
Abstract
The aim of the present research was to examine the toxic influence of different doses of zearalenone (ZEN) on the liver, especially oxidative stress induced by ZEN on the liver. A total of 48 pregnant Sprague-Dawley rats were randomly assigned into 4 treatments groups with 12 animals in each. The rats were fed with a normal diet treated with 0 mg/kg (control), 50 mg/kg (treatment 1), 100 mg/kg (treatment 2), or 150 mg/kg (treatment 3) ZEN in feed on gestation days (GDs) 0–7 and then all the rats were fed with a normal diet on GDs 8–20. The experimental period lasted 21 days. The results showed that exposure to ZEN induced increase in aspartate amino transferase, alanine aminotransferase, and alkaline phosphatase activities and decrease in total protein and albumin content in a dose-dependent manner and also induce decrease in superoxide dismutase and glutathione peroxidase activities and increase in malondialdehyde content in a dose-dependent manner in the serum and the liver. The increased transcription of cytochrome P450 2E1 (CYP2E1) was detected in the liver after exposure to ZEN. These results suggested that ZEN not only caused damage in the liver of pregnant rats in a dose-dependent manner but also induced the messenger RNA expression of CYP2E1 in the liver.
Introduction
Zearalenone (ZEN) is a mycotoxin that is mainly produced by various fungi of the Fusarium genus, which are frequently found in cereals such as maize, barley, oats, wheat, rice, and other plant products. The prevalence of ZEN in cereal has resulted in the worldwide contamination of food and animal feeds. 1,2 ZEN is rapidly absorbed following oral intake and is metabolized mainly in the liver and the intestine. It is transformed into α- and β-zearalenol, α- and β-zearalanol, and ZEN, all of which are subsequently conjugated to glucuronic acid. 3 ZEN is genotoxic and has been shown to have potent reproductive toxicity in humans and animals. 4–6
ZEN has been shown to be immunotoxic, 7–9 hepatonephrotoxic, 10–12 and induce apoptosis. 13,14 ZEN also induces DNA fragmentation, 13,15 micronuclei production, and chromosome aberrations. 16,17 Many xenobiotics and drugs were found mainly metabolized in the liver, which expresses the most prominent class of biotransformation enzymes, but other organs including the lungs, kidneys, and intestines may contribute to overall xenobiotic biotransformation. Differences in biotransformation enzyme activities alter the systemic bioavailability and subsequently the efficacy of drugs; they may also provide protection against certain xenobiotics and environmental pollutants, but can increase the toxicity of others. 18 ZEN is metabolized mainly in the liver, which was found to be hepatotoxic; it affected some enzymatic parameters of the hepatic function in rats 19 and in rabbits. 20 The degree of the damage to liver can be assessed by testing specific enzymes. 21
The toxic effects of ZEN and its metabolites have been primarily ascribed to its chemical structure, which resembles that of naturally occurring estrogens. 3 However, the exact underlying mechanisms remain largely unknown. Oxidative stress has been considered an important mediator of the toxic effects incurred after exposure to ZEN. In fact, recent investigations have shown the oxidative metabolism of ZEN in the liver of rodent and human in vitro systems. 22–24 Additionally, some reports have suggested that oxidative stress is a key determinant of ZEN-induced toxicity in vivo and in vitro. 5,8,25–27 Oxidative damage has been described in rats fed with diets containing high levels of ZEN. 28 Moreover, it has been demonstrated that ZEN and it metabolites induces lipid oxidation and increases the production of malondialdehyde (MDA) in several cell lines. 27,29,30 Abid-Essefi et al. found that ZEN induced oxidative damage by enhancing lipid peroxidation in the two cell lines (Vero and Caco-2 cells), since ZEN increased MDA formation in a concentration-dependent manner. 31
The early events of pregnancy are associated with rapid changes in the expression of genes required for nutrient transport, cellular remodeling, angiogenesis, as well as cell proliferation and migration. 32,33 The aim of this research was to examine the toxic influence of different doses of ZEN on the liver of pregnant rats during the early gestation. For this purpose, the feature of liver damage was examined by the measurement of some enzymatic parameters of the hepatic function. In addition, the oxidative stress indices, glutathione peroxidase (GPx), superoxide dismutase (SOD), and lipidperoxidation product (MDA) activities were determined in serum and in liver homogenates. Moreover, the effect of the messenger RNA (mRNA) expression of cytochrome P450 2E1 (CYP2E1) and UDP-glucuronosyltransferase (UGT) 1A1 (UGT1A1) induced by ZEN were examined in the liver.
