
Abstract
The aim of this study was to evaluate the attenuating effect of given selenium and/or magnesium on ethanol-induced oxidative stress, disturbances of liver function and cholesterol metabolism. Forty male rats were divided into five groups: C – control, Et – intoxicated with alcohol (15% solution in drinking water), Et + Mg, Et + Se, Et + Mg + Se – intoxicated with alcohol and supplemented with selenium (0.4 mg Se/l water), magnesium (100 mg Mg/l water) and combination of Se and Mg, respectively. The experiment was carried out over the 3 months. The results show that the chronic ingestion of alcohol induces lipid peroxidation and histopathological changes in liver. Supplementation with magnesium only partially alleviates oxidative stress and damages in this tissue. The both selenium alone and combination of magnesium and selenium significantly elevated total antioxidant status (TAS) in serum, activity of glutathione peroxidase and ratio of reduced glutathione to oxidized glutathione (GSH/GSSG) in liver and retarded oxidative stress and histopathological changes in this tissue. Chronic administration of ethanol (alone and with magnesium) resulted in significant decrease in the serum total cholesterol and retardation in the body weight gain in comparison with the control group. In the groups supplemented with selenium and selenium and magnesium simultaneously, concentration of total cholesterol in serum and body gains was similar to the control group. Supplementation of Se or selenium and magnesium simultaneously significantly enhances antioxidant defence and is more effective against alcohol-induced oxidative stress, disturbance of liver function and cholesterol metabolism than the separate use of magnesium.
Introduction
In Europe, where alcohol consumption per capita is the highest (compared with other continents), its abuse constitutes particularly serious social and health problems. It is estimated that 23 million Europeans are addicted to alcohol. Drinking and alcohol abuse usually initiate in adolescence or early adulthood, when the risk of addiction is the highest and health consequences may be particularly serious due to long-term exposure to alcohol over the entire life.1,2 Liver is the main site of ethanol metabolism and the most damaged organ. Alcoholic liver disease represents a spectrum of clinical symptoms and morphological changes that range from fatty liver, hepatic inflammation and necrosis to progressive alcoholic cirrhosis and is one of the major cause of disease and death worldwide. 3 Liver damages due to abuse of the alcohol are associated with the formation of toxic products of the ethanol metabolism such as acetaldehyde, and many reactive oxygen species (superoxide radical, hydrogen peroxide, hydroxyl and 1-hydroxyethyl radicals) result in oxidative stress that initiates cell injury.4,5 Oxidative stress and liver injury are partially caused as a result of redox changes in hepatocytes associated with increased conversion of NAD to NADH during alcohol dehydrogenase and cytochrome P450 dependent pathway and subsequent aldehyde dehydrogenase dependent metabolism of ethanol to acetaldehyde and acetale.6,7
Many toxic effects of alcohol also appear to be mediated through imbalances in micronutrients leading to decrease in antioxidative defence and disturbances in function of cells. Levels of many nutrients and antioxidants, including magnesium and selenium are lower in alcoholics.8–11
Magnesium is an important cofactor in more than 300 enzymatic reactions, fourth most abundant extracellular cation and the second in intracellular fluid. Mg participates in metabolisms of lipids, proteins, carbohydrates and nucleic acids. It is also involved in adenosine triphosphate (ATP) synthesis; moreover, ATP requires magnesium in order to be stable. That is why this element is essential for cells to produce, store and use energy. Decrease of cellular magnesium level affects negatively on function of mitochondria and energy production. 9
Chronic intake of alcohol develops alcoholism, decreasing the level of magnesium in the body. Alcohol abusers are frequently malnourished and additionally lose intestinal contents of Mg through vomiting. Moreover, ethanol inhibits the antidiuretic hormone (ADH) and acts as an Mg diuretic, causing a prompt, vigorous increase in the urinary excretion of this element along with other electrolytes. Under physiological conditions, magnesium modulates the production and release of anti-inflammatory cytokines positively. Induced by ethanol, decreasing of cellular Mg2+ causes the release of proinflammatory cytokines. Inflammation plays a key role in the progression of alcoholic liver disease. Moreover, loss of hepatic Mg2+ by alcohol administration is associated with an increased collagen deposition and may spell consequence for the development of hepatic cirrhosis.9,11
Mg treatment abnormally decreases high activities of gamma-glutamyltransferase (GGT), aspartate aminotransferase (AST) and alanine aminotransferase (ALT) in serum of alcoholic patients and may help to restore liver function.12,13 Study of Hans et al. 14 has demonstrated that supplementation of magnesium restores hepatic activity of glutathione S-transferase (GST) and superoxide dismutase (SOD), serum levels of vitamin C and total thiols and decreases oxidative stress in diabetic rats treated with alloxan. Authors have concluded that magnesium is an important component of the antioxidant system and may be used as potential therapeutic agent to reduce the clinical diseases associated with increased oxidative stress.
