
Abstract
Environmental exposure to mercury can cause a number of adverse effects in humans including the disruption of endocrine function that may result in sex-specific effects. The present study was designed to characterize sex-specific effects of chronic inorganic mercury exposure on toll-like receptor (TLR) 2 and TLR4 and inflammatory signaling in the liver of prairie voles (Microtus ochrogaster). Following 10 weeks of exposure to mercury via drinking water, effects on protein expression levels of TLR2 and TLR4 and the downstream p38 mitogen-activated protein kinase (p38 MAPK) and nuclear factor-kappa (NF-κB) signaling pathways were assessed. Using immunoblot analysis, we found that mercury exposure significantly enhanced the expression of TLR4 and activated p38 MAPK and NF-κB pathways in vole livers. This is the first report indicating that TLR4 may serve as a sensor for chronic mercury exposure in the liver. Further, compared to females, mercury-treated male voles exhibited significant increases in activated p38 MAPK and a greater extent of liver damage. Together, these findings establish sex-specific liver immunomodulation and cellular signaling following chronic inorganic mercury exposure. Furthermore, this study also supports the use of voles as biomarkers of environmental mercury contamination and offers a promising in vivo tool to test various therapeutic strategies for mercury detoxification.
Introduction
Human exposure to heavy metals is ubiquitous and is an important public health hazard. Industrial activities have produced generalized increases in environmental exposure to toxic metals via contaminated air and soil, and of atmospheric deposition of volatiles and dust.1,2 Consumption of top predators such as tuna is a concern since these animals are the penultimate step in a bioaccumulation process that can lead to high human exposures. 3 Excessive exposure to heavy metals may alter behavior through their actions within the central nervous system, and can cause neurological, pulmonary, renal, gastrointestinal, or hepatic damage in experimental animals and humans.4–6
Among the heavy metals, mercury is an especially important environmental and industrial pollutant since the elemental form is volatile and can easily enter the atmosphere. For example, emissions from landfills can contribute significant amounts of gaseous mercury emissions. 7 Mercury is highly toxic and, upon absorption into the blood stream, readily gets absorbed in various organs. Being lipid soluble, mercury accumulates in fat—particularly in nerve sheathes, brain, and liver. The liver is a major site of metabolism for mercury and accumulation in the liver can cause hepatocellular damage. 8 Mercury accumulation in the human liver due to chronic exposure is frequently considered in risk assessment for associated diseases.9–11 For example, some individuals, such as those with autistic spectrum disorders, are genetically predisposed toward problems of detoxification. 12 More recently, mercury has been recognized as an endocrine disruptor and can inhibit the actions of hormones or interfere with the normal functions of the nervous system. 13 It has been suggested that these effects are manifested either by affecting the hormone synthesis or preventing the binding of hormones to their receptors.14,15 Interestingly, there is evidence that toxic effects of heavy metals are manifested differently in men and women, however, the underlying mechanisms are poorly understood. 14
Mercury also plays a role in the induction or exacerbation of some autoimmune diseases; however, the mechanisms are not yet fully understood.16,17 Possible alterations in hormonal receptors or T-helper 1(Th1)/Th2 reactivity by mercury may constitute the trigger for autoimmunity in susceptible individuals. Other effects of mercury on the immune system include immune suppression and decreased host resistance.18,19 The immune response initiates liver damage by hepatic neutrophil accumulation and injury to some extent is caused by the activation of a local cytokine network. The liver has a complete innate immune system consisting of Kupffer cells as well as hepatic natural killer cells expressing toll-like receptor (TLR) sensors which are involved in the recognition of microbe-derived products as well as cell damage-associated products that initiate subsequent inflammatory responses.20,21 Activation through TLR4 leads to the induction of various intracellular signaling pathways including the mitogen-activated protein kinases (MAPKs), which in turn activate transcription factors such as NF-κB, regulating the proinflammatory response. 22 The inactive, p65/p50 NF-κB dimer is normally sequestered in the cytoplasm, bound to inhibitory kappa B (IκBα). Following cellular activation, kinases including p38 MAPK are upregulated, followed by phosphorylation of IκBα by IκB kinase and subsequent polyubiquitination. 23 Proteolytic degradation of the ubiquitinated complex results in the release of active NF-κB, which can then translocate into the nucleus and bind κB sites in the promoter region of various genes. 24
Evidence is emerging that TLR4 may be a multifunctional receptor involved in innate immunity in the liver. 25 Further, liver is responsive to hormones and expresses both estrogen and androgen receptors and thus may be a target of endocrine disruptive chemicals. 26 Unfortunately, there is little information available about the effects of mercury on innate immune signaling in the liver and any related sex-specific changes.
