
Abstract
The present study was undertaken to evaluate the possible ameliorating effect of butylated hydroxyl toluene (BHT), associated with ferric nitrilotriacetate (Fe-NTA)-induced oxidative stress and liver injury in mice. The treatment of mice with Fe-NTA alone enhances ornithine decarboxylase activity to 4.6 folds, protein carbonyl formation increased up to 2.9 folds and DNA synthesis expressed in terms of [3H] thymidine incorporation increased to 3.2 folds, and antioxidants and antioxidant enzymes decreased to 1.8–2.5 folds, compared with the corresponding saline-treated controls. These changes were reversed significantly (p < 0.001) in animals receiving a pretreatment of BHT. Our data show that BHT can reciprocate the toxic effects of Fe-NTA and can serve as a potent chemopreventive agent.
Introduction
Oxidative stress results when the balance between the production of reactive oxygen species (ROS) overrides the antioxidant capability of the target cell, resulting in the occurrence of oxidative damage due to the interaction of reactive oxygen with cellular macromolecules. 1,2 ROS are formed in cells during many endogenous and exogenous metabolic processes. ROS may attack and modify membrane lipids, proteins and DNA, giving rise to a large number of altered and damaged products, which may result in the initiation and progression of carcinogenesis by induction of gene mutations, chromosomal damage and cytotoxic effects. 3,4
Ferric nitrilotriacetate (Fe-NTA) is an iron complex of a chelating agent which has earlier been demonstrated to accumulate more redox active iron in hepatocytes and has been shown to be a potent hepatic tumor promoter. Sakurai and Cederbaum 5 have revealed that Fe-NTA causes apoptosis in hepatocytes and promotes liver fibrosis. Fe-NTA is a potent hepatic tumor promoter which acts through the generation of oxidative stress. 6 In addition, Fe-NTA induces oxidative stress by downregulating hepatic NAD(P)H: quinine oxidoreductase activity and produces protein carbonyl contents. 7 Administration of Fe-NTA has also been reported to result in hepatic iron loading associated with extensive peroxidation of membrane lipids in vivo. 8
Diet has been suggested to have a significant impact on the process of the cancer development. 9,10 Past investigations suggest that several dietary constituents may be important factors capable of increasing or decreasing the cancer incidence. Thus, specific and predetermined manipulations of the diet may represent a promising and noninvasive approach to minimize cancer. These studies promoted us to search for a novel and effective agent that may suppress toxicity. Fruits, vitamins and other several herbs with diversified pharmacological properties have been shown to be a rich source of cancer chemopreventive agents. 11,12 Most of these agents intervene at the promotion stage of multistage carcinogenesis. 13
Nevertheless, many such agents having the capability to inhibit carcinogenesis both at the initiation and at the promotion stages possess antioxidant and free radical scavenging properties. 14 The prevention of Fe-NTA-induced hepatotoxicity by gallic acid has also been reported previously. 10 The inhibition of Fe-NTA-mediated carcinogenicity in mice by nordihydroguaiaretic acid (NDGA), a plant lignin, was observed in our earlier studies. 15
When administered prior to, or at the time of carcinogen exposure, the phenolic antioxidant butylated hydroxyl toluene (BHT) was found to be an effective inhibitor of carcinogenesis in several target organs. 16,17 Since liver is the largest gland and a major metabolic organ in the body, it is necessary to understand the effect of BHT on Fe-NTA-induced hepatic damage. This study was designed to provide sufficient amount of data to show the chemopreventive potential of BHT, utilizing a well-established animal model of oxidant-induced hepatic damage. In the present study, the effect of prophylactic treatment of BHT, a known antioxidant on Fe-NTA-induced hepatic oxidative stress, hyperproliferation and injury has been shown.
Materials and methods
Preparation of Fe-NTA
The Fe-NTA solution was prepared by the method given by Awai et al. 18
Animals and treatments
Male Swiss albino mice (4–6 weeks old) weighing 20–25 g, were used throughout this study. Animals were housed in an air-conditioned room and had free access to pellet diet and water ad libitum.
Study design
For various sets of biological studies, different groups of animals were used. A total of 60 mice were used in the study. For studying the effect of BHT on Fe-NTA-mediated induction of oxidative stress in liver, 30 mice were used. Another 30 mice were used for studying the effect of BHT on Fe-NTA-mediated [3H] thymidine incorporation into hepatic and renal DNA.
