
Abstract
Since prehistoric times Coccinia grandis has been used as traditional medicine for various diseases including diabetes, dyslipidemia, metabolic and digestive disorders. Although the rationality of efficacy as natural antioxidants with different bioactive compounds in Coccinia grandis against monosodium glutamate (MSG) induced hepato-cardiac damage remains to be disclosed. Six different solvent extracts of the leaves of Coccinia grandis were chosen to evaluate in vitro antioxidant and free radical (FR)-scavenging activity. Due to high antioxidant content and FR-scavenging property of ethanol extract of Coccinia grandis leaves (EECGL) and presence of different bioactive compounds in EECGL was further tested to evaluate in vivo hepato-protective and cardio-protective efficacy against MSG-induced anomalies. MSG-induced dyslipidemia, increased cell toxicity markers altered functional status and histopathological peculiarities of target organs were blunted by EECGL. Additionally, MSG incited increase level of interleukin (IL)-6, tumour necrosis factor (TNF)-α, IL-1β which activates nuclear factor kappa-B (NF-kB) guided inflammation via down regulation of IL-10; impaired redox-homeostasis subsequently promoted inflammation associated oxidative stress (OS) and increased vascular endothelial growth factor (VEGF) which provoked microvascular proliferation related cellular damage. On the contrary, increased lipid peroxidation and nitric oxide promotes reduced cell viability, deoxyribonucleic acid damage and apoptosis via activation of caspase 3. EECGL significantly reduced MSG-induced inflammation mediated OS and apoptosis via inhibition of pro-inflammatory factors and pro-apoptotic mediators to protect liver and heart. Therefore, it can be suggested that EECGL contributed competent scientific information to validate the demands for its use to treat MSG-induced hepato-cardiac OS mediated inflammation and apoptosis from natural origin.
Introduction
Food habits, food preference and energy homeostasis of the body are directly linked with each other which modify the sensation of taste. 1 In recent years, there is a drastic change in food habits globally which positively correlates with different toxicological impacts on human health. In fast-paced life, peoples have very less time for the preparing their meal and eating with minimal physical activities apart from their work; most of them are dependent on ‘readymade fast foods’ either cafeteria or local restaurant with minimal or without essential nutrients that the our body needs to stay healthy which ultimately change their food habits and dependency 2 ; and such type of foods are considered as junk foods. 3 Moreover, for enhancement of taste and palatability of food, food-processors immensely used MSG as flavour enhancer secretly. Although, varieties of protein rich foods, soups, processed meats, dietary supplements, salad dressings, pizza, hamburgers, fried chicken, chips, hot dog, pakora, chow mein, French fries, cheese chili, pav bhaji, momo and some other snakes also contains MSG. 4 Earlier study demonstrated that MSG has been directly associated with diabetes, neurodegenerative disorders, cardiovascular damage, nephrotoxicity, and liver damage. 5 In the last few years, MSG has been excessively used large-scale because of its demands and dependency on readymade fast food, increased urbanization, and improvement of living standards and industrial development of processed foods all over the world, mostly in Asian countries. 6 In addition, MSG is being used randomly by crossing its safe limit of use and acts as an inducer of OS via generation of reactive oxygen species (ROS) by altering the redox equilibrium, which is an independent factor for systemic damage by increasing lipid peroxidation with disturbing the balance between FR and endogenous antioxidant (EA) levels. 7 Alteration of redox-homeostasis may also take part in initiation of inflammatory response to promote systemic damage.2,7 However, lack of scientific evidence about possible molecular target mechanism of action by which MSG exerted deleterious impacts on liver and heart by causing OS mediated anomalies and till now there was not such effective viable strategy of management to fight against MSG-induced anomalous situations. Moreover, failure of effective treatment with synthetic drugs may be due to their different adverse effects and high costing value; therefore, the present study mainly focused on plant based supplement as an alternative of synthetic drug which have been used as folk medicine to treat various diseases with minimal adverse effects, easily available and cheap in cost since ancient times. 8 Coccinia grandis (L) voigt commonly known as Ivy gourd is one of the widely used medicinal plants belonging to cucurbitaceae family; it has been found all over the world but mostly Asian countries with its different beneficial effects on health. 9 Every part of the plant have ethnopharmacological relevance to treat various diseases including allergy, antimicrobial, hepatocellular disorders, pancreatic diseases, respiratory diseases, and kidney related disorders, asthma, ulcer, urinary tract diseases, allergy, and bronchitis. 10 In addition, our recent study proposed that Coccinia grandis have significant in vivo anti-oxidative, anti-dyslipidemic, anti-inflammatory, and anti-apoptotic activity due to the presence of different ethnopharmacologically relevant bioactive compounds. 7 Natural supplements are very suitable for prophylactic use in order to prevent harmful impacts to maintain our normal wellbeing. Hence, we got an interest to fulfill the paucity of studies by means of preliminary evaluation of in-vitro antioxidant and FR-scavenging activity of six different solvent extracts of the leaves of Coccinia grandis to identify most valuable solvent extract with high antioxidant content as well as higher FR-scavenging property; thereafter, we further checked whether it has any role against MSG-induced liver and heart damage in male Wistar rat or not. It is perhaps the first comprehensive study with a mechanistic approach which provides a strong unique strategy of management against MSG-induced hepato-cardiac anomalies with Coccinia grandis.
Materials and methods
Reagents and chemicals
The accompanying reagents were acquired from Sigma–Aldrich Chemicals Company (St. Louis, MO): Folin-Ciocalteu reagent (FCR), deoxyribose, Griess reagent, polyvinyl polypyrrolidone, vanillin, potassium persulphate (K2S2O8), trichloroacetic acid (TCA), ferric chloride (FeCl3), phosphate buffer, 2, 2-diphenyl-1-picrylhydrazyl (DPPH), 2-thiobarbituric acid (TBA), 2,2’-azino-bis(3-ethyl benzothiazoline-6-sulfonic acid) (ABTS), ascorbic acid, quercetin, caffeine, gallic acid, α-tocopherol, butylated hydroxytoluene (BHT), diosgenin, cholesterol, sodium meta periodate, 3-methyl-2-benzothiazolinone hydrazone hydrochloride (MBTH), sodium nitroprusside, ethylenediaminetetraacetic acid (EDTA), hydrogen peroxide (H2O2), 4-nitro blue tetrazolium chloride (NBT), Iron (III) chloride (FeCl3), phenazine methosulfate (PMS), β-nicotinamide adenine dinucleotide (NADH), sodium carbonate (Na2CO3), aluminium chloride (AlCl3), sodium nitrite (NaNO2), sodium hydroxide (NaOH), 2-Deoxy-D-ribose, sulfuric acid (H2SO4), sodium phosphate, ammonium molybdate. Petroleum ether, n-hexane, chloroform, methanol and ethanol were procured from Merck (Darmstadt, Germany) and water was used from Millipore Milli-Q system (Bedford, MA). MSG (L-Glutamic Acid Monosodium Salt extrapure, SRL chemicals), paraformaldehyde (Sigma, Munich, Germany), 5, 59-dithio-bis (2-nitro benzoic acid) (DTNB), Triton X-100, Tween 20 were purchased from Sigma Aldrich (St. Louis, MO, USA). Antibody NF-kB (p65), cleaved caspase 3, 4′,6-diamidino-2-phenylindole (DAPI) were purchased from Cell Signalling Technology (Danvers, MA, USA). 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), Bovine serum albumin (BSA) were purchased from Sigma Chemical Co. (St. Louis, MO, USA). Anti-rabbit IgG fluorescein isothiocyanate (FITC) was obtained from Santa Cruz (Santa Cruz, CA, USA). 2′,7′-dichlorofluorescein diacetate (DCFH-DA) was purchased from Thermo Fisher Scientific, Waltham, MA, USA. All the cell culture media, buffer, reagents, collagenase type II were purchased from Gibco (Waltham, MA, US) and all other reagents used for this study were of highest quality grade.
Collection, identification and authentication of the plant
Fresh matured leaves of Coccinia grandis were collected from Serampore College Campus and certified (Specimen No. SC/HPY/BM/AB/001) by the Botanical Survey of India, Central National Herbarium, Howrah, 711 103, India. Primarily, the leaves were washed with water and shed dried for 360 hours, after that made it into fine powder form and sieved through the 40 micron strainer. Finally, the powder form of the leaves of Coccinia grandis put away from water and kept in an air-tight container (stored at 4°C until further use).