Selenium is an essential micronutrient for humans and animals health. Deficiency of Se is believed to implicate in a many diseases for example: cardiomyopathy, enhanced cancer risk, disorder of liver, kidney and bone. Se functions in antioxidant defence as a component of glutathione peroxidase and other selenoproteins. 15 A number of investigations have revealed that selenium alone and in combination with other antioxidans (vitamin C, vitamin E and herbal) can decrease alcohol-induced hyperlipidemia and injury of tissues.16–18 The protective effects of selenium seem to be primarily associated with its presence in the glutathione peroxidase, which are known to protect the cells from damage by oxygen radicals. Supplementation with selenium makes tissues less susceptible to be injured by endogenously and exogenously generated oxidants. 19
Ethanol consumption induces changes in lipid homeostasis, but this influence is complex and results obtained by different authors are incoherent. Studies show that effects are depended on many factors such as: the kind of the alcohol, drinking pattern, quantity of consumed ethanol, age and diet.20–23
In our study, we used young rats at the threshold of adulthood (aged 6–7 weeks) in order to create an animal model of liver damage caused by chronic and relatively heavy alcohol consumption (four times a week for 3 months). The aim of the study was to determine whether drinking water supplementation by magnesium and selenium (in combination or separately) significantly enhanced the antioxidant defences and reduced alcohol-induced oxidative stress, contributed to normalization of liver function and lipid metabolism and both elements.
The purpose of study was also assessment whether selenium and magnesium act in synergistic way on the intensification of antioxidative defence mechanisms, alleviation of oxidative damages, normalization of liver function and changes in serum cholesterol level.
Methods
Chemicals
Ethyl alcohol (96%) was obtained from POCH (Gliwice, Poland). Sodium selenite Na2SeO3 and magnesium aspartate tetrahydrate was obtained from Sigma Chemical Co. (St. Louis, MO, USA) and Farmapol (Poland), respectively.
Animals
Forty male rats of Wistar strain (6–7 weeks old, initially weighing 160–200 g) were used in this study. Animals were bred and kept during experiments in Animal Laboratory of the Department of Pathomorphology, Wroclaw Medical University, Poland. The rats were housed in plastic cages, under standard laboratory conditions (room temperature 19–21°C, relative humidity 50–60%, in natural day/night cycle) and had free access to fluids and standard pellet diet for rodents (Labofeed B, Factory of Animal Feed, Kcynia, Poland).
Experimental design
Rats were randomly divided into 5 groups of 8 rats each: one represented the control (C) and the remaining four groups were treated with alcohol alone (Et) or intoxicated with alcohol and supplemented selenium (Et + Se), magnesium (Et + Mg) or selenium plus magnesium simultaneously (Et + Se + Mg). The protocol of the rats' treatment is given in Table 1.