Effect of BHT on Fe-NTA-mediated generation of hepatic oxidative stress, ornithine decarboxylase activity induction
For studying the effect of BHT on Fe-NTA-mediated generation of hepatic oxidative stress and ornithine decarboxylase (ODC) induction, 30 mice were taken and were divided into five groups of six mice each. Group I received saline and served as negative control. Group II received an oral treatment of only higher dose BHT and also served as control. Group III received only corn oil (vehicle of BHT) for a period of 1 week through the gavage. Groups IV and V received 1 mg and 2 mg/animal/day in 0.2 ml of corn oil of BHT, respectively, daily for 1 week orally. Twenty-four hours after the last treatment of BHT or corn oil, the animals of groups III, IV, and V received an intraperitoneal (i.p.) injection of 9 mg Fe/kg body weight of Fe-NTA.
All the 30 animals were killed 12 h after the treatment of Fe-NTA by cervical dislocation within a short period of 1 h. Just before killing, blood of these animals was collected in different test tubes from retro-orbital sinus for the estimation of serum transaminases, serum glutamate oxaloacetate transaminases (sGOT) and serum glutamate pyruvate transaminases (sGPT), and lactate dehydrogenase (LDH). Liver of these animals were collected and processed for the ODC activity and estimated for antioxidants including glutathione (GSH) and antioxidant enzymes: GSH peroxidase, GSH reductase, GSH S transferase, glucose-6-phosphate dehydrogenase and catalase.
Estimation of serum transaminases and LDH
Estimation of sGOT and sGPT activities
Estimation of sGOT and sGPT activities were determined by Reitman and Frankel colorimetric procedure. 19 The method involves incubation of 0.5 ml of 2 mM α-ketoglutatarate for 5 min at 37°C in a water bath. Then, 0.1 ml of serum was added and the volume was adjusted to 1 ml with sodium phosphate buffer. The reaction mixture was incubated at 37°C for 20 min. The reaction was terminated with an equal volume of trichloroacetic acid (10% w/v), stored in ice and then centrifuged at 4000g for 5 min To the supernatant, 0.5 ml of 2,4 dinitrophenylhydrazine (DNPH) (1 mM) was added. The reaction mixture was left for another 30 min at room temperature. Finally, the color developed by the addition of 5 ml of 0.4 sodium hydroxide and the product was read at 505 nm.
Estimation of LDH activity
The LDH activity was estimated by Wroblewski and La Due method. 20 Briefly, the reaction mixture containing 2.7 ml of phosphate buffer (0.1 ml, pH 7.4), 0.01 ml serum and 0.1 ml of reduced nicotinamide adenine dinucleotide was incubated for 20 min. To this 0.1 ml of pyruvate (2.5 mg/ml) was added. The absorbance was read for 5 min at an interval of 30 sec. The activity was expressed as milli International Unit.
Preparation of postmitochondrial supernatant and microsomes
Livers were quickly removed and perfused immediately with ice-cold saline (0.85% w/v sodium chloride) and homogenized in chilled phosphate buffer (0.1 M, pH 7.4) that contained 1.17% w/v potassium chloride (KCl), using a Potter Elvehjem homogenizer. The homogenate was filtered through a muslin cloth and was centrifuged at 800 g for 5 min at 4°C in refrigerated centrifuge (Eltek, RC 4100 D) to separate the nuclear debris The aliquot obtained was centrifuged at 10,500 g for 20 min at 4°C to obtain postmitochondrial supernatant (PMS), which was used as a source of enzymes. A portion of the PMS was centrifuged in an ultracentrifuge (Beckman, L7-55) at 105,000 g for 60 min at 4°C. This pellet was considered to be the microsomal fraction and was suspended in a phosphate buffer (0.1 M, pH 7.4) containing 1.17% w/v KCl.
Determination of ODC activity
ODC activity was determined by utilizing 0.4 ml of renal and hepatic supernatant fractions per assay tube and measuring the release of 14 CO2 from DL-[1- 14 C] ornithine by the method of Brien et al. 21 and as described by Athar et al. 22 The livers were homogenized in Tris-HCl buffer (pH 7.5, 50 mM) that contained ethylenediaminetetraacetic acid (EDTA) (0.1 mM), pyridoxal phosphate (0.1 mM), phenylmethylsulfonyl fluoride (1.0 mM), 2-mercapto-ethanol (1.0 mM), dithiothreitol (0.1 mM) and Tween 80 (0.1% w/v), at 4°C using a polytron homogenizer (Kinematica, AGPT 3000). ODC activity was expressed as pmol 14 CO2 released/h/mg protein.