Materials and methods
Chemicals
ZEN was purchased as pure crystals from Fermentek Ltd. (Jerusalem, Israel). The substance was stable for at least 8 months.
Animals and experimental design
Sprague-Dawley rats were obtained from Jilin University Laboratory Animal Centre (Changchun, China) and were acclimated for 1 week. Male rats weighed 300–325 g, and female rats weighed 190–210 g (9 weeks). The animals were maintained according to the National Research Council Institute of Laboratory Animal Resources. 34 The rats were kept in plastic rat cages and had free access to food and water. They were kept in the study room on a 12-/12-h light–dark cycle under a controlled temperature ranging between 22 and 26°C at a relative humidity of 47–55%. Male rats were used only as sires and were not treated. Throughout the study, male and female rats were fed with experimental animal ration (University Laboratory Animal Centre, Changchun, China). The feed was prepared in an experimental feed mill with high precision mixers, and all feeding stuffs were subjected to post-processing analytical control. All of the experimental procedures involving animals were carried out in compliance with the Polish legal regulations determining the terms and methods for performing experiments on animals (opinion of the Local Ethics Committee for Animal Experimentation No. 14/N of 26 January 2005). Nominal concentrations were compared using validated analytical methods, National Standards of People’s Republic of China (GB/T 19540—2004). 35 The findings are presented in Table 1.
ZEN concentrations in feed and number of treated rats per treatment.
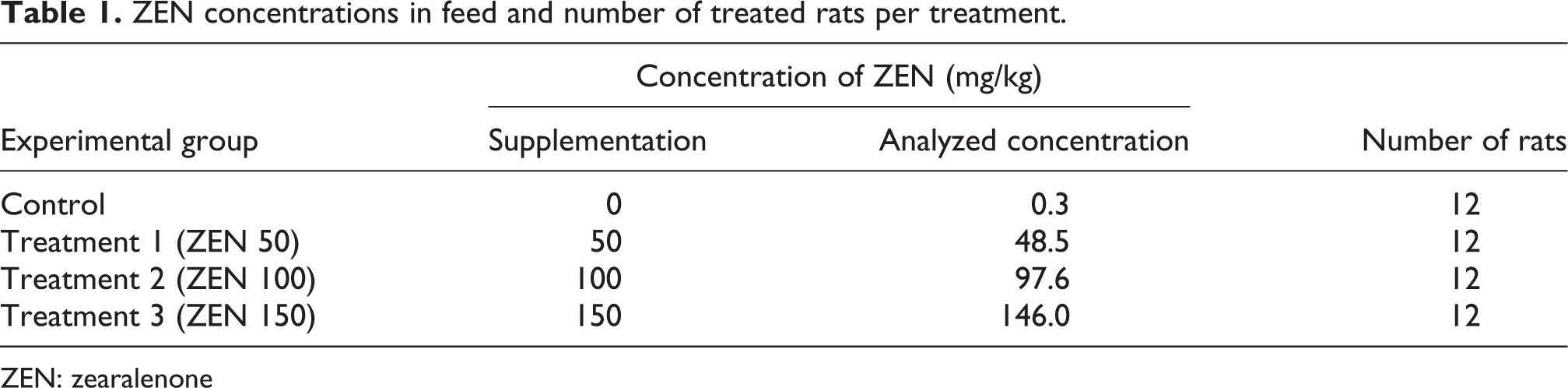
ZEN: zearalenone
Following acclimation, the females were mated (two females per male). Cohabitation began at approximately 4:30 p.m. on each mating day. The next morning, each female was examined at 8:00 a.m. for the presence of sperm in the vaginal lavage. A total of 48 sperm-positive females were presumed pregnant at gestation day 0 (GD 0), and a stratified random procedure was used to assign each animal to the control group or one of the three treatment groups. 36
All the 48 pregnant rats were randomly assigned into 4 treatment groups with 12 animals in each. The pregnancy of the treated rat was normal and the average weight of the animals in the four groups was similar (205.6, 205.8, 204.9, and 205.3 g, respectively) at the beginning of the experiment. The rats were fed with a normal diet treated with 0 mg/kg (control), 50 mg/kg (treatment 1; ZEN 50 group), 100 mg/kg (treatment 2; ZEN 100 group), or 150 mg/kg (treatment 3; ZEN 150 group) ZEN in feed on GDs 0–7 and then all the rats were fed with a normal diet on GDs 8–20. The experimental period lasted 21 days and the animals were killed at GD 20. For blood collection, microcentrifuge tubes were coated with 10 mL heparin sodium salt (20 U/mL in phosphate-buffered saline). Whole blood was collected from the submandibular site as described above and kept on ice for 30 min. Samples were centrifuged at 3000g for 10 min at 4°C. Serum layers were removed and placed into sterile microcentrifuge tubes and stored at –80°C until assay. Fragments of maternal liver were quickly removed, frozen in liquid nitrogen, stored at −80°C, and analyzed for gene expression by RNA extraction followed by quantitative reverse transcriptase polymerase chain reaction (RT-PCR). Other fragments were stored at −20°C for antioxidant enzymes measurements.
