The aim of this study was to investigate the protective effect of l-carnitine (l-CAR) in tamoxifen (TAM)-induced toxicity and antitumor activity. Adult female rats were randomly divided into four groups. Group I was served as control, groups II and III were treated with TAM (10 mg/kg, periorally) and l-CAR (300 mg/kg, intraperitoneally), respectively, while group IV was treated with both compounds. The treatment continued daily for 28 days. Administration of TAM resulted in significant increase in serum lipid profiles, liver enzymes, and bilirubin level. TAM produced a significant increase in lipid peroxides (LPO) level and nonsignificant change in nitrogen oxide (NO(x)) level accompanied with significant decrease in superoxide dismutase (SOD) activity of hepatic and uterus tissues and significant decrease in glutathione (GSH) content of uterus tissue. Administration of l-CAR for 1 h prior to TAM treatment decreased serum lipids and liver enzymes significantly and significantly increased SOD activity in liver and uterus tissues compared with TAM-treated group. Furthermore, it restored LPO and GSH levels and increased NO(x) level in uterus tissue. DNA fragmentation and the apoptotic marker, caspase-3, were not detected in the liver of all treated groups. Histopathologically, alterations in the liver and uterus structures after TAM treatment, which was attenuated after l-CAR administration. The antitumor effect and survival of the combined treatment of Ehrlich ascites carcinoma (EAC)-bearing mice was less than each one alone. l-CAR interestingly increased survival rate of EAC-bearing mice more than TAM-treated group. In conclusion, l-CAR has beneficial effects regarding TAM toxicity; however, it interferes with its antitumor effect.
Breast cancer is the leading cause of cancer deaths in women worldwide.1 Tamoxifen (TAM; 1-(4-(2-dimethyl-aminoethoxy) phenyl)—1,2-diphenyl-1-butene), a nonsteroidal antiestrogen drug, is a selective estrogen receptor modulator used in prevention and treatment of all stages of hormone-responsive breast cancer.2 Besides long treatment with TAM and its widespread uses, attention has been focused on its adverse effects, particularly liver and endometrium toxicity.3 According to studies, adjuvant TAM significantly increases women’s risk of subsequently developing endometrial carcinoma.4 Fatty liver was observed in more than 30% of breast cancer patients who received TAM treatment.5 Hepatotoxicity has been described with toxic hepatitis, multifocal hepatic fatty infiltration, submassive hepatic necrosis, and cirrhosis.6 In addition, TAM leads to oxidative liver damage and has been elucidated to be a hepatocarcinogen in rodents, which is due to overproduction of oxygen radical during TAM metabolism.7 It was demonstrated that TAM caused DNA adducts in rat liver and in the endometrium.8,9
l-CAR (l-trimethyl-3-hydroxy-ammoniabutanoate) is quaternary ammonium compound biosynthesized from amino acids lysine and methionine.10 It plays an essential role in human intermediary metabolism.11 The most prominent function lies in its role in the transport of activated long-chain fatty acids from the cytosol to the mitochondrial matrix where β oxidation and production of cellular energy takes place.12l-CAR has antioxidant properties and can reduce oxidative stress and acts as a free-radical scavenger. l-CAR has protective effects against acute and chronic doxorubicin toxicity in rat.13 In addition, it significantly inhibited preneoplastic lesions and hepatocarcinogenesis in animals.14l-CAR has a radioprotective and antidiabetic effects that attributed to its potent antioxidant activity.15l-CAR prevents oxidative stress and regulates nitric oxide (NO(x)).16 Also, l-CAR has a protective effect on the activity of mitochondrial enzyme succinate dehydrogenase as well as the activity of enzymes involved in defense against oxidative damage and 3-nitropropionic acid-induced neurotoxicity.16,17 In addition, cosupplementation of l-CAR (300 mg/kg) and lipoic acid (100 mg/kg) for 28 days improve the quality of living during the later stages of life by preventing free radical-induced damage to the proteins.18l-CAR supplementation is potentially beneficial with valproate-induced physiological disturbances.19
Therefore, the current study was designed to test the ability of l-CAR to protect the liver and endometrial tissues of TAM-treated rats from toxicity without impediments to the antitumor effect.
Materials and methods
Materials
Animals
Adult female Wistar albino rats weighing 120–170 g were obtained from the Egyptian Organization for Biological Products and Vaccines (VACSERA, Giza, Egypt) and female Swiss albino mice weighing 18–20 g were obtained from animal facility, Pharmacology unit, National Cancer Institute (NCI), Cairo University, Egypt. Animals were kept under standard conditions and were allowed free access to a standard requirement diet and water ad libitum. Animals were kept under a controlled lighting condition (light: dark, 13:11h). The animals’ treatment protocol has been approved by the animal care committee of the National Cancer Institute, Cairo University, Egypt.
Drug
TAM citrate was a kind gift from Medical Union Pharmaceuticals Company (Leiras/MUP, Cairo, Egypt). It is obtained as white powder soluble in sterile water and was diluted to the required concentration before use.
Chemicals
l-CAR (Sigma-Tau, Rome, Italy) was a generous donation from the Pharmacology Unit, National Cancer Institute (NCI), Cairo University, Egypt. All other chemicals and solvents used were of the highest purity grade available.
Methods
Toxicity study
Experimental design
In total, 24 female rats were divided randomly into 4 groups (6 animals in each group). Rats of group I was administered with 0.5 ml of normal saline and served as a control group. Rats of group II were administered with TAM (10 mg/kg, periorally (p.o.)).20 Rats of group III were injected with l-CAR (300 mg/kg, intraperitoneally (i.p.)).21 Rats of group IV were injected with l-CAR (300 mg/kg, i.p.); after 1 h, rats were treated with TAM (10 mg/kg, p.o.). The treatment schedule was continued once per day for 28 consecutive days.
Twenty-four hours after the last treatment, animals were anesthetized after exposure to ether in desiccators kept in a well-functioning hood. Blood samples were collected by heart puncture, and serum samples were separated. Livers and uterus were quickly excised, washed with saline, blotted with a piece of filter paper, and homogenized using a Branson sonifier (250, VWR Scientific, Danbury, Connecticut, USA). The homogenates were centrifuged at 800g for 5 min at 4°C to separate the nuclear debris. The supernatant so obtained was centrifuged at 10,500g for 20 min at 4°C to get the postmitochondrial supernatant, which was used to assay superoxide dismutase (SOD) activity.