Lipid peroxidation
The assay for hepatic microsomal lipid peroxidation was done following the method of Wright et al. 23 The amount of malonaldehyde formed in each of the samples was assessed by measuring the optical density of the supernatant at 535 nm using a spectrophotometer against a blank reagent. The results were expressed as nmol MDA formed/h/g tissue at 37°C using a molar extinction coefficient of 1.563105 M/cm.
Estimation of glutathione
Reduced glutathione (GSH) in liver was assayed by the method of Jollow et al. 24 An aliquot of 1.0 ml of hepatic PMS (10% w/v) was precipitated with 1.0 ml of sulfosalicylic acid (4% w/v). The yellow color developed was read immediately at 412 nm on a spectrophotometer (Milton Roy Model-21D). The amount of GSH was expressed as mmol/g tissue.
Assay of antioxidant enzymes
Glucose 6-phosphate dehydrogenase activity
The activity of glucose 6-phosphate dehydrogenase was assayed by the method of Zaheer et al. 25 The changes in absorbance were recorded at 340 nm and the enzyme activity was calculated as nmol NADP reduced/min/mg protein using a molar extinction coefficient of 6.223103 M/cm.
GSH reductase activity
GSH reductase activity was assayed by the method of Carlberg and Mannervik 26 as modified by Mohandas et al. 27 The assay system consisted of 1.65 ml phosphate buffer (0.1 M, pH 7.6), 0.1 ml EDTA (0.5 mM), 0.05 ml oxidized GSH (1 mM), 0.1 ml NADPH (0.1 mM) and 0.1 ml of hepatic PMS (10% w/v) in a total volume of 2.0 ml. The enzyme activity was quantitated at 25°C by measuring the disappearance of NADPH at 340 nm and was calculated as nmol NADPH oxidized/min/mg protein using a molar extinction coefficient of 6.223103 M/cm.
Catalase activity
Catalase activity was assayed by the method of Claiborne. 28 Briefly, the assay mixture consisted of 1.95 ml phosphate buffer (0.05 M, pH 7.0), 1.0 ml hydrogen peroxide (H2O2; 0.019 M), 0.05 ml and 0.025 ml of hepatic PMS (10% w/v), respectively, in a final volume of 3.0 ml. Changes in absorbance were recorded at 240 nm. Catalase activity was calculated in terms of nmol H2O2 consumed/min/mg of protein.
GSH-S-transferase activity
GSH-S-transferase activity was measured by the method of Habig et al., 29 as described by Athar et al. 30 The reaction mixture consisted of 1.425 ml phosphate buffer (0.1 M, pH 6.5), 0.2 ml reduced GSH (1 mM), 0.025 ml CDNB (1 mM), 0.3 ml and 0.05 ml of hepatic PMS (10% w/v), respectively, in a total volume of 2.0 ml. The changes in absorbance were recorded at 340 nm and enzyme activity was calculated as nmol CDNB conjugate formed/min/mg protein using a molar extinction coefficient of 9.63103 M/cm.
H2O2 assay
H2O2 was assessed by H2O2-mediated horseradish peroxidase–dependent oxidation of phenol red by the method of Pick and Keisari 31 with a slight modification. Hepatic microsomes, 2.0 ml (10% w/v) each, were suspended in 1.0 ml of solution containing phenol red (0.28 nM), horseradish peroxidase (8.5 U), dextrose (5.5 nM) and phosphate buffer (0.05 M, pH 7.0) and were incubated at 37°C for 60 min. The quantity of H2O2 produced was expressed as nmol H2O2/h/g tissue based on the standard curve of H2O2-oxidized phenol red.
Effect of BHT on Fe-NTA-mediated induction of protein carbonyl
Determination of protein carbonyl content
Protein oxidation was measured by an estimation of carbonyl groups performed according to the method described by Levine et al. 32 with slight modifications using 2,4-DNPH reagent. Then, 0.1 ml of serum was treated with 0.5 ml of 10 mM DNPH and dissolved in 2 M HCl as a sample or with 0.5 ml of 2 M HCl as a control blank.
Effect of BHT on Fe-NTA-mediated [3H] thymidine incorporation in hepatic DNA (DNA synthesis)
For studying [3H] thymidine incorporation in hepatic DNA another 30 mice were used, animal treatment protocol and the dose regimen were the same as described above. However, 18 h after the treatment of Fe-NTA, these animals were given an i.p. injection of [3H] thymidine (20 µCi/animal). After 2 h of administration of [3H] thymidine, these animals were killed by cervical dislocation. Liver was quickly removed, cleansed for extraneous material and homogenized in distilled water for their further processing and the separation of DNA. The incorporation of [3H] thymidine in hepatic DNA was done by the method of Smart et al. 33 as described by Iqbal et al. 6 The amount of DNA in the filtrate was estimated by Giles and Myers’ diphenylamine method. 34 The amount of [3H] thymidine incorporated was expressed as d.p.m./mg DNA.