Determination of biochemical parameters
The biochemical parameters were determined with the automated biochemical analyzer (SYNCHRON CX®4 PRO model, Beckman Coulter Inc., USA) using commercial diagnostics kits (Biosino Bio-Technology and Science Inc, Beijing, China). Parameters were determined for the following biochemical characteristics of serum: aspartate amino transferase (AST), alanine aminotransferase (ALT), alkaline phosphatase (ALP), total protein (TP), albumin (ALB), and total bilirubin (TBIL).
Determination of antioxidative parameters
The antioxidative parameters were determined with ultraviolet spectrophotometer (UV-2410PC model, SHIMADZU, Japan) using commercial diagnostics kits (Nanjing Jiancheng Biotechnology Co., Ltd, Jiangsu, China). The parameters evaluated in the serum and liver were MDA, SOD, and GPx.
Total RNA isolation, integrity, and quantification
Total RNA was isolated from tissue samples (30 mg liver tissue) using TRIzol reagent according to the manufacturer’s instructions (Tiangen Biotech (Beijing) Co., Ltd, Beijing, China). The dried RNA pellets were resuspended in 50 μL of diethyl-pyrocarbonate-treated water. The concentration and purity of the total RNA was determined using a spectrophotometer at 260/280 nm. First-strand complementary DNA (cDNA) was synthesized from 5 μg of total RNA using oligo dT primers and Superscript II reverse transcriptase according to the manufacturer’s instructions (Tiangen Biotech (Beijing) Co., Ltd). Synthesized cDNA was diluted five times with sterile water and stored at −80°C before use.
Quantitative RT-PCR
Assays were performed within a few days (≤1 week) following total tissue RNA isolation and quantification. Real-time quantitative RT-PCR was used to detect the expression of CYP2E1 and UGT1A1 genes in the liver using SYBR Premix Ex Taq (Takara, Shiga, Japan). Real-time PCR was also performed on liver samples. Reaction mixtures were incubated in the ABI PRISM 7500 real-time PCR system (Applied Biosystems, Foster City, California, USA). The program set was 1 cycle at 95°C for 30 s and 40 cycles at 95°C for 5 s and at 61°C for 34 s. Dissociation curves for each PCR reaction were analyzed with dissociation curve 1.0 Software (Applied Biosystems) to detect and eliminate possible primer–dimers and nonspecific amplification. The relative expression ratios of the target genes were calculated based on the efficiencies and quantification cycle deviations of unknown samples versus controls and expressed in comparison with the reference genes according to Pfaffl. 37 The primer sequences and product sizes are shown in Table 2.
Primers used for quantitative real-time PCR.