Based on these reports, we hypothesized that chronic mercury exposure might induce TLR responses in the liver, affecting downstream inflammatory signaling leading to immune activation and hepatocyte damage. To test this hypothesis, we characterized the effects of chronic mercury exposure on TLR expression and inflammatory signaling and examined sex-specific effects on liver immune responses in prairie voles. Voles are potentially useful sentinel animals for monitoring environmental toxins.27,28 Cobb et al. documented elevated levels of mercury in various tissues following the exposure to organic mercury. 29 Furthermore, studies in voles and other mammals show that the liver seems to be especially prone to mercury accumulation.11,29 In the present study, TLR2 and TLR4 protein expression levels and key inflammatory molecules including those involved in p38 MAPK/nuclear factor κB (NF-κB) pathways were assessed in mercury-treated voles. We found that mercury exposure led to enhanced expression of TLR4 in both sexes, and there are sex-specific changes in downstream inflammatory signaling.
Materials and methods
Animal husbandry
Liver tissues used in the present study were collected from prairie voles of both sexes that were used in previously described behavioral and neurochemical studies on the effects of chronic metals exposure on central nervous system function. 30 Voles were bred and reared on-site in a captive breeding-colony housed in United States Department of Agriculture (USDA)-approved facilities. General animal care was provided by laboratory animal resources personnel and animals were monitored daily. Voles were maintained at ~21°C with a 14:10 light: dark cycle. Breeding pairs were housed in plastic cages (20 × 25 × 45 cm) containing corncob bedding with Timothy hay as nesting material. Animals had ad libitum access to drinking water and food (Purina rabbit chow supplemented with black-oil sunflower seeds). After weaning (~20 days of age), pups were housed as same-sex pairs in shoebox style plastic cages (10 × 17 × 28 cm), also with ad libitum access to food and drinking water. Veterinary staff was available for consultation regarding animal health and welfare. All experimental manipulations and animal handling procedures were approved by the Oklahoma State University Center for Health Sciences Institutional Animal Care and Use Committee.
Experimental subjects
Experimental subjects were sexually naive adult male and female prairie voles. The breeding colony is out-crossed periodically with wild stock, and the animals used in this study were of the F3 and F4 generations relative to most recent out-crossing. After weaning, males were housed in a single-sex room until used in experiments, while females were housed in the same room as the breeders. However, during the experimental manipulations, subjects and control animals of both sexes were housed in a single room. The average age at which the animals were killed was similar for both sexes (males 142.2 ± 8.4; females 144.0 ± 7.3 days)
Mercury treatments and tissue collection
Each same-sex pair was randomly assigned to groups that received either the standard drinking water available to the general colony, or drinking water containing 60 ppm HgCl2. Solutions were provided to the animals in Nalgene bottles (~300 ml) outfitted with rubber stoppers and metal sipper tubes. Both standard and mercury-containing bottles were changed at normal intervals with the remainder of the animal colony unless contaminated with bedding materials, in which case fresh solutions were provided. At each bottle change, both the old bottles and the replacement bottles were weighed to the nearest 0.1 g and the difference between weight on and weight off was assumed to represent the total liquid ingested. An unavoidable limitation of housing voles in pairs is that it is not possible to determine exactly how much liquid each animal consumes, and thus it was assumed that each animal consumed half of the total. Details of water and mercury consumption are reported elsewhere. 30 After 10 weeks of mercury exposure, the animals were euthanized by carbon dioxide inhalation and the left posterior lobes of each animal’s liver were removed and immediately chilled on ice. Livers were cut into two sections with one half stored at −80°C until used in assays and the other half fixed in formalin for histopathological analyses.