Statistical analysis
Values were expressed as mean ± standard error of the mean. The level of significance between different groups is based on Dunnett’s t test followed by analysis of variance (ANOVA). One-way ANOVA was used to calculate the statistical significance between various groups. A value of p < 0.05 was considered to be statistically significant.
Results
Effect of BHT on Fe-NTA-mediated generation of hepatic oxidative stress and ODC induction
The effect of pretreatment of BHT to mice on Fe-NTA-mediated induction of hepatic ODC activity is shown in Figure 1. The treatment of Fe-NTA alone results in about 4.6-fold increase in hepatic ODC activity as compared to the corresponding saline-treated control. The pretreatment of BHT to mice at dose level of 1 mg/animal/day marginally reduced the Fe-NTA-mediated hepatic ODC induction to about 1.21-fold but at a higher dose of 2 mg/animal/day of BHT ODC activity was reduced to about 1.59-fold compared with Fe-NTA-treated groups. The pretreatment of BHT on the whole reduced the ODC activity up to 64% in liver as compared to corresponding Fe-NTA-treated groups.
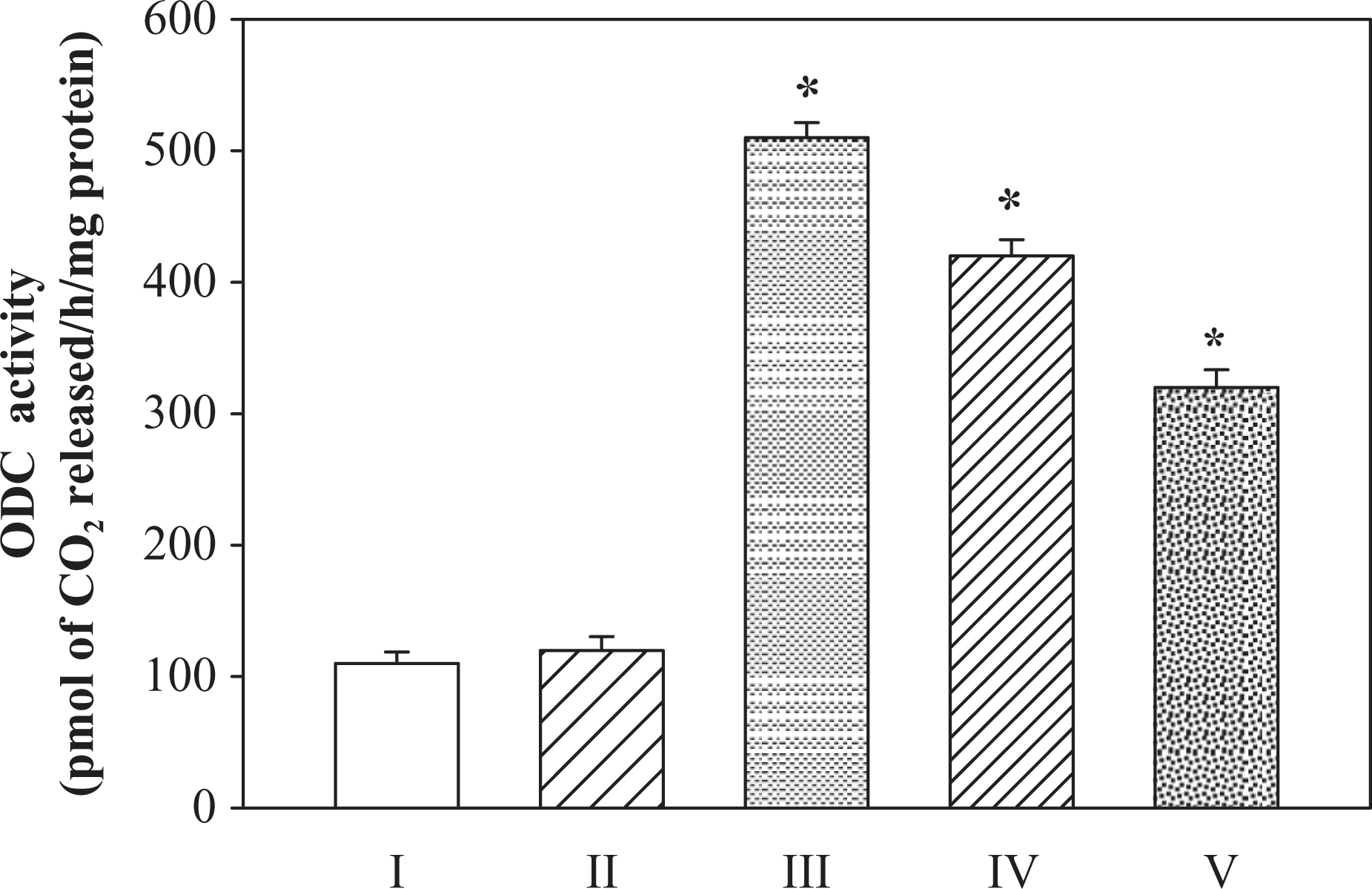
Inhibitory effect of BHT on Fe-NTA-induced hepatic ODC activity. I-saline, II-BHT (D2), III-Fe-NTA, IV-BHT(D1) + Fe-NTA, V-BHT (D2) + Fe-NTA. Data represents mean ± SE of six animals. Dose regimen and treatment protocol are described in the text. *Represents p value < 0.05. ODC: ornithine decarboxylase; BHT: butylated hydroxyl toluene; Fe-NTA: ferric nitrilotriacetate; SE: standard error; D1: dose1; D2: dose2.
