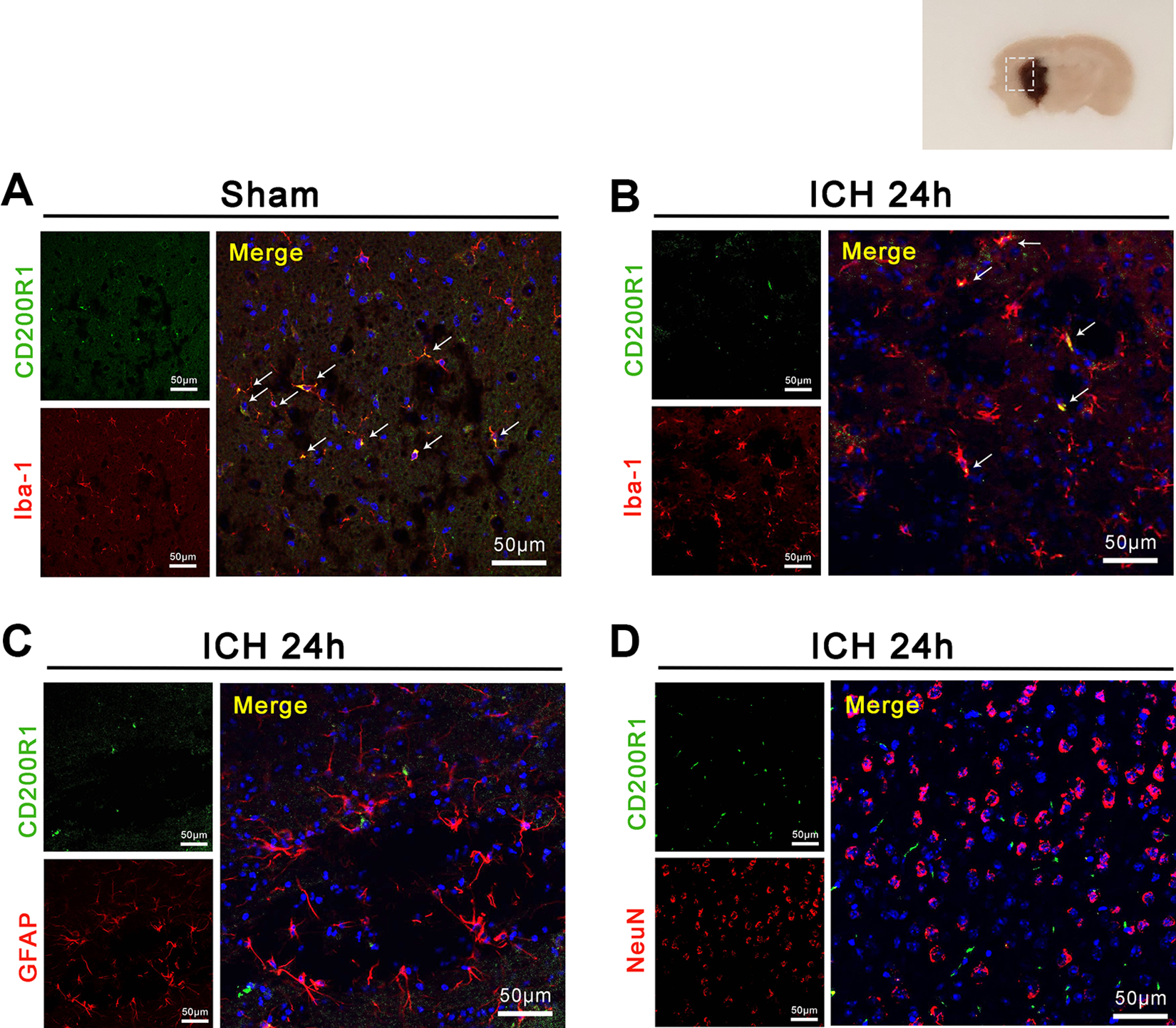
Localization of CD200R1 in brain. (A–B) CD200R1 was detected on Iba-1 positive cells in sham and ICH groups. (C-D) CD200R1 did not co-localize with GFAP or NeuN.
