
Abstract
Objective:
To investigate how nuclear factor-E2-related factor 2 (Nrf2) involved in the protective effect of isoflurane (Iso) preconditioning in oxygen glucose deprivation (OGD)-induced cortical neuron injury.
Methods:
Primary mouse cortical neurons were divided into Control, ML385 (an Nrf2 inhibitor), Iso, Iso + ML385, OGD, ML385 + OGD, Iso + OGD, and Iso + ML385 + OGD groups. Lactate dehydrogenase activity (LDH) release and oxidative stress indexes were quantified. 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay was used to detect cell viability, Annexin V-FITC/propidium iodide (PI) staining to measure cell apoptosis, dichloro-dihydro-fluorescein diacetate (DCFH-DA) method to test reactive oxygen species (ROS), and quantitative reverse-transcription polymerase chain reaction (qRT-PCR) and Western blotting to evaluate genes and protein expression.
Results:
Iso preconditioning reduced LDH release and inhibited cell cytotoxicity in OGD-induced cortical neurons, which was abolished by ML385. Iso preconditioning increased the Nrf2 nuclear translocation in cortical neurons. Meanwhile, Iso decreased the OGD-induced apoptosis with the down-regulations of Bax and Caspase-3 and the up-regulation of Bcl-2, which was reversed by ML385. OGD enhanced the level of ROS and malondialdehyde (MDA) in cortical neurons, but reduced the activities of superoxide dismutase (SOD) and glutathione peroxidase (GSH-Px), which were aggravated in ML385 + OGD group and mitigated in Iso + OGD group. No observable difference was found between OGD group and Iso + ML385 + OGD group regarding apoptosis-related proteins and oxidative stress-related indexes.
Conclusion:
Iso preconditioning up-regulated Nrf2 level to play its protective role in OGD-induced mouse cortical neuron injury.
Introduction
Cerebral stroke is the second leading cause of human death and the third leading cause of human disability around the world, posing a heavy burden on patients and their families. 1 Generally speaking, this disease mainly includes two types, ischemic stroke and hemorrhagic stroke, with the former accounting for approximately 80% of stroke incidence. 2 The occurrence of ischemic stroke results from brain tissue necrosis induced by the lack of blood and oxygen supply to the brain. 3 In the prevention and treatment of cerebral ischemia, Isoflurane (Iso) preconditioning has been widely applied because of its feasibility, noninvasiveness and obvious curative effects. 4 Commonly, Iso is considered as a common volatile anesthetic having preconditioning and post-conditioning effects on multiple organs, including brain, 5 and its preconditioning has been shown to alleviate the neurological deficit, brain edema and infarct volume of ischemic animal model. 5 Studies also found the neuro-protective involvement of genes in the Iso preconditioning. 6,7
Nuclear factor-E2-related factor 2 (Nrf2), a member of cap’ n’ collagen family has highly conserved leucine zipper motif, and is well-established as an important endogenous antioxidant factor widely distributed in various organs of the body. 8 When Nrf2 is stimulated by oxidative stress or nucleophiles, it will dissociate from Kelch-like ECH-associated protein 1 (Keap1) and enter into the nucleus, in which Nrf2 binds to the antioxidant response element (ARE) in the target gene promoter to facilitate the expression of antioxidant enzymes. 9 The oxygen glucose deprivation (OGD) in vitro, aiming to remove oxygen and glucose and simulating the process of brain ischemia neuron injury, has become the most classical model in the study of ischemic brain injury toward providing reliable and technically straightforward evidence. 6,10 In recent years, accumulating evidence supported the important role of Nrf2 in OGD-induced neuron injury. 11 –13 Additionally, emulsified Iso was found to be able to affect reactive oxygen species (ROS) and activate Nrf2-antioxidant response element signaling pathway, and thereby alleviating the myocardial injury caused by ischemia-reperfusion in mice. 14 To date, several inhibitors of the Nrf2 have been identified, such as ML385, Brucea chinensis, clonazepa propionate, luteolin, all-trans retinoic acid, and flavonoid molecular compounds. 15 Among them, ML385 has been widely used in the neuroprotective effect. 16,17 However, it is still unknown whether Nrf2 can affect the protective role of Iso preconditioning in OGD-induced cortical neuron injury. Hence, this study was conducted to find out the possible answer.
