
Abstract
Traumatic brain injury (TBI) is a devastating disorder causing negative outcomes in millions of people each year. Despite the alarming number of brain injuries and the long-term detrimental outcomes that can be associated with TBI, treatment options are lacking. Extensive investigation is underway, in hopes of identifying effective treatment strategies. Among the most state-of-the-art strategies is cell replacement therapy. TBI is a seemingly good candidate for cell replacement studies because there is often loss of neurons. However, translation of this therapy has not yet been successful. It is possible that a better understanding of endogenous neurogenic mechanisms after TBI could lead to more efficacious study designs using exogenous cell replacement strategies. Therefore, this study was designed to examine the number and migration of immature neurons at 1 and 7 d after a fluid percussion TBI. The results show that the number of immature neurons increases from 7 d after a fluid percussion injury (FPI), and there is ectopic migration of doublecortin (DCX+) immature neurons into the hilar region of the dentate gyrus. These results add important data to the current understanding of the endogenous neurogenic niche after TBI. Follow-up studies are needed to better understand the functional significance of elevated neurogenesis and aberrant migration into the hilus.
Introduction
Traumatic brain injury (TBI) is a major cause of death and disability in the United States and throughout the world. Treatment options are lacking for those who suffer from subsequent neurological symptoms, some of which can be lifelong. TBI is a complex injury with numerous symptoms and disabilities that can have a devastating impact on the individual, as well as an enormous socioeconomic toll. Despite the alarmingly high incidence rates, treatment options are lacking, and a full mechanistic understanding of the various types of TBIs remains elusive.
There are a number of different experimental therapies for treating TBI. These include pharmacological, steroidal, anti-inflammatory, cellular replacement, and behavioral methods. 1 –5 Although a number of compounds have been tested in phase 3 clinical trials, none have proven to be efficacious at significantly improving outcomes. Advances in technology bring forth new treatment options that are being readily explored in experimental and clinical settings.
One of these new technologies, stem cell therapy, is being investigated to treat TBI as well as a number of other neuropathologies. 3,6 –8 One of the reasons that stem cell therapy is such an appealing option for TBI is that TBI can be associated with a loss of neurons, both at the site of the injury and sites distal to the injury. In the latter case, the neuronal loss is often the result of counter-coup forces and/or Wallerian degeneration. Thus, it is believed that some of the resultant cognitive deficits may be explained by the loss of neurons and that replacing these neurons using exogenous stem cells or stimulating endogenous neuronal growth could improve or restore function.
Major obstacles have thus far prevented successful clinical implementation of stem cell therapy for TBI or other disorders. For example, after a TBI, there is release of a poorly understood milieu of inflammatory proteins as well as a number of other biochemical changes in the central nervous system. Moreover, despite a growing number of studies on the subject, 9–10 there is still a lack of understanding of how TBI influences the endogenous neuronal stem cell population, the understanding of which could help further stem cell therapy. Several studies using different animal models of TBI have demonstrated an alteration to adult neurogenesis following TBI. 11,12 Such alterations include increases and/or decreases to the number and survival of newborn neurons, 11,12 as well as aberrant growth and integration of the newborn neurons into the existing circuitry. 11,13,14 Therefore, understanding how TBI alters the endogenous neurogenic niche could improve the strategies of exogenous stem cell treatments.
One area of interest in TBI is the hippocampus. The hippocampus is an important structure involved in, among other things, learning and memory. The hippocampus is an area of interest because neurogenesis continues in the hippocampus well into the adulthood. The importance of this adult hippocampal neurogenesis is not entirely understood. However, manipulations that have positively altered adult hippocampal neurogenesis, such as exercise and an enriched environment, have resulted in improvements to learning, memory, spatial awareness, and affect. Neurogenic changes have also been linked to detrimental outcomes, including depression, fear responsiveness, increased seizure susceptibility, seizure generation, seizure facilitation, and the potential for development of epilepsy 15 ; development of epilepsy may possibly be due to improper migration and/or integration of new neurons into the existing hippocampal circuitry. 11,15 –17 In order to better understand the temporal effects of TBI on neurogenesis in the hippocampus, we used the fluid percussion injury (FPI) model and examined the quantity and location of newborn neurons in the hippocampus at 1 and 7 d after FPI or after a sham FPI. The hippocampi from these mice were processed for immunohistochemistry and labeled for immature neurons using antidoublecortin (DCX); the number of these cells was quantified, as were DCX-labeled cells in the hilus.