Effect of BHT on Fe-NTA-mediated changes in transaminases and LDH
The effect of pretreatment of BHT on Fe-NTA-mediated enhancement in the activities of serum transaminases (sGOT and sGPT) and LDH is shown in Figure 2. Fe-NTA administration led to about 2.2 and 3.2-fold enhancement in the values of both sGOT and sGPT and 3.4-fold increase in LDH activity, respectively. Administration of Fe-NTA to mice receiving a pretreatment of BHT, however, led to a significant diminution in these values. The effect was dependent on the dose of BHT administered; though both doses of BHT used, a significant reduction in the values of sGOT and sGPT and LDH activities could be observed. Higher dose of BHT resulted in bringing down these indices to almost 1.7-, 1.4- and 3.0-fold, respectively, as compared to the Fe-NTA-treated group. There was up to 32–68% decrease in the activities of these enzymes compared to Fe-NTA groups.

Modulatory effect of BHT on Fe-NTA-induced hepatic enhancement in the levels of sGOT, sGPT and LDH. I-Saline, II-BHT (D2), III-Fe-NTA, IV-BHT (D1) + Fe-NTA, V-BHT (D2) + Fe-NTA. Data represents mean ± SE of six animals. Dose regimen and treatment protocol are described in the text. sGOT: serum glutamate oxaloacetate transaminase; sGPT: serum glutamate pyruvate transaminase; LDH: lactate dehydrogenase; BHT: butylated hydroxyl toluene; Fe-NTA: ferric nitrilotriacetate; SE: standard error; D1: dose1; D2: dose2.
Effect of BHT on Fe-NTA-induced changes in antioxidants
The effect of pretreatment of mice with BHT on Fe-NTA-mediated depletion in the level of hepatic GSH and on the activities of various GSH metabolizing enzymes including GSH reductase, GSH-S-transferase, GSH peroxidase, catalase are shown in Table 1. The treatment of Fe-NTA alone results in the depletion of hepatic GSH and decreases the activities of GSH reductase and GSH-S-transferase to the level of 1.9-, 2.5- and 2.1-fold in liver compared with the corresponding saline-treated controls. Pretreatment with BHT resulted in the significant recovery of these enzymes. At a higher dose of 2 mg/animal/day of BHT, there was significant increase (p < 0.001) in the levels of GSH (1.53 fold) compared with Fe-NTA group. GSH reductase, GSH-S-transferase activity was significantly increased to 1.92- and 2.08-fold, respectively.