Western blotting
Western blot analysis was performed for TLR4, TLR2, MAPK p38, NF-κB, and IκBα proteins derived from vole liver tissue extracts. For TLR4, TLR2, and p38, a total protein whole lysate was used. 31 NF-κB and IκBα were estimated in nuclear and cytoplasmic fractions of the tissue extracts, respectively, prepared from tissue samples using nuclear–cytosol protein extraction kits as per the manufacturer’s instructions (Active Motif, Carlsbad, CA, USA). Protein content of the lysates was determined by bicinchonic acid (BCA) assay (Pierce Chemical Co., Rockland, IL, USA). Samples containing 60 μg of protein were separated using 4–12% Bis-Tris (Invitrogen, CA, USA) gels and then transferred onto 0.2 μM nitrocellulose membranes (Bio-Rad, CA, USA). For immunoblotting, the membrane was blocked in 5% non-fat dry milk in a solution of Tris-buffered saline containing 0.1% Tween-20 (TBST) and then incubated with anti-TLR4 (goat, M-16) or anti-TLR2 (rabbit, H-175) polyclonal antibodies (1:500 dilutions, Santa Cruz Biotechnology, Santa Cruz, CA, USA) overnight at 4°C. The membranes were then washed and incubated with anti-goat or anti-rabbit alkaline phosphatase-conjugated secondary antibodies (1:30,000, Sigma, St. Louis, MO, USA). Signal visualization was via chemiluminescence (Pierce Chemical Co., Rockland, IL, USA). Images were captured digitally with Alpha InnoTech instrumentation (Alpha InnoTech Corp. San Leandro, CA, USA) and quantified using Image J (NIH Image J, Scion Corp.). Similar methods were used to probe the membranes using rabbit antibodies to phospho-p38 (pp38) and p38 MAPK, phospho-p65 NF-κB and p65 NF-κB, and phospho-IκBα and mouse anti-body to IκBα (1:1000 dilutions, Cell Signaling Technology, MA, USA). Positive staining for GAPDH or for histone was considered evidence of purity of the cytosolic and nuclear fractions, respectively (data not shown). Finally, all membranes were re-probed with anti-β-actin rabbit polyclonal antibody at a 1:2000 dilution (Sigma, St. Louis, MO, USA) as a loading control. The band intensities for the proteins of interest were normalized to those of β-actin in the same sample. The phosphorylated forms of proteins were expressed as a ratio relative to the total unphosphorylated forms.
Data presentation and statistical analysis
Data are presented as means ± SEM. All Western blot assays were repeated twice with similar trends. Representative images are depicted in the results. Comparison between treatments and sex effects were analyzed by two-way analysis of variance (ANOVA) followed by Student Newman-Keuls analysis (SNK) whenever appropriate using Statistica software (San Diego, CA, USA). A p value <0.05 was considered statistically significant in all analyses.
Histopathological assessment of livers
After euthanasia, livers were collected from 10 mercury-treated and 10 control voles of each sex and processed for histopathological evaluation. Most livers (n = 40) were fixed using 24-h immersion in 10% neutral buffered formalin, embedded in paraffin, and 5-μm sections were collected onto charged glass slides. Additional livers were sectioned as unfixed frozen tissue on a cryostat. Tissue from both sectioning methods then was stained with hemotoxylin and eosin (H and E) and evidence of hepatic damage was assessed by a pathologist blind to the treatments associated with each liver. 32 Hepatic damage was qualitatively scored from one (little to no evidence of hepatic damage with few lymphocytes) to three (extensive damage, many lymphocytes) and the number of animals in each category within each sex for each treatment group were used as the basic unit of measure. Micrographic images were captured using a Nikon Eclipse Binocular Microscope fitted with a Nikon Digital Camera model DXM1200F. Chi-squared analyses were used to assess the data in three different combinations: each treatment × sex group separately (3 × 4, 10 d.f.); grouped by treatment only (3 × 2, 4 d.f.); and grouped by sex only (3 × 2, 4 d.f.).