Materials and methods
Ethics statement
This study was conducted in compliance with Guide for the Care and Use of Laboratory Animals. 18 All animal experiments were performed under the supervision of Medical Ethics Committee of Laboratory Animals in our hospital. C57BL/6J mice were purchased from Jackson Laboratory (Bar Harbor, ME) and kept under a 12 h light/12 h darkness cycles with sufficient supply of water and food.
Primary culture of mouse cortical neurons
Neonatal mice (1–3 days after birth) were sacrificed to isolate their cortex, which was cut up and placed in improved DMEM medium (80% Dulbecco’s modified eagle medium with 10% fetal bovine serum, 10% F12 medium, and 10 g/mL Pen/Strep). Cortex was digested by trypsin for 25 min, mixed with 50% horse serum, and ground by a fire-polished Pasteur pipette. Cell suspension was centrifuged for 3 min, filtered (Steriflip100 M, Millipore, Billerica, MA) and transferred to polylysine-coated wells. For immunofluorescence microscopy, polylysine-coated glass coverslips were employed to plate cells. Incubation in vitro was conducted at 37°C with 5% CO2 and 95% air for 10–12 days. There were eight groups: Control, ML385, Iso, Iso + ML385, OGD, Iso + OGD, ML385 + OGD, and Iso + ML385 + OGD.
Iso preconditioning
For Iso preconditioning, cortical neurons (for 10–12 days in vitro) were placed in a humidified exposure chamber with an inlet and an outlet for the anesthetic mixture. 19 The exposure chamber was placed in an incubator at 37°C. The mixture 95% air/5% CO2 was transported into an Isotec 3 Iso vaporizer (GE Healthcare, Waukesha, WA) so that the 1.5% Iso/93.5% air/5% CO2 can be delivered to experiment box by rate of 0.4 L/min for 3 h. Following Iso preconditioning, neurons were washed, 1× fresh medium was added, and cells were transferred into a standard humidified CO2 incubator. As for neurons in Control, ML385, OGD, and ML385 + OGD groups, the medium replacement and/or replenishment were conducted in line with preconditioned cultures.
Establishment of OGD-induced injury models
OGD-induced injury models were established at 24 h after Iso preconditioning. 6 For OGD process, the culture medium was replaced by glucose-free buffer, which was pretreated for 5 min in 100% N2. Neuron cells were exposed to 100% N2 for 10 min in an airtight chamber, which was closed and placed at 37°C for 2 h at the condition of hypoxia (<1% oxygen content). Cells in Control group were incubated in the same conditions except for the existence of 4.5 g/L glucose and normoxic condition. After induction by OGD, glucose-free buffer was replaced with fresh culture medium, which was kept under normal conditions for 24 h. Nrf2 inhibitor ML385 (7.5 μM) was added before the performance of OGD until 24 h after OGD.
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay
Cells were inoculated onto 96-well plates by 1 × 104 cells per well. The culture medium was removed, 200 µL of serum-free culture medium was added, and 20 µL of MTT (Code No: M6494, Invitrogen™, USA) was added for 4 h of incubation, followed by the addition of 200 µL dimethyl sulfoxide (DMSO) and 10 min of oscillation. Automatic enzyme-linked immunosorbent assay (ELISA) system was used to determine the absorbance value at the wavelength of 570 nm (OD570 nm). The experiment was repeated three times.
Lactate dehydrogenase activity (LDH) assay
The LDH cytotoxicity assay (Code No: C20300, Invitrogen™, USA), a colorimetric assay that provides a simple and reliable method for determining cellular cytotoxicity, was used to measure the LDH. LDH content in culture medium (released LDH) and in neuron cells was determined. Intracellular LDH was measured after the cells were lysed by 0.5% Triton X-100TM solution) was quantified. The percentage of released LDH in the total LDH (released LDH plus intracellular LDH) was calculated and used to reflect the cell injury.