Inhibitory effects of BHT on Fe-NTA-mediated hepatic levels of GSH, the activity of GSH-metabolizing enzymes and antioxidant enzymes.a,b
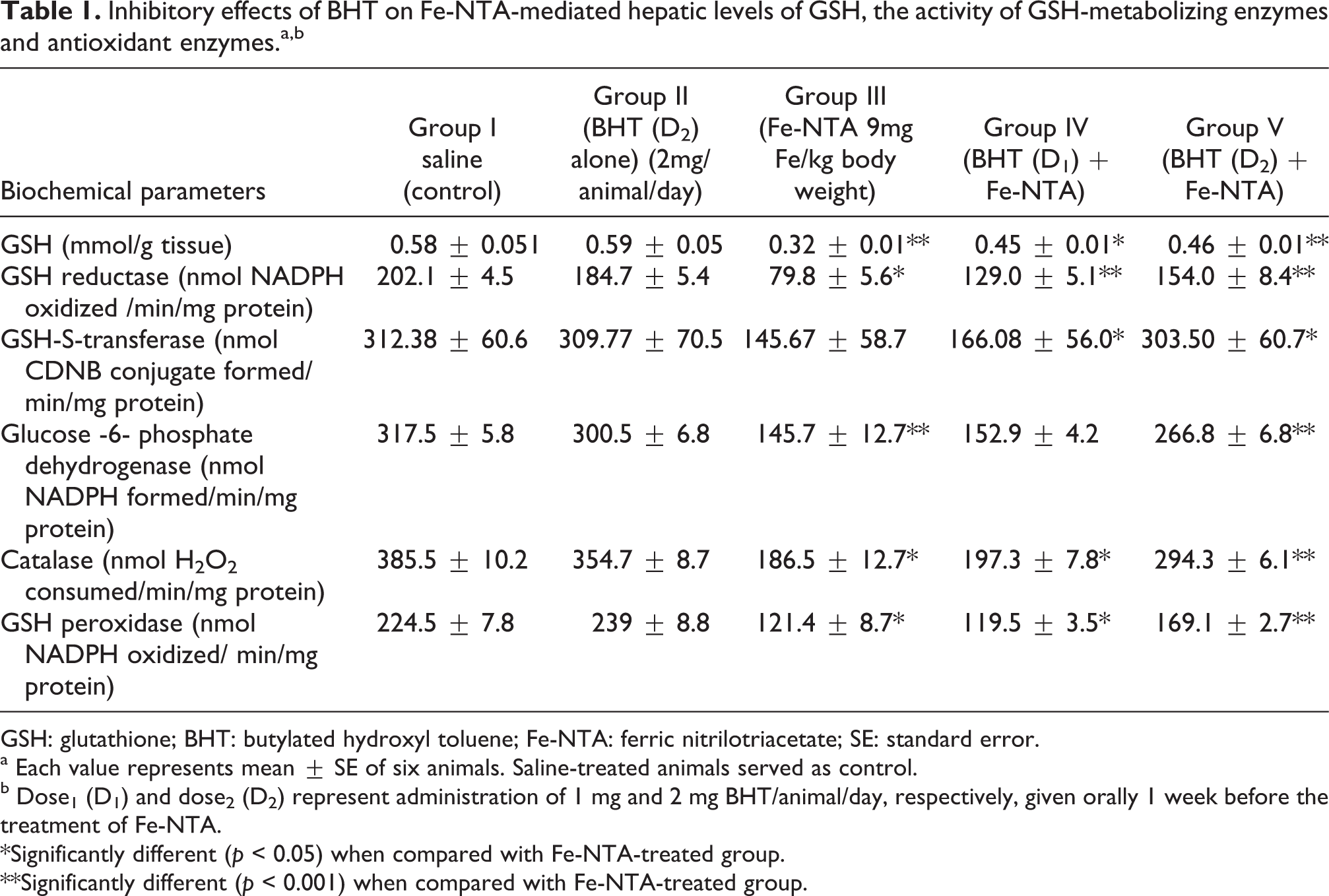
GSH: glutathione; BHT: butylated hydroxyl toluene; Fe-NTA: ferric nitrilotriacetate; SE: standard error.
a Each value represents mean ± SE of six animals. Saline-treated animals served as control.
b Dose1 (D1) and dose2 (D2) represent administration of 1 mg and 2 mg BHT/animal/day, respectively, given orally 1 week before the treatment of Fe-NTA.
*Significantly different (p < 0.05) when compared with Fe-NTA-treated group.
**Significantly different (p < 0.001) when compared with Fe-NTA-treated group.
The Fe-NTA treatment alone results in significant decrease (p < 0.05) in the activities of other hepatic antioxidants enzymes including glucose 6-phosphate dehydrogenase, catalase and GSH peroxidase to about 2.1-, 2.0- and 1.8-fold, respectively, in the liver of the corresponding saline-treated controls. Pretreatment of mice with BHT resulted in the significant recovery of these enzymes. There was significant increase to1.83-, 1.57-, 1.39-fold, respectively, in the activities of these enzymes at the higher dose of BHT compared with Fe-NTA-treated group. Upon pretreatment with a high dose of BHT, the levels of antioxidants were increased to 51–97% in liver as compared to Fe-NTA-treated group.
