
Abstract
Our group previously demonstrated that administration of a CD34-negative fraction of human non- hematopoietic umbilical cord blood stem cells (UCBSC) 48 h after ischemic injury could reduce infarct volume by 50% as well as significantly ameliorate neurological deficits. In the present study, we explored possible mechanisms of action using next generation RNA sequencing to analyze the brain transcriptome profiles in rats with ischemic brain injury following UCBSC therapy. Two days after ischemic injury, rats were treated with UCBSC. Five days after administration, total brain mRNA was then extracted for RNAseq analysis using Illumina Hiseq 2000. We found 275 genes that were significantly differentially expressed after ischemic injury compared with control brains. Following UCBSC treatment, 220 of the 275 differentially expressed genes returned to normal levels. Detailed analysis of these altered transcripts revealed that the vast majority were associated with activation of the immune system following cerebral ischemia which were normalized following UCBSC therapy. Major alterations in gene expression profiles after ischemia include blood-brain-barrier breakdown, cytokine production, and immune cell infiltration. These results suggest that UCBSC protect the brain following ischemic injury by down regulating the aberrant activation of innate and adaptive immune responses.
Keywords
Introduction
Each year, over 750,000 individuals in the United States suffer a stroke, making it a leading cause of morbidity and mortality. The primary treatment for stroke is administration of tissue plasminogen activator (tPA). Unfortunately, the vast majority of stroke victims are unable to reach hospital emergency rooms within the 3- to 5-h time-window following stroke onset required for effective tPA therapy. Therapies that expand the time window of treatment are therefore much needed.
We previously isolated a CD34-negative population of non-hematopoietic umbilical cord blood stem cells (UCBSC) within human umbilical cord blood that is highly expandable and exhibit stem cell-like properties 1 . Systemic administration of UCBSC 48 h after ischemic brain injury resulted in a significant reduction in infarct volume and an amelioration in neurological severity score as well as limb placement function in the rat MCAO model of stroke 1,2 . Interestingly, very few UCBSC were observed in the brain 28 days post transplantation, suggesting a neurotrophic or anti-inflammatory function of UCBSC therapy. Indeed, flow cytometry analysis revealed a reduction in infiltrating immune cells following ischemic stroke and UCBSC treatment 2 . The use of stem cells derived from the umbilical cord blood/matrix have garnered recent attention for their immunomodulatory and neuroprotective properties 3 –6 . Animal and human studies of ischemic brain injuries demonstrate that the activation of the immune system and inflammation are important pathological process following ischemia 7 –9 .
In the present study, we investigated the immunological mechanisms by which UCBSC exert their neuroprotective effects, focusing on blood-brain-barrier (BBB)/extracellular matrix (ECM) function, immune cell infiltration, and secondary apoptosis. We postulate that UCBSC therapy modulates the immune system to reduce the infiltration of immune cells into the brain following ischemic injury, thereby reducing secondary injury. To address this issue we performed RNA sequencing (RNAseq) to study the brain transcriptome change in rats following ischemic injury and after the systemic administration of UCBSC.
Materials and Methods
Animal Model of Acute Focal Ischemic Stroke
To induce acute focal transient ischemic stroke, adult Sprague Dawley rats (175.0 ± 25.0 g; Groups: Normal, MCAO, MCAO+UCBSC; n = 5/group) were anesthetized with a cocktail (0.85 ml/kg) of ketamine (75 mg/ml Ketaject; Phoenix Pharmaceuticals, Burlingame, CA, USA) and xylazine (10 mg/ml Xyla-ject; Phoenix Pharmaceuticals) intramuscularly. Each rat’s head was stabilized in a Koph head holder on Stereotaxic Frame (Tujunga, CA, USA) in a supine position after the fur of the neck area was shaved and sterilized. A midline incision in the skin of the neck was made to expose subdermal structures. The right common carotid artery was exposed and traced distally to its bifurcation, the superficial branch of which is the external carotid artery (ECA). The right ECA was cauterized and cut to be able to introduce the thread occluder for the temporary ligation. The right internal carotid artery (rICA) has an extracranial branch before it enters the cranium—the pterygopalatine artery—which was temporarily ligated for a complete ischemic lesion. The rECA was cut at its distal stem so that a silicon-coated surgical thread could be inserted into its lumen. The thread was inserted into the bifurcation, up into rICA, to occlude the middle cerebral artery (MCAO). The occluder is kept in place for 1 h before removal. The open arteries were then cauterized, the wound closed, and the animal monitored until fully sternal and recovered. All procedures were approved by the IACUC at the University of Minnesota (Protocol Number: 133-31062A). Considerations for the ethical use of animals in this study as well as alternatives to the use of animals were submitted to IACUC prior to final approval and authorization.