Materials and Methods
Mice
Six-week-old male, C57Bl/6 (Jackson Laboratories, Bar Harbor, ME) mice (N = 16) were divided into 2 groups: FPI (N = 8) and sham (N = 8). Mice in each of these groups were randomly assigned to either the 1 d (N = 8) or the 7 d (N = 8) postinjury groups, such that each group at each time point had 4 mice. All protocols were approved by the Baylor Scott and White institutional animal care and use committee (IACUC).
FPI
All mice underwent surgery as previously described. 11,18,19 Briefly, a 2-mm hole was drilled, with dura intact, into the skull over the left parietal cortex (anteroposterior: +1.5 mm; mediolateral: −1.2 mm). A female luer lock was connected to the hole in the skull. Animals in the FPI group received a pressure pulse (1.5 to 1.7 atm) from the FPI apparatus. Sham animals received identical treatment except that no pressure pulse was delivered.
Tissue Preparation
At 1 or 7 d after FPI, mice were sacrificed with an overdose of Euthasol® (Virbac, Forth Worth, TX, USA), followed by transcardial perfusions with saline and paraformaldehyde (PFA), as previously described. 20 After postfixing within the skull for 24 h, brains were removed and postfixed 48 h in PFA. Brains were then stored in phosphate-buffered saline (PBS) and cut in serial sections on a vibratome (Ted Pella Inc., Redding, CA) at 50 µm. We selected 4 to 5 slices containing the anterior two-thirds of the hippocampus from each animal. Using Paxinos Mouse Atlas, we determined the distance from bregma for each hippocampal section to ensure equal sampling throughout the anterior/posterior portions of the hippocampus.
DCX Immunohistochemistry
DCX immunohistochemistry was performed as previously described (Santa Cruz Biotechnology, Dallas, TX). 21 –23 Briefly, tissue was incubated, rotating in the dark at room temperature, for 24 h in PBS containing 0.005% Tween (Sigma-Aldrich, St. Louis, MO), 5% normal horse serum (Vector Labs, Burlingame, CA), and DCX antibodies to the N and C termini (1:500 each, Santa Cruz Biotechnology, Dallas, TX). The tissue slices were rinsed 3 times for 5 min each in 0.01 M PBS, then incubated for 90 min, rotating in the dark at room temperature in PBS containing 0.005% Tween, 5% normal horse serum, and fluorescently tagged donkey antigoat antibody (1:200; Alexa Flour 555; Invitrogen, Carlsbad, CA). After 90 min, the tissue slices were rinsed 3 times for 5 min each in 0.01 M PBS, mounted onto glass slides, allowed to dry overnight, and cover slips were applied with Fluoromount-G™ (EMD Millipore, Billerica, MA). The slides were coded, and the images of DCX labeling in the dentate gyrus were captured by a rater blind to the condition of the mice. The coded images were given to a second rater who performed the physical cell counts.
Quantification of DCX+ Cells
DCX-labeled cells were quantified in the subgranular zone (the granule cell layer and the border between the hilus and granule cell layer) as previously described. 11 Briefly, grids of 5,000 μm2 boxes (50 × 100 μm) were superimposed along the hilar/granule cell layer border. Each grid was numbered, and a random number generator was used to randomly select ∼ 60% of the grids from within which to count DCX-labeled cells. The number of grids to be counted was determined using the StereoInvestigator software (MBF Bioscience Inc., Williston VT). These principles are consistent with stereological quantification and ensure random and unbiased sampling throughout the tissue. A 1-way analysis of variance was performed (SPSS V. 9; IBM, Armonk, NY), with post hoc Bonferroni analysis to examine differences between groups after FPI. It is pertinent to note that the analysis was initially performed separately for each hemisphere. Analysis comparing the ipsilateral to the contralateral hemisphere revealed no significant differences (not shown). Therefore, ipsilateral and contralateral hemispheres were combined, and the data presented are comprised of bilateral changes in DCX+ cell numbers.