Preparation of extraction of the leaves of Coccinia grandis
Extraction was carried out at room temperature under normal conditions. 10,000 mg of shade dried powder of leaves of Coccinia grandis was extracted in 0.5 litre of different solvent separately such as aqueous (AECGL: aqueous extract of Coccinia grandis leaves), petroleum ether (PEECGL: petroleum ether extract of Coccinia grandis leaves), n-hexane (nHECGL: n-hexane extract of Coccinia grandis leaves), chloroform (CECGL: chloroform extract of Coccinia grandis leaves), methanol (MECGL: methanol extract of Coccinia grandis leaves) and EECGL for one week in the normal laboratory temperature, and the extract was centrifuged for 15 minutes at 4000 rotation per minute (rpm). Supernatant was taken as extracts of Coccinia grandis leaves, concentrated using a rotary evaporator at 40°C and dried in lyophilizer. Extracts were kept at −20°C for further research work. 7
Phytochemical screening of six different solvent extracts of the leaves of Coccinia grandis
Six different solvent extracts of the leaves of Coccinia grandis were utilized for fundamental screening of the presence of phytoconstituents according to standard techniques.11,12
Total antioxidant activity
To determine total antioxidant activity of each of the six different solvent extracts of the leaves of Coccinia grandis, in each hard glass test tube 300 µl of test sample was taken and 3000 µl of a reagent mixture of 0.6 M sulfuric acid, 28 mM sodium phosphate and 4 mM ammonium molybdate was added to it; kept it for 90 minutes at 95°C. Absorbance was estimated at 695 nm. Total antioxidant activity was expressed in mg/g of dry extract with ascorbic acid as standard. 13
Determination of total phenolic content
The total phenolic content in each of the six different solvent extracts of the leaves of Coccinia grandis were determined by using the FCR. 1000 µl of test sample was taken in each test tubes and 5000 µl of 1:10 diluted FCR was added to it; 4000 µl of 0.7 M Na2CO3 solution was added to the same test tube and absorbance was recorded at 760 nm. The total phenolic content was expressed in mg/g of dry extract with gallic acid as standard. 13
Determination of total flavonoid content
To determine flavonoid content in each of the six different solvent extracts of the leaves of Coccinia grandis, 100 µl of the sample extract was taken in each test tube and 300 µl of distilled water was added to it; in the same test tube 5% solution of 30 µl of NaNO2 was added and incubated at 25°C for 5 minutes. After the incubation, 10% solution of 30 µl of AlCl3 was added and the mixture of the test tube was treated with 200 µl of 1 mM NaOH solution followed by dilution with 1000 µl of distilled water and the absorbance was measured at 510 nm. The total flavonoid content was expressed in mg/g of dry extract with quercetin as standard. 13
Determination of total alkaloid content
Alkaloid content in each of the six different solvent extracts of the leaves of Coccinia grandis were estimated by taking 1500 µl of the test sample extract into each hard glass test tube and made up to 10000 µl with distilled water followed by the application of 1000 µl of 0.01 M sodium meta periodate, 500 µl of 0.1 M acetic acid to it; kept the hard glass test tube in 100oC water bath for 10 minutes. After that 2000 µl of 0.01 M MBTH was added in each of the test tubes and again placed it into the same water bath for 2 minutes. Cooled the test tube and made up to 25000 µl with double distilled water. Finally, the blue colour complex was measured at 630 nm in spectrophotometer and expressed in mg/g of dry extract by using caffeine as standard. 14
Determination of total sterol content
The spectrophotometric estimation of sterol was performed by taking 1000 µl of each of the six different solvent extracts in each of the test tube and 5000 µl of chloroform was added to it. About 2000 µl of Liebermann-Burchard reagent was added to each test tube, mixed well and kept at dark for 15 minutes; finally, the green colour complex was estimated by using spectrophotometer at absorbance of 640 nm. The total sterol content was expressed in mg/g of dry extract and cholesterol was used as standard. 15
Determination of total tannins content
To determine tannins content in each of the six different solvent extracts of the leaves of Coccinia grandis, in each test tube 500 µl of test sample, 10 mg of polyvinyl polypyrrolidone and 500 µl of distilled water was taken; the entire content of each of the test tube was incubated at 4oC for 4 hours. After that, test tubes were centrifuged at 5000 rpm for 5 minutes and from each test tube 20 µl of supernatant was taken. The supernatant contained phenolic free tannins and then finally the content of the supernatant was measured spectrophotometrically at 725 nm. The total tannins content was expressed in mg/g of dry extract with gallic acid as standard. 16
Determination of total saponin content
Saponin content in each of the six different solvent extracts of the leaves of Coccinia grandis were determined by taking 50 µl of each of the solvent extract in 250 µl of distilled water; 250 µl of vanillin reagent and 2500 µl of 72% of H2SO4 was added to each of the test tube. The entire content was kept at 60oC for 10 minutes. After that, it was cooled under tap water and absorbance was recorded at 544 nm. The total saponin content was expressed in mg/g of dry extract with diosgenin as standard. 16
DPPH radical scavenging activity
50 µl of 0.16 mM DPPH solution and 50 µl of each of the six different solvent extracts of the leaves of Coccinia grandis or standard ascorbic acid were mixed in a test tube. The content was then incubated for 30 minutes at dark and estimated at 517 nm; 50% inhibitory concentration (IC50) was expressed as mg/ml of extract. 17
Determination of scavenging activity of ABTS•+ radical cation
The ABTS•+ radical cation scavenging activity of each of the six different solvent extracts of the leaves of Coccinia grandis were estimated. In a test tube 7 mM ABTS with 2.45 mM K2S2O8 and kept it in a dark room for 16 hours to produce ABTS radical cation. 300 µl of ABTS solution was taken in six separate test tube, 50–150 µg of test sample extract was added to each of the test tube and final volume was made up to 1000 µl by using ethanol; absorbance was noted at 734 nm by using spectrophotometer where ascorbic acid used as standard antioxidant. IC50 was expressed as mg/ml of extract. 18
Hydroxyl radical (HO•) scavenging
HO• scavenging activities of each of the six different solvent extracts of the leaves of Coccinia grandis were estimated. In each test tube 100 µl of 1 mM EDTA, 10 µl of 10 mM FeCl3, 100 µl of 10 mM H2O2, 360 µl of 10 mM deoxyribose and 1000 µl of test sample extract or standard ascorbic acid (1-10μg/ml) were taken; to it 330 µl of 50 mM phosphate buffer (pH 7.4), 100 µl of 1 mM ascorbic acid were added and kept the entire content at 37°C incubator for 1 hour. 1000 µl from each of the test tubes was taken in another fresh test tube and mixed it with 1000 µl of 10% TCA, 1000 µl of 0.5% TBA solution to get a pink chromogen. Absorbance was estimated at 532 nm. IC50 was expressed as mg/ml of extract with standard ascorbic acid. 13
H2O2 scavenging assay
In each test tube 400 µl of test sample extract was taken and to it 600 µl of 40 mM H2O2 was added. Finally, the volume of each test tube was made up to 2000 µl by using a 50 mM phosphate buffer (pH 7.4) and kept it for 40 minutes at a 30°C incubator. The percentage scavenging activities of each of the six different solvent extracts of the leaves of Coccinia grandis were determined at 230 nm and IC50 values were compared α-tocopherol as standard in mg/ml of extract. 19
Nitric oxide (NO•) radical scavenging
Griess reagent was used to measure the generation of nitric oxide. 4000 µl test sample extract with 1000 µl of 5 mM sodium nitroprusside solution was taken to each of the test tubes and kept the entire content 27°C for 2 hours. After that, 2000 µl from each of the test tube was taken in another fresh test tube and 1200 µl of Griess reagent was added to it; mixed well and finally the absorbance was noted at 550 nm. The values were compared with standard ascorbic acid and IC50 values were expressed in mg/ml as compared to ascorbic acid. 13
Reducing power ability
To measure reducing power ability of each of the six different solvent extracts of the leaves of Coccinia grandis, in each test tube 1000 µl of test sample extract, 2500 µl of phosphate buffer (pH 6.6) and 2500 µl of 1% potassium ferricyanide were taken and kept the entire content at 50°C for at least half an hour. Then, 2500 µl of 10% TCA was added to each of the test tubes and centrifuged at 3000 g for at least 10 minutes. After centrifugation, 2500 µl of the supernatant was collected and diluted it with 2500 µl of distilled water; mixed well with 500 µl of 0.1% FeCl3 solution and finally absorbance was measured at 700 nm. IC50 values were expressed in mg/ml of extract with BHT as a standard. 20
Superoxide anion radical scavenging and superoxide dismutase (SOD) activity
Each of six different solvent extracts of the leaves of Coccinia grandis was added, one by one to test tube with 1000 µl of 156 μM NBT solution in phosphate buffer with pH 7.4, 1000 µl of 468 μM NADH in phosphate buffer with pH 7.4, and finally 100 µl PMS solution (60 μM prepared in phosphate buffer, pH 7.4) was added to it. After 5 minutes, absorbance at 560 nm was estimated and results were compared with standard ascorbic acid (10–50 μg/ml). SOD activity was expressed as nKat/ min/ mg of protein. 21
Catalase (CAT) activity
CAT activity of each of the six different solvent extracts of the leaves of Coccinia grandis were studied by measuring the absorbance at 240 nm. 500 µl 10 mM hydrogen peroxide was added in test tube with 500 µl of the test sample and 2000 µl of 0.1 M sodium phosphate buffer with pH 6.8. CAT activity was expressed as nKat/min/mg of protein. 22
Method of preparation of MSG
1.2 g of MSG was dissolved in 20 ml of distilled water in every day just before the treatment period.