UCBSC Culture
CD34 negative UCBSC were isolated and grown as previous described
1
. Briefly, umbilical cord blood was collected after delivery and, within 4 h, mononuclear cells were separated by centrifugation at 500 × g for 30 min in a Ficoll-hypaque density gradient (1.077 g/cm3) (Sigma, St. Louis, MO, USA). The cells were then grown in Dulbecco’s modified Eagle medium DMEM (Invitrogen, Waltham, MA, USA) / MCDB-201(Sigma) mix with 10% fetal bovine serum (Invitrogen), 10–4 M of
Stem Cell Delivery
Two days post-MCAO, animals in the treatment group were injected with 1 million UCBSC suspended in 500 μL of sterile saline delivered via saphenous vein under isoflurane anesthetization. Immunosuppressant drugs (e.g. Cyclosporine A) were not administrated as we have previously observed beneficial effects of UCBSC therapy in the absence of immunosuppressive agents 10 .
Behavioral Testing
Neurological function was assessed by a neurological severity score (NSS) test 11 on day 2 and day 7 post-MCAO. The NSS test consists of eight subtests evaluating motor and sensorimotor function of the animals’ limbs. Left and right side limbs were each scored as either 2 (normal function), 1 (impaired function), or 0 (unable to perform). The scores from these subtests were tallied for the right side and left side limbs and analyzed using a Student’s t-test.
Tissue Processing and RNA Sequencing
Immediately following NSS testing on day 7, the rats were deeply anesthetized with ketamine (150 mg/kg). We chose to do the RNAseq at 7 days because we wished to compare the transcriptome results with our previous flow cytometry study 2 . Rats were then transcardially perfused with phosphate buffer saline and the hemisphere ipsilateral to the MCAO was isolated. Total RNA was extracted from all 15 brains using the RNeasy Plus Mini Kit (Qiagen, Germantown, MA, USA) according to manufacturer’s instructions. Each sample was obtained in triplicate (one animal had duplicate samples only) for a total of 44 total RNA samples submitted for sequencing.
RNA samples were analyzed at the University of Minnesota Biomedical Genomics Center. Briefly, total RNA samples were quantified using the RiboGreen fluorometry assay and RNA integrity was checked using capillary electrophoresis (Agilent BioAnalyzer 2100; Agilent, Santa Clara, CA, USA). Libraries were generated from 1 μg of total RNA. Polyadenylated coding mRNA in each sample was isolated and reverse transcribed using random primers. The resulting paired-end cDNA libraries were subsequently sequenced using the HiSeq 2000 (Illumina, San Diego, CA, USA). For each sample, at least 10 million paired-end reads of 50 base pairs were performed in four lanes.
Bioinformatics Analysis
FASTQ files for each sample were first combined before analysis. Raw sequences were analyzed using a customized pipeline (gopher-pipelines; https://bitbucket.org/jgarbe/gopher-pipelines/overview) developed and maintained by the Minnesota Supercomputing Institute. Briefly, quality controls were performed on each FASTQ files using FastQC (v0.11.5) before and after adapter trimming with Trimmomatic (v0.33). Post-trimming sequences were aligned to Rattus norvegicus reference genome using HISAT2 (v2.0.2). Transcript abundance was then estimated using subread (v1.4.6) 12,13 . Pathway analysis of differentially expressed genes was performed by functionally annotating the genes, performing an overrepresentation enrichment test using BiNGO package, and visualizing using Cytoscape 14 .