Effect of BHT on Fe-NTA-induced lipid peroxidation
The effect of pretreatment of mice with BHT on Fe-NTA-mediated enhancement of hepatic microsomal lipid peroxidation and generation of H2O2 is shown in Figure 3. The treatment of Fe-NTA alone enhanced the hepatic microsomal lipid peroxidation to about 1.9-fold of the corresponding saline-treated control; whereas there was an enhancement of H2O2 generation more than 2.2-fold. However, the pretreatment with BHT inhibited the dose dependency on Fe-NTA-mediated enhancement of hepatic microsomal lipid peroxidation as well as H2O2 generation. Although, there was a significant reduction in oxidant generation on both the doses of BHT studied, but at the higher dose of BHT, the level of hepatic microsomal lipid peroxidation reduced to about 1.5-fold, respectively, as compared with the corresponding Fe-NTA-treated group. This dose also induced about 1.5-folds reduction in the H2O2 generation in liver as compared with the corresponding Fe-NTA-treated groups. Fe-NTA-mediated enhancement in lipid peroxidation was reduced up to 68% in liver by treatment with BHT.

Modulatory effect of BHT on Fe-NTA-induced hepatic enhancement in the level of LPO and H2O2 generation. I-Saline, II-BHT (D2), III-Fe-NTA, IV-BHT (D1) + Fe-NTA, V-BHT (D2) + Fe-NTA. Data represents mean ± SE of six animals. Dose regimen and treatment protocol are described in the text. *Represents p value < 0.05, **Represents p value < 0.001. LPO: lipid peroxidation; H2O2: hydrogen peroxide; D1: dose1; D2: dose2.
Effect of pretreatment of BHT to mice on Fe-NTA-mediated enhancement of [ 3 H] thymidine incorporation
The effect of pretreatment of BHT to mice on Fe-NTA-mediated enhancement of [ 3 H] thymidine incorporation in hepatic DNA is shown in Figure 4. The treatment of Fe-NTA alone resulted in about 3.2-fold enhancement in the incorporation of [ 3 H] thymidine in hepatic DNA, when compared with their corresponding saline-treated controls. However, in BHT-pretreated animals, this enhancement was significantly less as compared with Fe-NTA alone treatment group. The decrease in the [ 3 H] thymidine incorporation was dependent on the dose of BHT used, as observed with the ODC induction studies. By providing a higher dose of 2 mg/animal/day of BHT, DNA synthesis was reduced to about 2.8-fold compared with Fe-NTA-treated groups. Pretreatment of BHT reduced the [ 3 H] thymidine incorporation up to 35% in liver as compared with the corresponding Fe-NTA-treated group.

Inhibitory effect of BHT on Fe-NTA-induced hepatic DNA synthesis. I-saline, II-BHT(D2), III-Fe-NTA, IV-BHT(D1) + Fe-NTA, V-BHT(D2) + Fe-NTA. Data represents mean ± SE of six animals. Dose regimen and treatment protocol are described in the text. *Represents p value < 0.05, **Represents p value < 0.001. BHT: butylated hydroxyl toluene; Fe-NTA: ferric nitrilotriacetate; SE: standard error; D1: dose1; D2: dose2.
Effect of pretreatment of mice with BHT on Fe-NTA-mediated induction of protein carbonyl
The effect of pretreatment of mice with BHT on Fe-NTA-mediated increase in 2,4-DNPH incorporation into cytosolic proteins representing protein carbonyl formation is shown in Table 2. Fe-NTA alone treatment resulted in a significant increase of up to 2.9-fold in the 2,4-DNPH incorporation into cytosolic proteins in liver of mice (p < 0.05). However, in BHT pretreated animals, this increase in the incorporation of 2,4-DNPH was lesser as compared to Fe-NTA alone-treated group. The decrease in the incorporation of 2,4-DNPH into cytosolic protein was dependent on the dose of BHT used. At the higher dose of BHT of 2 mg/animal/day, the decrease in the incorporation of 2,4-DNPH into cytosolic protein was about 2.3-fold in the liver of mice as compared with Fe-NTA-treated group. Pretreatment of higher dose of BHT resulted in 44% decrease in the level of protein carbonyl compared with the corresponding Fe-NTA-treated groups. Fe-NTA-mediated induction of protein carbonyl was reduced up to 44% in liver.
Inhibitory effects of BHT on Fe-NTA-mediated enhancement of protein carbonyl.a,b

BHT: butylated hydroxyl toluene; Fe-NTA: ferric nitrilotriacetate; SE: standard error.
a Each value represents mean ± SE of six animals. Saline-treated animals served as control.
b Dose1(D1) and dose2(D2) represent administration of 1 mg and 2 mg BHT/animal/day, respectively, given orally 1 week before the treatment of Fe-NTA.