In vivo hepatoprotective activity of EECGL
32 male adults (120–125 g) albino rats (Wistar strain) were chosen randomly and divided into four equal groups (n=8) and treated as: NC, normal control; MSG, treated with MSG in 0.6 mg/g body weight/day, orally 7 (which equates to 3.6g/60 kg of humans respectively); EECGL, treated with EECGL in 0.6 mg/g body weight/day, orally 7 (which equates to 3.6g/60 kg of humans respectively); MSG + EECGL, treated with MSG (0.6 mg/g body weight) + EECGL (0.6 mg/g body weight) orally for 28 days. The experimental conditions and experimental procedure were performed according to the ethical guidelines suggested by Serampore College Institutional Animal Ethics Committee, West Bengal, India (Project Approval No. 07/P/S/IAEC/2017, 24/P/S/SC/IAEC/2019) registered under Committee for the Purpose of Control And Supervision of Experiments on Animals (CPCSEA), Government of India (Reg. No. 1946/PO/Re/S/17/CPCSEA). Animals were maintained in an environmentally controlled animal house (temperature 24 ± 3oC) and in a 12 hours light/dark schedule with free access to water supply. The rats of all groups were supplied with a control diet prepared with 71% carbohydrate, 18% protein, 7% fat, and 4% salt mixture and water ad libitum. 2
Blood collection and serum preparation
After completion of the experimental period (on the 29th day), overnight fasted animals of different groups were anesthetized with intraperitoneal injection (ip) of ketamine (87 mg/kg body weight) suggested by IAEC. Blood was drawn by cardiac puncture and serum was prepared for measuring different biochemical parameters. 2
Biochemical assay
After preparation of serum from blood, different biochemical parameters were assessed by ACCUREX Biomedical Pvt. Ltd., Mumbai. Parameters were aspartate transaminase (AST), alanine amino transaminase (ALT), alkaline phosphatase (ALP), total cholesterol (TC), triglycerides (TG), very low density lipoprotein (VLDL), low density lipoprotein (LDL), and high density lipoprotein (HDL). VLDL and LDL were measured by Friedewald’s formula: VLDL-C = Triglyceride/5 and LDL-C = Total cholesterol-(VLDL-C + HDL-C). 23 Atherogenic index (AI) was measured by the formula: TG/HDL-C. 24 AST to Platelet Ratio Index (APRI) was calculated as APRI = (AST in IU/L)/(AST upper limit of normal in IU/L)/(Platelets in 103/μl) based on the meta-analysis in hepatology by Lin et al. 25
Serum lactate dehydrogenase (LDH) was evaluated by means of industrially accessible standard test packs (Stanbio Laboratory, USA) utilizing UV double beam spectrophotometer (Shimadzu Model 1800) and creatine kinase-MB (CK-MB) was assessed utilizing an economically accessible enzyme-linked immunosorbent assay kit with a microplate reader (Thermo Fisher Scientific, Waltham, MA, USA) as indicated by the directions of the maker.
Determination of cytokine level
Serum VEGF (RayBiotech), IL-6, IL-10, IL-1β, TNF-α and transforming growth factor (TGF)-β (eBioscience) levels were estimated by rat enzyme-linked immunosorbent assay (ELISA) kit by using microtiter plate reader. The assays were performed as per the detailed instructions of the manufacturer.
Preparation of tissue extract and isolation of hepatocytes
Liver and heart were isolated for preparation of tissue homogenate from all groups of animals for estimation of enzymatic and nonenzymatic antioxidant and OS parameters with protease inhibitor cocktail. 2 Further, isolation of hepatocytes and cardiomyocytes were achieved by two step collagenase type-II digestion perfusion technique.26,27
Estimation of lipid peroxidation and nitric oxide (NO)
The development of thiobarbituric acid reactive substance (TBARS) as a result of lipid peroxidation was estimated by TBA test 28 and the absorbance was estimated at 530 nm using ELISA Reader (Thermo Scientific). The outcomes were expressed as nmoles of TBARS per milligram of protein utilizing the molar extinction coefficient (1.56 × 105 cm2/mM). NO decomposes rapidly in aerated solutions to form stable nitrite/nitrate products. Griess reaction was utilized for the estimation of the nitrite accumulation by previously published method 29 and the absorbance was recorded at 550 nm using ELISA Reader (Thermo Scientific). NO was expressed in the sample as µmole/mg of protein.
Estimation of SOD
Nitroblue tetrazolium method was utilized for the estimation of SOD activity, which depends on the hindrance of NBT decreased by SOD. 29 The relative absorbance was measured in 560 nm and then converted into unit of SOD activity/mg of protein.
Estimation of CAT
CAT was estimated as per a well-established method of Bhattacharjee et al., 29 by the decomposition of H2O2 at 240 nm. The distinction in absorbance per unit time was utilized as a proportion of CAT activity and values were expressed as U/mg of protein.
Estimation of glutathione (GSH)
DTNB was utilized for estimation of GSH level. The absorbance of the decreased chromogen was pursued spectrophotometrically at 412 nm. GSH level was determined by utilizing a standard curve and expressed as nmoles/mg of protein. 30
Estimation of protein
BSA was used as standard for the estimation of protein content in liver and heart (both tissue homogenate and isolated cells). 31
Cell viability assay
MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide)] assay was used to decide the feasibility of cells as indicated by the instruction manual of MTT assay kit (Thermo Fisher Scientific, Waltham, MA, USA).
Histopathological analysis by hematoxylin and eosin stain
The liver and heart were fixed in 10% buffered formalin, processed, and embedded in paraffin for hematoxylin-eosin (H & E) staining and microscopic photographs were taken for analysis. The microscopic slides were analysed by using a compound microscope with camera attachment (Carl Zeiss, Primostar model) and photographs of the respective sections were taken. 2
Activity of NF-kB (p65) and caspase 3
NF-kB (p65) and caspase 3 activity were determined by standard kit (Abcam) as per the instruction manual. Optical density was measured at wavelength of 450 nm for NF-kB (p65) and 405 nm for caspase 3 activity by using ELISA plate reader.
Immunohistochemistry
Immunohistochemistry was carried out on paraffin sections with NF-kB (p65) and cleaved caspase 3 antibody. Briefly, Xylene was used to deparaffinize the sections and followed by permeabilization by treating with 0.1% Triton X100. Then unmasking of antigens was performed by heating the sections at 90oC for 10 minutes in a 10 mM citrate buffer, pH 6. After cooling at room temperature for 30 minutes each section was treated with the diluted primary antibodies overnight at 4oC. After that, the sections were then washed with PBS and incubated with appropriate dilution of secondary antibody tagged with FITC. Nuclei were stained by using DAPI. Fluorescent signals were viewed under a microscope (Olympus IX81). To observe any nuclear translocation of cleaved caspase 3, the colour of FITC was merged with the corresponding DAPI stained nuclei. Quantification of cleaved caspase 3 nuclear translocation was done by evaluating the colour intensity using Image J software. 32
Statistical analysis by Statsdirect 3.0 software
Data were expressed as Mean ± SE. Kruskal–Wallis nonparametric analysis of variance (ANOVA) test was performed to observe whether or not scores of various groups differ significantly. To test inter-group significance difference, Mann–Whitney U multiple comparison tests were performed to discover the correlation between the study variables. StatsDirect 3.0 (United Kingdom) was utilized for statistical analysis. Differences were considered significant if p < 0.05. 7
Results and discussion
Liver is the important organ in the body, which helps to detoxify different xenobiotic compounds and protects our body from different ailments; there is a link between the liver and heart via the circulatory system. In addition, recent report stated that MSG has also been linked with systemic damages via generation of ROS and thereby OS. 7 In pathophysiological condition, different antioxidants play an important function. However, increased lipid peroxidation and nitric oxide disturbed the normal homeostasis by altered the ratio of FR/EA which further leads to OS and thereby systemic damage because of lower level of intracellular antioxidants to combat against this anomalous situation. Likewise, in the present study, MSG also induced OS and disrupted redox-status to provoke hepato-cardiac anomalies. Therefore, attentiveness has been concentrated on antioxidants from natural sources with minimal or without any side effects to fight against the FR to protect liver and heart. Moreover, the in vitro experimental model was not enough to investigate and distinguish the antioxidant and FR scavenging properties of any plant extract; because such experimental model does not judge the appropriate parameters required for living organisms. Hence, in vivo experiments are more appropriate to evaluate the antioxidant and FR scavenging activity of any plant extracts to protect the living body from FR mediated damage. Therefore, the present study was demonstrated the hepato-protective and cardio-protective efficacy of EECGL in MSG induced anomalies in rat model by restoring normal lipid profile, reducing the level of cellular toxicity markers, suppressing inflammatory responses, regulating the balance between FR/EA and inhibiting programmed cell death. Due to major side effects of synthetic drugs, the present study was designed with plant-based supplementation as an alternative to synthetic drugs which further offer protective impacts against OS-induced cellular damage by MSG. In this context, the present study was highlighted on the antioxidant and FR-neutralizing activity of Coccinia grandis both in vitro and in vivo to reduce the chances of occurrence of OS, which regulates the redox-equilibrium and redox-guided cellular signalling to protect two different systems against MSG-induced anomalous situation.
