
Abstract
Introduction
Several approaches have been used to prevent the unwanted and harmful physiological effects of toxicants like NO. Much attention in this regard has been paid to the protective effects of natural products such as oils rich in ω-3 fatty acids. A number of investigations have demonstrated that diet supplemented with fish oil (FO) enriched in ω-3 fatty acids has profound beneficial health effects against various pathologies, 16 including cardiovascular diseases (CVD), respiratory diseases, diabetes, depression, cancers, inflammatory and immune renal disorders. 17 Preliminary reports indicate that FO prevents nephrotoxicity. 18 Not lacking far behind, plant sources of polyunsaturated fatty acid (PUFA), such as flaxseed oil (FXO; >50% ALA), have shown great promise in fighting against many diseased conditions. Dietary supplementation of FXO is being used to reduce hyperlipidemia. 19 It is also being used as a cardioprotective agent. 20 FXO has been recently linked with bone development in experimental animals. 21
Considering the potential clinical benefits of FO and FXO, the present study was undertaken to examine the effect of prior administration of FO and FXO on the ARG-induced changes on the enzymes of renal brush border membrane (BBM), carbohydrate metabolism, phosphate transport and antioxidant parameters. We hypothesize that FO and FXO would prevent ARG-induced nephrotoxicity due to their intrinsic biochemical and antioxidant properties that would lead to improved metabolism and antioxidant defense mechanism in the kidney. The results obtained indicate that the dietary supplementation with FO and FXO markedly enhanced/ameliorated ARG-induced nephrotoxicity parameters. The activities of enzymes of carbohydrate metabolism, BBM and phosphate transport in renal cortical BBM were significantly enhanced by feeding FO and FXO to ARG-administered rats. Furthermore, FXO showed similar protective effects as of FO and thus can be used as a replacement of FO for people with strict vegetarian habits.
Materials and methods
Chemicals and drugs
The following chemicals were used in this study: FO (Menhaden, Sigma Chemical Co., St Louis, MO, USA), FXO (Canada), sodium nitrite (Sigma Chemical Co., USA) and Gries reagent (Sigma Chemical Co., USA). All other chemicals used were of analytical grade and were purchased either from Sigma Chemical Co. (USA) or Sisco Reseach Laboratory (Mumbai, India).
Diet
A nutritionally adequate laboratory pellet diet was obtained from Ashirwad Industries (Chandigarh, India). Pellets were finely crushed and mixed with 15% FO and 15% FXO and stored in airtight containers. Vitamin E as
Experimental design
The animal experiments were conducted according to the guidelines of Committee for Purpose of Control and Supervision of Experiments on Animals (CPCSEA), Ministry of Environment and Forests, Government of India. Adult male Wistar rats (eight rats per group) weighing 150–180 g fed with standard rat chow (Ashirwad Industries) and water ad libitum were conditioned for 1 week before the start of the experiment. Rats were divided into four groups. They were fed on a normal diet (control and ARG) or diet containing 15% FO (ARG + FO) or 15% FXO (ARG + FXO). After 15 days, ARG was administered intraperitoneally (500 mg/kg bw/day) for 10 days to all the groups except control. The control animals received an equivalent volume of normal saline. The rats were killed 24 h after the last injection under light ether anesthesia. Urine samples were collected for 4 h in the standard metabolic cages a day before killing (Figure 1). Blood samples were collected and the kidneys were harvested and processed for the preparation of homogenates and BBM vesicles (BBMVs) as described below.

Experimental design. ND: normal diet; FO: fish oil; FXO: flaxseed oil; ARG:
Serum/urine chemistries
Serum/urine parameters
Serum samples were deproteinated with 3% trichloroacetic acid in a ratio of 1:3, left for 10 min and then centrifuged at 2000g for 10 min. The protein-free supernatant was used to determine inorganic phosphate ( 32 Pi) and creatinine. The precipitate was used to quantitate total phospholipids. Levels of blood urea nitrogen (BUN) and cholesterol were determined directly in serum samples. Glucose was estimated by o-toluidine method using a kit from Span diagnostics (Mumbai, India). These parameters were determined by standard procedures as mentioned in a previous study. 22
Preparation of homogenates
The kidneys were decapsulated and kept in ice-cold 154 mM NaCl and 5 mM Tris-HEPES (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid) buffer, pH 7.5. The cortical and medullary regions were carefully separated and homogenized (as mentioned above) in 50 mM mannitol buffer to get 10% (w/v) homogenate.
One part of the homogenate was centrifuged at 2000g for 10 min at 4°C and the supernatant was saved for assaying the enzymes of carbohydrate metabolism, while the second part was centrifuged at 3000g for 15 min at 4°C and the supernatant was used for the assay of free-radical scavenging enzymes and the third part was used for the estimation of total thiol (–SH) and lipid peroxidation (LPO).
Preparation of BBM
Kidney BBM was prepared from the whole cortex homogenate using the MgCl2 precipitation method as described by Yusufi and Dousa. 23 The final membrane preparations were suspended in 300 mM mannitol, pH 7.4. Freshly prepared BBMVs were used for the transport of 32 Pi and some portion was saved and stored at −20°C until further analysis for BBM enzymes. Each sample of BBM was prepared by pooling the tissues from two to three rats.