PCR: polymerase chain reaction; CYP2EI: cytochrome P450 2E1; UGT1A1: UDP-glucuronosyltransferase (UGT) 1A1.
Data analysis
Statistical analysis was performed using the Statistical Package for Social Science 18.0 (SPSS 18.0; SPSS Inc., Chicago, Illinois, USA). One-way analysis of variance was used to detect differences among groups and the means were compared by Duncan’s multiple comparison test. The variability degree of the results is expressed as mean ± SEM. p < 0.05 was considered significant.
Results
The effect of ZEN on the biochemical parameters in the serum of pregnant rats
The biochemical data are presented in Table 3. The activity levels of AST, ALT, ALP, and TBIL in the serum gradually increased with the concentration of ZEN treatment in a concentration-dependent manner. The results from the serum biochemical analysis revealed that treatment with ZEN (50, 100, or 150 mg/kg) significantly increased the serum activity levels of AST, ALT, ALP, and TBIL compared with those in the control group (p < 0.05), and the serum activity levels of AST, ALP, and TBIL in ZEN 150 group were significantly higher than those in ZEN 50 and ZEN 100 groups (p < 0.05). The serum activity level of ALT in ZEN 100 and ZEN 150 groups were higher than that in ZEN 50 group (p < 0.05). Conversely, the levels of serum TP and ALB in all the ZEN-treated groups (treatments 1–3) were significantly decreased compared with those in the control group (p < 0.05), and the levels of serum TP and ALB in ZEN 100 and ZEN 150 groups were significantly lower than those in ZEN 50 group (p < 0.05).
Effect of ZEN on biochemical parameters in serum of pregnant rats.a

ZEN: zearalenone; AST: aspartate amino transferase; ALT: alanine aminotransferase; ALP: alkaline phosphatase; TP: total protein; ALB: albumin; TBIL: total bilirubin.
aMeans within a line with different superscript letters are significantly different at p < 0.05.
The effect of ZEN on antioxidative parameters in the serum and the liver of pregnant rats
In order to evaluate the role of oxidative stress on the effects induced by different concentrations of ZEN, we measured several enzymatic indicators of oxidative stress in the serum and the liver. On the other hand, activities of enzymatic markers for oxidative stress were altered by ZEN treatment. The effects of different treatments on MDA, SOD, and GPx activities in the serum and the liver are depicted in Tables 4 and 5. The activities of the antioxidant enzymes GPx and SOD in the serum and the liver were decreased while the levels of MDA were increased with the concentration of ZEN improved in a dose-related manner. Two ZEN treatment groups (100 or 150 mg/kg) showed a significant decrease in SOD and GPx in the serum and the liver compared with levels in the control group (p < 0.05), and the activities of GPx and SOD in the serum and the liver in ZEN 100 and ZEN 150 groups were significantly lower than those in ZEN 50 group (p < 0.05). In contrast, MDA levels in all the ZEN-treated groups were significantly increased compared with those in the control group (p < 0.05), and the levels of MDA in the serum and the liver in ZEN 100 and ZEN 150 groups were significantly higher than those in ZEN 50 group (p < 0.05).
Effect of ZEN on antioxidative parameters in serum of pregnant rats.a

ZEN: zearalenone; MDA: malondialdehyde; SOD: superoxide dismutase; GPx: glutathione peroxidase.
aMeans within a line with different superscript letters are significantly different at p < 0.05.
Effect of ZEN on antioxidative parameters in liver of pregnant rats.a

ZEN: zearalenone; MDA: malondialdehyde; SOD: superoxide dismutase; GPx: glutathione peroxidase.
aMeans within a line with different superscript letters are significantly different at p < 0.05.
The effects of ZEN on the mRNA expression of CYP2E1 and UGT1A1 in the liver of pregnant rats
The abundance of mRNA for two enzymes (CYP2E1 and UGT1A1) was measured by qRT-PCR as shown in Table 6. Compared with the control group, there was a significant increase (p < 0.05) of CYP2E1 mRNA levels in all ZEN-treated groups. The CYP2E1 mRNA levels of liver in ZEN 100 and ZEN 150 groups were significantly higher than that in ZEN 50 group (p < 0.05). There was an upward trend in the UGT1A1 mRNA levels in the ZEN-treated groups, but the difference was not significant (p > 0.05) when compared with the control group.