Read counts generated by subread were filtered to include genes with counts per million >5 and subsequently analyzed in R 15 . The remaining reads were then normalized and log transformed using edgeR. Heat maps were generated using the log transformed values with pheatmap package. Hierarchical clustering was performed using Euclidean distances and average linkage clustering method. Principle component analysis (PCA) was performed in R using prcomp and visualized using ggplot2 and rgl packages. Immune cell subtype analysis was performed using CIBERSORT (https://cibersort.stanford.edu/) following the recommended settings for RNA-Seq data. Two-sided Student’s t-test was performed to assess the CIBERSORT results 16 . All P-values are listed in the supplementary files. P-values ≤ 0.05 were considered as significant differences.
Results
UCBSC Therapy Ameliorates Behavioral Deficits
Two days after MCAO, all animals exhibited sensorimotor deficits assessed by the NSS test. At this time-point, no significant differences were observed between the two treatment groups ( Fig 1 ). Five days after the systemic administration of UCBSC (7 days post-MCAO), a significant improvement in NSS score was observed in ischemic rats treated with UCBSC, relative to untreated ischemic rats (t-test P = 0.046).

UCBSC therapy ameliorates neurological deficits induced by MCAO. (A) The Neurological Severity Scores (NSS) were similar for the two groups of animals 2 days post MCAO but prior to the UCBSC therapy. Five days after UCBSC cell therapy (day 7 after MCAO), NSS scores improved significantly in the UCBSC treated group (t-test P = 0.046). (B, C, and D) PCA of RNA-seq data show MCAO-treated samples cluster separately from Normal and MCAO+UCBSC groups.
UCBSC Therapy Globally Diminishes the Immunological Response to Stroke
To gain insights into the mechanisms of action underlying UCBSC therapy in ischemic brain injury, we compared the brain transcriptome in normal, MCAO, and MCAO+UCBSC rats. After initial filtering, a total of 12,788 genes were used for differential expression (DE) analysis. PCA shows the MCAO group is clearly clustered separate from both control and MCAO+UCBSC treatment groups (
Fig 1A

Gene expression analysis following MCAO and UCBSC treatment. (A) Scatter plot of significantly DE genes that return to normal expression levels after UCBSC treatment. Black dotted line represents expression levels of transcripts from normal brain. Red dots are expression levels of transcripts from MCAO group. Aqua colored circles are expression levels of transcripts after UCBSC treatment. X-axis indicates log10 counts per million (cpm) level from normal brain. Y-axis represents corresponding expression levels for the same transcript from MCAO group or UCBSC group. (B) Gene expression patterns following MCAO and UCBSC treatment.
UCBSC Therapy Normalizes Gene Transcripts Related to BBB and ECM Function
Because the breakdown of the extracellular matrix (ECM) and blood-brain-barrier (BBB) allows immune cells to penetrate into the brain following ischemic injury, we specifically investigated genes related to the BBB functions ( Fig 3A ; Supplementary Table ST5, 6). We also identified genes associated with extracellular matrix (GO:0031012) from the Gene Ontology Consortium ( Fig 3B ). The results show most genes associated with BBB and ECM were upregulated following MCAO treatment and normalized by treatment with UCBSC (Supplementary Table ST5, 6).