Assay of carbohydrate metabolism enzymes
The activities of the enzymes involving oxidation of NADH or reduction of NADP were determined spectrophotometrically on Cintra 5 fixed at 340 nm using 3 ml of assay in a 1-cm cuvette at room temperature (28–30°C). The enzyme assays of lactate dehydrogenase (LDH, E.C.1.1.1.27), malate dehydrogenase (MDH, E.C.1.1.1.37), malic enzyme (ME, E.C.1.1.1.40), glucose-6-phosphate dehydrogenase (G6PDH, E.C.1.1.1.49), glucose-6-phosphatase (G6Pase, E.C.3.1.3.3) and fructose-1,6-bisphosphatase (FBPase, E.C.3.1.3.11) activities were studied as described by Khundmiri et al. 22 Hexokinase (HK) was estimated by the method of Crane and Sols and the remaining glucose was measured by the method of Nelson. 24,25
Assay of BBM marker enzymes and lysosomal marker enzymes
The activities of alkaline phosphatase (ALP), leucine amino peptidase (LAP), γ-glutamyl transferase (GGTase), sucrase and acid phosphatase (ACPase) were determined as described by Farooq et al. 26
Assay of enzymes involved in free radical scavenging
Superoxide dismutase (SOD, E.C.1.15.1.1) was assayed by the method of Marklund and Marklund. 27 Catalase (CAT, E.C.1.11.1. 6) activity was assayed by the method of Giri et al. 28
LPO and total –SH group estimation
Total SH groups were determined by the method of Sedlak and Lindsay. 29 LPO were determined by the method of Ohkawa et al. 30
Transport of 32 Pi
Measurement of 32 Pi (Bhabha Atomic Research Center, India) uptake in BBMVs was carried out at 25°C by rapid filtration technique as described by Yusufi et al. either in the presence or absence of Na gradient. 31 Uptake was initiated by the addition of 30 µl of incubation medium containing 100 mmol/l mannitol, 100 mmol/l NaCl/KCl, 5 mmol/l Tris-HEPES, pH 7.5, 0.1 mmol/l K2 H 32 PO4 to 15 µl BBM suspension (50–100 µg protein) and incubated for the desired time intervals (see Results section). The uptake was stopped by the addition of 3 ml ice-cold stop solution (containing 135 mmol/l NaCl, 5 mmol/l Tris-HEPES and 10 mM sodium arsenate, pH 7.5) and filtered immediately through 0.45 µm MF-Millipore Membrane, mixed cellulose esters, Hydrophilic (DAWP) Millipore (USA) and washed three times with the stop solution using a Cornwall type syringe (Wheaton, Illinois, USA). Correction for nonspecific binding to filters was made by subtracting from all data the value of corresponding blank obtained by filtration of the incubation buffer without vesicles. The radioactivity of dried filters was measured by liquid scintillation counting (Rackbeta, LKB, Wallac, Sweden) with 10 ml scintillation fluid (Cocktail T, SRL, India).
Determination of tissue nitrite
Levels of tissue nitrite were quantified using the Griess assay. 32 Kidney homogenates were centrifuged at 5000g for 10 min. Supernatant (500 µl) was removed and an equal volume of modified Griess reagent was added and the samples were incubated in the dark for 20 min. Standard curves were generated using 0–20 µM sodium nitrite. Absorbance was measured in standards and samples at 550 nm. Concentrations of nitrite were determined from the standard curve, which was linear over the entire concentration range tested. Values were expressed as nanomols per gram tissue.
Statistical analyses
All the data are expressed as mean ± SEM for at least four to five different preparations. Statistical evaluation was conducted by one-way analysis of variance. A probability level of p < 0.05 was selected as indicating statistical significance. Most of the changes between various groups were compared with the control values for better understanding and clarity.
Results
The present work was undertaken to study the detailed mechanism of ARG-induced nephrotoxicty and other deleterious effects and its possible protection by feeding ω-3 fatty acid–enriched oil diets to the rats. To address our hypothesis, the effect of ARG alone and in combination with FO or FXO was determined on nephrotoxicity parameters with serum biomarker enzymes of oxidative stress, BBM and carbohydrate metabolism in the rat kidney tissues. Na-dependent phosphate transport was also determined in renal cortical BBM to assess the role of kidney in the conservation of Pi transport under different experimental conditions.
Effect of dietary FO and FXO on ARG-induced nephrotoxicity parameters in the serum and urine
In general, the rats remained clinically well throughout the study. There was no significant difference in daily food intake and body weights between control and other experimental rats (data not shown). ARG treatment to control rats resulted in significant increase in serum creatinine (Scr), BUN, cholesterol, glucose and phospholipids but decrease in Pi compared with control rats (Table 1). These changes were associated with profound phosphaturia, proteinuria and glucosuria (Table 2) accompanied by decrease in creatinine clearance (Table 2). Feeding of FO or FXO diet to ARG-administered (FO + ARG and FXO + ARG) rats resulted in significant reversal of various ARG elicited deleterious effects on serum and urine parameters. Both FO and FXO diets prevented ARG-induced increase in Scr, BUN, glucose and cholesterol and decrease in serum Pi (Table 1). FO/FXO diet appears to greatly improve renal functions in a similar manner as evident by the increase in creatinine clearance and decrease in the excretion of protein, phosphate and glucose in the urine (Table 2).
Effect of FO and FXO on serum parameters with ARG treatmenta

FO: fish oil; FXO: flaxseed oil; ARG:
a Results (specific activity expressed as micromoles per milligram protein per hour) are mean ± SEM for five different preparations.
b Significantly different from control.
c Significantly different from ARG at p < 0.05 by one-way analysis of variance. Values in parentheses represent percentage change from control.
Effect of FO and FXO on urine parameters of rats with ARG treatmenta
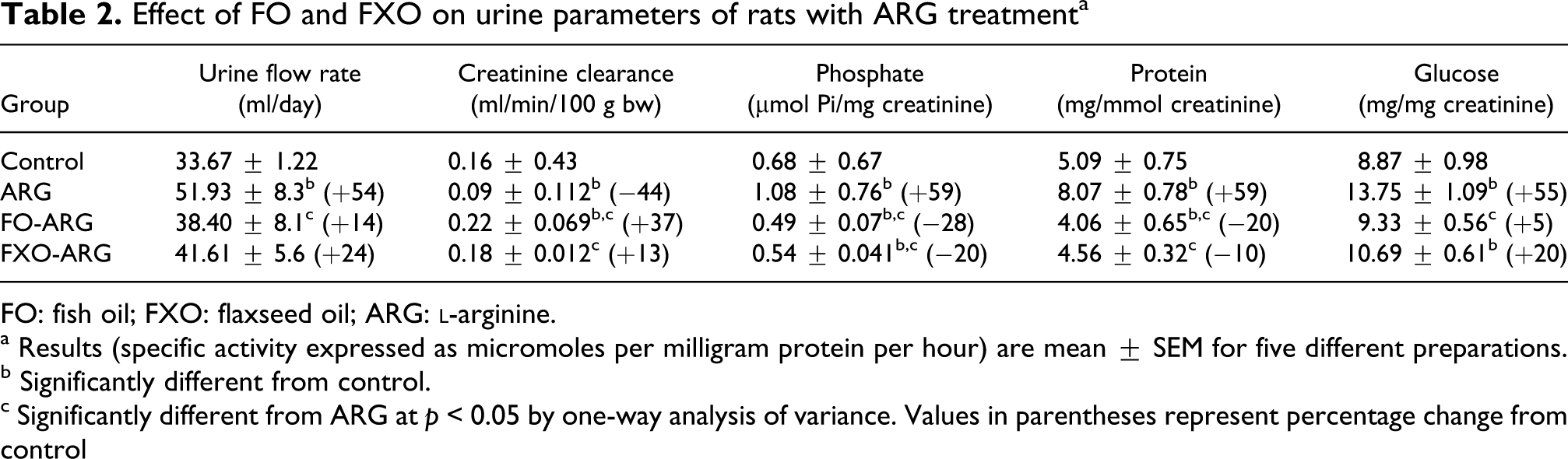
FO: fish oil; FXO: flaxseed oil; ARG:
a Results (specific activity expressed as micromoles per milligram protein per hour) are mean ± SEM for five different preparations.