Biochemical procedures in blood, liver, and uterus tissues
Lipid profile was determined using a standard commercial kit (Spectrum Diagnostics, Cairo, Egypt). Serum triglycerides (TGs), total cholesterol (CH), and high-density lipoprotein-cholesterol (HDL) were determined according to the methods described by Tietz et al.,22 Ellefson and Caraway, 23 and Warnick and Wood,24 respectively. Low-density lipoprotein-cholesterol (LDL) was calculated by subtracting the HDL-cholesterol from CH described by Terpstra et al.25 Aspartate aminotransferase (AST) and alanine aminotransferase (ALT) activities were determined according to the method described by Zilva and Pannall26 using spectrophotometric kit (Spectrum Diagnostics, Cairo, Egypt). Total and direct bilirubin levels were determined according to the method followed by Malloy and Evelyn.27
SOD activity, malondialdehyde (MDA), and reduced glutathione (GSH) content in liver and uterus homogenates were determined according to the methods followed by Minami and Yoshikawa,28 Buege and Aust,29 and Ellman,30 respectively. Total nitrate/nitrite (NO(x)) was measured as stable end product, nitrite, according to the method of Miranda et al.31
Determination of DNA fragmentation: Agarose gel electrophoresis
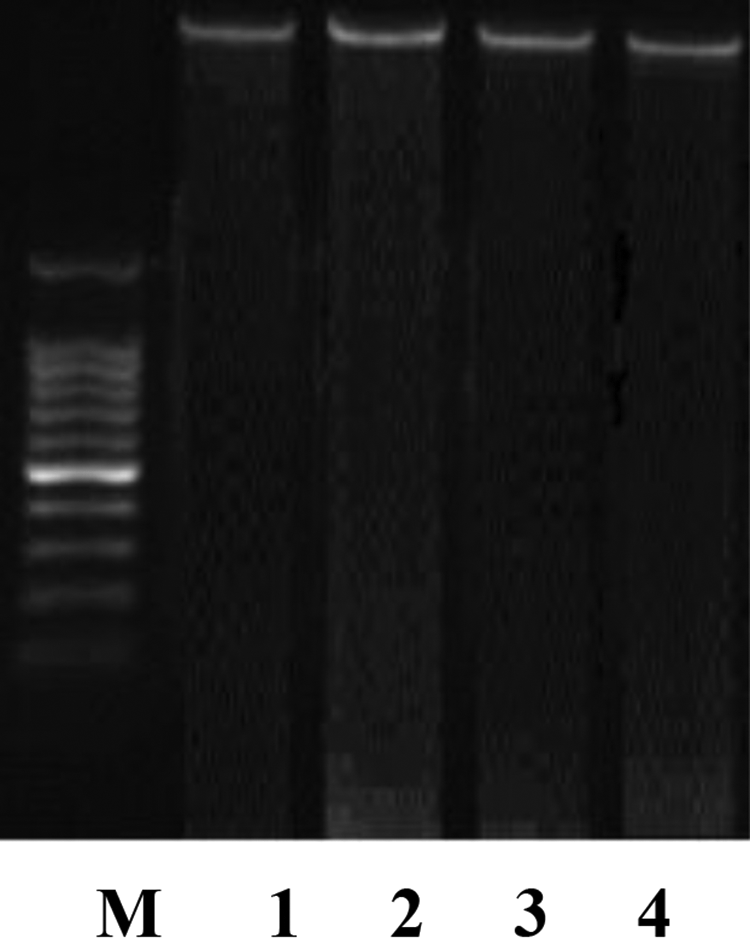
According to the method followed by Katoh et al.,32 the liver tissue was homogenized and lysed in a cold lysis buffer (10 mM Tris-hydrochloric acid (HCl), 5 mM disodium ethylenediaminetetraacetic acid (EDTA), and 0.5% Triton X-100, pH 8.0) for 10 min at 4°C. The DNA was sequentially extracted twice using half volumes of phenol/chloroform and incubated at 55°C for 10 min. After centrifugation at 800g for 20 min, the upper layer was incubated with proteinase K at 37°C for 60 min followed by incubation with ribonuclease at 37°C for 60 min. The DNA was precipitated by adding 10 M ammonium acetate and 100% ethanol and maintained at −20°C overnight. DNA was collected by centrifugation at 15,000g for 20 min, air-dried, and resuspended in Tris–EDTA buffer (10 mMTris-HCl, 5 mM EDTA, pH 7.4). Agarose gel electrophoresis33 was carried out for the analysis of DNA fragmentation. The resulting DNA preparations were electrophoresed through a 1.4% agarose gel containing ethidium bromide using Tris–boric acid–EDTA buffer (pH 8.3) at 40 V for 5 h. Equal quantities of DNA (based on optical density measurements at 260 nm) were loaded in each lane, and a molecular DNA marker was used as a molecular mass standard. DNA fragmentation was visualized and photographed under ultraviolet illumination.
Determination of apoptotic markers: Western blotting
The livers were homogenized with phosphate-buffered saline followed by centrifugation at 105,000g 4°C for 15 min. Protein content in the resulting supernatant was determined using Bradford reagent (Thermo scientific, Waltham, Massachusetts, USA). Equal volume of supernatant was mixed with 1× loading buffer and 5 µl of β-mercaptoethanol and boiled for 5 min. Proteins were separated by sodium dodecyl sulfate polyacrylamidegel electrophoresis and then electrotransferred onto polyvinylidene fluoride membrane using semi-dry transfer apparatus (Biometra, Germany).The membrane was blocked according to manufacturer’s instruction of chromogenic Western max detection kit (Ameresco, USA). After blocking with dilution buffer + 1% Tween-20 (DBT) + 1% bovine serum albumin, the membrane blots were incubated with indicated primary antibody (in DBT) at 4°C over night, then washed three times with DBT buffer, and incubated with horseradish peroxidase-conjugated secondary antibody for 4 h at room temperature. After washing three times with DBT buffer, the protein bands were visualized by 3,3′-diaminobenzidine (DAB) chromogen, which gives brown precipitate at the reaction site, specific protein bands on these transferred membranes were detected using the following antibodies: purfied anti-β-actin antibody was obtained from (BioLegend, San Diego, California, USA) and anti-mousecaspase 3 monoclonal antibody were obtained from (Bioscience, San Jose, California, USA).Relative expression of proteins was evaluated by normalizing the expression of proteins with quantitative housekeeping protein β-actin.34
Histopathological study
The samples were fixed in 10% neutral-buffered formalin, dehydrated through alcohols, cleared in xylene and then embedded in paraffin wax. Sections (5 mm thick) were stained with hematoxylin and eosin.35
Antitumor activity
Percentage survival of animals
Forty female albino mice inoculated with EAC cells (2.5 × 106cells/0.1 ml). Each mouse was injected i.p. with EAC cells. After 24 h of cell inoculation, 40 mice were classified into 4 groups. Group I animals were injected with 0.2 ml of saline and served as control. Group II animals were injected once with TAM (10 mg/kg, i.p.). Group III animals were injected once with l-CAR (300 mg/kg, i.p.). Group IV animals were injected once with l-CAR (300 mg/kg, i.p.) followed by TAM dose (10 mg/kg, i.p.) with an in-between interval of 60 min. The change in the percentage survival of animals was recorded daily during a period of 45 days following treatment.