Phytoconstituents of Coccinia grandis
Phytochemical screening is of paramount importance in identifying new sources of therapeutically valuable compounds having medicinal significance, to make the best and judicious use of available natural wealth. Yield of the six different solvent extracts of the leaves of Coccinia grandis were 15.44% (EECGL), 12.2% (MECGL), 10.33% (CECGL), 7.44% (PEECGL), 6.1% (nHECGL), and 8.5% (AECGL) respectively. Phytochemical screening of the six different solvent extracts of the leaves of Coccinia grandis revealed the presence of different phytochemicals (Figure 3A); EECGL showed maximum number of phytocompounds having different medicinal values. Furthermore, our recent high-performance thin-layer chromatography (HPTLC) and gas chromatography–mass spectrometry (GC-MS) study uncovered that ethanol extract of Coccinia grandis leaves have different bioactive phytocompounds with different therapeutic activities such as β-carotene, (E)-γ-atlantone, aR-turmerone, linolenic acid, β-turmerone, germacron, ergost-5-en-3β-ol, δ-tocopherol, farnesol and polyprenol have also been present in leaves of Coccinia grandis with different therapeutic activities. 7 Since, carotenoids, phytosterols, alkaloids and triterpenoids were present in Coccinia grandis in the present study thus it can be assumed that those active compounds may also be present in the Coccinia grandis or more prominently EECGL which was well corroborated with our earlier report. 7
Total antioxidant activity, total flavonoid, phenolic, sterol, alkaloid, tannins and saponin content of six different solvent extracts of the leaves of Coccinia grandis
Quantitatively higher amounts of secondary metabolites present in plants play a crucial role in protecting against OS mediated damage via antioxidant defence mechanism. 33 Therefore, quantitative estimation of antioxidant in six different solvent extracts of the leaves of Coccinia grandis may provide significant biological knowledge. In the present study, total antioxidant activity (Figure 1A) was significantly higher in EECGL (MECGL, CECGL, AECGL: p < 0.01; PEECGL, nHECGL: p < 0.001) as compared with the rest of the solvent extracts. Although, each of the solvent extract of Coccinia grandis leaves have significant antioxidant activity but the activity was not as much as the EECGL. Additionally, estimation of phytochemicals quantitatively specified the amount of the substance present in extracts; upon quantitative estimation of phytochemicals, we have found that each of the solvent extract of the leaves of Coccinia grandis was a good source of phenolic (Figure 1B), flavonoid (Figure 1C), sterol (Figure 1D), alkaloid (Figure 1E), tannins (Figure 1F) and saponin (Figure 1G) compounds; but the content was not as much as compared to EECGL. Sequential order of quantitative profile of phytochemicals present in six different solvent extracts of the leaves of Coccinia grandis has been shown in Figure 3B.

Quantitative profiles of six different solvent extracts of the leaves of Coccinia grandis. In vitro (A) total antioxidant activity, (B) total phenolic content, (C) total flavonoid content, (D) total sterol, (E) total alkaloid, (F) total tannins, (G) total saponin content of six different solvent extracts of the leaves of Coccinia grandis. Values were expressed as Mean ± SE; significance based on Kruskal Wallis test (##p < 0.01, ###p < 0.001). Significance level based on Mann-Whitney U multiple comparison tests: a-EECGL vs. MECGL, b-EECGL vs. PEECGL, c-EECGL vs. nHECGL, d-EECGL vs. CECGL, e-EECGL vs. AECGL, f-MECGL vs. PEECGL, g-MECGL vs. nHECGL, h-MECGL vs. CECGL, i-MECGL vs. AECGL, j-PEECGL vs. nHECGL, k-PEECGL vs. CECGL, l-PEECGL vs. AECGL, m-nHECGL vs. CECGL, n-nHECGL vs. AECGL, o-CECGL vs. AECGL [*p < 0.05, **p < 0.01, ***p < 0.001].
However, considering the qualitative and quantitative profile it can be suggested that higher antioxidant activity of EECGL due to high content of phytosterol, alkaloid, flavonoids, phenolic, tannins and saponin in EECGL which was well corroborated with earlier reports.34,35 Therefore, it can be speculated that the solvent of ethanol is able to dissolve more amounts of these compounds than other solvents. Moreover, it is debatable whether the total carotenoids, triterpenoid, phytosterol, alkaloid, phenolic, flavonoids, tannins, saponin along with some other compound acts as FR scavenger via antioxidant activity. This can be cleared after investigating the FR neutralizing activity of the individual solvent extract in the later part of this study.
Reducing power ability of six different solvent extracts of the leaves of Coccinia grandis as FR scavenger
Reducing power ability of the extract is directly correlated with antioxidant activity due to the presence of phenolic compounds. 33 In the present investigation, antioxidants present in each of the extract was able to reduce more potassium ferricyanide to potassium ferrocyanide and ultimately formed Fe2+ complex by showing higher reducing power ability. Furthermore, it has been observed that EECGL showed significantly (p < 0.05) higher reducing power ability as compared to the standard BHT; although the rest of the solvent extracts showed significantly (p < 0.01) higher reducing power ability (Figure 2A) as compared to the standard but the level was not as much as EECGL. Higher reducing power ability of extract served as a major indicator of antioxidant activity. Therefore, to impair the initiation of chain, peroxide decomposition, reducing ability and neutralizing the FR, antioxidants play a beneficial role. 36

In vitro FR scavenging activity of six different solvent extracts of the leaves of Coccinia grandis. (A) Reducing power, (B) DPPH scavenging ability, (C) ABTS•+ scavenging ability, (D) superoxide anion radical scavenging activity, (E) hydrogen peroxide scavenging activity, (F) nitric oxide radical scavenging activity, (G) hydroxyl radical scavenging activity, (H) SOD activity, (I) CAT activity of six different solvent extracts of the leaves of Coccinia grandis. Values were expressed as Mean ± SE; significance based on Kruskal Wallis test (##p < 0.01). Significance level based on Mann-Whitney U multiple comparison tests: a-EECGL vs. MECGL, b-EECGL vs. PEECGL, c-EECGL vs. nHECGL, d-EECGL vs. CECGL, e-EECGL vs. AECGL, f-MECGL vs. PEECGL, g-MECGL vs. nHECGL, h-MECGL vs. CECGL, i-MECGL vs. AECGL, j-PEECGL vs. nHECGL, k-PEECGL vs. CECGL, l-PEECGL vs. AECGL, m-nHECGL vs. CECGL, n-nHECGL vs. AECGL, o-CECGL vs. AECGL, p-EECGL vs. BHT/Ascorbic acid/α-tocopherol, q-MECGL vs. BHT/Ascorbic acid/α-tocopherol, r-PEECGL vs. BHT/Ascorbic acid/α-tocopherol, s-nHECGL vs. BHT/Ascorbic acid/α-tocopherol, t-CECGL vs. BHT/Ascorbic acid/α-tocopherol, u-AECGL vs. BHT/Ascorbic acid/α-tocopherol [*p < 0.05, **p < 0.01,***p < 0.001].
DPPH radical scavenging activity of six different solvent extracts of the leaves of Coccinia grandis as FR scavenger
To determine the anti-oxidative activity of the antioxidants present in the different solvent of the leaves of Coccinia grandis we have used DPPH as a substrate. 33 In the present experimental set up, it was revealed that in presence of hydrogen donating antioxidant, the purple colour of DPPH solution was reduced (Figure 2B). Moreover, the reduction of purple colour indicates the FR scavenging activity of the extracts. Whereas the ascorbic acid as standard showed DPPH radical scavenging activity was very close to EECGL and but the difference between these two groups were statistically significant (p < 0.05). On the other hand, the other five solvent extracts of the leaves of Coccinia grandis showed significant (p < 0.01) differences in DPPH radical scavenging activity as compared to the standard ascorbic acid but not up to the level of EECGL. Therefore, it can be suggested that EECGL has great source of hydrogen-donor capacity apart from the rest of the solvent extracts; which acts as antioxidant with higher FR scavenging activity whereas the other five solvent extracts with lower FR activity mainly due to reduced levels of hydrogen donating capacity or presence of higher amounts of pro-oxidants.
ABTS•+ scavenging activity of six different solvent extracts of the leaves of Coccinia grandis as FR scavenger
The total antioxidant activity of six different solvent extracts of the leaves of Coccinia grandis were performed by estimating the decolourization of ABTS•+. ABTS•+ radical scavenging activity was depended on presence of the flavonoid and phenolic constituents 33 ; the present study already showed that each of the six different solvent extracts of the leaves of Coccinia grandis contains flavonoid and phenolic compounds. However, the present experimental set up showed higher ABTS•+ radical scavenging activity (Figure 2C) due to presence of higher amount of phenolic and flavonoid content in the six different solvent extracts by impaired the formation of ABTS•+. Furthermore, EECGL showed significantly (p < 0.05) higher ABTS•+ radical scavenging activity as compared to the standard ascorbic acid, whereas rest of the solvent extracts did not show the same activity as exhibited by EECGL.
Superoxide anion radical scavenging activity of six different solvent extracts of the leaves of Coccinia grandis as FR scavenger
Superoxide anion is a toxic substance in the biological system because it can be decomposed to form as singlet oxygen and hydroxyl radicals. 37 The superoxide anion scavenging activity (Figure 2D) occurred by inhibition of the oxidation of NBT to form formazan. In the present study, each of the extracts showed superoxide anion radical scavenging activity with ascorbic acid as reference antioxidant; but the superoxide anion scavenging activity was higher in EECGL as compared with the rest of the extracts. Therefore, it can be inferred that high solubility of the antioxidant compounds in EECGL inhibits the formazan formation and thereby exhibits higher FR-neutralizing activity.