Results
DCX-labeled Cells Are Significantly Increased in the Dentate Gyrus 7 D After FPI
The results demonstrated a trend toward an increase in DCX labeling at 1 d after FPI (P = 0.0806), compared to sham (Fig. 1a to c), and a significant increase in DCX-labeled cells in the dentate gyrus at 7 d after FPI (P < 0.05), compared to sham (Fig 1a to c).
Doublecortin (DCX) labeling in the hippocampal dentate gyrus at 1 d after fluid percussion injury (FPI). (a) A sham mouse is shown to represent the normal staining pattern of DCX+ cells in the dentate gyrus. (b) DCX labeling is shown from a mouse at 7 d after FPI to illustrate an increase in immature neurons at this time point after FPI. (c) A graph of the mean number of DCX+ cells is shown. As can be seen in this figure, the number of DCX+ cells is increasing 1 d after FPI, and the increase is significant at 7 d after FPI. *P < 0.05. Scale bars = 25 µm.
Ectopic Cells Are Increased in the Hilus at 7 D after FPI
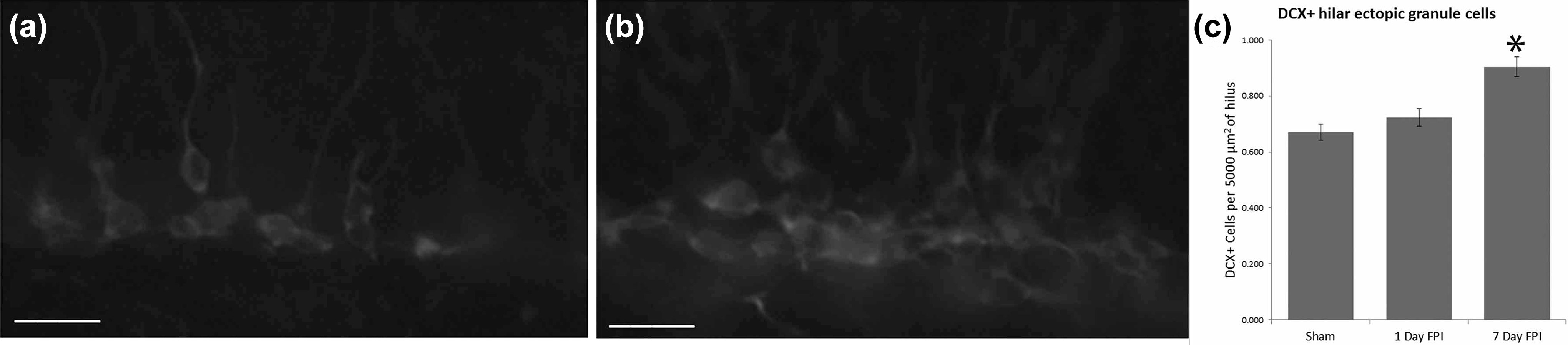
The results demonstrated an increased number of DCX-labeled cells in the hilus at 7 d after FPI (P < 0.04), compared to sham (Fig. 2a to c). There was no significant difference at 1 d after FPI (Fig. 2a to c).

Hilar ectopic cells in the dentate gyrus after fluid percussion injury (FPI). (a) A representative photomicrograph showing a sham mouse at 7 d after FPI. (b) A photomicrograph is provided to illustrate an increase in the number of DCX+ cells, as well many hilar ectopic cells (arrowheads) in the dentate gyrus of a mouse at 7 d after FPI. (c) A graph of the mean number of hilar basal dendrites reveals a significant increase in the number of hilar ectopic granule cells at 7 d after FPI. *P < 0.04. Scale bars = 50 µm in (a) and (b).
Discussion
The results from this study demonstrate that an FPI in mice results in an increase in the number of DCX-labeled cells in the dentate gyrus, as well as an increase in the appearance of DCX-labeled hilar ectopic cells. The finding of an early increase in neurogenesis after an FPI is consistent with findings from several different models of TBI, 12,24 –26 and the observation of hilar ectopic cells within a week of an FPI in mice is novel.