b Significantly different from control.
c Significantly different from ARG at p < 0.05 by one-way analysis of variance. Values in parentheses represent percentage change from control
Effect of FO and FXO on ARG-induced alterations on metabolic enzymes in renal cortex and medulla
The effect of ARG, FO/FXO diets and their combined treatment was determined on the activities of various enzymes of carbohydrate metabolism in renal cortex and medulla.
Effect on carbohydrate metabolism in renal cortex
As shown in Tables 3 and 4, ARG treatment to control rats significantly increased the activity of LDH but decreased MDH, HK, G6Pase and FBPase activities in the renal cortex (Table 3). When ARG treatment was extended to FO and FXO-fed rats, ARG-induced alterations in metabolic enzyme activities were not only prevented by FO and FXO diets, but G6Pase remained significantly higher in FO- and FXO-ARG compared with control as well as ARG rats in the renal cortex.
Effect of FO and FXO on the activities of LDH, MDH, G6Pase, and FBPase in homogenates of cortex and medulla with ARG treatmenta
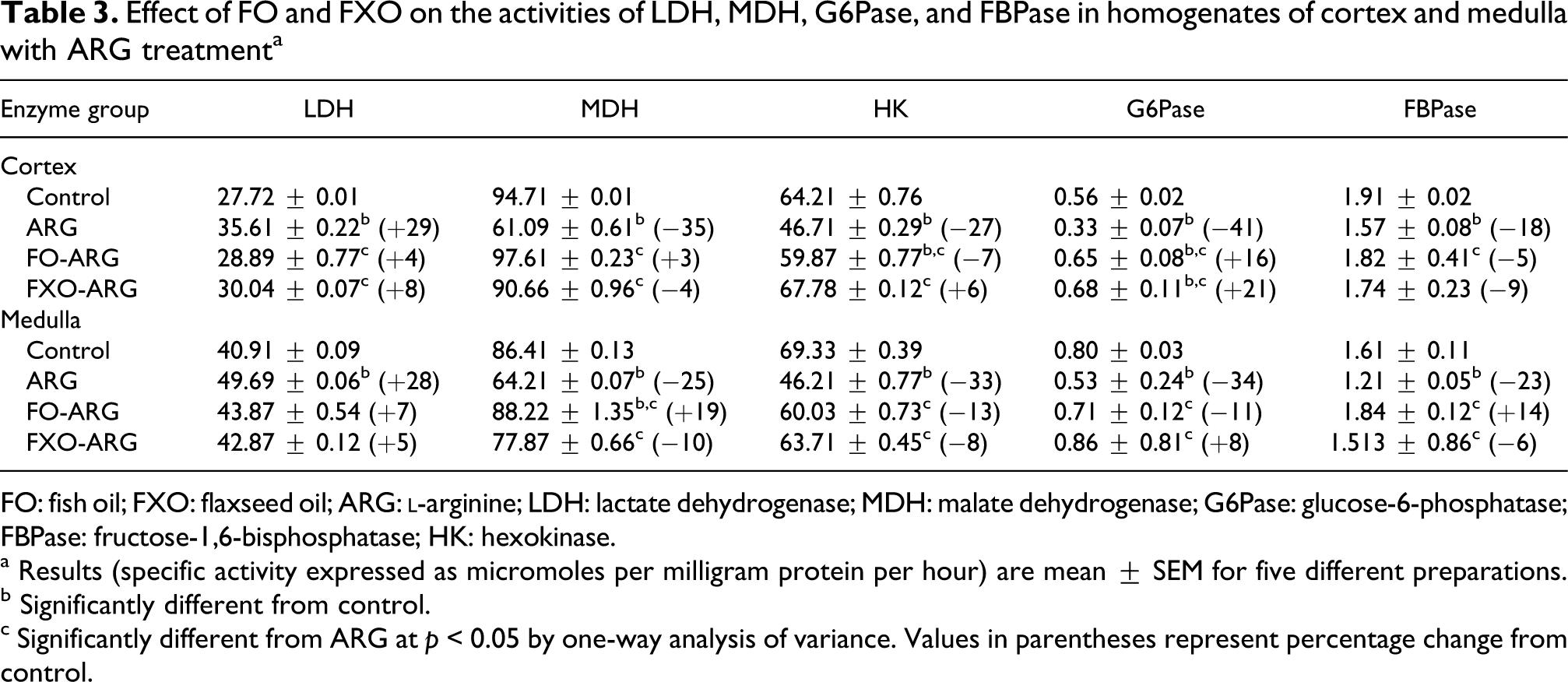
FO: fish oil; FXO: flaxseed oil; ARG:
a Results (specific activity expressed as micromoles per milligram protein per hour) are mean ± SEM for five different preparations.
b Significantly different from control.
c Significantly different from ARG at p < 0.05 by one-way analysis of variance. Values in parentheses represent percentage change from control.