H2O2 scavenging activity of six different solvent extracts of the leaves of Coccinia grandis as FR scavenger
H2O2 scavenging activity of the plant extract was attributed to its phenolic content which can contribute electrons to the H2O2 and thereby converted it to water. 38 The capacity of the extracts to effectively neutralize the H2O2 to water was compared with α-tocopherol as reference antioxidant and the present investigation showed EECGL has high H2O2 scavenging activity (Figure 2E). However, the H2O2 scavenging activity of EECGL was very close to the reference antioxidant. However, H2O2 is not always reactive with very low concentration but if a high amount of H2O2 is generated then it can cause cellular toxicity by producing intracellular hydroxyl radicals. Hence, neutralizing of H2O2 is very important for maintaining the OS free environment in biological system. 33 It can be speculated that higher H2O2 scavenging activity of EECGL may be due to more solubility of the phytocompounds in it.
Nitric oxide radical (NO•) scavenging activity of six different solvent extracts of the leaves of Coccinia grandis as FR scavenger
NO• scavenging activity was measured by the amount of NO produced from sodium nitroprusside reacts with oxygen to generate stable nitrite and nitrate ions. The present study showed that presence of FR scavenger in six different solvent extracts of the leaves of Coccinia grandis reacts with oxygen and thereby attenuated the formation of nitrite. Furthermore, the nitrite ion reacts with sulphanilamide of Griess reagent to form a diazotized molecule which was estimated in spectrophotometry. 33 The ethanol extract of Coccinia grandis leaves demonstrated strong NO• scavenging action (Figure 2F), although rest of the extracts also have good NO• scavenging properties but EECGL showed stronger NO• scavenging action than the rest of the solvent extracts. However, the present experiment also reported that EECGL showed significantly (p < 0.05) higher nitric oxide radical scavenging activity as compared to the reference antioxidant ascorbic acid. Considering the biochemical data, it can be assumed that decreased nitrite concentration in presence of EECGL is attributed to prevent formation of nitrite from NO•. Therefore, it can be hypothesized that each of the extracts with its endogenous antioxidants compete with the oxygen to react with NO and inhibit the generation of stable nitrite/nitrate by reduced the formation of diazotized molecule but higher nitric oxide scavenging activity in EECGL.
HO• scavenging activity of six different solvent extracts of the leaves of Coccinia grandis as FR scavenger
Fe2+ as catalytic component mediated degradation of deoxyribose by Fenton reaction produced HO•. To prevent the degradation of deoxyribose, it should be necessary to contain FR-scavenger in the extracts to neutralize its reactivity. 33 To investigate the HO• scavenging activity (Figure 2G) of Coccinia grandis by measuring the inhibition of the degradation of deoxyribose by the FRs produced through the Fenton reaction, we have found that each of the solvent extract of the leaves of Coccinia grandis prevent the generation of HO• with ascorbic acid as positive control. Moreover, all the six solvent extracts possessed good HO• scavenging activity but the scavenging activity was higher in EECGL. Furthermore, EECGL showed significantly (p < 0.05) higher HO• scavenging activity as compared to the reference antioxidant ascorbic acid. Therefore, it can be inferred that higher ability of EECGL to scavenge HO• and other FRs have been directly proportional to its antioxidant property due to the presence of different phytoconstituents and their solubility in the ethanol extract as evidenced by both qualitative and quantitative phytochemical profiles.
SOD and CAT activity of six different solvent extracts of the leaves of Coccinia grandis
Human and animal cells ensure themselves against the FR damage by endogenous antioxidant enzymes such as SOD and CAT. 39 The superoxide anion scavenging activity of the six different solvent extracts was additionally supported by SOD activity. CAT catalyses the deterioration of H2O2 to water and oxygen by acting as a first line safe guard antioxidant enzyme with SOD. 40 In the present study, EECGL showed higher SOD (Figure 2H) and CAT (Figure 2I) activity which further offer to protect human cells from FR-mediated damage. The FR scavenging activity and antioxidant potential of six different solvent extracts of the leaves of Coccinia grandis were also well supported by the presence of SOD and CAT activity. However, six different solvent extracts of the leaves of Coccinia grandis exhibited a good source of SOD and CAT activity but the endogenous enzyme activity was higher in EECGL.
Selection of extract on the basis of antioxidant and FR-scavenging properties for the in vivo experiment
Sequential order of in vitro antioxidant and FR scavenging activity of six different solvent extracts of the leaves of Coccinia grandis has also been shown in Figure 3C and 3D. Considering both qualitative and quantitative phytochemical data, total antioxidant and FR-neutralizing activity of Coccinia grandis, it revealed that each of the solvent extracts have a great source of antioxidant and able to scavenged the FR but it was not as much as EECGL exhibited in the present study. Moreover, it can be postulated that high antioxidant and FR-neutralizing activity of EECGL due to higher solubility of phytoconstituents (as evidenced by earlier published HPTLC and GCMS analysis of EECGL) 7 with different health-promoting activities. Hence, the synergistic impacts of phytoconstituents present in the EECGL acts as an antioxidant with diverse health promoting impacts may be responsible for its viable restorative impacts on MSG induced anomaly by inhibiting OS mediated inflammation and apoptosis.

Phytochemical profile, sequential order of quantitative profile of phytochemicals, in vitro antioxidant and FR scavenging activity of six different solvent extracts of the leaves of Coccinia grandis. (A) Phytochemical profile of six different solvent extracts of the leaves of Coccinia grandis (‘+’ indicated as presence and ‘−’ as absence). (B) Sequence of order of quantitative profile of phytochemicals present in six different solvent extracts of the leaves of Coccinia grandis. (C) Sequence of order of in vitro FR scavenging activity of six different solvent extracts of the leaves of Coccinia grandis. (D) Sequence of order of in vitro enzyme activity of six different solvent extracts of the leaves of Coccinia grandis.
EECGL ameliorates MSG-induced dyslipidemia and cell damage by regulation the balance of LDL/HDL level and cellular toxicity markers
In the present study, MSG-induced hyperlipidaemia was established by significant increase (p < 0.01) levels of TC, TG, VLDL and LDL by 56.90%, 45.47%, 49.98% and 62.88% respectively (Figure 4A) and decrease level of (p < 0.01) HDL by 68.92% in rat model as compared to the control (NC) group. Additionally, MSG-induced alteration of the ratio of LDL/HDL leads to dyslipidemia which was well supported by 5.91 folds higher AI (Figure 4B) (p < 0.01) in MSG group as compared to NC group. Disturbances in LDL and HDL level in MSG group confirmed hyperlipidemia mediated lipid metabolic aggravations, finally increased lipogenesis and decreased lipolysis which also corroborated with earlier study. 41 Moreover, MSG + EECGL group significantly (p < 0.01) increase HDL level by 62.81% and decrease TC, TG, VLDL, LDL level by 40.86%, 36.21%, 31.12% and 53.14% respectively; on the other hand, high AI was significantly (p < 0.01) reduced by the EECGL supplementation by 4.23 folds as compared with that in the MSG-treated group. There was no such significant difference (NS) observed in the EECGL group as compared to the NC group. The antidyslipidemic activity of EECGL was well corroborated with earlier studies7,42; which offered to protect MSG-induced dyslipidemia via restored the LDL/HDL balance and reduction of hypercholesterolemia by lipogenesis may be via liver X receptor dependent signalling. 43 Although the present study did not observe any significant changes in daily food intake (data not shown) among different groups of rats.