Annexin V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) staining
The cell apoptosis was detected by Annexin-V-FITC/PI (Code No: V13242, Invitrogen™, USA). In brief, cells to be detected were adjusted to density of 1 × 106 cells/mL. Next, 200 µL cell suspension was taken for centrifugation at 1000 rpm × 5 min (4°C), washed with pre-cooled PBS (1 mL) twice, and centrifuged again at 1000 rpm × 5 min (4°C). Then, cells were suspended in 100 µL binding buffer, mixed with 2 µL Annexin-V-FITC (20 μg/mL), placed on ice for 15 min, and transferred to testing tube with 400 µL PBS. Before loading, 1 µL PI (50 μg/mL) was added into each sample and detection was done within 2 min. In the scatter diagram of the flow cytometer, the right upper quadrant necrotic cells (FITC+/PI+), and right lower quadrant apoptotic cells (FITC+/PI−).
Quantitative reverse-transcription polymerase chain reaction (qRT-PCR)
TRIzol™ reagent (Code No: 15596026, Invitrogen™, USA) was employed to extract total RNA in each group. The primers were synthesized by Takara Biomedical Technology Co., Ltd. (Beijing, China) and were listed in Table 1. PrimeScript™ RT Master Mix (Code No: RR036Q, Takara Biomedical Technology Co., Ltd., Beijing, China) was used for reverse transcription of RNA to cDNA (50 ng/μL). Reaction solution was taken for qRT-PCR according to instructions of One Step TB Green® PrimeScript™ RT-PCR Kit (Code No: RR066A, Takara Biomedical Technology Co., Ltd., Beijing, China) . The relative mRNA expression target genes were calculated using 2− △△Ct method with GAPDH as reference gene.
Primers sequences used in qRT-PCR.

Western blotting
Total protein in cortical neurons was extracted, quantified for protein concentration with a Pierce™ BCA protein assay kit (Code No: 23225, Thermo Scientific™, USA), and adjusted for loading volume with deionized water. Separateion gel and concentrated gel were prepared using 10% SDS. Samples and loading buffer were mixed, boiled for 5 min at 100°C, bathed in icy water, centrifuged, and loaded to each lane with pipettes for electrophoresis sepamouseion. The protein on the gel was transferred to the cellulose nitmousee membrane, which was blocked in 5% skimmed milk for 1 h. Next, primary antibody was added for overnight incubation at 4°C, including anti-Nrf2 antibody (ab137550, 1/1000 dilution), anti-Histone H3 antibody—nuclear loading control (ab1791, 1/1000 dilution), anti-Bax antibody (ab32503, 1/2000 dilution), anti-cleaved Caspase-3 antibody (ab2302, 1 µg/ml), anti-Bcl-2 antibody (ab59348, 1/500 dilution), and anti-GAPDH antibody-loading control (ab9485, 1/2500 dilution). Then, membrane was washed with PBS at room temperature for 3 times × 5 min, followed by addition of secondary antibody Goat Anti-Rabbit IgG H&L (ab175781, 1:10000 dilution) for 1 h of incubation at room temperature. The membrane was washed again with PBS at room temperature for 3 times × 5 min and soaked in ECL reaction solution (Pierce, USA) for 1 min at room temperature. Subsequently, the liquid solution was removed and the membrane was covered by cling film for exposure and development in a dark environment. All antibodies were purchased from Abcam (USA).
Dichloro-dihydro-fluorescein diacetate (DCFH-DA) assay measures intracellular reactive oxygen species (ROS)
Cortical neurons were collected at logarithmic growth phase, washed with PBS, and mixed with 10 μmol/L DCFH-DA serum-free medium (200 μl/well, Code No: HY-D0940, MedChemExpress, China), which should cover all bottom-level cells. Neurons were cultured for 30 min at 37°C with 5% CO2, washed with PBS once, digested by trypsin, and collected for centrifugation at the mouse of 1200 r/min (5 min). Next, neurons were washed twice with PBS, suspended in 300 µl PBS, and detected using a flow cytometer (Beckman coulter, USA) (excitation wavelength 488 nm and emission wavelength 525 nm).