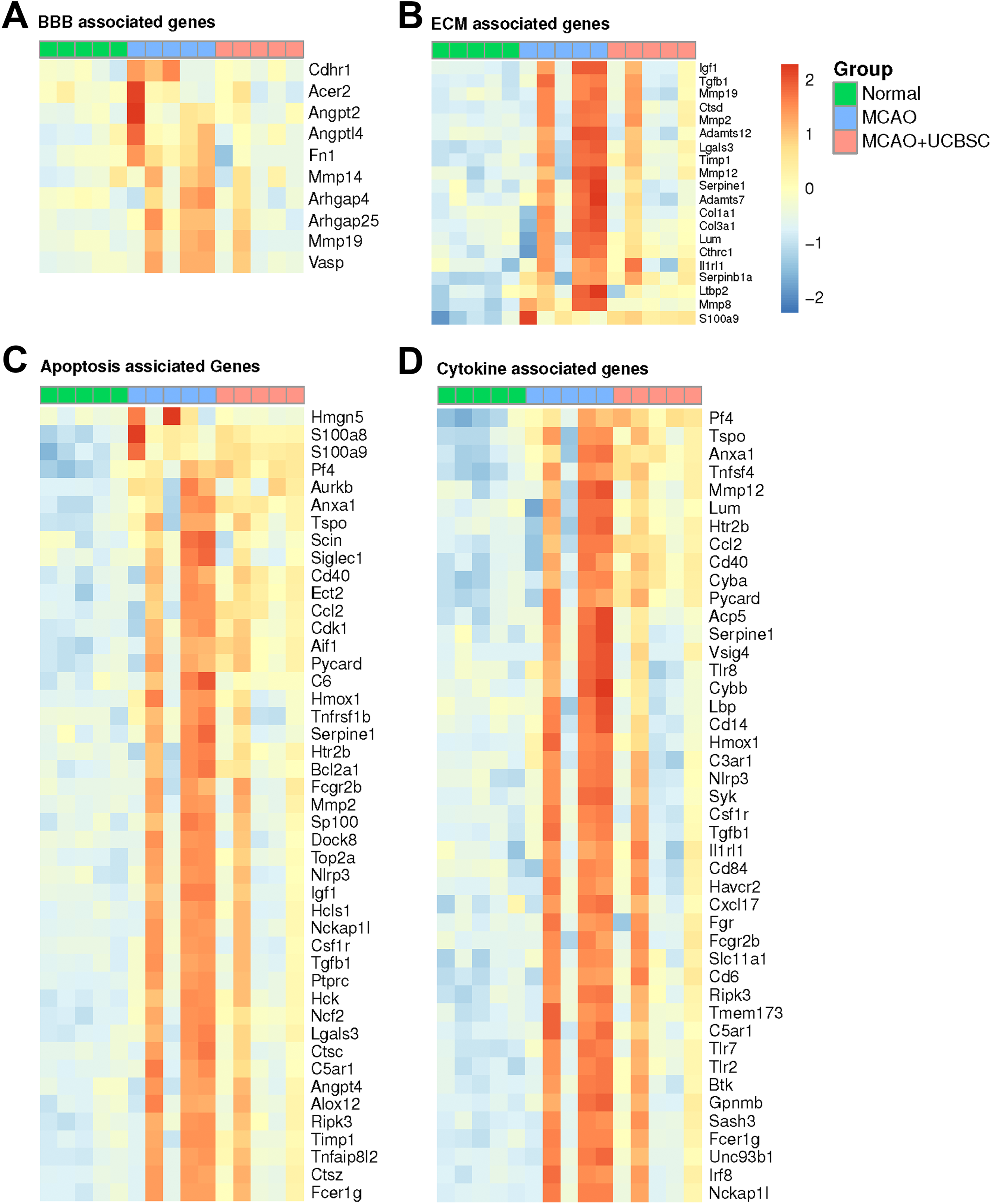
Heatmap of (A) BBB, (B) ECM, (C) apoptosis, and (D) cytokine associated genes. The expression pattern of these genes return to normal expression levels after UCBSC treatment. Red indicates relative increase in expression and blue indicates relative decrease in expression.
UCBSC Therapy Reduces Apoptotic Gene Transcripts
To assess if UCBSC can reduce cell death after MCAO, we investigated genes related to apoptosis. We identified genes associated with the regulation of apoptotic process (GO:0042981) from the Gene Ontology Consortium. The results show most genes associated with apoptosis were upregulated following MCAO treatment and normalized with UCBSC therapy (
Fig 3C
UCBSC Therapy Normalizes Gene Transcripts Related to Cytokine Production
Cytokines are involved in stroke-induced cerebral inflammation. We identified genes associated with regulation of cytokine production (GO:0001817) from the Gene Ontology Consortium plotted as a heatmap ( Fig 3D ). Results demonstrate many genes associated with pro-inflammatory cytokines were upregulated following MCAO treatment and normalized by UCBSC (Supplementary Table ST8).