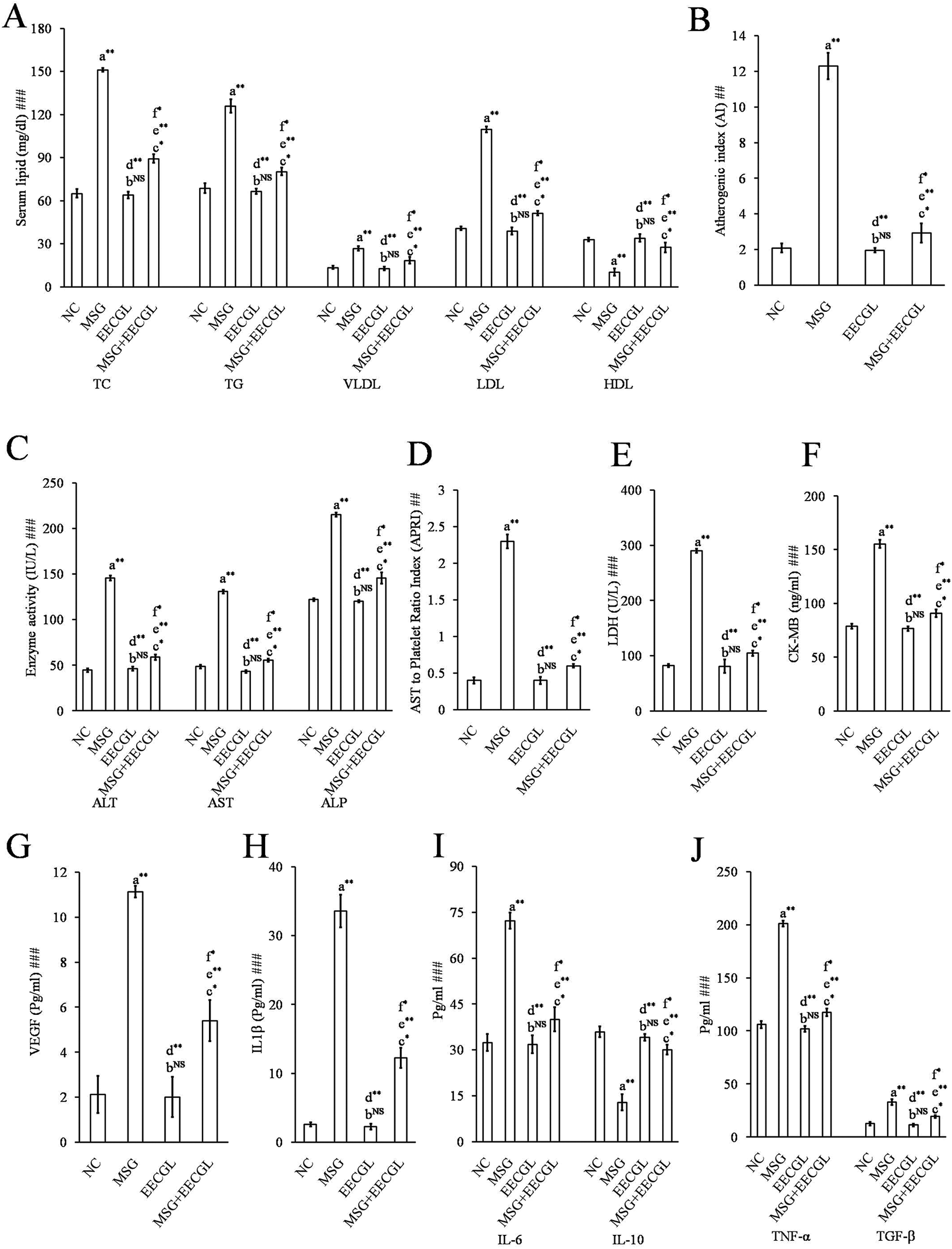
Effects of EECGL on MSG-induced dyslipidemia, hepato-cardiac toxicity, inflammation and fibrogenesis. (A) Lipid profile, (B) atherogenic index, (C) ALT, AST, ALP, (D) APRI, (E) LDH, (F) CK-MB, (G) VEGF, (H) IL-1β, (I) IL-6 and IL-10, (J) TNF-α and TGF-β. Values were expressed as Mean ± SE; significance based on Kruskal Wallis test (##p < 0.01, ###p < 0.001). Significance level based on Mann-Whitney U multiple comparison tests: a-NC vs. MSG, b-NC vs. EECGL, c-NC vs. MSG + EECGL, d-MSG vs. EECGL, e-MSG vs. MSG + EECGL, f-EECGL vs. MSG + EECGL [*p < 0.05, **p < 0.01, NS: not significant].
In addition we have also checked some preliminary cellular toxicity markers ALT, AST, ALP, LDH and CK-MB to evaluate the correlation between MSG-induced hepato-cardiac anomalies because increased levels of these enzymes indicates tissue damage. After completion of experiment ALT, AST and ALP levels (Figure 3C) were estimated and it was observed that the serum ALT, AST and ALP levels were significantly increased (p < 0.01) by 69.37%, 62.87% and 43.32% respectively in the MSG group as compared to the NC group. In addition, APRI (Figure 4D) was also significantly increased (p < 0.01) by 5.75 folds in MSG fed rats as compared to the control. According to Lin et al., 25 APRI is an important marker of fibrosis or cirrhosis, APRI ≥2 indicate 36% sensitive, 93% specific for severe fibrosis and 46% sensitive, 91% specific for cirrhosis which was observed in the MSG fed rats; on the other hand, APRI <0.7 were not sensitive or specific enough to determine level of fibrosis or cirrhosis which was observed in NC, EECGL and MSG + EECGL fed rats respectively. Therefore, it can be suggested that EECGL acts as an alternative to anti-fibrotic/anti-cirrhotic supplement against MSG induced liver damage which was well corroborated with TGF-β and VEGF level in the later part of the study.
Moreover, LDH (Figure 4E), CK-MB (Figure 4F) were significantly (p < 0.01) increased by 71.63% and 49.24% respectively, in the MSG group as compared to the NC group. Therefore altered hepato-cardiac toxicity markers by the treatment of MSG indicated systemic damage and similar types of results were also documented in earlier studies.7,42,44 However, MSG + EECGL group significantly (p < 0.01) reduce the augmented levels of ALT (59.71%), AST (57.58%), ALP (32.29%), LDH (63.91%), CK-MB (41.59%) and APRI (3.83 folds) towards normalcy as compared to the MSG group recommended as a therapeutic efficacy of EECGL in MSG-induced cellular toxicity. There was no significant difference of such parameters observed in the EECGL group as compared to the NC group. Therefore, it can be suggested that EECGL showed hepatoprotective activity by reducing the cellular toxicity markers to protect the liver from MSG-induced damage.
EECGL suppressed MSG-induced inflammation mediated systemic damage by inhibiting NF-kB and VEGF signalling
Pro-inflammatory cytokines play a pivotal role in the development of inflammation, steatosis, fibrosis of liver 45 which further stimulate cardiac damage 7 ; moreover in hepatic fibrosis also stimulated by the influence of TGF-β. 41 In the present study, IL-1β (Figure 4H), IL-6 (Figure 4I) TNF-α and TGF-β (Figure 4J) were significantly (p < 0.01) increased by 92.25%, 55.18%, 47.33% and 61.44% respectively, and IL-10 (Figure 4I) was significantly (p < 0.01) decreased by 64.05% in MSG fed rats as compared to the NC group. Pro-inflammatory mediators TNF-α, IL-1β and IL-6 were increased on binding of specific sequence of promoter region of the target gene with NF-kB. 7 As anti-inflammatory cytokines IL-10 was reduced it failed to inhibit the NF-kB and thereby the binding of pro-inflammatory factors to bind with the target gene. Therefore, it can be demonstrated that the disturbances in pro-inflammatory/anti-inflammatory factors induced inflammation primarily via the activation of NF-kB signalling pathway which was also evidenced by immunohistochemistry data in the later portion of the study. Moreover, up regulation of pro-fibrogenic TGF-β provoked acute necro-inflammation, steatosis and fibrosis. 46 It can be proposed that MSG-induced increase TGF-β promoted inflammation mediated fibrosis in the present study was well corroborated with earlier report. 47 On the contrary, MSG + EECGL group significantly (p < 0.01) blunted the serum IL-1β, IL-6, TNF-α and TGF-β by 63.49%, 44.62%, 41.59% and 41.15% respectively, and escalated the IL-10 level by 57.10% as compared to the MSG group (no significant difference in the EECGL group alone as compared to the NC group). The present study also demonstrated EECGL has noteworthy anti-inflammatory potential via regulating the pro-inflammatory and anti-inflammatory cytokine levels by partial inhibition of NF-kB signalling (as evidenced by immunohistochemistry data). Therefore, it can be suggested that EECGL inhibit the binding of pro-inflammatory cytokines to the target gene via increased anti-inflammatory IL-10 to blocks the NF-kB signalling.
Furthermore, we also measured most important angiogenic cytokine VEGF which is the key pro-angiogenic factor during chronic liver injury and may act as fibrogenic during the development of liver cirrhosis. In the present study, VEGF (Figure 4G) level was significantly (p < 0.01) increased by 80.94% in the MSG fed group as compared to the NC group. VEGF triggered the pro-angiogenic activity mainly by binding to two high-affinity tyrosine kinase receptors, VEGFR-1 and VEGFR-2.48–50 Therefore, it can be suggested that up regulation of VEGF indicated chronic liver injury in MSG fed rats. Our results therefore confirmed that the VEGF signalling pathway may trigger the microvascular proliferation associated with liver fibrogenesis, thereby contributing to the remodelling of liver architecture. It has been found that inhibition of either VEGFR-1 or VEGFR-2 significantly attenuated liver fibrogenesis accompanied by angiogenesis suppression. 51 Interestingly, treatment of EECGL significantly (p < 0.01) decreased the serum VEGF by 51.44% in MSG-fed rats (no significant difference in the EECGL group alone as compared to the NC group). These data first uncovered that the anti-fibrotic effect of EECGL could act via the inhibition of high-affinity tyrosine kinase receptors to further influence liver fibrosis associated angiogenesis. Additionally, inflammation also altered the redox-balance via activation of pro-inflammatory cytokines or vice versa and thereby systemic damage2,7 which was uncovered after investigating the OS related parameters in the later part of the study.