Results
Effects of mercury on TLR4 and TLR2 protein expression
The effects of mercury exposure on the TLR4 and TLR2 protein expression in the livers of male and female voles are shown in Figures 1 and 2. There was a main effect of treatment (F 1, 28 = 11.87, p < 0.003) for TLR4. Mercury-treated voles exhibited significantly higher TLR4 expression compared with their respective control groups (Figures 1A and 2A). There was no sex difference (F 1, 28 = 0.52, p = 0.48) and no interaction (F 1, 28 = 0.67, p = 0.42) for TLR4. TLR2 protein expression was slightly higher in male voles treated with mercury, however, this difference was not statistically different as there was no effect of treatment (F 1, 28 = 2.07, p = 0.16) or sex (F 1, 28 = 0.90, p = 0.35) and no interaction (F 1, 28 = 1.99, p = 0.17). Overall, the expression of TLR2 protein in voles treated with mercury or water did not differ (Figures 1A and 2B).

Western blot results for inflammatory proteins in voles that received either 60 ppm HgCl2 solutions or unadulterated water as their sole drinking water sources for 10 weeks. Representative blots from both sexes in each treatment group are presented. Blots within a row are all from the same membrane; blots within a column are all from the same subject. Membranes were stripped and re-probed to produce each row. (A) Toll-like receptors (TLR) 4 and 2; (B) phosphorylated and total p38 MAP-kinase; (C) phosphorylated and total IκBα; (D) phosphorylated and total nuclear factor-κB (NF-κB). β-Actin was probed on each membrane to serve as a loading control.
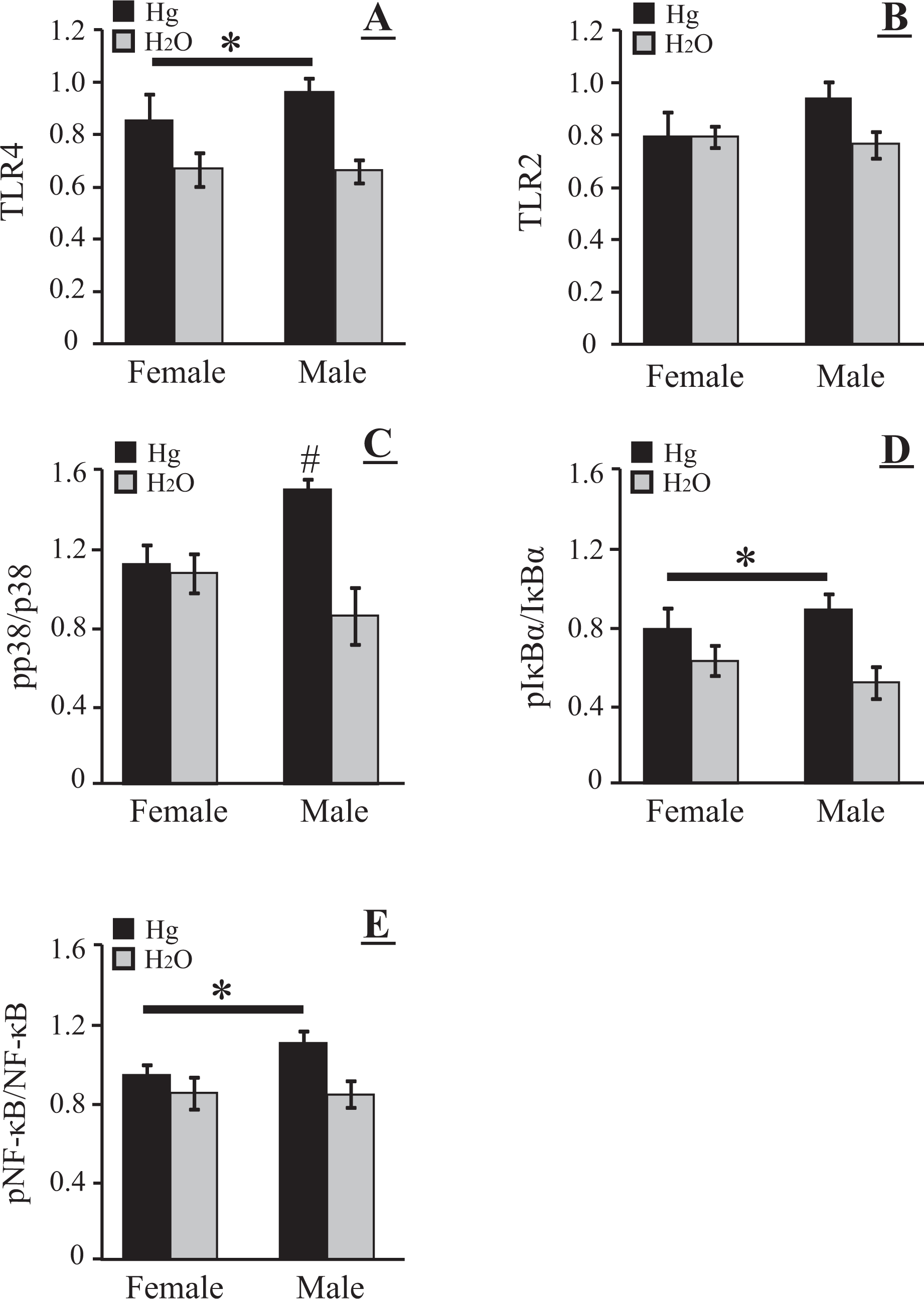
Effects of chronic mercury ingestion on inflammatory proteins in vole liver. (A) Toll-like receptor (TLR) 4; (B) TLR-2; (C) ratio of phosphorylated to total p38 MAPK (pp38/p38); (D) ratio of phosphorylated to total IκBα; (E) ratio of phosphorylated to total nuclear factor-κB (NF-κB). Mercury-treated (dark bars) and