CD200Fc Protected BBB Integrity at 24 h after ICH
To examine whether CD200R1 plays a vital role in BBB protection after ICH, we tested the permeability of the BBB in mice with Evans blue and brain edema with brain water content. The results showed that the accumulation of Evans blue increased in our mice at 24 h after ICH, compared with sham. Mice injected with normal IgG showed no significant difference from the nontreatment group. Also, ICH mice that received CD200Fc treatment had a lower release of Evans blue than ICH with IgG. (Fig. 3A). The brain water content was higher in ICH mice than sham mice. Besides, mice that received CD200Fc treatment showed significant improvement in both ipsilateral basal ganglia and cortex after ICH. (Fig. 3B).

Evans blue assay and brain water content. (A) Accumulation of Evans blue increased at 24 h after ICH (∗∗p < 0.01 versus sham). Evans blue decreased in mice treated with CD200Fc (#p < 0.05 versus ICH). N=6 mice/group. (B) Brain water content showed significant increase in ipsilateral basal ganglia and cortex at 24 h after ICH while was reversed by CD200Fc treatment (∗∗p < 0.01 versus sham, #p < 0.05 versus ICH). N=6 mice/group. Values are expressed as mean ± SD.
CD200Fc Improved Neurobehavioral Outcomes
A 1 week a neurobehavior test was performed in order to see whether CD200Fc influenced the functional recovery after ICH. Foot-fault and corner turn tests showed a significant difference between ICH groups and CD200R1 treatment groups on the first 7 days after ICH (Fig. 4A and B). The rota-rod test showed a significant improvement in CD200Fc treatment group from day 2 to day 7 (Fig. 4C). The Garcia test showed neuronal function improvement at 24 h after ICH in the CD200Fc-treated group (Fig. 4D).

Neuronal behavior test at 24 h after ICH. (A–C) Long-term tests showed significant improvement in CD200Fc-treated mice. (D) Short-term Garcia test showed significance improvement in CD200Fc-treated mice. (∗p < 0.05 versus sham, ∗∗p < 0.01 versus sham, #p < 0.05 versus ICH, ##p < 0.01 versus ICH. Values are expressed as mean ± SD).
CD200Fc Upregulated Dok1