UCBSC Therapy Returns Immune Cell Subtypes to Normal Levels
We next investigated the immune cell infiltration after MCAO using CIBERSORT with the lm22 immune gene signature set ( Fig 4 ). The absolute score estimated by CIBERSORT show significantly increased immune cell infiltration after MCAO (0.97 vs. 0.47; MCAO vs. Control; two-sided t-test P = 0.039). An intermediate effect of UCBSC was observed where treated animals were not statistically significant from MCAO (0.97 vs. 0.60; MCAO vs. MCAO+UCBSC; P = 0.126) nor controls (0.47 vs. 0.60; Control vs. MCAO+UCBSC; P = 0.160).
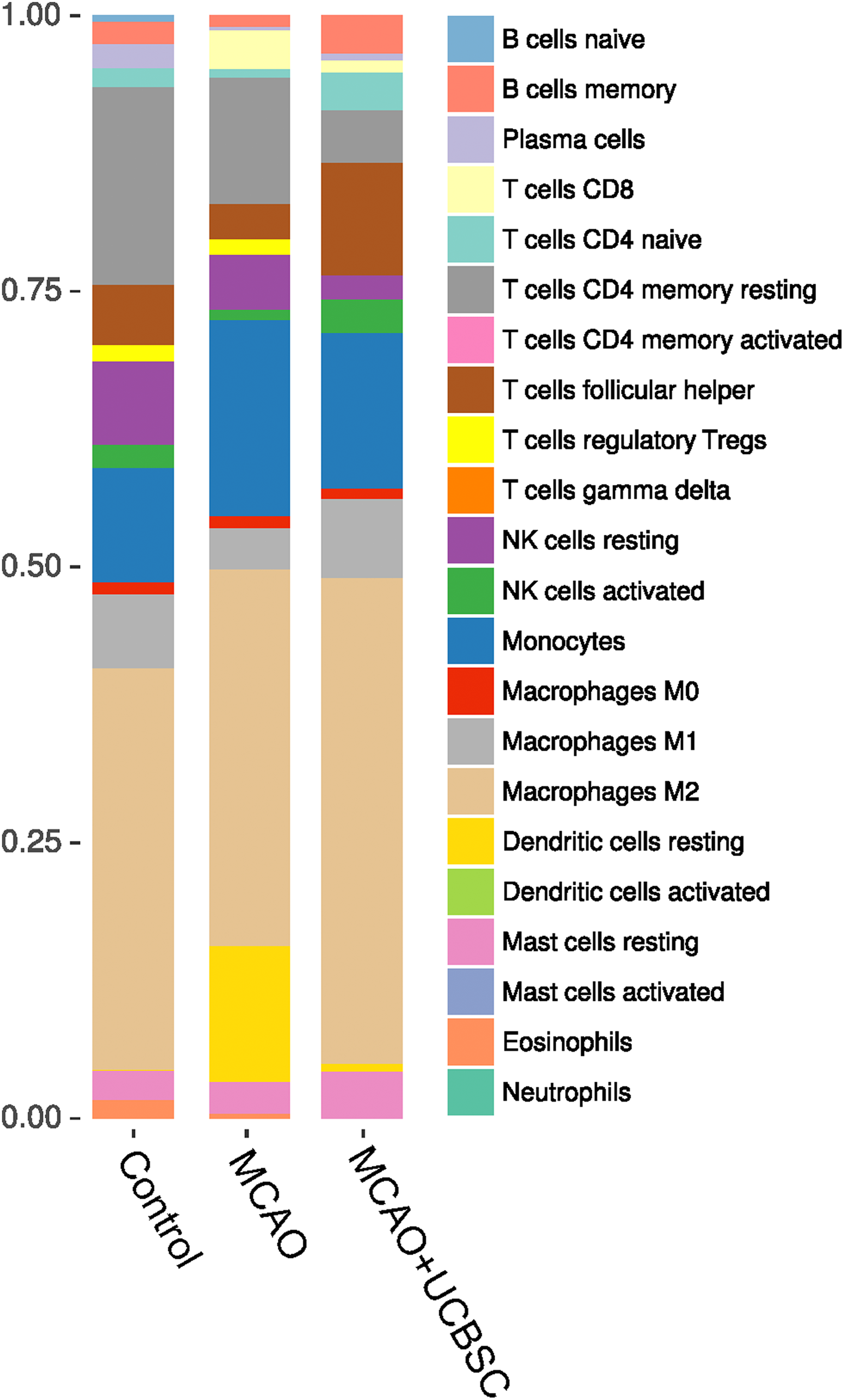
CIBERSORT analysis of control, MCAO, and UCBSC treatment. The relative percentage of different immune cell types were deconvoluted from the RNA-seq data using CIBERSORT.