control (light bars) means (±SEM) are presented. An asterisk above a line indicates a significant effect of mercury ingestion (p < 0.05) on a given measure, but no differences between male and female responses. #Indicates mercury-treated males had a higher ratio of phosphorylated to total p38 MAPK than did either control males or mercury-treated females.
Effect of mercury on p38 MAPK
Mercury ingestion did not alter the total p38 MAPK protein expression in vole livers (treatment F 1, 28 = 0.52, p = 0.48; sex F 1, 28 = 0.79, p = 0.38; interaction F 1, 28 = 0.44, p = 0.51) however, mercury ingestion did change patterns of phosphorylation of p38 MAPK (Figures 1B and 2C). Although there was no significant effect of sex on p38 MAPK phosphorylation (F 1, 28 = 0.55, p = 0.46), there was a significant treatment effect (F 1, 28 = 10.79, p < 0.004) and a significant sex by treatment interaction (F 1, 28 = 8.23, p < 0.01). SNK analysis revealed that male voles that received mercury in drinking water displayed higher ratios of pp38 MAPK/p38 MAPK than did either similarly treated females (p < 0.02) or males that received just water to drink (p < 0.002)
Effect of mercury on IκBα
Mercury ingestion significantly altered the expression of IκBα protein in vole livers. There were main effects of both treatment (F 1, 28 = 16.47, p < 0.001) and sex (F 1, 28 = 94.96, p < 0.001) and a significant interaction (F 1, 28 = 18.72, p < 0.001). Males in either treatment condition displayed higher levels of total IκBα protein than corresponding females (both p’s < 0.002) and, in addition, males that received mercury displayed higher IκBα levels than control males that received water (p < 0.001). Mercury exposure-induced phosphorylation of IκBα in the cytoplasm as determined by Western blot analysis. Mercury-treated voles exhibited significantly higher pIκBα/IκBα (F 1, 28 = 9.29, p < 0.01) than control voles (Figure 1C and 2D).