Tumor volume
Forty female mice were used and solid tumor was transplanted subcutaneously in the right thigh of the lower limb of each mouse. Mice with a palpable tumor mass (100 m3) that developed within 7 days after implantation was divided into 4 groups: Group I: 10 mice were injected i.p. 4 times every other day with normal saline and served as control group. Group II: 10 mice were injected 4 times every other day with TAM (10 mg/kg, i.p.). Group III: 10 mice were injected 4 times every other day with l-CAR (300 mg/kg, i.p.). Group IV: 10 mice were injected with l-CAR (300 mg/kg, i.p.), 60 min later, injected with TAM (10 mg/kg, i.p.) 4 times every other day. The change in tumor volume was measured every other day using a Vernir caliper and calculated using the following equation according to Osman et al.36
where π equals 22/7, A and B denote the minor and major tumor axis, respectively.
Statistical analysis
Differences between obtained values (mean ± SEM; n = 6) were carried out by one-way analysis of variance (ANOVA) followed by the Tukey–Kramer multiple comparison test. The value of p ≤ 0.05 was taken as a criterion for a statistically significant difference.
Results
Effect on serum lipid profile
Treatment of rats with TAM (10 mg/kg for 28 days) produced a significant increase in serum levels of TG, CH, and LDL by 85.36, 40.78, and 129.07%, respectively, as compared to the control group. The combined effect of l-CAR and TAM resulted in a significant decrease in levels of TG by (25.88%) as compared to the TAM group, but it is significantly high in comparison to control group. On the other hand, the levels of CH and LDL were significantly decreased and reached the normal level (Table 1).
Effect of TAM, l-CAR, and their combination on the level of TG, CH, HDL and LDL in serum of female rats.a
aThe values are expressed as mean ± SEM of six rats per group. p0.05: significanta using ANOVA followed by Tukey–Kramer as a post ANOVA test. TAM (10 mg/kg, p.o.) and l-CAR (300 mg/kg, i.p.) were administered for 28 consecutive days. In the groups treated with two drug regimens, the animals were treated with l-CAR 1 h prior to TAM for 28 days.
bSignificantly different from the control group.
cSignificantly different from TAM-treated group.
Effect on serum liver function
AST, ALT, total bilirubin, and direct bilirubin of rats treated with TAM were significantly elevated by 20.85, 86.57, 104.65,and 172.09%, respectively, compared to the control group. On other hand, treatment with both l-CAR and TAM significantly decreased AST (19.12%), ALT (54.78%), total bilirubin (45.08%), and direct bilirubin (28.21%), as compared to the TAM-treated group, which still significantly increased compared to the control group (Table 2).
Effect of TAM, l-CAR, and their combination on the level of AST, ALT, total bilirubin and direct bilirubin in serum of female rats.a
aThe values are expressed as mean ± SEM of six rats per group. p0.05: significant using ANOVA followed by Tukey–Kramer as a post ANOVA test. TAM (10 mg/kg, p.o.) and l-CAR (300 mg/kg, i.p.) were administered for 28 consecutive days. In the groups treated with two drug regimens, the animals were treated with l-CAR 1 h prior to TAM for 28 days.
bSignificantly different from the control group.
cSignificantly different from TAM-treated group.
Oxidative status markers
Figure 1(a) to (d) shows the effects of TAM, l-CAR, and their combination on the activity of SOD, level of LPO, GSH content, and level of NO(x), respectively, in liver and uterus tissues. TAM treatment induced a significant decrease in the activity of SOD by 30.06 and 20.31% in liver and uterus, respectively. Administration of l-CAR prior to TAM induced significant increase in SOD activity (44.8%) in liver tissue and no significant change in uterus compared to the TAM-treated group (Figure 1(a)).
Treatment with TAM showed significant increase by 102.5and 118.44% in LPO levels of liver and uterus, respectively. Treatment with both l-CAR and TAM induced no significant decrease in LPO level of liver compared to the TAM-treated group while induced significant decrease in the level of LPO (34.63%) in uterus compared with the TAM-treated group; however, the value is still significantly high when compared with the control group (Figure 1(b)).
Effect of TAM, l-CAR, and their combination on (a) SOD expressed as microgram per gram tissue SOD activity, (b) LPO expressed in nanomoles per gram tissue LPO level, (c) reduced GSH expressed in micromoles per gram tissue GSH level, (d) total NO(x) expressed in nanomole per gram tissue NO(x) level in female rats for 28 consecutive days. Data are presented as percentage of control ± SEM of six rats per group. a′ and b′ denote significantly different from the control group and TAM-treated group at p0.05 using ANOVA followed by Tukey–Kramer as a post ANOVA test. TAM (10 mg/kg, p.o.) and l-CAR (300 mg/kg, i.p.) were administered for 28 consecutive days. In the groups treated with two drug regimens, the animals were pretreated with l-CAR 1 h prior to TAM and the administration was continued as described above. TAM: tamoxifen; l-CAR: l-carnitine; SOD: superoxide dismutase; LPO: lipid peroxides; GSH: glutathione; NO(x): nitrate/nitrite; ANOVA: analysis of variance; p.o.: periorally; i.p.: intraperitoneally.
Administration of TAM resulted in a significant decrease by 62.38% in uterus GSH level and no significant change in liver (Figure 1(c)). On the other hand, administration of l-CAR prior to TAM induced significant increase (100%) in GSH level of uterus compared with the TAM-treated group; however, the value is significantly decreased compared with the control group, while the value of GSH was still significantly high in liver compared with the control group.