Deeper analysis among the subtypes of immune cell and inflammatory cell within the CIBERSORT identified a significant increase in transcripts specific for monocytes, resting dendritic cells, CD8- and naive CD4-T cells, as well as a decrease in transcripts specific for M2 macrophages following MCAO, relative to controls. Gene transcripts associated with neutrophils ( Fig 5A ), natural killer cells ( Fig 5B ), microglia/macrophages ( Fig 5C ), B-cells ( Fig 5D ), mast cells ( Fig 5E ), CD4 T-cells ( Fig 5F ) and regulatory T-cells ( Fig 5G ) were significantly increased after MCAO, and UCBSC therapy reversed these transcripts back to normal levels.

Box plot showing RNA expression levels change of (A) Neutrophil, (B) Natural Killer cell, (C) Macrophage, (D) B-cell, (E) Mast cell, (F) T-cell, and (G) Regulatory T-cell related gene transcripts after MCAO (Red) and UCBSC treatment (Green). * denotes statistical significance between the three groups P < 0.05 (edgeR, false-discovery-rate adjusted).
Discussion
Consistent with our previously published reports 2 , the data presented here suggest UCBSC is an effective treatment for ischemic stroke and acts through modulation of the immune system. We hypothesize that a breakdown of the BBB and ECM results in increased infiltration of immune cells, leading to secondary damage at the site of infarction. We observed that UCBSC treatment rescued sensorimotor deficits following stroke and systematically normalized gene expression alterations caused by MCAO. Our results suggest the underlying mechanism of UCBSC treatment in protecting the brain following a stroke is through dampening the immune response and immune cell infiltration. These changes are consistent with reports by other investigations on ischemic brain injury 17 –20 .
Stroke and BBB/ECM Function
The initial pathophysiological processes that permit the penetration of immune cells into the brain parenchyma following ischemic injury is the breakdown of the BBB and ECM. Cell adhesion molecules engage immune cells and initiate their penetration across the BBB 21 . After cerebral ischemia, we observed an upregulation in the expression of ICAM1, which is part of the immunoglobulin superfamily, and is involved in many processes including inflammation, immune responses and intracellular signaling. Immediately following ischemia, ICAM1 expression is localized to capillary endothelium of intra-parenchymal blood vessels, but expression is increased in leukocytes in the weeks following 22 –24 . Interestingly, cell-surface expression levels of ICAM1 correlated with infiltration of CD4 + or effector memory T-cells 25 , suggesting that modulation of ICAM1 expression by UCBSC may play a role in the observed reduction in infiltrating immune cells. Preclinical treatment of ischemia with anti-ICAM1 antibodies reduced edema and infarct size 26 , although this effect was not translated into the clinic 27 .
Matrix metalloproteinase (MMP) enzymes are responsible for the breakdown of the ECM by degrading tight junctions and basal lamina proteins, leading to immune cell infiltration 28 . We observed an upregulation of transcripts of several MMPs (including -2, -8, -12, -14, and -19) at 7 days post-ischemia that was normalized with UCBSC treatment. In rodent models of cerebral ischemia, elevation of MMP-2, MMP-8, and MMP-14 have been observed 29 –31 . Interestingly, an increase in expression of MMP regulator Timp1 was observed in untreated ischemic animals, suggesting that BBB damage has peaked and vascular remodeling may begin 31 . Further investigation is warranted to determine if UCBSC treatment inhibits BBB breakdown or mediates rapid vascular repair.