Dok1, the downstream protein of CD200R1, decreased at 24 h after ICH, which was reversed by treatment with CD200Fc. (Fig. 5)
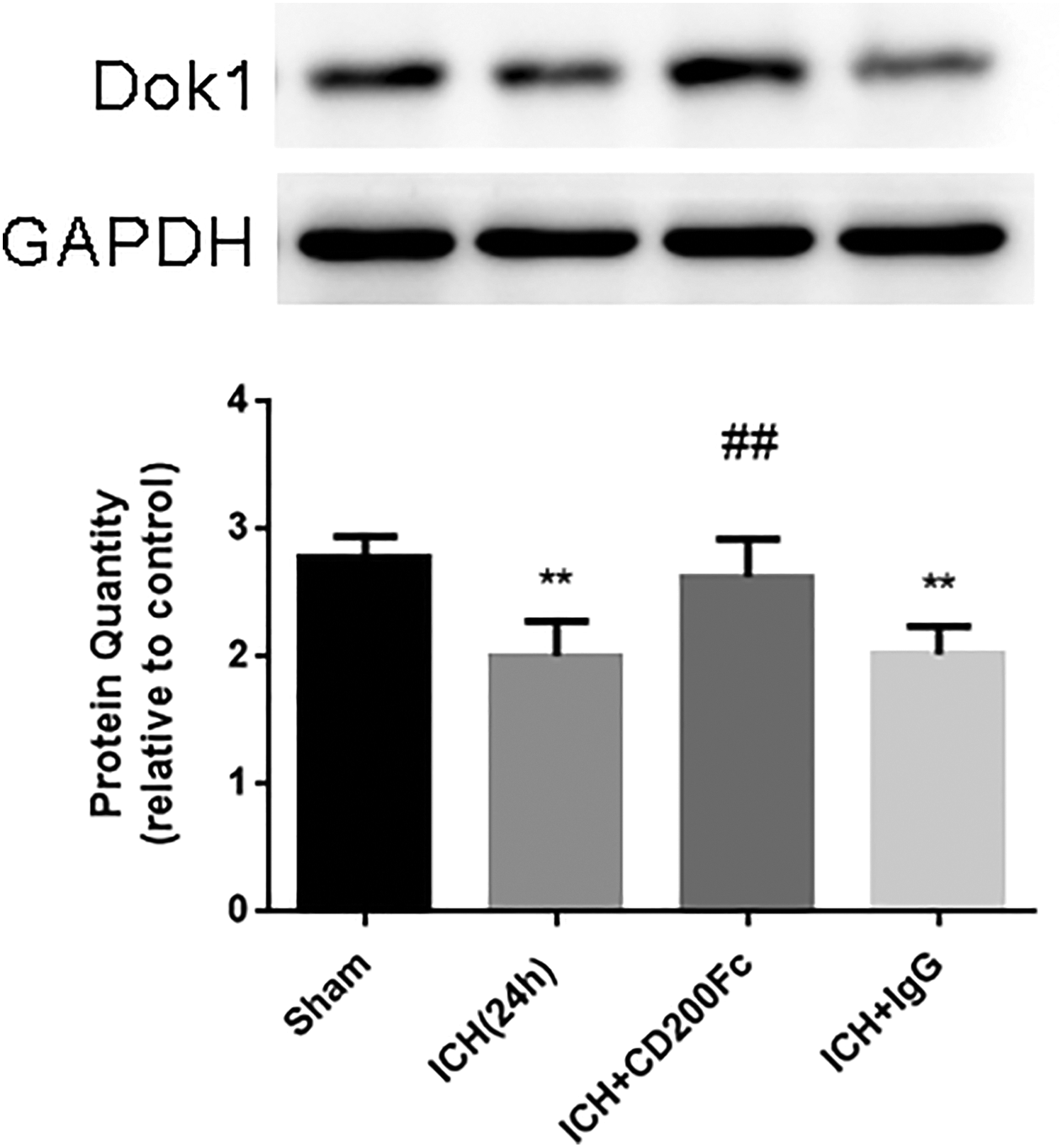
Dok1 expression after ICH and different treatments. Dok1 expression decreased at 24 h after ICH. While treatment with CD200Fc showed significant improvement (∗∗p < 0.01 versus sham, ##p < 0.01 versus ICH. Values are expressed as mean ± SD).
Discussion
Though intracerebral hemorrhage is the leader of mortality and morbidity in all subtypes of stroke, there are few effective treatments for improving functional outcome in patients with ICH 16 . In this study, we found that administration of CD200Fc attenuated BBB disruption at 24 h and improved functional neurological outcomes of mice in the first week after ICH. CD200, the ligand of CD200 R family, is a surface protein widely expressed in central nervous system 17 . The decrease of interaction between CD200 and its receptor may promote the proinflammatory effect of microglia 18 . The decrease of CD200 expression and exaggeration of microglial response was reported in several disease models such as multiple sclerosis 19 and Parkinson’s disease 20 . Though a microglia response has been noted, little is known about the modification of the CD200 receptors. Studies have shown a decrease of CD200 R on macrophages in patients with Parkinson’s disease 21 as well as on microglia in Alzheimer’s disease patients 22 . However, no significant alteration of CD200 R expression was found in patients with multiple sclerosis 23 . In the present study, we found that both CD200 and its receptor decreased after ICH in mice. A recent study showed that the disruption of the BBB might reduce CD200 expression by interleukin (IL)-6 and tumor necrosis factor (TNF)-α produced by the microglia 24 . Though CD200 expression is better understood, there is still a substantial gap in the understanding of why there would be a decrease in CD200R1. According to some studies, anti-inflammatory cytokines, such as IL-4 and IL-13 stimulate CD200 R expression on microglia 22,25 . This stimulation may be related to the activation of microglia and ultimately responsible for BBB disruption and neurobehavior deficit after ICH.