Effect of FO and FXO on the activities of G6PDH, ME, and ACPase in the homogenates of cortex and medulla with ARG treatmenta
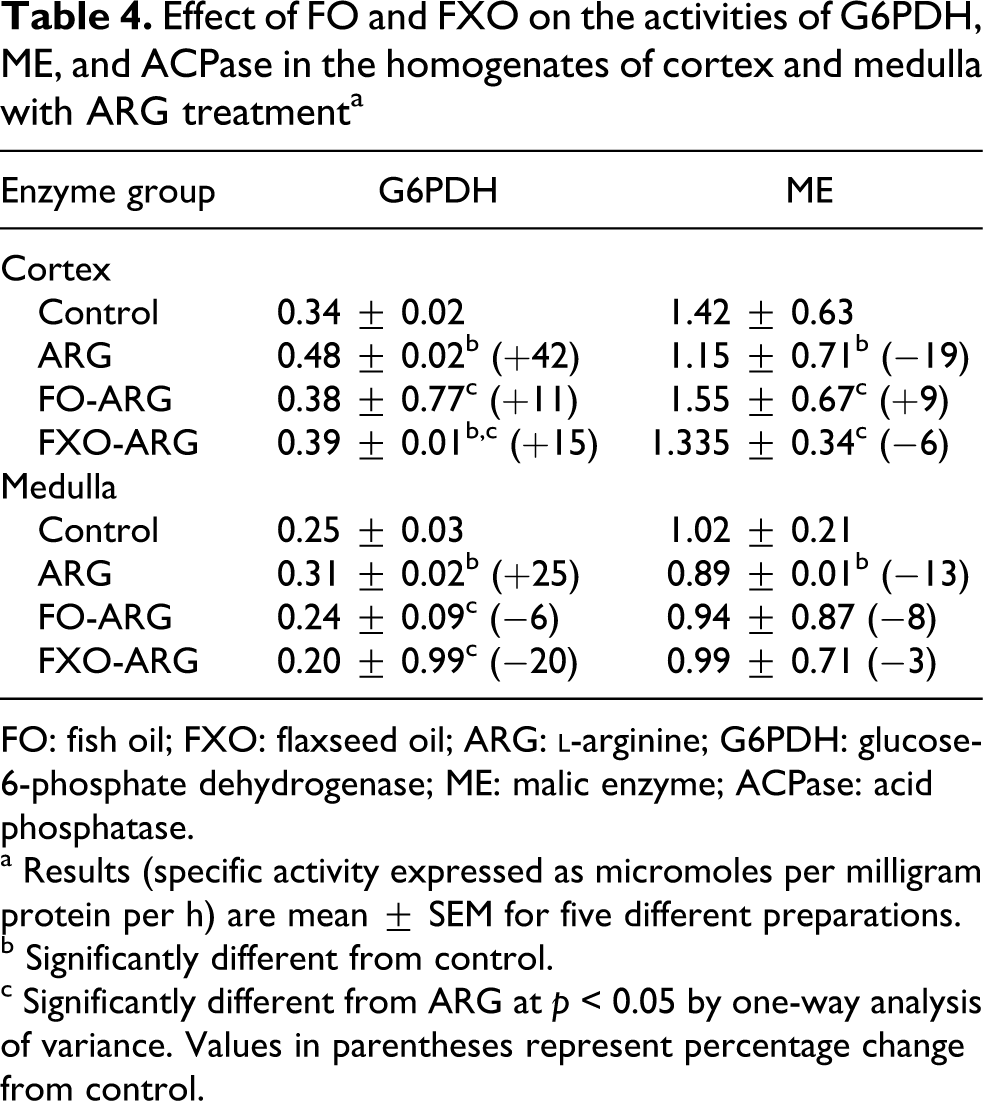
FO: fish oil; FXO: flaxseed oil; ARG:
a Results (specific activity expressed as micromoles per milligram protein per h) are mean ± SEM for five different preparations.
b Significantly different from control.
c Significantly different from ARG at p < 0.05 by one-way analysis of variance. Values in parentheses represent percentage change from control.
The effect of FO, FXO and ARG combined was also determined on G6PDH and NADP-ME, source of NADPH production needed in various anabolic reactions (Table 4). ARG treatment to control rats significantly increased G6PDH activity but decreased ME activity. ARG-elicited increase in G6PDH activity was normalized to near control values by both FO and FXO diets. ARG-induced ME activity decrease was arrested by both FO and FXO diet.
Effect on carbohydrate metabolism in renal medulla
The activities of LDH, MDH, HK, G6Pase and FBPase in medullary homogenates were similarly affected by ARG treatment (Table 3) as in the cortical homogenates. Feeding of FO and FXO to ARG-treated rats prevented ARG-induced alterations in various enzyme activities. Similar to cortex, G6PDH activity increased, whereas ME activity decreased by ARG in the medulla (Table 4). FO and FXO diet to ARG-treated rats decreased G6PDH activity and increased ME activity.
Effect of dietary FO and FXO on ARG-induced alterations in biomarker enzymes of BBM and lysosomes
To assess the structural integrity of certain organelles, for example, plasma membrane (BBM) and lysosomes, the effect of ARG alone and in combination with FO or FXO diets was determined on biomarker enzymes of BBM and lysosomes in the homogenates of renal cortex and medulla and isolated BBM preparations from renal cortex.
Effect of ARG alone and with FO or FXO diet on the biomarkers of BBM and lysosomes in the homogenates
The activities of ALP, γ-glutamyl transferase (GGTase), LAP and ACPase were determined under different experimental conditions in the homogenates of renal cortex and medulla (Table 5). ARG treatment to control rats caused significant reduction in the specific activities of ALP (−35%), GGTase (−57%) and LAP (−52%) in cortical homogenate. The prior feeding of FO or FXO diet with ARG treatment prevented ARG-elicited decrease in BBM enzyme activities. As can be seen from the data, ARG-induced decrease in BBM enzyme activities was similarly prevented by FO or FXO diet. The activity of ACPase was also decreased (−45%) by ARG in the cortical homogenates, and FO/FXO diet was able to prevent the decrease in the enzyme activity in a similar manner (Table 5).
Effect of FO and FXO on the biomarkers of BBM and lysosomes in homogenates of cortex and medulla with ARG treatmenta

FO: fish oil; FXO: flaxseed oil; ARG:
a Results (specific activity expressed as micromoles per milligram protein per hour) are mean ± SEM for five different preparations.
b Significantly different from control.