Several studies have previously demonstrated altered numbers of immature cells in the dentate gyrus in different models TBI. 12,24 –26 Notably, the severity and/or type of injury was found to impact differential changes in hippocampal neurogenesis, 12 and altered neurogenesis was found to be associated with cognitive impairment. 27 –29 Interestingly, Robinson et al. 11 used the same model as in the current study and found no significant differences in the DCX-labeled cells in the dentate gyrus at 30 d after an FPI. 11 There are several possible explanations for this discrepancy. First, it is possible that while there is an initial increase in neurogenesis after FPI, many of these cells either do not survive or become functionally integrated. Previous studies have demonstrated that TBI alters the functional integration of newborn neurons. 13 From the context of cell replacement, it is important to understand the expected functional survival of the endogenous population of newborn neurons. It would similarly be important to understand approximately what percentage of the newborn neurons die, so that more accurate survival expectations can be formulated. Studies that incorporate a retroviral vector to label newborn neurons, and/or bromodeoxyuridine labeling, combined with anti-neuronal nuclei (NeuN) double labeling at 30+ d after FPI (the duration of time for integration and maturation of newborn neurons) could further address this possibility.
Another possible explanation for the discrepancy in neurogenesis at 7 and 30 d post-FPI could be that the increased neurogenesis observed 7 d after FPI is transient and returns to near baseline levels by 30 d after FPI. In this case, it is important to understand whether, at time points later than 30 d, neurogenesis is significantly decreased. Indeed, previous studies in epileptogenesis models have demonstrated early increases, followed by long-term deficits in neurogenesis that are associated with behavioral impairments. 30 From a translational perspective, these latter studies would be extremely important for determining at what time point it would be most advantageous to consider cell transplantation or other methods to stimulate endogenous neurogenesis and/or methods to ensure better survival/integration of the newly born cells.
Previous studies have demonstrated a rapid inflammatory response in the hippocampus in this model of TBI. 31 It is possible that rapid hippocampal inflammation plays a role in the changes to neurogenesis. Such changes could include increased survival factors, such as cytokines and chemokines that can also serve as growth factors. Considering that a vast majority of newborn granule cells will die before becoming functionally integrated into circuitry, 11 it is possible that the elevated cytokines and chemokines might prolong the survival of some of these recently born neurons. A similar hypothesis has been previously postulated in models of epilepsy, in which hippocampal neurogenesis is also initially increased. 15 –17 Such enhanced survival might also contribute to aberrant hippocampal circuit formation. 15 –17 Considering that we have recently demonstrated a TBI-induced alteration to the radial glial-like processes in the dentate gyrus and that these cells provide a scaffold for the normal growth and integration of newborn granule cells, 11,21,22 it is possible that the alterations we have observed to hippocampal neurogenesis are related to hippocampal inflammation, including the activation of the radial glial-like astrocytes.
The observation of an increase in DCX+ hilar ectopic cells illustrates another important consideration for cellular transplantation studies. The possibility that cells migrate into aberrant regions or circuits must be addressed. In the case of seizures, hilar ectopic granule cells have been demonstrated to contribute to a hyperexcitable recurrent hippocampal circuitry. 32,33 This circuit has also been demonstrated anatomically 15 –17 and in previous studies using the same FPI model. It has been demonstrated that this model has anatomical alterations consistent with aberrant hippocampal circuitry. 11 Interestingly, the FPI model used in the present study has also been shown to have increased seizure susceptibility 30 d after FPI. 18 Therefore, aberrant growth and integration of newborn neurons, whether endogenous or transplanted, can contribute to abnormal brain functioning, further underscoring the necessity to fully elucidate endogenous neurogenic processes.
When transplanting cells into a normal brain, there is a relatively reasonable expectation that the implanted cells will grow along the established, normal brain circuitry. However, cell transplantations are only required in cases of extreme neuropathology. Therefore, studies that provide insight into the endogenous neurogenic processes in disease models are important for providing experimental evidence for optimization of stem cell therapy strategies. The current study demonstrates an increased number and ectopic growth of newborn neurons 7 d after FPI. Follow-up studies are required to fully appreciate the timing of integration as well as molecular and/or anatomical cues that promote both normal and ectopic growth of newborn neurons.
Footnotes
Acknowledgments
The author would like to thank Sanjib Mukherjee and Kyle Wade for their technical support.
Ethical Approval
The protocols in this study were approved by the relevant ethics committee (see Materials and Methods).
Statement of Informed Consent
There are no human subjects in this article and informed consent is not applicable.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, and/or authorship of this article.