Treatment of animals with TAM treatment induced nonsignificant change in level of NO(x) in the liver and uterus compared with the control group. Treatment with both l-CAR and TAM induced significant increase 29.26% in the level of NO(x) in uterus tissue compared to TAM-treated group (Figure 1(d)).
DNA fragmentation
Figure (2) shows the gel electrophoresis of DNA in liver tissue of female rat treated with TAM (10 mg/kg, p.o.) and/or l-CAR (300 mg/kg, i.p.) for 28 consecutive days did not show any change indicating no DNA fragmentation.
Gel electrophoresis of DNA in hepatic tissue, M: molecular weight marker, lane 1: liver DNA of normal control female rat, lane 2: liver DNA of female rat treated with TAM (10 mg/kg, p.o.), lane 3: liver DNA of female rat treated with l-CAR (300 mg/kg, i.p.), lane 4: liver DNA of female rat treated with l-CAR 1 h prior to TAM for 28 consecutive days. TAM: tamoxifen; l-CAR: l-carnitine; p.o.: periorally; i.p.: intraperitoneally.
Effect on the level of caspase 3 protein
The level of caspase 3 protein was determined by Western immunoblotting analysis. As it is observed there was no marked alteration in the level of caspase 3 protein in all treated groups (Figure 3).
Effect of TAM, l-CAR, and their combination on caspase 3 in hepatic tissue using blot technique. TAM (10 mg/kg, p.o.) and l-CAR (300 mg/kg, i.p.) were administered for 28 consecutive days. In the groups treated with two drug regimens, the animals were pretreated with l-CAR 1 h prior to TAM for 28 days. TAM: tamoxifen; l-CAR: l-carnitine; p.o.: periorally.
Histopathological findings
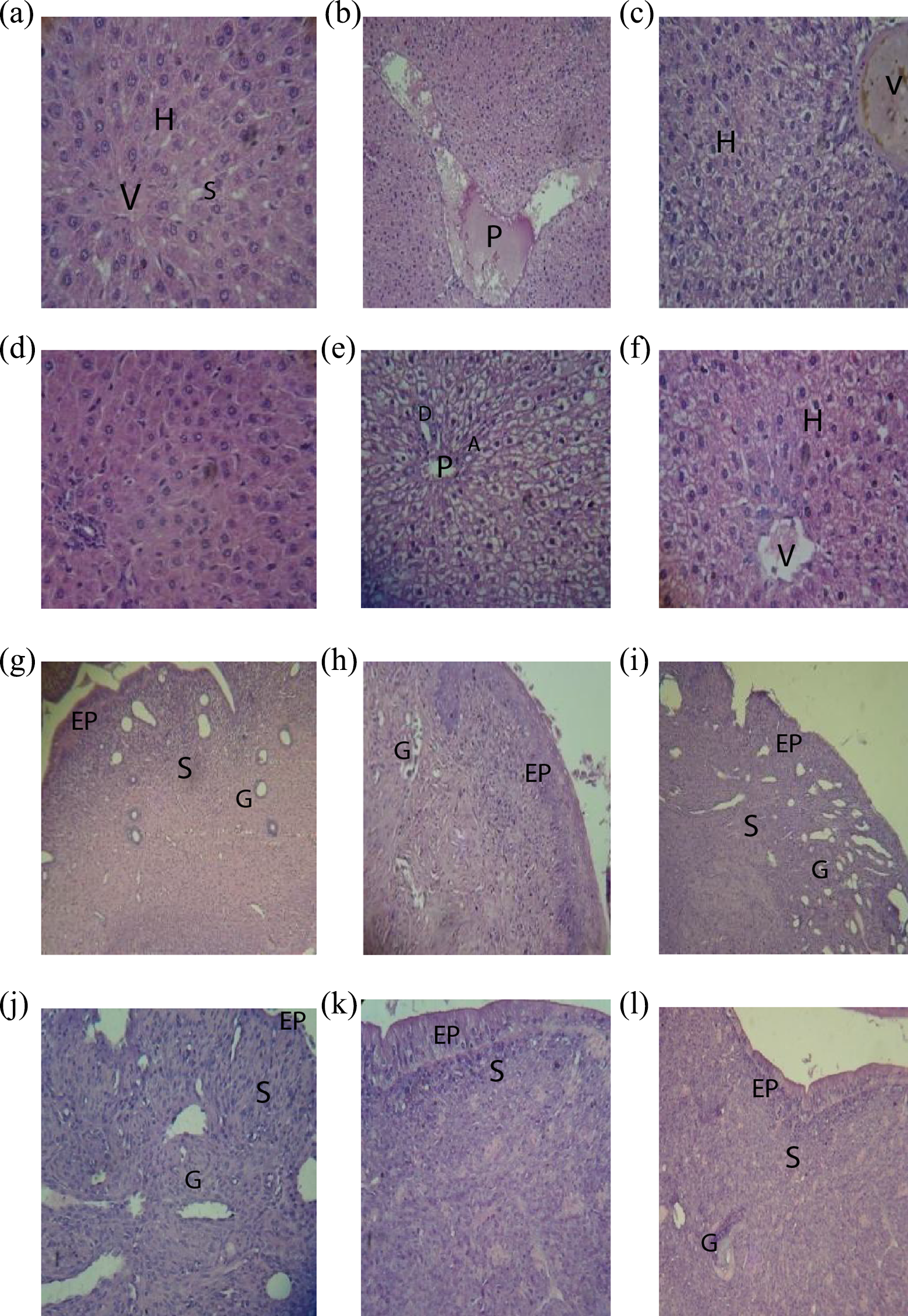
The protection effect of l-CAR against TAM-induced hepatotoxicity was further confirmed by conventional histopathological examination. Liver sections in the control group showed branching cords of hepatocytes and are radiating from the central vein (V). The hepatocytes (H) have vesicular nuclei, and some binucleated cells are separated by sinusoids (S) lined by flat endothelial cells and kupffer cells Figure 4(a). In Figure 4(d), liver section of rat liver treated with l-CAR group showed similar histological structure as control. Figure 4(b) and (c) photomicrograph of rat liver of TAM-treated group showed most of H around dilated and congested portal vein (P), and that the H around it have empty cytoplasm and dark nuclei. In addition, thickened and congested V, and the H with vacuolated cytoplasm were detected. In combined group, l-CAR and TAM showed relatively normal H around P, bile duct (b), and hepatic artery (A). However, the remaining cells are moderately swollen and vacuolated (Figure 4(e) and (f)).