The activities of BBM enzymes similar to the cortex were also lowered in the medulla by ARG administration although to a lower extent (Table 5). The consumption of FO/FXO in combination with ARG treatment resulted in the reversal of ARG-induced decrease in ALP (−19%), GGTase (−33%) and LAP (−42%) in the medulla. The activity of ACPase was not affected by ARG and ARG + FO; however, it was found to be lower in ARG + FXO rats (−18%) compared with the control rats.
Effects of ARG, ARG + FO and FXO diet on BBM markers in isolated BBMV
The effects of ARG, FO and FXO on BBM marker enzymes were further analyzed in BBMV preparations isolated from the renal cortex (Figure 2). The data show a similar activity pattern of BBM enzymes as observed in cortical homogenates. However, the magnitude of the effects was much more pronounced in BBMV than in cortical homogenates. Activities of ALP (−67%), GGTase (−53%) and LAP (−51%) have profoundly declined by ARG treatment when compared with control rats. FO and FXO dietary supplementation similar to the effect in the homogenates appeared to lower the severity of the ARG treatment. ARG-induced decrease in BBM enzyme activities was significantly prevented by dietary FO and to greater extent by FXO diet.
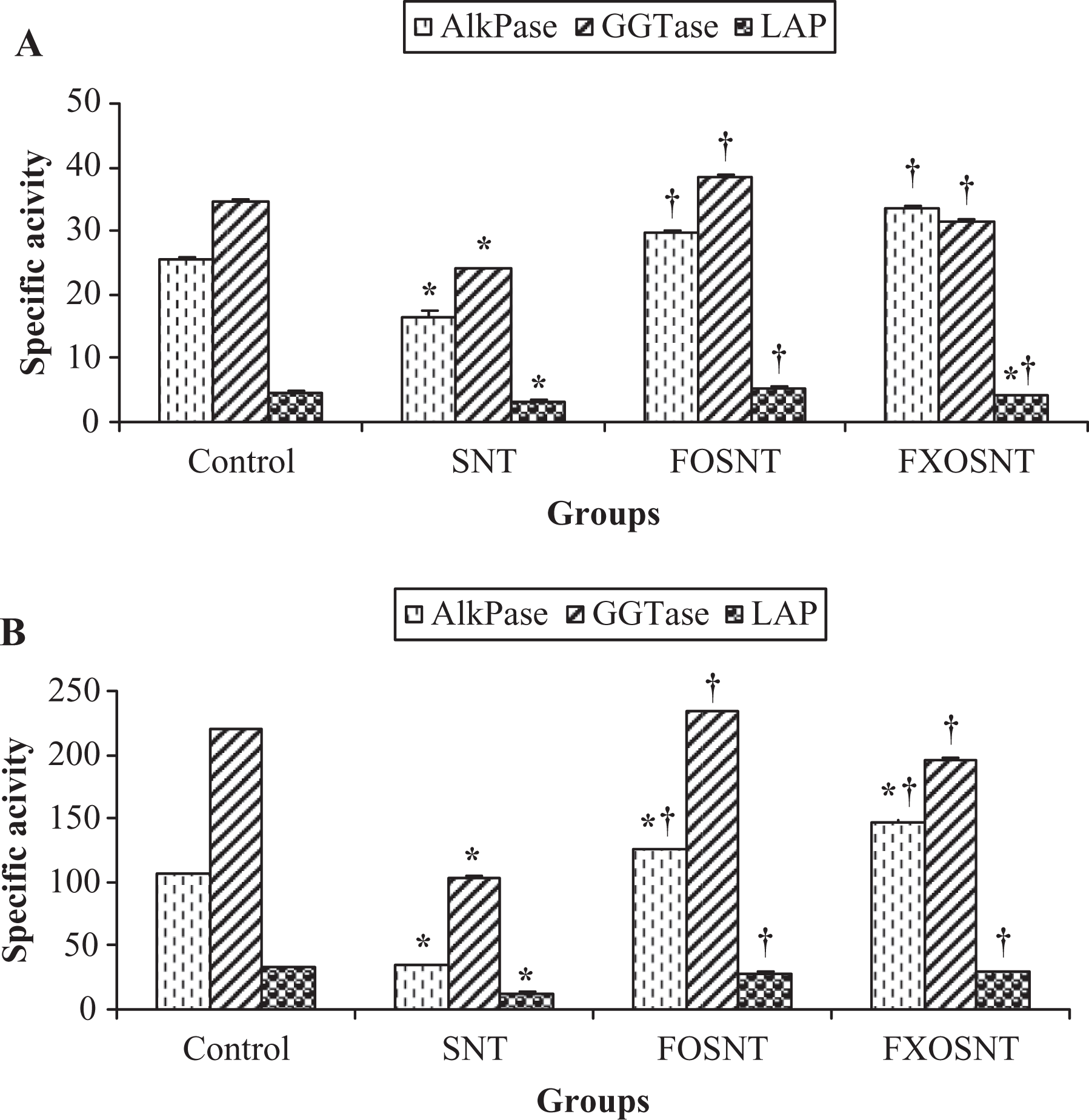
Effect of FO and FXO on the activities of ALP, GGTase, and LAP in (a) cortical homogenates and (b) BBM with ARG treatment. Results (micromoles per milligram protein per hour) are mean ± SEM for five different preparations. *Significantly different from control, †significantly different from ARG at p < 0.05 by one-way analysis of variance. FO: fish oil; FXO: flaxseed oil; ALP: alkaline phosphatase; GGTase: γ-glutamyl transferase; LAP: leucine amino peptidase; BBM: brush border membrane; ARG:
Effect of dietary FO and FXO on ARG-induced alterations in the antioxidant defense parameters in renal cortex and medulla
It is evident that reactive oxygen species (ROS) generated by various toxicants are the important mediators of cellular injury and pathogenesis of various diseases. Primary components of oxidative stress and cellular injury response include elevation of LPO, depletion of glutathione (GSH) and suppression of antioxidant enzymes. To ascertain the role of antioxidant system in the ARG-induced toxicity, the effect of ARG is observed on the oxidative stress parameters. ARG enhanced LPO and significantly altered antioxidant enzymes both in cortex and in medulla, although differently (Table 6). LPO measured in terms of the levels of MDA significantly enhanced in the cortex (+46%) and medulla (+40%) to similar extent whereas total-SH declined in these tissue (−29% to −37%). ARG treatment caused marked increase in SOD (+58%) and glutathione peroxidase (GSH-Px, +30%) activities but decrease in CAT (−16%) activity in the renal cortex. However, in the medulla, the activity of SOD (−58%) and CAT (−28%) significantly decreased, whereas the activity of GSH-Px (+45%) was significantly increased by ARG administration alone.