The BBB is a system composed of microvascular endothelium, astrocytes, basement membrane, pericytes, and neurons 26,27 . In a thrombin-induced ICH injury model, blocking the downstream activation of thrombin alleviated BBB disruption and promoted brain microvascular endothelial cell and astrocyte regeneration 28 . This experimental design allows for a new approach for studying neuroprotection after cerebral hemorrhage and the pathologies of secondary brain injury.
Microglia get activated very soon after ICH and play a vital role in secondary injury 29,30 . Yan et al. 31 , found that microglia can not only work on neurons or other glia but also alter the microvascular structure. As more evidence continues to suggest that microglia are powerful players in innate immunity and maintain homeostasis in the nervous system, microglia should not be separated from the BBB 30 . When over-activated, microglia accumulate around the blood vessels, and destruction of microvascular structures takes place, including atrophy of endothelial cells, thickening of basement membrane, vacuolization, and degeneration of pericytes. The accumulation of these complications results in the changes of BBB permeability and exudation around the blood vessels 32,33 . Previous studies have demonstrated that, CD200 is expressed mainly on the neurons, but also vascular endothelial cells 6 and astrocytes 34 . In the present study, we found CD200R1 is expressed on microglia but not on neurons or astrocytes, which indicated that the CD200–CD200R1 pathway might be a bridge between neurovascular units and microglia. Thus, further studies need to be carried out to determine whether CD200Fc protects the BBB or improves neurological functions through regulating microglia.
Since CD200 fusion protein has been used in several in vivo experiments 8,35 –37 , we treated mice with CD200Fc at a dosage accordingly 36,37 . Following treatment, Evans blue, brain water content and a series of neurobehavioral tests were applied to estimate the treatment effect of CD200Fc. We found that CD200Fc treatment resulted in a significant reduction of BBB permeability, Evans blue accumulation and brain water content was decreased in CD200Fc-treated mice, compared with the nontreatment group and the IgG-treated group. This finding coincided with the improvement of the neurobehavioral scores of the CD200Fc-treated mice. Among the long-term neurobehavioral tests, foot-fault and corner turn tests showed a significant difference following the first day after administration of CD200Fc, while the rota-rod test showed a significant difference following the second day.
There are a few limitations to our study. We directly focused on BBB function but did not look into the anti-inflammation or anti-apoptotic effect of CD200R1, although anti-apoptosis of CD200R1 function is quite widely studied in tumor and autoimmune diseases. Furthermore, the detailed mechanism about how the signaling pathways of CD200/CD200R1 changed with or without CD200Fc treatment in ICH was not demonstrated in our study. Previous studies have confirmed in both in vivo and in vitro studies using rat models, that when CD200 R binds to CD200, Y297 tyrosine in the PTB domain of its intracellular NPXY sequence is rapidly phosphorylated, then the phosphorylated inhibitory molecule Dok1 and Dok2 proteins are recruited and bound to Ras-GAP and SHIP. This cascade furtherly inhibits RAS and its downstream ERK, JNK, and P38 MAPK activation 38 –40 . As a downstream molecule of ERK, MMP-9 has been proven to be important in BBB permeability 41 –43 . Here we assume that activation of CD200 R protects BBB through ERK/MMP-9 signaling pathway. Thus, future studies are needed to investigate which downstream signals take charge in ICH and what changes take place during the treatment.
Footnotes
Ethical Approval
The animal study protocol was approved by the Institutional Animal Care and Use Committee of Zhejiang University.
Statement of Human and Animal Rights
All procedures were performed according to the protocol approved by the Institutional Animal Care and Use Committee, Zhejiang University.
Statement of Informed Consent
There are no human subjects in this article, and informed consent is not applicable.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the National Natural Science Foundation of China (NSFC; 81471168) to Feng Gao, NSFC (81500991) to Lu-sha Tong.