Effects of ZEN on the mRNA expression of CYP2E1 and UGT1A1 in pregnant rat liver.a

mRNA: messenger RNA; CYP2EI: cytochrome P450 2E1; UGT1A1: UDP-glucuronosyltransferase (UGT) 1A1.
aMeans within a line with different superscript letters are significantly different at p < 0.05.
Discussion
Although aflatoxins and ochratoxins have been the focus of many studies over the last decade, fusariotoxin like ZEN have received little attention. This has changed recently, with fusarium toxins including ZEN becoming the focus of intense study.
The toxicity of ZEN manifests in reproductive tracts, 38,39 livers, 20 and kidneys. The degree of damage to such tissues or in the whole body can be assessed by testing specific enzymes. 21 In a clinical diagnosis, determination of the transaminases is of great importance. 40 For example, the intracellular enzyme AST is released into the blood in proportion to the level of damaged cells. In liver function disorders, ALT is also a specific and sensitive indicator of acute hepatic necrosis. 41 ALP in tissues takes part in the transport of phosphate ions through cell membranes. Pathological ALP levels increase in liver diseases. 42
In the present study, analysis of the results revealed statistically significant differences for AST, ALT, and ALP activities between the control group and groups of rats that were given different doses of ZEN during GDs 0–7. This reflected initial hepatocellular damage as described by Čonková et al. 20 Changes in these biological parameters were observed in similar studies. Maaroufi et al. 19 reported some modifications of hematological and biochemical parameters after rats were treated intraperitoneally (i.p.) with 1.5, 3, or 5 mg/kg body weight (b.w.) of ZEN. Compared with controls, blood biochemical markers such as bilirubin (total and conjugated), ALT, AST, and ALP were all significantly increased in ZEN-treated animals. These increased levels indicated liver toxicity. Mice that were orally given ZEN at 40 and 500 mg/kg of body mass showed significantly increased blood levels of AST, ALT, and ALP after 48 h. 43 ZEN (2.7 mg/kg b.w.) treatment significantly increased ALT, AST, and ALP activities after 48 h and two weeks, whereas ALT was also significantly increased after 1 week. 44
Most serum proteins are synthesized in the liver, and the total level of proteins in the blood is a main test of liver function. The levels of these proteins may change due to some diseases or certain types of tissue damage. Protein synthesis of the liver was tested by sampling TP and ALB levels. Mice treated with ZEN showed a significant decrease in TP and ALB. The decreased levels of TP and ALB may be due to the inhibition of transfer RNA-synthetase that is accompanied by reduced protein synthesis. 45
In the present study, the activities of TP and ALB gradually decreased as the concentration of ZEN increased. Furthermore, results showed statistically significant differences between the control group and the groups treated with ZEN. Protein synthesis was affected by ZEN, and this further confirmed that the liver was damaged. Similar results were observed by Liang et al., 46 who demonstrated that after different single doses of ZEN (25, 50, and 100 mg/kg) were injected into mice, the levels of TB and ALB were decreased. In addition, male Balb/c mice that received an oral treatment of ZEN (40 mg/kg i.p.) for 28 days had significantly decreased TP and ALB activities in their testes. 5
Oxidative stress is a disturbance in the balance between pro-oxidants and antioxidants that results in potential cell damage. The targets of oxidative damage are usually critical biomolecules such as nucleic acids, protein, and lipids. 31,47 Lipid peroxidation is one of the cellular pathways involved in oxidative damage. The capability of ZEN to cause oxidative stress by inducing lipid peroxidation was demonstrated by Kouadio et al. 29
Antioxidant enzymes are a major defense system against liver injury and carcinogenesis. GPx is an important component of cellular antioxidant defense, and the decrease in GPx activity was considered as a key indicator of oxidative injuries. In the present study, the activities of the antioxidant enzymes GPx and SOD in the serum and the liver were significantly decreased in ZEN 100- and ZEN150-treated rats compared with those in the control group (p < 0.05). These results are in agreement with the findings of Ben Salah-Abbès et al., 5 who reported that male Balb/c mice orally treated for 28 days with ZEN (40 mg/kg i.p.) had significantly decreased GPx, SOD, and CAT activities in their testes. The serum level of GPx was not significantly influenced in ZEN 50 group compared with the control group (p > 0.05). The result may be explained by the low dose of ZEN use. Stadnik et al. found that there were no changes observed in GPx activities in groups that were given low doses of ZEN (1, 2, or 3 mg/kg b.w). 48 In the current study, ZEN was found to decrease the levels of GPx and SOD in the liver. The decreased level of GPx may be explained by the conjugation of GPx with ZEN or its metabolites. 49
MDA is an end product of lipid peroxidation that is considered a late biomarker of oxidative stress and cellular damage. 50 In the current study, MDA in all the ZEN-treated groups was significantly increased with respect to the control group. This result is consistent with Ben Salah-Abbès et al., 5 who found that male Balb/c mice orally treated for 28 days with ZEN (40 mg/kg, i.p.) had significantly increased MDA activity in their testes.