Photomicrograph of effect of TAM, l-CAR, and their combination on hepatic and endometrium toxicities in female rats; (a): normal liver of control rats, (b,c): liver of rats treated with TAM (10 mg/kg, p.o. for 28 days); (d): liver of rats treated with l-CAR (300 mg/kg, i.p., for 28 days); (e,f): liver of rats treated with l-CAR 1 h prior to TAM for 28 consecutive days. (g): normal uterus of control rats, (h): uterus of rats treated with l-CAR, (i,j): uterus of rats treated with TAM, (k,l): uterus of rat treated with l-CAR 1 h prior to TAM for 28 consecutive days. (a–f, j) × 400 magnification; (g–i, k, and l) × 100 magnification H&E staining. H: hepatocytes; V: central vein; P: portal vein; EP: epithelium; S: stroma; G: endometrial glands; TAM: tamoxifen; l-CAR: l-carnitine; p.o.: periorally; H&E: hematoxylin and eosin.
Photomicrograph of rat endometrium of control group shows surface columnar epithelium (EP) and highly cellular stroma (S) containing numerous corkscrew endometrial glands (G) (Figure 4(g)). Section of rat endometrium of l-CAR-treated group showed no histological changes as compared to the control group (Figure 4(h)). Photomicrograph of rat endometrium of TAM-treated group showing degenerated surface epithelium (EP), partially ulcerated and edematous S containing tortuous G, hyperplasia S containing cells with deeply stained nuclei and endometrial glands (G) with its wide lumen (Figure 4(i) and (j)). Photomicrograph of rat endometrium of l-CAR with TAM-treated group showed recovery of the surface EP with nearly normal cellularS and most of the endometrium histologically architural (Figure 4(k) and (l)).
Antitumor activity
Percentage survival of animals
Figure (5) showed the percentage survival of EAC-bearing mice treated with TAM (10 mg/kg single i.p.) and/or l-CAR (300 mg/kg single i.p.). None of the control untreated tumor-bearing mice survived on day 23, while on day 24 of the experiment, none of the animals were survived in TAM-treated group. On the other hand, on day 41, none of the mice treated with l-CAR were survived, whereas none of the mice treated with l-CAR and TAM survived at day 19.
Effect of TAM, l-CAR, and their combination on the percentage survival rate of EAC-bearing mice. TAM (10 mg/kg, i.p.) and l-CAR (300 mg/kg, i.p.) were administered once. In the groups treated with two drug regimens, the animals were treated with l-CAR 1 h prior to TAM. TAM: tamoxifen; l-CAR: l-carnitine; i.p.: intraperitoneally; EAC: Ehrlich ascites carcinoma.
Tumor volume
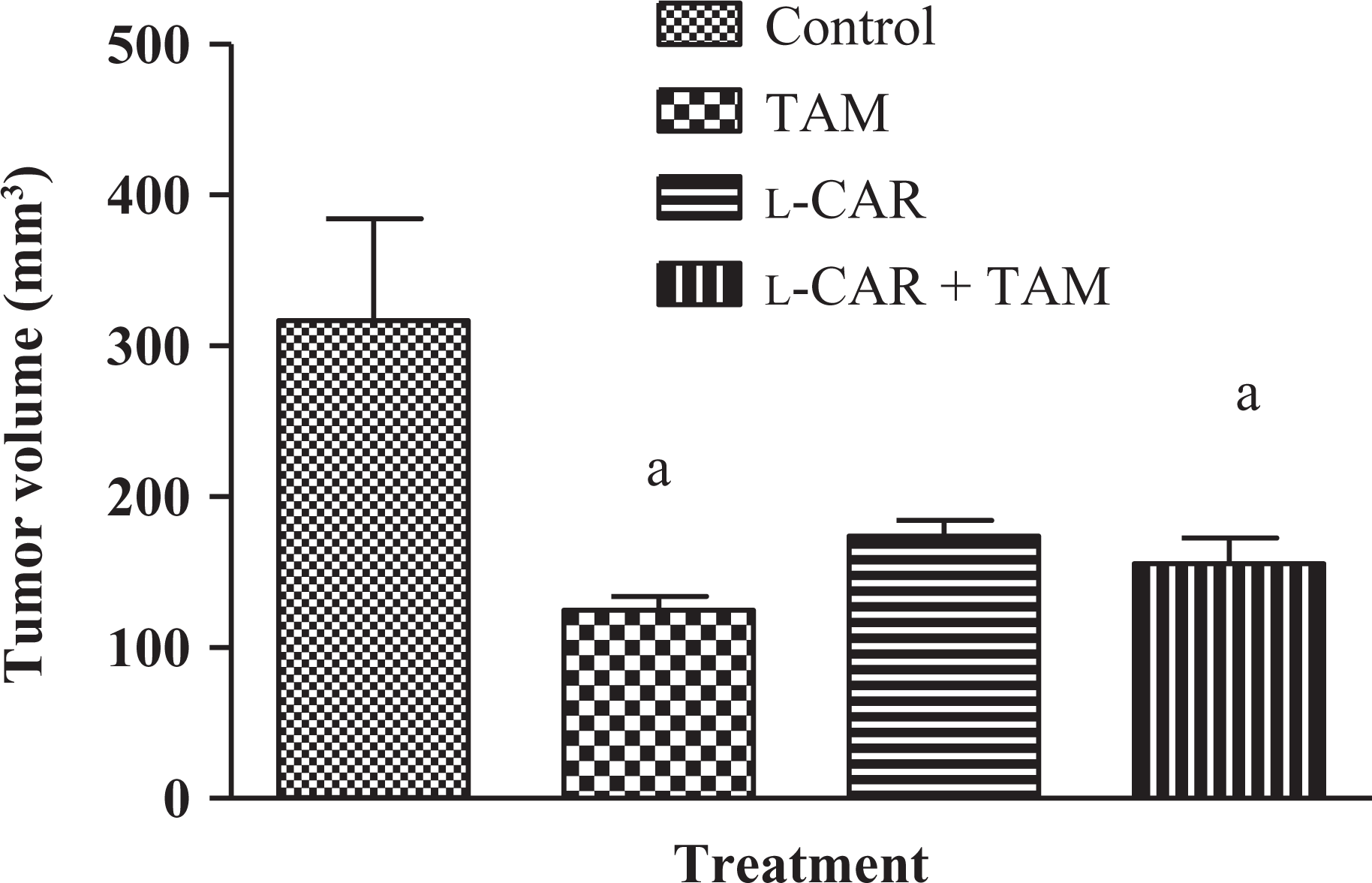
Figure (6) showed the effect of TAM, l-CAR, and their combination on the growth of solid Ehrlich carcinoma. The tumor volume of the control group showed progressive increase, whereas treatment of mice four times every other day with TAM (10 mg/kg, i.p.) resulted in a significant decrease in tumor volume as compared to control group. l-CAR alone induced nonsignificant decrease in tumor volume compared with the control group. Pretreatment with l-CAR in the combined group resulted in a significant decrease in tumor volume compared with the control group.