Effect of mercury on nuclear NF-κB
There were no main effects and no interaction on total NF-κB protein expression in vole livers after mercury ingestion (treatment F 1, 28 = 0.34, p = 0.57; sex F 1, 28 = 0.86, p = 0.36; interaction F 1, 28 = 0.05, p = 0.82). Phosphorylation of p65 NF-κB (pNF-κB) was shown in the nuclear fraction as determined by Western blotting (Figure 1D), and mercury ingestion significantly affected the ratio of pNF-κB/NF-κB (Figure 2E; F 1, 28 = 6.67, p < 0.02): there was no main effect of sex (F 1, 28 = 1.33, p = 0.26) and no interaction (F 1, 28 = 1.53, p = 0.23).
Pathological examination of livers
The majority of females (15/20), regardless of treatment, displayed minimal (n = 10) to no (n = 5) apparent liver damage. Livers from these females were characterized by radial distribution of hepatocytes around the large central vein, organized in a compact manner within hepatic lobules with normal appearing sinusoids lined by endothelial cells (Figure 3A), although there often was evidence of minimal hepatocellular damage (Figure 3B). Among the remaining females, three of the mercury-treated and two water-treated animals displayed moderate liver damage (Figure 3C). In contrast, half of the males (10/20), including four water-treated animals, displayed evidence of greater hepatocellular damage, again regardless of treatment. In these animals, hepatocellular damage ranged from moderate to extensive showing necrotic changes (Figure 3D), the latter characterized by ballooning degeneration of hepatocytes with acidophil bodies or Malory hyaline, focal microvesicular steatosis. The damage was seen mainly around the central veins (zones 2 and 3) and less around the portal areas (zone 1). There was focal central vein damage with sinusoidal dilatation and there was an occasional lymphocyte. No significant parenchymal inflammation in the form of lymphocytes, plasmahistiocytic cells or neutrophils was seen in any of the livers. There was no evidence of cholestasis or any significant fibrosis.

Representative photomicrographs of prairie vole liver sections used for histological examination of potential effects of chronic mercury consumption. Each micrograph is an example of tissue showing one of the four damage classifications. (A) No hepatocellular damage; (B) mild hepatocellular damage; (C) moderate hepatocellular damage; (D) maximum hepatocellular damage. H&E, ×400.
Although males tended toward greater liver damage, χ2 analyses revealed no significant group differences when data were grouped by animal sex (χ2 = 3.14, d.f. = 4), treatment (mercury vs. H2O; χ2 = 1.65, d.f. = 4), or sex × treatment combinations (χ2 = 5.83, d.f. = 10).
Discussion
There is a rising incidence of nonalcoholic fatty liver disease (NAFLD) in the United States and other developed countries and one of the factors to which this is attributed is industrial pollution. 33 Recently, using data from the National Health and Nutrition Examination Survey (NHANES) for 2003–2004, researchers found detectable levels of 196 different pollutants in the blood or urine of adults. Among these pollutants, mercury was commonly found and whole blood total mercury was associated with increased liver enzymes and thus may be associated with liver disease. 34 Such results highlight the need for increased understanding of various pathogenic effects of environmental pollutants, such as the effects of mercury on the immune system and its resultant effects on various organ systems. Exposure to sub-lethal levels of inorganic mercury is known to alter immunocompetence.18,35,36 For example, the pathogenic deposition of metals in a host increases the susceptibility to infections as evidenced by a reversal of genetic resistance to Chlamydia-induced arthritis after mercury exposure. 16 Mercury-induced autoimmunity in genetically susceptible rodents has been widely reported. 37 However, sex-based differences in the effects of chronic mercury exposure on innate immune signaling and inflammatory pathways in different organ systems have received relatively little attention. The present study was designed to test the effects of chronic mercury exposure on inflammatory signaling in the livers of male and female prairie voles.