EECGL diminished MSG-induced OS by modifying redox status and increased survivability of cells
Accumulating evidence suggested that pro-inflammatory responses have been directly linked with ROS generation and thereby OS mediated systemic anomalies.2,7 Generation of ROS boosted the level of lipid peroxidation which disrupted the intracellular antioxidant/FR balance. 52 Figure 5A and 5B have been shown in vivo antioxidant and FR neutralizing activity of Coccinia grandis. The lipid peroxidation by generation of TBARS in liver tissue homogenate and isolated hepatocytes were significantly (p < 0.001) increased by 4.08 and 4.35 folds respectively, and in heart tissue homogenate and isolated cardiomyocytes were significantly (p < 0.001) increased by 5.69 folds and 6.25 folds respectively in the MSG group as compared to the NC group but no such significant difference was found in the EECGL group as compared to the NC group. Moreover, NO level in liver tissue homogenate and isolated hepatocytes were significantly (p < 0.001) increased by 4.17 and 4.45 folds respectively, and in heart tissue homogenate and isolated cardiomyocytes were significantly (p < 0.001) increased by 6.03 folds and 6.98 folds respectively in the MSG group as compared to the NC group. On the other hand, in both liver tissue homogenate and isolated hepatocytes, MSG + EECGL group significantly (p < 0.001) blunted the elevated levels of TBARS (liver tissue homogenate by 50.91%, isolated hepatocytes by 50.43%, heart tissue homogenate by 65.37% and isolated cardiomyocytes by 69.48%) and NO (liver tissue homogenate by 43.96%, isolated hepatocytes by 47.80%, heart tissue homogenate by 70.69% and isolated cardiomyocytes by 73.74%) as compared to the MSG group. Furthermore, we have also measured the EA levels, in which SOD activity was significantly (p < 0.001) decreased in MSG group by 1.74 folds both in liver homogenate and isolated hepatocytes, and 1.95 folds in heart tissue homogenate and 1.99 folds in isolated cardiomyocytes as compared to the NC group; but EECGL significantly (p < 0.01) increased these activity by 33.83% in both liver tissue homogenate and isolated hepatocytes, and 38.76% in heart tissue homogenate and 39.97% in isolated cardiomyocytes. In addition, another first line defence enzyme CAT was significantly (p < 0.01) decreased by 1.41 folds in liver tissue homogenate, 1.40 folds in isolated hepatocytes, 1.62 folds in heart tissue homogenate and 1.59 folds in isolated cardiomyocytes in MSG fed animals as compared to the NC group of animals. Furthermore, EECGL significantly (p < 0.01) increased these activities by 21.79% in liver tissue homogenate, 21.07% in isolated hepatocytes, 30.86% in heart tissue homogenate and 32.69% in isolated cardiomyocytes in the MSG + EECGL group as compared to the MSG group. Furthermore, GSH were significantly (p < 0.01) decreased by 1.59 folds in liver tissue homogenate, 1.54 folds in isolated hepatocytes, 1.66 folds in heart tissue homogenate and 1.67 folds in isolated cardiomyocytes in the MSG group as compared to the NC group; on the contrary EECGL significantly (p < 0.01) increased the GSH level by 34.56% in liver tissue homogenate, 18.17% in isolated hepatocytes, 32.78% in heart tissue homogenate and 30.79% in isolated cardiomyocytes as compared to the MSG group. However, all over the experiments there was no such significant difference observed in the EECGL alone group as compared to the NC group of animals.

Effect of EECGL on MSG induced OS-related parameters. (A) Bar diagram represents the TBARS, NO, SOD, CAT, GSH level in NC, MSG, EECGL and MSG + EECGL treated groups from liver tissue homogenate and isolated hepatocytes, respectively. (B) Bar diagram represents the TBARS, NO, SOD, CAT, GSH level in NC, MSG, EECGL and MSG + EECGL treated groups from heart tissue homogenate and isolated cardiomyocytes, respectively. Values were expressed as Mean ± SE; significance based on Kruskal Wallis test (##p < 0.01). Significance level based on Mann-Whitney U multiple comparison tests: a-NC vs. MSG, b-NC vs. EECGL, c-NC vs. MSG + EECGL, d-MSG vs. EECGL, e-MSG vs. MSG + EECGL, f-EECGL vs. MSG + EECGL [*p < 0.05, **p < 0.01, ***p < 0.001, NS: not significant].
In normal physiological state, there is equilibrium between the intracellular antioxidants and endobiotics or xenobiotics; however, MSG provoked generation of intracellular reactive free radicals and electrophiles which deranged the normal physiological state of the body and thereby leads to OS mediated cellular damage. In the present study, we demonstrated that EECGL reduced MSG induced increased lipid peroxidation and nitric oxide by restoring the levels of endogenous antioxidants and thereby attenuated OS mediated damage; which offers possible in vivo antioxidant and anti-oxidative potential of EECGL against MSG induced imbalance of FR and EA, either via acts as FR-scavenger or modulating redox-guided cellular signalling.
However, hepatocellular viability (Figure 6A) were significantly (p < 0.001) reduced by 57.35% in MSG group as compared to the NC group but EECGL significantly (p < 0.001) increased this level by 42% as compared to the MSG group (no significant difference was observed in only EECGL group as compared to the NC group). On the other hand, viability of cardiomyocytes (Figure 6B) was significantly (p < 0.001) reduced by 47.35% in MSG group as compared to the NC group but EECGL significantly (p < 0.001) increased this level by 26.25% as compared to the MSG group (no significant difference was observed in only EECGL group as compared to the NC group). The decreased viability of both hepatocytes and cardiomyocytes may be due to MSG-induced FR generation, which is further restored by EECGL supplementation due to its FR scavenging properties in vivo. Thus, it can be inferred that EECGL has a defensive role in preventing MSG-induced altered redox-status and thereby loss of viability of hepatocytes and cardiomyocytes.

Effects of EECGL on MSG induced changes in cell viability and histopathology of liver and heart. (A) Viability of hepatocytes in NC, MSG, EECGL and MSG + EECGL group, (B) viability of cardiomyocytes in NC, MSG, EECGL and MSG + EECGL group, (C) histopathology of liver section of NC, MSG, EECGL and MSG + EECGL group, (D) histopathology of heart section of NC, MSG, EECGL and MSG + EECGL group. Values were expressed as Mean ± SE; significance based on Kruskal Wallis test (###p < 0.001). Significance level based on Mann-Whitney U multiple comparison tests: a-NC vs. MSG, b-NC vs. EECGL, c-NC vs. MSG + EECGL, d-MSG vs. EECGL, e-MSG vs. MSG + EECGL, f-EECGL vs. MSG + EECGL [*p < 0.05, **p < 0.01, ***p < 0.001, NS: not significant].
EECGL rehabilitated MSG induced changes in tissue architecture
Histological findings of the present study showed (Figure 6C) MSG-induced alteration in hepatic tissue architecture by using H & E staining. Normal radiating pattern with intact central lobule was observed in the NC and EECGL group alone, no such other pathological disturbances were observed in this section; degeneration in the hepatocytes, atrophic changes, marked necrosis, ruptured central lobule, hepatocellular inflammation, haemorrhage, absence of hepatic sinusoid, enlarged hepatocytes, blurred boundaries between cells, hepatic steatosis and fibrosis were seen in MSG fed group which was well corroborated with the data of pro-inflammatory factors, more specifically TGF-β, VEGF, and APRI. Elevated levels of cell toxicity markers, hepatic fibrosis markers and OS stimulated the MSG-induced hepatic tissue damage. On the contrary, EECGL restored the normal architectural view of liver in the MSG fed animals which further suggested the potential hepato-protective role of EECGL.
Additionally, no pathological findings were observed in the cardiac tissue section of control as well as EECGL group (Figure 6D). However, severe necrotic changes with inflammatory response, occasional loss of cardiac muscle fibres were observed in MSG fed group which well corroborated leakage of LDH, inflammatory response and OS to promote tissue damage; but the supplementary group beneficially restored the normal architectural view of the heart in MSG fed group which further suggested the possible cardio-protective role of EECGL.
EECGL attenuated MSG-induced activation of NF-kB (p65) and caspase 3 via inhibiting NF-kB signalling and caspase 3 signalling
Different cellular response associated physiological disturbances are mediated by reduction/oxidation regulation and one of the important redox-sensitive factors is NF-kB play a crucial role in development of inflammatory responses via different cytokines and immune responsive genes. It is generally concealed in the cytosol of stimulated cells and in response to the OS it translocates into the nucleus. ROS triggered the activation of NF-kB signalling in cytosol and nuclear translocation. 53 NF-kB has also played an important role in inflammation mediated cellular damage and death. 54 To substantiate MSG induced inflammation and systemic injury NF-kB (p65) activity was estimated by spectrophotometry and nuclear translocation by immunofluorescence method. The present study showed liver tissue homogenate, isolated hepatocytes, heart tissue homogenate and isolated cardiomyocytes of MSG fed rats exhibited higher NF-kB (p65) (Figure 7A, 7B) activity. NF-kB (p65) activity was significantly (p < 0.01) increased in MSG group by 95.44% in liver tissue homogenate, 96.15% in isolated hepatocytes, 96.95% in heart tissue homogenate and 97.41% in isolated cardiomyocytes as compared to the NC group. This result demonstrated that NF-kB (p65) activation was involved in MSG-induced OS mediated cellular inflammation which further justified the finding of immunohistochemistry data (Figure 8A, 9A). MSG + EECGL intervention group significantly (p < 0.01) reduced increased NF-kB (p65) activity by 90.10% in liver tissue homogenate, 90.31% in isolated hepatocytes, 86.28% in heart tissue homogenate and 85.88% in isolated cardiomyocytes as compared with the MSG-treated group; only EECGL treatment did not show any significant effect.