Quantification of oxidative stress indexes (MDA, SOD and GSH-Px)
The detection kits from Abcam (USA) were used to quantify malondialdehyde (MDA, Code No: ab118970), superoxide dismutase (SOD, Code No: ab65354) and glutathione peroxidase (GSH-Px, Code No: ab138881) of cortical neurons.
Statistical methods
Statistical analysis was conducted using SPSS 21.0 (SPSS, Inc, Chicago, IL, USA). Measurement data were expressed as mean ± standard deviation (SD). All experiments were repeated three times using three replicate wells in each independent experiment. Comparison among multiple groups was analyzed using one-way ANOVA, while inter-group difference was compared using Turkey HSD’s test. P < 0.05 was regarded as statistical significance.
Results
Released LDH and cell viability of cortical neurons in each group
As shown in Figure 1(a), compared with Control group, cortical neurons in OGD groups had the obvious LDH release, especially in ML385 + OGD group (all P < 0.05). Iso preconditioning reduced the LDH release in OGD-induced cortical neurons, which was reversed by Nrf2 inhibitor ML385 (both P < 0.05). Cortical neuron viability was detected using MTT assay (Figure 1(b)). According to the results, Iso preconditioning significantly increased the viability of OGD-induced cortical neurons, while ML385 can further aggravate the decreased viability of OGD-induced cortical neurons (both P < 0.05). OGD group and Iso + ML385 + OGD group presented no significant difference in terms of cell viability (P > 0.05). Besides, the released LDH and cell viability of cortical neurons were not statistical differences among the Control, ML385, Iso, and Iso + ML385 groups.

LDH release (a) and cell viability (b) of cortical neurons in each group. The same letters showed no statistical significance, P > 0.05, while different letters indicated the statistical significance, P < 0.05. Data was expressed as mean ± standard deviation (SD) of three replicate wells per condition from three independent experiments.
Nrf2 expression in cortical neurons of each group
Western blotting was employed to detect Nrf2 protein expression in cortical neurons (Figure 2(a)). As a result, Iso could enhance the translocation of Nrf2 into nucleus, which was abolished by ML385. Furthermore, OGD-induced cortical neurons had decreased translocation of Nrf2 into nucleus as compared the Control group. ML385 further reduce the translocation of Nrf2 into nucleus in OGD-induced cortical neurons, but Iso preconditioning increased the translocation of Nrf2 into nucleus in OGD-induced cortical neurons (P < 0.05). The mRNA expression of Nrf2 was detected by qRT-PCR (Figure 2(b)). Accordingly, OGD inhibited Nrf2 mRNA expression in cortical neurons, and Iso preconditioning and ML385 up-regulated and down-regulated Nrf2 mRNA expression, respectively (P < 0.05). Compared with OGD group, Iso + ML385 + OGD group presented no significant difference regarding the Nrf2 mRNA expression in cortical neurons (P > 0.05).

Nrf2 expressions in the cortical neurons of each group. (a) Nrf2 expression in nucleus measured by Western blotting. (b) Nrf2 mRNA expression quantified by qRT-PCR. The same letters showed no statistical significance, P > 0.05, while different letters indicated the statistical significance, P < 0.05. Data was expressed as mean ± standard deviation (SD) of three replicate wells per condition from three independent experiments.
Apoptosis of cortical neurons in each group
Annexin V-FITC/PI staining was performed to evaluate the apoptosis of cortical neurons (Figure 3). Compared with the groups not induced by OGD, the other OGD-induced groups were enhanced in cortical neuron apoptosis (all P < 0.05). Compared with OGD group, ML385 + OGD group increased while Iso + OGD group decreased significantly in cortical neuron apoptosis (both P < 0.05). However, Iso + ML385 + OGD group showed no significant difference concerning the apoptosis as compared with the OGD group (P > 0.05).

Apoptosis of cortical neurons detected after Annexin V-FITC/PI staining. The same letters showed no statistical significance, P > 0.05, while different letters indicated the statistical significance, P < 0.05. Data was expressed as mean ± standard deviation (SD) of three replicate wells per condition from three independent experiments.