Effect of TAM, l-CAR, and their combination on the tumor volume of SEC-bearing mice. Data are expressed as mean ± SEM of six mice per group. p 0.05: significanta using ANOVA followed by Tukey–Kramer as a post ANOVA test.a: significantly different from the control group. TAM (10 mg/kg, i.p.) and l-CAR (300 mg/kg, i.p.) were administered four times. In the groups treated with two drug regimens, the animals were treated with l-CAR 1 h prior to TAM. TAM: tamoxifen; l-CAR: l-carnitine; ANOVA: analysis of variance; i.p.: intraperitoneally.
Discussion
TAM is an anti-estrogenic drug widely used for the treatment of all stages of breast cancer in spite of its hepatic and endometrium toxicities in rats3,37 and human.38 In the present study, TAM induced a significant increase in TG, CH, and LDL. Our results are in agreement with previous studies.39,40 They reported that the increased lipid profile after TAM administration was probably due to reduction in the hepatic ability of lipid β-oxidation that ultimately enhanced hepatic fat content, hypertriglyceridemia and increase in the rate of CH biosynthesis in the liver by TAM. Increase in TG level within normal range in most of the patients, while severe hypertriglyceridemia developed in a small number of them was reported in postmenopausal patients receiving adjuvant TAM.41
Our study demonstrated a significant increase in serum activity of AST, ALT, and level of bilirubin in TAM-treated rats. Our results are in agreement with the previous findings of Jain et al.42 and Kumarappan et al.43 who reported similar elevation in liver function. TAM has much higher affinity for hepatic tissue, its active metabolite, 4-hydroxytamoxifen induces conformational changes in the lipid bilayers of cell membranes, an effect that can lead to an increase in the membrane permeability and facilitates the passage of cytoplasmic enzymes outside the cells leading to the increase in the aminotransferase activities in blood.5l-CAR, in this study, significantly decreased the levels of TG, CH, and LDL and significantly inhibited the rise in liver function compared with the TAM-treated group. l-CAR is an essential cofactor in the transport of long-chain fatty acids from the cytosol to mitochondria for subsequent β-oxidation and production of cellular energy. In addition, it exhibits antioxidant and antiapoptotic activity.21,44 Previous study reported restoration of lipid profiles by l-CAR in rats.45
In consistent with previous studies,5,42,46 our study showed a significant increase in LPO with significant decrease in antioxidant levels in hepatic and endometrium tissue after TAM administration. TAM impaired β-oxidation of fatty acids6 and mitochondrial functions as it acts as an uncoupling agent and a powerful inhibitor of mitochondrial electron transport chain eventually resulting in mitochondrial oxidative damage.47 TAM decreased the activities of catalase, glutathione peroxidase, SOD, and content of GSH in liver; the decreased glutathione peroxidase activity leads to hydrogen peroxide (H2O2) accumulation in the liver, which in turns inactivates SOD.48 The decreased antioxidant power of TAM-intoxicated rats could be attributed to impairment of hexose monophosphate shunt and thereby reduced nicotinamide dinucleotide phosphatase availability, and the ability to recycle substances as GSH disulfide to GSH is decreased.3 However, Perumal et al.20 have shown that TAM inhibited LPO triggered by free radicals in 7,12 dimethylbenz(a)anthracene) induced peroxidative damage in rat mammary carcinoma.
Reduction in levels of LPO and elevation in antioxidants in l-CAR-treated rats suggested that l-CAR scavenges free radicals generated during oxidative stress. l-CAR had an effective superoxide anion radical scavenging, H2O2 scavenging, total reducing power, and metal chelating on ferrous ions activities.10,49 NO(x) has been implicated in various aspects of cancer biology, including both pro- and anti-tumor functions.50 In our study, neither TAM nor l-CAR alone had an effect on NO(x) level; however, significant increase in NO(x) in uterus tissue was observed when we use two drugs in combinations. We assume that this dual modulatory effects were not notable until both drugs were used resulting in significant increase in NO(x).
Our study showed no change in the expression of caspases 3 and intact genomic DNA was observed in normal hepatic tissue treated with TAM. The concept that TAM has favorable antineoplastic effect without any adverse effect on human normal breast tissue was indicated in benign biopsies of patients receiving TAM daily.51,52l-CAR was reported as an important antiapoptotic mediator.53l-CAR was reported to induce apoptosis in hepa1c1c7 cells by regulating Fas ligands and inhibiting the expression of Bcl-2 and inducing the upregulation of caspase 9 and caspase 3.54
Savitha and Panneerselvam55 have also observed that the l-CAR protected DNA-repairing enzymes, thereby preventing DNA strand break by maintaining thiol-containing compound and improving the GSH redox status, in addition, l-CAR decreased the levels of oxidative stress-mediated DNA damage during aging in rats. However, our results indicated histopathological changes in the architecture of the liver and uterus in TAM-treated group, which might be attributed to impaired β-oxidation of fatty acids and generation of ROS.5 It was found that TAM-induced DNA adduct formation in rat liver and uterus through metabolic activation to α-hydroxytamoxifen, which is sulfonated, forming an electrophilic carbocation that is capable of reacting with DNA yields a DNA adduct.56
l-CAR clearly ameliorated the histopathological changes induced by TAM and showed relatively normal and recovery of liver and uterus tissues. These findings support previous results indicating the antioxidant activity exhibited by l-CAR. In the present study, we have tested the effect of administration of TAM against induction of mammary carcinoma in female mice. Our study demonstrated that TAM decreased tumor volume and increased survival rate compared with the untreated control. Gabri et al.57 reported a significant reduction in tumor volume and a significant increase in the mean survival time of EAC-bearing animals in TAM-pretreated group. l-CAR as a monotherapy had an inhibitory effect on tumor volume.58 Niang and Melka59 reported that acetyl-l-CAR prolonged the survival of mice with the ascitic form of leukemia L1210 and l-CAR treatment prolonged survival of rats with adriamycin-induced heart failure by improving the myocardial metabolism of fatty acids.60
The combined effect of TAM and l-CAR on the survival in our study was less than the individual drug. It seems that l-CAR antagonize both the antitumor effect and survival of TAM upon administration of both drugs together. The antitumor effect and survival of the combined drug was less than each one alone. Interestingly, l-CAR increased survival rate of EAC-bearing mice more than TAM-treated group. The antagonistic effect of l-CAR with TAM may be due to interference with TAM metabolism, increase its excretion61 or may be due to drug interaction, which affects the cytotoxic effect of TAM. Also, this effect might be due to the antioxidant activity of l-CAR.10
Conclusion
This study illustrates that although l-CAR ameliorates TAM-induced hepatic and endometrium toxicities by preventing oxidative stress and enhancing antioxidant enzymes, it interferes with its antineoplastic activity.