*Significantly different (p < 0.05) when compared with Fe-NTA-treated group.
Discussion
Fe-NTA is a known renal carcinogen. However, little is known about its carcinogenic potential in liver. Considerable progress has been made relating free radicals, especially ROS, in living cells to pathogenicity of various diseases including diabetes mellitus, atherosclerosis, Alzheimer’s disease, Parkinson’s disease, aging and carcinogenesis. 35 Humans could be significantly affected if exposed to NTA in drinking water or to its metal complexes. Dietary intake of natural antioxidants, therefore, has been suggested to be useful against the effects of these mutagenic and carcinogenic agents in minimal amounts that are not toxic to the body.
A large number of agents including natural and synthetic compounds have been identified as having potential cancer chemopreventive value inhibiting mutagenesis hyperproliferation or induce apoptosis or differentiations, which are the critical characteristics of chemoprevention. 36 Identification of naturally occurring carcinogens and anticarcinogens provides new strategies for cancer prevention.
One of the known phenolic antioxidants, BHT, has been commonly used in foods, cosmetics and pharmaceutical products to prevent oxidation of liable lipid compounds, and it also modifies certain enzyme system. 16 Similar to previous reports which have shown protective effect of BHT in various animal models, a dose-dependent decrease in the Fe-NTA-mediated oxidative stress in liver of BHT-pretreated mice was also observed in this study. Our dose selection for BHT in minimum amounts does not show any toxic effect in mice and also suggests that these doses may not be toxic to human consumption.
In the present study, we observed that Fe-NTA treatment enhances ODC activity, DNA synthesis, inhibits GSH, activities of GSH-metabolizing enzymes, transaminases (sGOT and sGPT) and antioxidant enzymes in liver. In addition to these, Fe-NTA treatment induces hepatic microsomal lipid peroxidation and generation of H2O2.
Our data shows that pretreatment of animals with two different doses of BHT suppressed Fe-NTA-mediated induction of ODC activity and DNA synthesis, reversed the decrease in the activities of GSH-metabolizing enzymes, antioxidant enzymes and depleted the level of reduced GSH in liver. Also, there was decrease in the levels of protein carbonyl formation and lipid peroxidation in a group of mice pretreated with BHT. The inhibitory effect of BHT against Fe-NTA-mediated hepatic damage as obser-ved in the present study may be due to the counteracting effect of its antioxidant property, which may be related to its ability to prevent the in vivo activation of carcinogens to reactive forms and increasing detoxification of reactive intermediate by inducing relevant enzymes such as GSH-S-transferase, GSH peroxidase, GSH reductase, glucose 6-phosphate dehydrogenase, catalase and so on.
The exact mechanism by which BHT suppresses Fe-NTA-mediated hepatic toxicity and hyperproliferation is not known. In the present study, higher activities of sGOT and sGPT have been found in response to iron-induced oxidative stress. Such increased activities might be attributed to the leakage of these enzymes from the injured liver cells into the blood stream because of the altered liver membrane permeability.
ODC activity and [3H] thymidine incorporation are widely used as biochemical markers to evaluate tumor-promoting potential of an agent. 21 Although ODC and polyamines are necessary for cellular proliferation, induction of ODC is involved in tumor promotion and cell transformation, and cultured tumor cells often contain high levels of ODC. Therefore, ODC is considered as an attractive target in both chemotherapy and chemoprevention. It has also been reported that induction of ODC activity and DNA synthesis plays an important role in carcinogenesis. 37 A sharp decrease in Fe-NTA-mediated enhancement in hepatic protein carbonyl, ODC activity and [3H] thymidine incorporation by the pretreatment of BHT also suggests the protective role of BHT against Fe-NTA-induced hepatotoxicity in mice.
Conclusion
The results presented in this study clearly indicate that pretreatment of animals with BHT ameliorates Fe-NTA-mediated hepatic toxicity and thereby suppresses hyperproliferation. The major pathway for its action and efficacy seems to be its effectiveness to intercept the oxyradical and protect cellular membrane damage against the deleterious effect of ROS. In summary, present data suggest that BHT may be an effective chemopreventive agent and may offer protection against Fe-NTA-mediated hepatic damage in mice.
Footnotes
Conflict of Interest
The authors declared no conflict of interest.
Funding
This research project was supported by a grant from the “Research Center of the Center for Female Scientific and Medical Colleges,” Deanship of Scientific Research, King Saud University.