CYP2E1 plays an important role in the toxicity of many chemicals and of ethanol, and CYP2E1 produces oxidant stress. Indeed, a number of studies using cultured cells have established the importance of CYP2E1 in oxidative stress. 51
The mechanism by which CYP2E1 induces or potentiates liver toxicity is believed to be through the production of reactive oxygen species such as superoxides, hydrogen peroxide, and hydroxyl radicals. Bai and Cederbaum reported that in vivo administration of CYP2E1 adenovirus in mice produced significant liver injury compared with LacZ-infected mice as shown by histopathology, markers of oxidative stress, and elevated transaminase levels. 52 In the present study, CYP2E1 expression was significantly increased in groups treated with ZEN compared to the control group. As demonstrated in the present study, the activities of MDA in the serum and the liver increased in ZEN-treated groups, and the activities of SOD and GPx decreased. This suggested that elevated CYP2E1 levels may be a risk factor for xenobiotic metabolism, oxidative stress, and long-term hepatic injury.
UGT isozymes detoxify metabolites, drugs, toxins, and environmental chemicals via conjugation to glucuronic acid. UGT1A1 is one of the most extensively characterized UGT isoforms, and plays an important physiological role in the clearance of bilirubin and xenobiotics. 53,54 In the present study, ZEN had no significant effect on the mRNA levels of UGT1A1. Meissonnier et al. found that both the enzyme activity and the protein expression of the UGT 1A subfamily were unchanged in piglets regardless of the dose of AFB1 in the feed. 55
Bilirubin is the catabolic byproduct of heme proteins, such as β-globin, and CYP enzymes. Accumulation of bilirubin in the blood is potentially hepatotoxic and neurotoxic. In the current study, the activity of TBIL in serum was significantly increased in all ZEN-treated groups compared with the control group. This result is consistent with Abbès et al. who found that an orally administered single dose of ZEN (40 or 500 mg/kg b.w.) significantly increased blood levels of TBIL. 43 Given the same conditions, Kaplan demonstrated that the rise of serum levels of TBIL and BILD was possibly due to its degeneration in the liver, because the activity levels of these enzymes would increase as a result of degeneration in the hepatic tissues and perturbation of biliary system. 56
Conclusion
In conclusion, the results of the current study indicated that ZEN not only caused damage in the liver of pregnant rats in a dose-dependent manner according to both biochemical and antioxidant parameters but also induced the mRNA expression of CYP2E1 in the liver. Thus, treatment with ZEN induced hepatic degeneration.
Footnotes
Acknowledgments
We would like to thank Rui Gao, Zhe Qu, Shaoping Nie, and Min Liu of the Institute of Animal Nutrition at the Northeast Agricultural University for their technical assistance.
Conflict of interest
We declare that we have no financial and personal relationships with other people or organizations that can inappropriately influence our work, there is no professional or other personal interest of any nature or kind in any product, service and/or company that could be construed as influencing the position presented in, or the review of, this manuscript.
Funding
This work was supported by the National Basic Research Program (2012CB124703) and the China Agriculture Research System (CARS-36).