Footnotes
Conflict of interest
The authors declared no conflicts of interest.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
References
1.
BuijsCde VriesEGEMouritsMJEWillemsePHB. The influence of endocrine treatments for breast cancer on health-related quality of life. Canc Treat Rev2008; 34: 640–655.
2.
MatsuokaHTsubakiMYamazoeYOgakiMSatouTItohTTamoxifen inhibits tumor cell invasion and metastasis in mouse melanoma through suppression of PKC/MEK/ERK and PKC/PI3K/Akt pathways. Exp Cell Res2009; 315: 2022–2032.
3.
StanleyLACarthewPDaviesRHigginsonFMartinEStylesJA. Delayed effects of tamoxifen in hepatocarcinogenesis-resistant Fischer 344 rats as compared with susceptible strains. Canc Lett2001; 171: 27–35.
4.
SinghMNStringfellowHFParaskevaidisEMartin-HirschPLMartinFL. Tamoxifen: important considerations of a multi-functional compound with organ-specific properties. Canc Treat Rev2007; 33: 91–100.
5.
AlbukhariAAGashlanHMEl-BeshbishyHANagyAAAbdel-NaimAB. Caffeic acid phenethyl ester protects against tamoxifen-induced hepatotoxicity in rats. Food Chem Toxicol2009; 47: 1689–1695.
6.
ParvezSTabassumHRehmanHBanerjeeBDAtharMRaisuddinS. Catechin prevents tamoxifen-induced oxidative stress and biochemical perturbations in mice. Toxicology2006; 225: 109–118.
7.
Senkus-KonefkaEKonefkaTJassemJ. The effects of tamoxifen on the femalegenital tract. Canc Treat Rev2004; 30: 291–301.
8.
ChaoHLiuJHongHLinJChenCChengT. L-carnitine reduces doxorubicin induced apoptosis through a prostacyclin-mediated pathway in neonatal rat cardiomyocytes. Int J Cardiol2011; 146: 145–152.
9.
WangSWangLYinJWangZFuPPYuH. Light-induced toxic effects of tamoxifen: a chemotherapeutic and chemopreventive agent. J Photochem Photobiol2009; 201: 50–56.
10.
GulcinI. Antioxidant and antiradical activities of l-carnitine. Life Sci2006; 78: 803–811.
11.
DoberenzJHircheFKellerUEderK. Pivalate lowers litter sizes and weights in female rats independent of its effect on carnitine status. Reprod Toxicol2007; 24: 83–88.
12.
VamosEVorosKVecseiLKlivenyiP. Neuroprotective effects of l-carnitine in a transgenic animal model of Huntington’s disease. Biomed Pharmacother2010; 64: 282–286.
13.
Sayed-AhmedMM. Role of carnitine in cancer chemotherapy-induced multipleorgan toxicity. Saudi Pharm J2010; 18: 195–206.
14.
HoangBXGraeme ShawDPhamPLevineS. Restoration of cellular energetic balance with L-carnitine in the neuro-bioenergetic approach for cancer prevention and treatment. Med Hypotheses2007; 69: 262–272.
15.
MansourHH. Effect of l-carnitine on endothelial dysfunction markers in diabetic-irradiated rats. Int J Toxicol Appl Pharmacol2013; 3(1): 1–9.
BiniendaZKAliSF. Neuroprotective role of l-carnitine in the 3-nitropropionic acid induced neurotoxicity. Toxicol Lett2001; 125: 67–73.
18.
SethumadhavanSChinnakannuP. Carnitine and lipoic acid alleviates protein oxidation in heart mitochondria during aging process. Biogerontology2006; 7(2): 101–109.
19.
El saidFGZeghiebrFE. The effect of L-carnitine on sodium valproate-induced physiological disturbances in Rats. Egypt J Hosp Med2007; 28: 347–354.
20.
PerumalSSShanthiPSachdanandamP. Combined efficacy of tamoxifen and coenzyme Q10 on the status of lipid peroxidation and antioxidants in DMBA induced breast cancer. Mol Cell Biochem2005; 273: 151–160.
21.
MuthuswamyADVedagiriKGanesanMChinnakannuP. Oxidative stress mediated macromolecular damage and dwindle in antioxidant status in aged rat brain regions: role of l-carnitine and dl-α-lipoic acid. Clin Chim Acta2006; 368: 84–92.
22.
TietzNWBodenTStepletonJD. An improved method for determination of lipase in serum. Am J Clin Pathol1959; 31: 148.
23.
EllefsonRDCarawayWT. Lipids and lipoproteins. In: TietzED (ed) Fundamentals of Clinical Chemistry. 2 nd. New York, Philadelphia: W.B. Saunder, 1976, p. 506.
24.
WarnickGWoodP. National cholesterol education program recommendation for measurement of high density lipoprotein cholesterol: executive summary. Clin Chem1995; 41: 1427–1433.
25.
TerpstraAHMSanchez-MunizFJWestCEWoodwardCJH. The density profile and cholesterol concentration on serum lipoprotein in domestic and laboratory animals. Comp Biochem Physiol1982; 71B: 669–673.
26.
ZilvaJFPannallPR. Plasma enzyme in diagnosis. In: Clinical chemistry in diagnosis and treatment. London, UK: Lioydluke, 1979, Chapter 17, p. 338.
27.
MalloyHTEvelynKA. The determination of bilirubin with the photoelectriccolorimetric method. J Biol Chem1937; 119: 481–490.
28.