Effect of FO and FXO on the enzymatic and nonenzymatic antioxidant parameters in homogenates of cortex and medulla with ARG treatmenta

FO: fish oil; FXO: flaxseed oil; ARG:
a Results (specific activity expressed as micromoles per milligram protein per hour) are mean ± SEM for five different preparations.
b Significantly different from control.
c Significantly different from ARG at p < 0.05 by one-way analysis of variance. Values in parentheses represent percentage change from control.
We have previously shown that the diet supplemented with FO enriched in ω-3 fatty acids has been shown to reduce uranyl nitrate- and gentamicin-induced nephrotoxicity parameters by strengthening antioxidant defense mechanism, the protective effect of both FO and FXO enriched in ω-3 fatty acids was determined on ARG-induced oxidative stress parameters .The results indicate that FO and FXO dietary supplementation was able to ameliorate the ARG-induced oxidative damage in both renal cortex and medulla. ARG-induced increase in LPO and decrease in total-SH were not observed by feeding FO or FXO diet to ARG-treated rats. The activity of SOD and CAT in the cortex and the activity of CAT and GSH-Px in the medulla, respectively, remained significantly higher in FO/FXO + ARG rats compared with ARG and/or control rats. The results indicate marked protection by both FO/FXO diet against ARG-induced oxidative damage to renal tissues.
Effect of dietary FO and FXO on ARG-induced alterations in Na-gradient-dependent transport of 32Pi in BBMVs isolated from renal cortex
The bulk of filtered Pi in the kidney is reabsorbed by its proximal tubule. The Na-gradient-dependent {Naoutside (Nao)>Nainside (Nai)} transport of Pi in renal proximal tubule across its luminal BBM is an initial and regulatory step. The Pi is transported by the secondary active transport mechanism, which requires expenditure of energy in the form of ATP that is generated by the cellular metabolism. The uptake of 32 Pi was determined in the presence and absence of Na gradient in the initial uphill phase (30 s) and after equilibrium at 120 min in the BBM preparations. The rate of concentrative uphill uptake of 32 Pi in the presence of a Na gradient (NaCl in the medium) was markedly decreased by ARG treatment (Table 7). However, the uptake of 32 Pi at the equilibrium phase (120 min) when Nao = Nai was not significantly different between the two groups. Also Na-independent uptake (in the absence of a Na gradient, when NaCl in the medium was replaced by KCl where Ko > Ki) of 32 Pi at 30 s and 120 min was also not affected by the ARG treatment, indicating specific alterations only when Na gradient was present. When ARG treatment was extended to FO and FXO feeding rats, ARG-induced decrease in 32Pi transport was not observed.
Effect of FO and FXO on uptake of 32 Pi in BBMV from whole cortex with ARG treatmenta
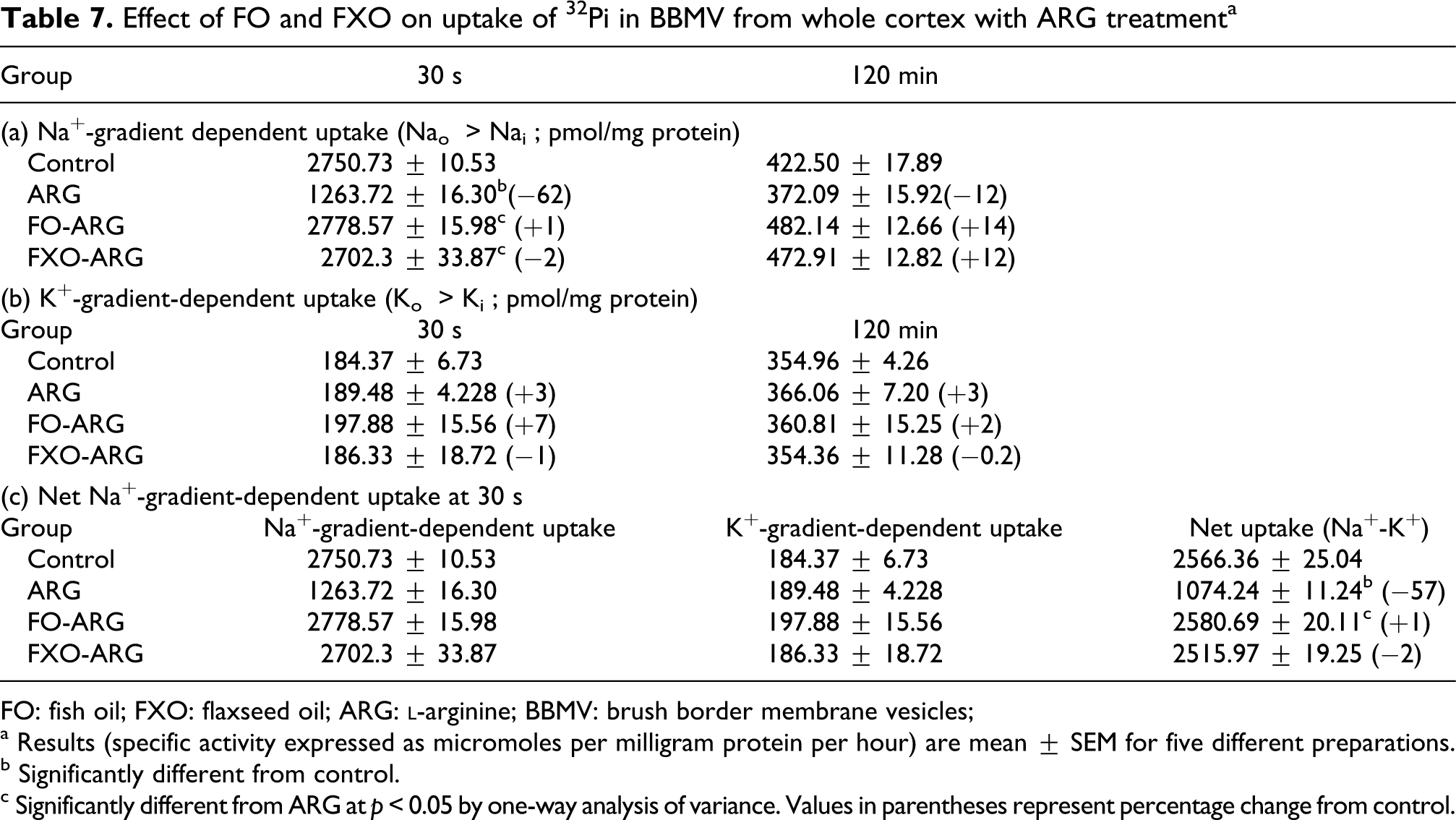
FO: fish oil; FXO: flaxseed oil; ARG:
a Results (specific activity expressed as micromoles per milligram protein per hour) are mean ± SEM for five different preparations.