Expression of apoptosis-related proteins in cortical neurons of each group
The result of qRT-PCR detection was presented in Figure 4(a) to (c). OGD can significantly induce the up-regulated mRNA expressions of Bax and Caspase-3 and decrease the Bcl-2 expression in cortical neurons (all P < 0.05), which was most significant in ML385 + OGD group. Iso preconditioning significantly improved the expression of apoptosis-related genes in OGD-induced neurons with the down-regulations of Bax and Caspase-3 and the up-regulation of Bcl-2. Compared with OGD group, Iso + ML385 + OGD group showed no significant difference in Bax, Caspase-3 and Bcl-2 mRNA expression. Besides, no statistical difference in these mRNA expressions of apoptosis-related genes was found among the Control, ML385, Iso, and Iso + ML385 groups. Western blotting analysis demonstrated the protein expression of Bax, Cleaved-Caspase-3, and Bcl-2 in consistency with their mRNA expression (Figure 4(d) to (g)).
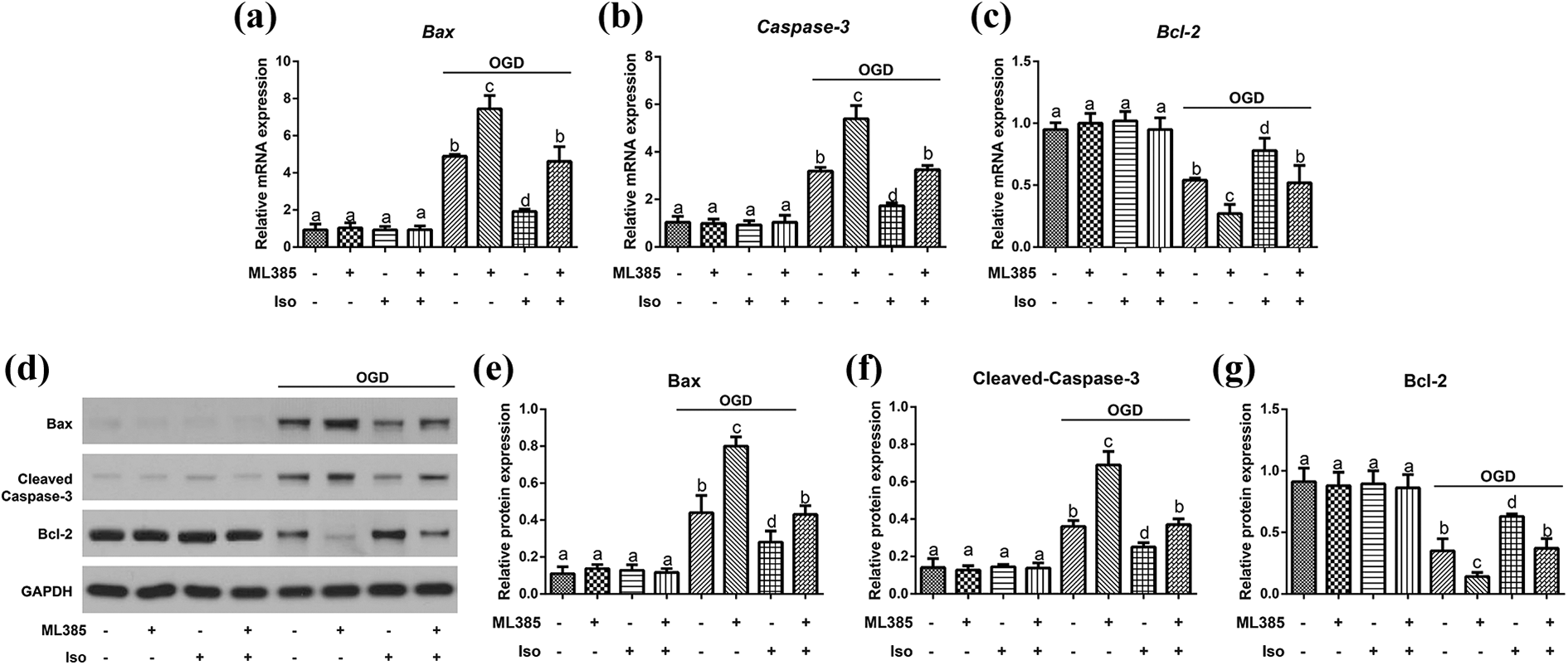
Expression of apoptosis-related proteins in cortical neurons of each group. (a to c) The mRNA expression of Bax, Caspase-3 and Bcl-2 measured by qRT-PCR. (d to g) The protein expression of Bax, Cleaved-Caspase-3 and Bcl-2 quantified by Western blotting. The same letters showed no statistical significance, P > 0.05, while different letters indicated the statistical significance, P < 0.05. Data was expressed as mean ± standard deviation (SD) of three replicate wells per condition from three independent experiments.