MinamiMYoshikawaH.simplified assay method of superoxide dismutase activity for clinical use. Clin Chim Acta1979; 92: 337–342.
MirandaKMEspeyMGWinkDA. A rapid, simple Spectrophotometric method for simultaneous detection of nitrate and nitrite. Nitric Oxide2001; 5: 62–71.
32.
KatohKIkataTKatohSHamadaYNakauchiKSanoTInduction and its spread of apoptosis in rat spinal cord mechanical trauma. Neurosci Lett1996; 216: 9–12.
33.
YokozawaTDongE. Role of ginsenoside-Rd in cisplatin induced renal injury: special reference to DNA fragmentation. Nephron2001; 89: 433–438.
34.
SalamiSKarami-TehraniF. Biochemical studies of apoptosis induced by tamoxifen in estrogen receptor positive and negative breast cancer cell lines. Clin Biochem2003; 36: 247–253.
35.
HartzH. Simultaneous histologic fixation and gross demonstration of calcification. Am J Clin Pathol1947; 17: 750–755.
36.
OsmanAMSayed-AhmedMMKhayyalMTEL-MerzabaniMM. Hyperthermic potentiation of cisplatin on solid Ehrlich carcinoma. Tumori1993; 79: 268–272.
37.
KarkiAMantylaEHirsimakiYKarlssonSToikkanenSHirsimakiP. Comparison of the effects of tamoxifen and toremifene on rat hepatocarcinogenesis. Arch Toxicol2000; 74: 249–256.
38.
AkcayTDinnerYKayaliRColgarUOralECakatayU. Effects of hormone replacement therapy on lipid peroxides and oxidation system in postmenopausal woman. J Toxicol Environ Health2000; 59: 1–5.
39.
Akhondi-MeybodiMMortazavy-zadahMRHashemianZMoaiediM. Incidence and risk factors for non-alcoholic steatohepatitis in females treated with tamoxifen for breast cancer. Arab J G2011; 12: 34–36.
40.
LeeMKimJKimJKangKKongGLeeM. Gene expression profiling of murine hepatic steatosis induced by tamoxifen.Toxicol Lett2010; 199: 416–224.
41.
LiuCLYangTL. Sequential changes in serum triglyceride levels during adjuvant tamoxifen therapy in breast cancer patients and the effect of dosereduction. Breast Canc Res Treat2003; 79: 11–16.
42.
JainAKSwarnakarNKGoduguCSinghRPJainS. The effect of the oral administration of polymeric nanoparticles on the efficacy and toxicity of tamoxifen. Biomaterials2011; 32: 503–515.
43.
KumarappanCVijayakumarMThilagamEBalamuruganMThiagarajanMSenthilSProtective and curative effects of polyphenolic extracts from Ichnocarpusfrutescense leaves on experimental hepatotoxicity by carbontetrachloride and tamoxifen. Ann Hepato2011; 10: 63–72.
44.
Gómez-AmoresLMate ARevilla ESanta-MarıaCVázquezCM. Antioxidant activity of propionyl-l-carnitine in liver and heart of spontaneously hypertensive rats. Life Sci2006; 78: 1945–1952.
45.
SidoriakNGVolginDV. Effect of L-carnitine on lipid peroxidation and lipidcomposition in blood serum in hemic hypoxia. Ukr Biokhim Zh1996; 68: 54–61.
46.
TabassumHRehmanHBanerjeeBDRaisuddinSParvezS. Attenuation of tamoxifen-induced hepatotoxicity by taurine in mice. Clin Chim Acta2006; 370: 129–136.
KakkarRManthaSRadhiJPrasadKKarlaJ. Antioxidant defense system in diabetic kidney. Life Sci1997; 60: 667–679.
49.
KalaiselviTPanneerselvamC. Effect of l-carnitine on the status of lipidperoxidation and antioxidants in aging rats. J Nutr Biochem1998; 9: 575–581.
50.
OstadSNManeshiASharifzadehMAziziE. Effect of 17-β estradiol on the expression of inducible nitric oxide synthase in parent and tamoxifen resistantT47D breast cancer cells. Iran J Pharmaceut Res2009; 8: 125–133.
51.
de LimaGRFacinaGShidaJYCheinMBCTanakaPDardesRCEffects of low dose tamoxifen on normal breast tissue from premenopausal women. Eur J Canc2003; 39: 891–898.
52.
WalkerKJPrice-ThomasJMCandlishWNicholsonRI. Influence of the antiestrogen tamoxifen on normal breast tissue. Br J Canc1991; 64: 764–768.
53.
MorettiSFamularoGMarcelliniSBoschiniASantiniGTrinchieriVl-carnitine reduces lymphocyte apoptosis and oxidant stress in HIV-1-infected subjects treated with zidovudine and didanosine. Antioxid Redox Signal2002; 4: 391–403.
54.
FanJPKimHSHanGD. Induction of apoptosis by l-carnitine through regulation of two main pathways in Hepa1c1c7 cells. Biomed Life Sci2009; 36: 365–372.
55.
SavithaSPanneerselvamC. Mitigation of age-dependent oxidative damage to DNA in rat heart by carnitine and lipoic acid. Mech Ageing Dev2007; 128: 206–212.
56.
CostaGMarquesMMFuXChurchwellMIWangYDoergeDREffect of N, N-didesmethyl tamoxifen upon DNA adduct formation by tamoxifen and α-hydroxytamoxifen. Canc Lett2007; 257: 191–198.
57.
GabriMSOsmanAMEl-SayedMMNegmSA. Prophylactic effect of tamoxifen against induction of mammary carcinoma. Egypt J Hosp Med2004; 14: 104–114.
58.
Sayed-AhmedMMShaarawySShoumanSAOsmanAM. Reversal of doxorubicin-induced cardiac metabolic damage by l-carnitine. Pharmacol Res1999; 39: 289–296.
59.
NiangMMělkaM. Effect of acetyl-l-carnitine on leukemia L1210, resistant to mitoxantrone. Acta Med2000; 43: 125–128.
60.
KawasakiNDaeleeJShimizuHUedaT. Long–term l-carnitine treatment prolongs the survival in rats with adriamycin-induced heart failure. J Card Fail1996; 2: 293–299.
61.
OlszowyZPlewkaACzechENowickaJPlewkaDNowaczykGEffect of l-carnitine supplementation on xenobiotic-metabolizing hepatic enzymes exposed to methanol. Exp Toxicol Pathol2006; 57: 427–435.