b Significantly different from control.
c Significantly different from ARG at p < 0.05 by one-way analysis of variance. Values in parentheses represent percentage change from control.
Effect of dietary FO and FXO on ARG-induced alterations in the nitrite concentration in renal cortex and medulla
ARG increased the tissue concentrations of nitrite as shown in Figure 3(a) and (b), while the accumulation of nitrite being greater in renal medulla. Both FO and FXO improved the values to near control in renal medulla but only partially in renal cortex.

Effect of FO and FXO on tissue nitrite levels in (a) cortical and (b) medullary homogenates with ARG treatment. Results (nanomoles per gram tissue) are mean ± SEM for five different preparations. *Significantly different from control, †significantly different from ARG at p < 0.05 by one-way analysis of variance. FO: fish oil; FXO: flaxseed oil; ARG:
Discussion
ARG is a semiessential, proteinogenic amino acid and has been recognized as a naturally occurring molecule. 33 It is involved in major essential metabolic pathways and plays a key role in regulating inflammatory processes. 34 –39 Alternatively, ARG may enhance the synthesis of potentially detrimental NO from iNOS, which countered the neuroprotective effects of iNOS inhibitor in several studies. 1,40 Increased production of NO has been described in various human cancers, which might contribute to tumor development by favoring neoangiogenesis, tumor metastasis, resistance to DNA-damaging drugs and tumor-related immunosuppression. 41,42
Recently, the use of naturally occurring dietary substances for the prevention of various chronic diseases and against various toxicities is gaining interest. ω-3 fatty acid rich FO and polyphenol-rich green tea are shown to have several beneficial health effects against various pathologies including cancer, CVD and immune inflammatory disorders. 16,43 Watkins et al. have also suggested the therapeutic use of ω-3 PUFA and flavonoids against inflammation that is mediated by environmental pollutants. 44
In view of the above-mentioned points, the present work was undertaken to test the hypothesis whether dietary FO and FXO rich in ω-3 FA would be able to protect against ARG-induced nephrotoxicity/gastrotoxicity/hepatotoxicity. The effects of ARG treatment and dietary FO and FXO were determined on various nephrotoxicity parameters and on the enzymes of carbohydrate metabolism, BBM and oxidative stress in kidney and intestine of the rat.
As can be seen from the Results section, a single dose of ARG produced a typical pattern of nephrotoxicity manifested by increased Scr and elevated BUN accompanied by polyuria, proteinuria, glucosuria, phosphaturia and decreased creatinine clearance, indicating that a significant damage to kidney has occurred by ARG exposure. Both FO and FXO diet given 15 days prior to and during ARG administration prevented ARG-induced alterations in various serum/urine parameters. ARG-induced increase in Scr/BUN, phospholipidosis, proteinuria and glucosuria were all absent in FO/FXO fed ARG-treated rats. Thus, the results clearly indicated that ARG-induced alterations in serum/urine parameters were prevented similarly by both FO and FXO diets. The preventive effects of FO and FXO diet on ARG-treated rats were made possible because of the increase in serum Pi, glucose and decrease in Scr, BUN by FO/FXO alone. A positive balance of cholesterol and phospholipids (essential membrane components) in serum may facilitate the repair and regeneration of the membrane as required after ARG exposure.
The activities of BBM marker enzymes were determined to examine the structural and functional damage caused by ARG to BBM integrity and any protection against ARG actions exerted by FO/FXO diet. As shown in the Results section, the activities of ALP, GGTase and LAP markedly decreased by ARG treatment to control rats in renal cortical BBM preparations. However, the activities of ALP and GGTase were significantly increased by FO/FXO diet alone. Thus, FO/FXO diets to ARG-treated rats were able to prevent the decrease in the activities of these enzymes. Dietary fatty acids are known to be incorporated in the membrane causing changes in the lipid composition, which in turn affect the biophysical characteristics of the membrane and the activities of certain membrane-associated enzymes. 45,46 Thus, by the virtue of their influence on membrane physiology, dietary ω-3 PUFA might have played an important role in reducing the membrane perturbing effects of ARG. A similar positive impact of ω-3 PUFA against cyclosporine A-induced changes in membrane structure and function has been reported earlier. 45
The reabsorption of Na+ by renal proximal tubular BBM is considered to be the major function of the kidney because the transport of other ions and various solutes depend directly or indirectly on Na+ reabsorption. 47,48 Since these transports depend on the structural integrity of BBM and available energy as ATP provided by various metabolic pathways, it is imperative that any alterations to BBM integrity and metabolic pathways caused by toxic insult or otherwise would determine the rate of renal transport functions. 48 As shown in the Results section, the activities of various enzymes involved in glycolysis, TCA cycle, gluconeogenesis and HMP shunt pathway were significantly affected by ARG treatment although differently. Taken together, ARG significantly increased the activity of HK and LDH (glycolysis), whereas the activity of MDH (TCA cycle) significantly decreased. The marked decrease in MDH activity indicates an impaired oxidative metabolism of glucose and decreased ATP production and hence depressed renal transport function. Although the actual rates of glycolysis and other pathways were not determined, the increased activity of LDH along with simultaneous decline in TCA cycle enzymes suggests a shift in energy production from mitochondria to anaerobic glycolysis due to the mitochondrial damage caused by ARG. The decreased activities of gluconeogenic enzymes FBPase and G6Pase may be the result of decrease in TCA cycle enzyme activities. This can be further explained by the fact that the reduced activities of TCA cycle enzymes especially that of MDH may have reduced the production of oxaloacetate from malate, which is required not only for the continuation of TCA cycle but also for gluconeogenesis. The oxidative conversion of glucose or glucose-6-phosphate to 6-phosphogluconate by G6PDH of HMP shunt pathway was also lowered in part due to mitochondrial dysfunction by ARG. FO/FXO diet normalized the activities of the metabolic enzymes, when they were given to ARG-treated rats, which prevented ARG-induced decrease in various enzymes involved in glucose degradation and production.