Effect of EECGL on MSG induced activation of NF-kB (p65) and caspase 3. (A) NF-kB (p65) and caspase 3 activity of liver tissue homogenate and isolated hepatocytes, (B) NF-kB (p65) and caspase 3 activity of heart tissue homogenate and isolated cardiomyocytes. Values were expressed as Mean ± SE; significance based on Kruskal Wallis test (##p < 0.01). Significance level based on Mann-Whitney U multiple comparison tests: a-NC vs. MSG, b-NC vs. EECGL, c-NC vs. MSG + EECGL, d-MSG vs. EECGL, e-MSG vs. MSG + EECGL, f-EECGL vs. MSG + EECGL [*p < 0.05, **p < 0.01, ***p < 0.001, NS: not significant].
Furthermore, to understand the level of phosphorylation of NF-kB (p65) and translocated it into the nucleus upon MSG treatment and whether the EECGL potentially suppressed the phosphorylation and nuclear translocation of NF-kB (p65), immunohistochemistry was carried out. The present investigation showed MSG-induced nuclear translocation of NF-kB (p65); whereas NF-kB (p65) remained localized in the cytosol of hepatocytes and cardiomyocytes in NC (3.33% nuclei were NF-kB positive in case both liver and heart) and EECGL (3.27% nuclei were NF-kB positive in case of liver and 3.19% nuclei were NF-kB positive in case of heart) group. Moreover, in the MSG group showed maximum translocation of NF-kB with 43.33% nuclei NF-kB (p65) positive in case of liver and 18.18% nuclei were NF-kB positive in case of heart within the selected region of the section were observed. Accumulating evidence suggested that increased systemic injury was associated with augmented level pro-inflammatory IL-6 and TNF-α which directly linked OS by MSG.7,55 Moreover, IL-6 and TNF-α act as major contributors of hepatic and cardiac derangement through inflammatory pathways via this vicious cycle. 7 MSG increased pro-inflammatory cytokines which specifically bind with the promoter region of the NF-kB targeted gene to initiate inflammatory response. On the contrary, the translocation was decreased in MSG + EECGL group (11.66% nuclei were NF-kB positive within the selected region of liver section and 5.17% nuclei were NF-kB positive within the selected region of heart section) (Figure 8, 9). Therefore, EECGL supplementation in MSG fed group decreased nuclear translocation of NF-kB (p65) expressions. Therefore, it was observed that inhibition of NF-kB by EECGL occurred due to reduced production IL-6 and TNF-α. Hence, it indicates EECGL play a crucial role to prevent cellular inflammation by inhibiting the binding of pro-inflammatory cytokines to bind on the targeted region of NF-kB and thereby inflammatory signals.
In addition, redox-state and intracellular GSH help to regulates the activity of pro-apoptotic caspase 3 activity via S-glutathiolation but altered redox-status and reduced level of GSH may be a positive contributor of apoptosis via caspase activation. 56 In this context, to measure whether MSG has any effect on apoptosis via the activation of caspases due to alteration of redox-balance and decreased endogenous GSH, a specific peptide substrate was used to detect caspase 3 proteolytic activity. Caspase 3 activity (Figure 7A, 7B) was significantly (p < 0.01) increased by 92.39% in liver tissue homogenate, 92.98% in isolated hepatocytes, 93.51% in heart tissue homogenate and 94.16% in isolated cardiomyocytes as compared to the NC group. This result demonstrated that caspase activation was involved in MSG-induced cellular apoptosis which further justified the finding of immunohistochemistry data (Figure 8B, 9B). EECGL intervention group significantly (p < 0.01) reduced caspase 3 activity by 69.54% in liver tissue homogenate, 69.94% in isolated hepatocytes, 83% in heart tissue homogenate and 82.12% in isolated cardiomyocytes as compared with that MSG-treated group; while no such significant effect was observed in only EECGL treatment.

Nuclear translocation of NF-kB (p65) and cleaved caspase 3 by immunohistochemistry of NC, MSG, EECGL and MSG + EECGL treated groups of liver tissue. Nuclear translocation of (A) NF-kB (p65) and (B) cleaved caspase 3 of liver tissue sections were represented. The nuclei were stained by DAPI which appeared blue and NF-kB (p65) or cleaved caspase 3 was stained by FITC tagged secondary antibody which appeared green. The merged images showed the infiltration of green colour into the blue region which indicated the nuclear translocation of NF-kB (p65) or cleaved caspase 3. NF-kB (p65) or cleaved caspase 3 positive nuclei were estimated from the selected region marked by red colour. The intensity of DAPI and FITC was plotted using ImageJ software (NIH Image J system, Bethesda, MD) for quantification of fluorescence intensity. The intensity (in arbitrary unit) was taken along the Y-axis and distance (in pixels) was plotted along the X-axis. For NC, MSG, EECGL and MSG + EECGL groups the green line indicated the intensity of FITC and blue line indicated the intensity of DAPI.

Nuclear translocation of NF-kB (p65) and cleaved caspase 3 by immunohistochemistry of NC, MSG, EECGL and MSG + EECGL treated groups of heart tissue.nuclear translocation of (A) NF-kB (p65) and (B) cleaved caspase 3 of heart tissue sections were represented. The nuclei were stained by DAPI which appeared blue and NF-kB (p65) or cleaved caspase 3 was stained by FITC tagged secondary antibody which appeared green. The merged images showed the infiltration of green colour into the blue region which indicated the nuclear translocation of NF-kB (p65) or cleaved caspase 3. NF-kB (p65) or cleaved caspase 3 positive nuclei were estimated from the selected region marked by red colour. The intensity of DAPI and FITC was plotted using ImageJ software (NIH Image J system, Bethesda, MD) for quantification of fluorescence intensity. The intensity (in arbitrary unit) was taken along the Y-axis and distance (in pixels) was plotted along the X-axis. For NC, MSG, EECGL and MSG + EECGL groups the green line indicated the intensity of FITC and blue line indicated the intensity of DAPI.
Furthermore, to confirm the MSG-induced altered redox-homeostasis mediated apoptosis by the activation of caspase 3, immunofluorescence method was adopted (Figure 8, 9). MSG caused nuclear translocation of cleaved caspase 3; whereas cleaved caspase 3 remained localized in the cytoplasm of hepatocytes in NC group (3.95% nuclei was cleaved caspase 3 positive in case of liver and 2.63% nuclei were cleaved caspase 3 positive in case of heart) and EECGL (3.90% nuclei were cleaved caspase 3 positive in case of liver and 2.5% nuclei were cleaved caspase 3 positive in case of heart) fed group. In MSG group, 51.28% and 21.57% nuclei were cleaved caspase 3 positive within the selected region of liver and heart sections respectively; On the other hand, EECGL supplementation in MSG fed group decreased cleaved caspase 3 expression with 4.29% (in liver) and 6.86% (in heart) nuclear translocation of cleaved caspase 3 expressions. It can be speculated that MSG-induced generation of ROS and OS inactivated the c-Jun N-terminal kinase (JNK)-inactivating phosphatases to further promote the JNK activation and discharge of cytochrome c from the mitochondria for further activation caspase 3 mediated apoptosis 56 which was inhibited by EECGL. It can be hypothesized that, as TNF-α and OS are connected with a positive feedback loop therefore MSG induced OS augmented damage of deoxyribonucleic acid which further leads to the activation of TNF-receptor 1 (TNF-R1) signalling and ultimately caused programmed cell death. 57 Therefore, it can be postulated that EECGL prevent OS mediated programmed cell death via breaking the connection between TNF-α and OS, with partial inactivation of TNF-R1, NF-kB and caspase 3 signalling pathway. Figure 10 shows hypothetical target mechanism of action of Coccinia grandis to inhibit MSG-induced hepato-cardiac anomalies.

Possible target mechanism of action of Coccinia grandis to prevent MSG-induced hepato-cardiac anomalies.
Conclusions
The present investigation suggested from experimental data about the potential ameliorative efficacy of Coccinia grandis against MSG induced hepatic and cardiac derangements via blunting the expression of NF-kB, cleaved caspase 3 by regulating the balance of FR/EA to inhibit inflammation and apoptosis. Moreover, it was the first comprehensive report with molecular mechanism about the in vivo hepato-protective and cardio-protective impacts of Coccinia grandis with its natural antioxidants combats against MSG induced hepato-cardiac anomalies.
Footnotes
Acknowledgements
We are grateful to the University Grants Commission (UGC), Government of India for funding this study [Memo No.F.PSW-125/15-16 (ERO)] as a Minor Research Project. Authors are grateful to Prof. Dhrubajyoti Chattopadhyay, Vice Chancellor of Sister Nivedita University, Kolkata, for his continuous encouragement and valuable suggestions. Authors are indebted to Dr. Vansanglura, Principal of Serampore College, for his active administrative support and encouragement during the experiment. Authors are thankful to Ms. Debasmita Das, Mr. Sandipan Roy, Ms. Soume Chandra, Ms. Sohini Roy, Mr. Rajarshi Paul of Department of Physiology (UG & PG), Serampore College, Dr. Subrata Maiti and Mr. Rathin Maji for their technical help during this study.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Funding received from University Grants Commission (UGC), Government of India for funding this study [Memo No.F.PSW-125/15-16 (ERO)] as Minor Research Project.