Cytokine/Chemokine Modulation
Through gene ontology, we identified DE genes in our dataset associated with cytokine and chemokine production. Specifically, we observed an upregulation of Tgf-β1 and many interleukin receptors in ischemic rats 7 days following MCAO. The increased expression of Tgf-β1 is likely driven by the increase in macrophages and microglia at the site of infarction, as previous studies have demonstrated increased colocalization of Tgf-β1 signaling in Cd68 + cells 32 . Other cytokine specific DE genes we observed to be upregulated following MCAO include Ccl2, Hmox1, TLR2, Serpine1, and C3ar1. Similarly, increase expression of chemokines Pycard, Tlr4, Il4ra, Tlr7, and Csfl1r were increased in untreated MCAO rats. These cytokines and chemokines are likely to be upregulated due to the migration of microglia at the site of infarction 33 . Given the complex interactions and kinetics between the many pro- and anti-inflammatory cytokines and their receptors following injury, it is difficult to determine the possible mechanisms at a single time-point. However, possible sites of action may include the spleen, cervical lymph nodes, and other lymphoid organs where UCBSC may interact with immune cells.
Modulation of the Immune Cell Infiltration
Using the CIBERSORT analysis package, we were able to quantify transcripts associated to individual immune-cell subtypes. Similar to our previous study interrogating inflammation through flow cytometry within the ischemic hemisphere, we observed increased transcripts associated with innate immune system cells – NK cells, neutrophils, monocytes/macrophages, and resting dendritic cells; and cells associated with the adaptive immune system – CD8-T cells and naive CD4 + T-cells, and B-cells. In addition, a decrease in pro-recovery M2 macrophages was observed. The activation of the innate immune system triggered immediately after ischemic stroke and is manifested by the mobilization of NK cells, neutrophils, resident macrophages, and microglia infiltrating the infarct zone within minutes after ischemia 34,35 . In untreated MCAO rats we observed increased expression in genes (including Trem2, Csf1, and Irf8) that have been associated with activation and proliferation of microglia 33,36 .
Infiltration of peripheral immune cells into the site of infarction also plays an important role in the outcome of ischemia. Within 1 h, antigen presenting dendritic cells infiltrate the ischemic brain 37 –39 . In the ischemic brain hemisphere, a 20-fold elevation of dendritic cells was observed at 3 days post-ischemia, which remained elevated at 12-fold normal levels on day 7 37 . In the normal brain, CD4 + and CD8 + cytotoxic T cell lymphocytes cells are present at low levels; however, after ischemic injury, there is a dramatic influx of CD4 + cells, which can activate resident brain microglia and upregulate endothelial cell adhesion molecules, increasing BBB permeability 40 –42 . Perhaps through UCBSC treatment following MCAO, a normalization of the pro-recovery M2 macrophages/microglia is able to reduce inflammation, thereby reducing infiltration of peripheral adaptive immune cells. Further work is needed to determine the kinetics of inflammation following UCBSC therapy.
Ischemic Cascade
In addition to neuronal death via necrosis or immune cell infiltration, secondary apoptosis occurs in the oxygen-deprived penumbra surrounding the initial infarct zone 20 . We observed that transcripts related to apoptosis were upregulated following ischemic injury, specifically, Scamp2, Cebpa, Shc1, Sipa1, Tep1, Ucp2, Dap, Plp2, Ccnb1, and Sla. Furthermore, treatment with UCBSC seemed to restore these transcripts to control levels.
A previous study showed Cyclin-B1 (Ccnb1) to be upregulated and cytotoxic following ischemia through excitotoxicity 43 . Other cell cycle-related genes we found to be upregulated following ischemia were Cebpa, Acer2, and Tep1. The increased expression of these genes suggests that cell proliferation mechanisms are upregulated following ischemia. This upregulation may result from glial, immune cell, or neuronal proliferation, but this question needs to be pursued further.