To further correlate BBM integrity and ATP production, enzymes of oxidative metabolism were analysed in BBM preparations. The results indicate that ARG caused reduction in the enzyme activities of oxidative metabolism and BBM, whereas FO and FXO diet increased most of them would lead to decrease/increase production of ATP by ARG and FO/FXO, respectively. Thus, both the situations would affect transport functions of the kidney. Indeed, Na-dependent transport of Pi across renal cortical BBM was significantly declined by ARG exposure whereas increased by FO/FXO diets. ARG-induced decrease in Pi transport was also prevented by FO/FXO diets when given together (FO-/FXO-ARG), indicating a positive correlation between ARG and FO/FXO actions on Na-Pi transport.
It has been reported that some toxicants including certain drugs and heavy metals exert their effect by inducing the generation of ROS/reactive nitogen species (RNS). However, redox stress can occur as a result of either increased ROS/RNS generation or decreased antioxidant defense or both. Of them, SOD, CAT and GSH-Px constitute the main components of the antioxidant defense system. The present results demonstrate that ARG exposure causes increase in the activities of SOD and decrease in the activities of CAT and GSH-Px, accompanied by increased LPO and decreased total-SH in renal cortex, whereas all antioxidant enzymes were depressed in renal medulla.
The feeding of both FO and FXO diets with ARG treatment prevented ARG-induced increase in LPO and decrease in total-SH and antioxidant enzymes activities remained normal in FO-/FXO-ARG treated rats compared with control rats. Taken together, the present observations clearly demonstrate that FO/FXO diets were able to prevent many of the adverse alterations caused by ARG in the metabolic, BBM and antioxidant enzymes in the kidney. Dietary FO/FXO have been shown to improve antioxidant defenses in a variety of pathophysiologic situations, where the oxidant/antioxidant defense mechanism are decelerated. 49 –52 Thus, the changes in the cell antioxidant status and membrane fatty acid composition could significantly alter the ability of the cell to cope with ARG-induced toxicity.
The mechanism by which ARG causes these effects is not fully understood. Accumulating evidence suggests that oxygen free radicals, NO, inflammatory mediators, all have a key role in the development of ARG-mediated alterations. 53 –56 In general, NO is generated from ARG by an enzymatic pathway (NOS) originally demonstrated in vascular endothelial cells. 57 Under physiological conditions, constitutive NOS (endothelial nitric oxide synthase (eNOS)) results in a low level of NO, while in different inflammatory processes iNOS produces larger quantities of NO in various cell types. The activity of eNOS was shown to be increased early (24–48 h) then declined thereafter at 48 h, whereas iNOS activity was found to be increased after 24 and 48 h vs control and remained higher thereafter. 6,58 It has been suggested that both the NOS forms play important role in the development of ARG-induced toxic alterations. 6 It has been demonstrated that NO derived from ARG is a potential source of redox stress. It can be quickly cleared through reacting with superoxide (O2 −) to generate peroxynitrite (ONOO), while cells are in a pro-oxidative state. As a highly reactive species, ONOO can react via homolytic or heterolytic cleavage and generate secondary constituents of nitroxidative stress and highly ROS/RNS including NO2 +, NO2 − and OH radicals. The present results show that ARG caused increase in LPO and decrease in SH content, whereas the activities of antioxidant defense system, for example, SOD, GSH-Px and CAT were perturbed in the kidney. The results confirmed previous studies that showed that ARG caused morphological changes in the liver, kidney and some other tissues including pancreas, which were demonstrated to be mediated by perturbation in the oxidative stress parameters. 6 In contrast, FO and FXO, major source of ω-3 fatty acids by altering membrane fatty acid composition, appeared to affect membrane organization and functions. Both FO and FXO appear to accelerate repair and/or regeneration of injured organelles, for example, mitochondria, peroxisomes and increased activity of TCA cycle and BBM as evidenced by increased activities of the enzymes of carbohydrate metabolism. Most importantly, by activating endogenous antioxidant defense mechanism, FO and FXO provided protection from ARG-induced RNOS production.
We conclude that while ARG elicited deleterious nephrotoxic and other adverse effects by causing severe damage to renal mitochondria, BBM and other organelles and by suppressing antioxidant defense mechanism, dietary supplementation with FO/FXO enriched in ω-3 fatty acids caused improvement in nutrition/energy metabolism, BBM integrity, 32 Pi transport capacity and antioxidant defenses and thus prevented ARG-induced various deleterious effects. ARG-induced uncontrolled NO production and mediators such as ONOO and other RNOS appear to cause most of ARG-induced deleterious effects and FO/FXO elicited increased antioxidants counter these alterations. Based on our present observations and already known health benefits, we propose that dietary FO supplementation may provide a cushion for a prolonged therapeutic option against ARG-induced various adverse effects without harmful side effects.
Footnotes
Declaration of Conflict of Interest
The authors declared no conflicts of interest.
Funding
This work was supported by Indian Council of Medical Research (ICMR; New Delhi, India) through the award of Junior Research Fellowship (JRF)/Senior Research Fellowship (to SP and MWK); Council of Scientific and Industrial Research (CSIR; New Delhi, India) through the award of JRF (to SK). Financial support was also provided to the department by University Grants Commission (UGC-DRF), Department of Science and Technology (DST-FIST), and a research grant (58/21/2001-BMS) from ICMR (to ANKY).