Voles were selected as the animal model for several reasons. First, there are previous reports on the potential usefulness of this species as a sentinel for environmental contamination. 38 Second, chronic exposure to at least some forms of mercury increased tissue levels of the metal in vole liver. 29 Third, we recently reported sex-specific deficits in social behavior in these animals after chronic exposure to mercury, suggesting that metals may contribute to the development of autism-like symptoms. 30 Finally, since female prairie voles do not display an estrous cycle, we could examine both sexes without the necessity for surgical manipulation to reduce hormonal variability. 39 Related in vivo studies, assessing chronic inorganic mercury exposure on inflammation and liver toxicity in rodents have utilized 14-day exposure via drinking water or daily intraperitoneal injections for 20 days.18,35,36,40 However, the present study used voluntary mercury intake through consumption of drinking water for a substantially longer period of time, 10 weeks. This mode of exposure is biologically relevant given that mercury contamination is estimated to be largely through intake of contaminated food and water and can occur for extended periods of time.
In our study, mercury differentially affected TLR protein expression in the livers of both sexes as demonstrated by increased TLR4 expression but minimal change in TLR2. This is the first description of TLR expression in vole liver and, to our knowledge, is the first report of a mercury-induced change in TLR4 expression in the liver. The consequences of increased TLR4 expression in the vole liver remain to be determined. However, elevated TLR4 expression in mercury-exposed animals suggests a priming of inflammatory signaling. Mercury and lipopolysaccharide (LPS) that is present in the liver may synergistically induce activation of TLR4. TLR4 levels are known to increase in the liver following infection or disease-induced cellular damage.41–43 Any changes in TLR expression in the liver may modulate host susceptibility to disease. The TLRs recognize pathogen-associated molecular patterns to detect the presence of pathogens and also play a major role in the regulation of inflammation even under non-infectious conditions, such as injury, by their ability to recognize several endogenous TLR ligands termed as damage-associated molecular patterns that are up-regulated during liver injury. 22
Activation of TLR can result in the phosphorylation of p38 MAPK and, downstream, activation of IκBα. 23 In our study, male voles that received mercury in drinking water displayed significantly higher levels of pp38 MAPK than similarly treated females. Altered levels of pIκBα and pNF-κB p65, and, in males, pp38 MAPK in the liver following chronic mercury exposure provide further evidence of mercury-induced hepatic inflammatory responses that in some cases are sex-specific. These combined data indicate that TLR4, p38 MAPK, and NF-κB associated signaling pathways are sensitive to mercury exposure, especially in males. Collectively these signaling pathways induce pro- and anti-inflammatory cytokines that may be beneficial in the short term to modulate injury effects. However, prolonged or exaggerated activation of TLR may have deleterious effects, ultimately leading to pathophysiological changes. A similar sex disparity in the production of pro-inflammatory cytokines in the liver in response to a chemical carcinogen, diethylnitrosamine, has been reported. 44 Several in vitro and in vivo experiments similarly confirm divergent effects mediated by estradiol or testosterone on signal transduction pathways observed in immune and other cells.45–49 Tissue retention of mercury is known to be higher in male mice than in female mice exposed to mercury chloride, again indicating sex differences in mercury accumulation and toxicity.50,51
Mercury also can alter the phosphorylation of these signaling molecules in immune cells.18,35,36,52,53 For instance, continuous mercury exposure (2 weeks, in drinking water) enhanced p38 MAPK phosphorylation in mouse liver, but did not activate extracellular-regulated kinase (ERK). 36 Similarly, mercury exposure activated p38 MAPK in the murine macrophage cell line, J774A.1. 35 One study found that pre-exposure to mercury potentiates LPS-induced NF-κB activation in human monocytic cells. 52 However, in a separate study utilizing the J774A.1 cell line, mercury inhibited LPS-induced NF-κB nuclear translocation, while mercury alone upregulated expression of the proinflammatory cytokine, tumor necrosis factor-α. 35 Differential levels of mercury exposure may be responsible for observed differential effects on cell signaling in various studies and such differences also may depend on the subjects or cellular population under study.