Oxidative stress of cortical neurons in each group
As illustrated in Figure 5, the groups not induced by OGD showed no significant difference in the oxidative stress indexes, including ROS, MDA, SOD and GSH-Px. Compared with Control group, OGD group increased apparently in ROS content of cortical neurons (P < 0.05), but Iso preconditioning apparently reduced ROS content (P < 0.05). Compared with OGD group, ML385 + OGD group was further elevated in cortical neuron ROS content (P < 0.05), while Iso + ML385 + OGD group presented no obvious difference in ROS content (P > 0.05). Additionally, OGD can induce the increase of MDA content and the decreases of SOD and GSH-Px activities in cortical neurons, especially in ML385 + OGD group, which, however, was mitigated in Iso + OGD group (all P < 0.05). OGD group and Iso + ML385 + OGD group didn’t show significant difference regarding activity of SOD and GSH-Px and content of MDA (all P > 0.05).

Oxidative stress of cortical neurons in each group. ROS content (a) in cortical neurons assessed using DCFH-DA assay. ROS (b), MDA (c), SOD (d) and GSH-Px (e) in cortical neurons of each group. The same letters showed no statistical significance, P > 0.05, while different letters indicated the statistical significance, P < 0.05. Data was expressed as mean ± standard deviation (SD) of three replicate wells per condition from three independent experiments.
Discussion
Cerebral ischemia can lead to the generation of a great quantity of oxygen free radicals, the inactivation of multifunctional enzyme system, and the consumption of a great deal of antioxidants, thus causing damage to normal antioxidant defense capacity; however, inhalation anesthetics can significantly reduce the production of free radicals and inhibit lipid peroxidation to play a protective role in cerebral ischemia. 20 Pretreatment is to give a sub-lethal stimulation before noxious stimulation of tissues, inducing the activation of endogenous protection mechanism, which is a kind of protective strategy that can increase the tolerance of tissues or organs to the stimulation and injury after antagonizing. 10,21 LDH is abundant in cytoplasm and it cannot pass through the cell membrane under the normal conditions, but it can be released to the outside in damaged or dead cells. 22 In our study, Iso preconditioning can significantly reduce LDH release in OGD-induced cortical neurons, increase cell viability, alleviate cell oxidative stress, and decrease cell apoptosis, which was consistent with the findings from previous studies and further demonstrated the neuro-protective role of Iso preconditioning in cerebral ischemia. 23 –25
As an important nuclear transcription factor, Nrf-2 has been found critically involved in many diseases, including cerebral ischemia, since it participates in oxidative stress, regulates the antioxidant enzymes and phase II detoxification enzymes, as well as affects cell resistance to foreign chemicals and oxidative damage. 26,27 As for the activation mechanism of Nrf2, it was previously believed that electrophilic substances or ROS can decouple Nrf2 from Keap1 by modifying cysteine residues of Keap1, and free Nrf2 could enter the nucleus to activate Nrf2 signaling pathway. 28 In our study, we did observe the increased translocation of Nrf2 into nucleus in cortical neurons induced with or without OGD after Iso preconditioning. Meanwhile, the ML385 as an Nrf2 inhibitor could further reduce the Nrf2 expression in OGD-induced cortical neurons, which was also revered by the Iso preconditioning, demonstrating that Iso preconditioning could overcome the ML385 inhibition of Nrf2. The mentioned above suggested the protection of OGD-induced cortical neurons may be due to Iso-induced upregulation of Nrf2. Similarly, Park and his group also found the down-regulated Nrf2 protein level in cytoplasm and the up-regulated Nrf2 protein level in nucleus after general anesthesia with Iso, 29 showing that Iso preconditioning can play its regulatory role by affecting Nrf2 expression. Besides, Zhong et al. revealed Iso preconditioning could also suppress OGD-induced injury in hepatocytes through the regulation of Nrf2 signaling. 10 Moreover, treatment with Iso effectively increased the translocation of Nrf2 from cytoplasm to nucleus of mouse hippocampus. 29 ML385 is a specific Nrf2 inhibitor and it can directly interact with Nrf2 protein and bind to Neh1 domain of Nrf2 to block the integration of Nrf2-MAFG complex with promotor ARE sequence, eventually reducing the transcriptional activity of Nrf2. 29,30 In present study, ML385 can further increase LDH release in OGD-induced cortical neurons and reduce cell viability, demonstrating that Nrf2 inhibition can aggravate OGD-induced injury, but Iso preconditioning can ameliorate this phenomenon. In this regard, pro preconditioning may mediate Nrf2 to improve OGD-induced cortical neuron injury.