Reactive oxygen species (ROS) expression in the peri-infarct region has been shown to induce apoptosis following ischemia 20 . Inhibition of SHC1 following ischemia in rats has decreased infarct size, as well as penumbra-associated apoptosis, likely through a decrease in ROS creation 44,45 . In our study, we found that UCBSC lower the expression of SHC1 to non-infarct control levels, suggesting these UCBSC may decrease ROS production, thus preventing apoptosis induction.
Conclusion
In summary, we provide a systems approach to study the cellular mechanisms underlying the neuroprotective effects of UCBSC treatment for ischemic brain injury. Our transcriptomics analysis reveal UCBSC are able to reduce upregulation of transcripts associated with BBB permeability, thereby reducing infiltration and activation of immune cells, which we believe is responsible for the downregulation of apoptotic related genes. These data further suggest that the UCBSC therapy is modulating the microglia/macrophage population to promote a pro-recovery M2 phenotype. This modulation of the immune system occurs even when the UCBSC are administered 2 days after the ischemic injury, significantly expanding the therapeutic window currently available for tPA treatment. This study provides insights into understanding the cellular mechanisms of stroke and its treatment with UCBSC. The cellular mechanisms underlying the therapeutic benefit of these stem cells provides a basis for future clinical studies evaluating the safety and efficacy of these cells in treating patients with ischemic brain injury.
Supplemental Material
Supplemental_Material - Immunomodulation with Human Umbilical Cord Blood Stem Cells Ameliorates Ischemic Brain Injury – A Brain Transcriptome Profiling Analysis
Supplemental_Material for Immunomodulation with Human Umbilical Cord Blood Stem Cells Ameliorates Ischemic Brain Injury – A Brain Transcriptome Profiling Analysis by Maple L. Shiao, Ce Yuan, Andrew T. Crane, Joseph P. Voth, Mario Juliano, Laura L. Hocum Stone, Zhenghong Nan, Ying Zhang, Nicole Kuzmin-Nichols, Paul R. Sanberg, Andrew W. Grande and Walter C. Low in Cell Transplantation
Supplemental Material
Supplementary_figure_1 - Immunomodulation with Human Umbilical Cord Blood Stem Cells Ameliorates Ischemic Brain Injury – A Brain Transcriptome Profiling Analysis
Supplementary_figure_1 for Immunomodulation with Human Umbilical Cord Blood Stem Cells Ameliorates Ischemic Brain Injury – A Brain Transcriptome Profiling Analysis by Maple L. Shiao, Ce Yuan, Andrew T. Crane, Joseph P. Voth, Mario Juliano, Laura L. Hocum Stone, Zhenghong Nan, Ying Zhang, Nicole Kuzmin-Nichols, Paul R. Sanberg, Andrew W. Grande and Walter C. Low in Cell Transplantation
Footnotes
Acknowledgment
We thank Kyle Schaible for editorial assistance.
Ethical Approval
This study was approved by the Institutional Animal Care and Use Committee (IACUC) at the University of Minnesota.
Statement of Human and Animal Rights
All experimental procedures involving animals were conducted in accordance with the IACUC guidelines at the University of Minnesota.
Statement of Informed Consent
There are no human subjects in this article and informed consent is not applicable.
Declaration of Conflicting Interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: P.R. Sanberg and W.C. Low are founder and consultant, respectively, of Saneron CCEL Therapeutics, Inc., and are inventors on patents related to cord blood. N. Kuzmin-Nichols is an inventor on patents related to cord blood. PRS is the co-editor-in-chief of Cell Transplantation. Neither PRS nor any member of the editorial office or editorial board affiliated the authors’ institutions was involved with the review process and/or decision making of this manuscript. The remaining authors declare no conflict of interest.
Funding
The author(s) disclosed receipt of the following financial support for the research and/or authorship of this article: This work was supported in part by funds from NIH grant R41-NS056626, and from Suzanne M. Schwarz, and Walter and Ginger Bailey. Ce Yuan is supported by the MnDrive-UMII Graduate Fellowship.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.