Limitations of the present study may include having single time point observations. In future experiments, both early and late time point specific observations may be needed to study the relationship among various inflammatory markers. 54 Mercury-exposed males showed the greatest damage in hepatocytes. The histopathological changes showing hepatocyte necrosis with occasional focal microvesicular steatosis seen in these animals were similar to the observations made by others55,56 studying various animal models of chronic mercury exposure. However, we did not directly correlate tissue damage with the various inflammatory markers. Also, in the present study, the inflammatory markers were studied in total tissue extracts that can also mask the activity levels of individual cell types involved. Detailed studies are necessary to identify the specific cell populations in the liver that are responsible for the induction of inflammatory molecules on exposure to mercury. Kupffer cells (tissue macrophages in the liver) are likely involved given that they are the primary site of TLR4 expression and largely responsible for responses to infection and other insults in the liver leading to activation of liver natural killer T cells. 41 However, hepatic non-immune cell populations like hepatocytes also express TLRs.57,58 Finally, there was occasionally moderate liver damage among some water-treated animals, suggesting some minimum amount of aging degradation, despite the absence of the toxin. As such, it is possible that liver degradation associated with aging also might have obscured subtle effects.
The mechanism and implications of hormonal or sex-specific interaction on mercury-induced inflammation in the liver remain to be elucidated. We need to understand the agonistic or antagonistic effects mediated by these metal pollutants on various hormonal receptors expressed on various cells. Thus far, the known cytotoxic effects of mercury in the liver have been found to be mediated largely via generation of reactive oxygen species and glutathione depletion. 59 To our knowledge, this is the first study suggesting involvement of TLR4 during chronic mercury exposure that may play an important role in inflammation in the liver. Further investigation is needed to determine the mechanism by which mercury induces these changes in TLR4 expression in both sexes at the cellular level and the activation of p38 MAPK and NF-κB specifically in males.
Clinical implications of these results include sex disparities observed in the generation of various organ specific autoimmune diseases in various populations and also the development of chronic liver diseases or liver cancer specifically in males or post-menopausal females. 26 How many of these clinical entities may result from environmental chemical insults serving as co-factors for increasing their susceptibility to various liver diseases remains to be determined. In the future, it may be crucial to study specific signal transduction pathway perturbations as a risk assessment in a particular target organ that may be predictive of health endpoints. The present findings suggest that therapeutic modulation of TLR4 and downstream signaling molecules may be feasible strategies for reducing liver injury and inflammation post-chronic mercury exposure.
In summary, we have provided the initial characterization of TLR4, TLR2, p38 MAPK, IκBα, and NF-κB protein expression in vole liver. Also, we have demonstrated that TLR4-associated inflammatory signaling in the liver may play an important role in the initiation of mercury-induced changes in the liver. Furthermore, this study shows that male voles are particularly responsive to mercury-induced activation of p38 MAPK. Thus, studies related to sex specific effects of modulation on androgens or estrogens affecting inflammatory signaling in the liver need further investigation. Overall, these insights support the use of voles (particularly males) as sentinels for environmental mercury contamination and also an important in vivo model to study dose dependent toxic or immunomodulatory effects of mercury in various organs, thus, providing information that is critical to public health.
Footnotes
This study was financially supported by Cancer Sucks, Bixby, Oklahoma; Oklahoma State University Center for Health Sciences Intramural Research awards to RK (Dept. of Biochemistry and Microbiology) and to JTC (Dept. of Pharmacology and Physiology); and an NIH grant #HD48462 to JTC.