On the other side, the up-regulation of Nrf2 and the treatment with Nrf2 activator can significantly alleviate oxidative stress and inflammation in multiple diseases, like diabetic wound healing, 31 pulmonary injury 32 and anti-GBM glomerular nephritis. 33 Generally, ROS as a product of normal aerobic metabolism strikes a dynamic balance between its production and clearance, but the impairment of cell antioxidant mechanism can cause the accumulation of ROS and induce the cellular oxidative stress injury. 34 Malondialdehyde (MDA), a good marker of oxidative stress and free radical damage, can indirectly reflect the severity of free radical attack. 35 As we know, cells usually contain a variety of enzymes, such as catalase, GSH-Px, and SOD, which can act as a detoxifying system to inhibit ROS-induced cell injury. 36 In a previous study, the Nrf2 inhibitor ML385 was found able to partially eliminate the antioxidant effect of Metformin on spinal cord injury. 37 Also, ML385 could effectively reverse the protection of MaR1 in ulcerative colitis induced by dextran sulfate sodium through reducing MPO and ROS activity. 38 Here in this study, OGD-induced cortical neuron injury was aggravated by ML385 treatment, accompanied with the increased ROS and MDA content, as well as the decreased SOD and GSH-Px activities. However, in neurons pre-treated with Iso, the effect of ML385 was attenuated, which indicated that Iso preconditioning may affect Nrf2 to alleviate oxidative stress. As a matter of fact, accumulating evidence has demonstrated that oxidative stress can induce cell apoptosis, which was the most important pathological process after cerebral ischemia. 39 According to the study of Ma et al., the effect of Rosmarinic acid on cell apoptosis of SCI mice could be weakened by ML385. 17 Of note, Liu et al. also reported chlorogenic acid could ameliorate oxidative stress injury and apoptosis of cerebral cells and block the expression of apoptosis-related proteins (cleaved Caspase-3 and Caspase-9), which can also be affected by ML385. 40 To our knowledge, apoptosis is regulated by many genes, among which Bax and Bcl-2 are the two most important ones, as Bcl-2 gene family can control the release of apoptotic factors like cytochrome C in mitochondria and their over-expression can cause drug resistance of tumor cells. 41,42 Recently, Caspase-3 protein has attracted the attention of many researchers as apoptotic executioners, because it is one of the most important members of the caspase family. 43 In this study, ML385 could promote the apoptosis of OGD-induced cortical neurons with the up-regulated Bax and Caspase-3, and the reduced Bcl-2, which demonstrated that Iso preconditioning can mitigate OGD-induced cortical neuron apoptosis via regulating Nrf2.
In conclusion, Iso preconditioning can ameliorate the reduced translocation of Nrf2 into nucleus in OGD-induced cortical neurons. Nrf2 inhibitor ML385 can aggravate OGD-induced cortical neuron injury, which however can be reversed by Iso preconditioning, with decreased LDH release, increased cell viability, reduced cell apoptosis, and mitigated oxidative stress.
Footnotes
Acknowledgement
The authors appreciate the reviewers for their useful comments in this paper.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.