The protocol of the rats treatment
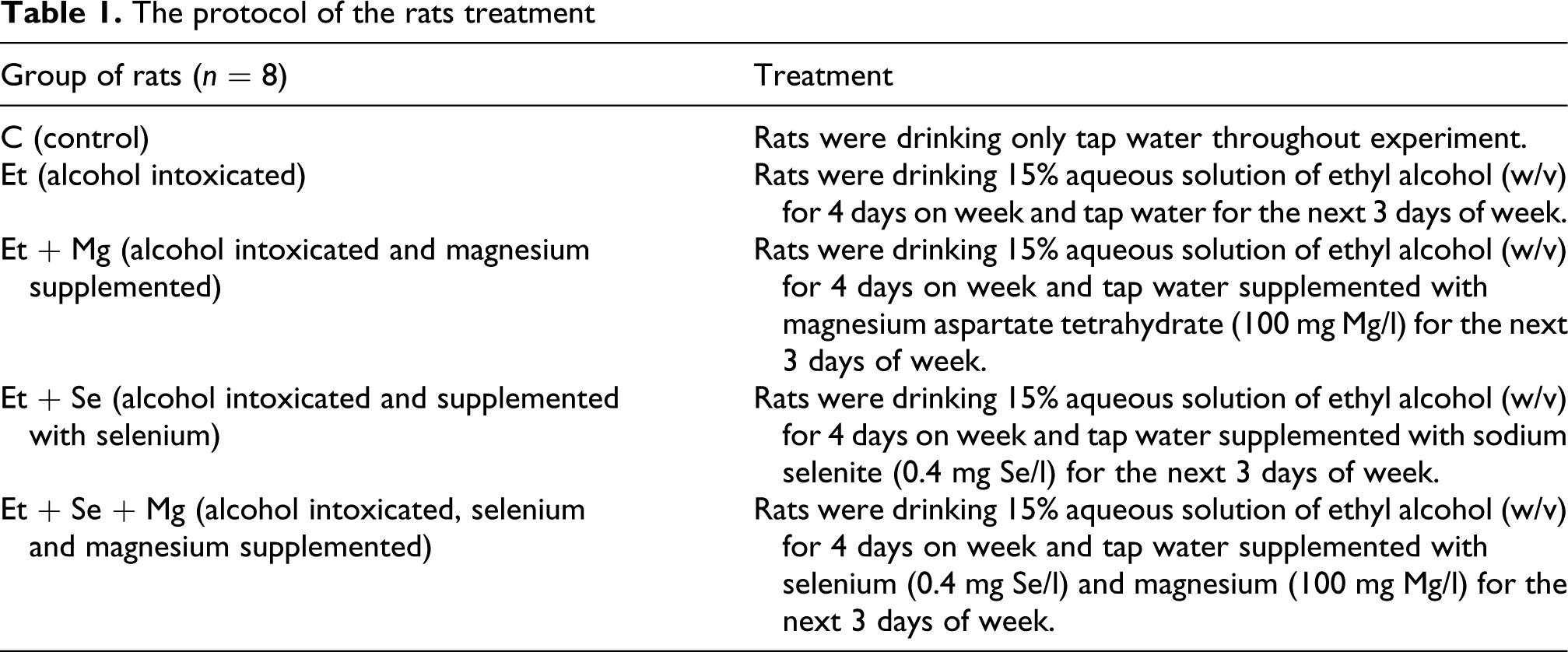
The experiment was carried out over 3 months. Throughout the treatment period, consumption of fluids was monitored twice a week, using the crude method of measuring the fluids remaining in the bottles as a measure of the previous time’s intake. Animals were weighed weekly. Average doses of the alcohol, Se and the Mg were converted to mg/kg body weight per week and per day and are given in Table 2.
Consumption of fluids, alcohol, magnesium and selenium by rats from control and experimental groups

Data are expressed as the mean (±SD) intake/per week/kg m.b. Bodyweight gain is expressed as mean (±SD)/rats. a, b, c, d significant differences between groups of rats, p < 0.05.
a - control group C significantly different from intoxicated with alcohol groups: Et, Et + Mg, Et + Se, Et + Se + Mg, p < 0.05.
* Rats were given alcohol four days a week, and Se and Mg only three days a week (during their abstinent periods).
Doses of alcohol
Rats received alcohol as 15% solution in drinking water (w/v), approximately 30 g ethanol/kg mass body, during four days of week (7.4 g/kg m.b/24 h) for 3 months. It may be tantamount to abuse of alcohol by people. The rate of ethanol oxidation is faster in rats than in humans and the animals need a higher dose to produce a comparable toxic effect. For example, the dose of 5 g EtOH/kg body weight/24 h is equivalent to consumption of about 0.7 l/day of 40% vodka by men, 24 although the dose of 11 g ethanol/kg m.b did not appear to result in ataxia or lethargy in the pregnant rats. 25 The liver samples of animals administered with 20% alcohol in the dose level was similar as in our experiment (7.9 g ethanol/kg body weight/24 h) for a period of 45 days showed microvesicular type and occasional macrovesicular type of fatty change. 26
Doses of selenium
The Recommended Dietary Allowance (RDA) for selenium is 55 µg/day for man and women. The tolerable upper intake level (UL) for adults is set at 400 µg/day based on selenosis as the adverse effect. 27 No observable adverse effect level (NOAEL) and lowest-observed-effect-level (LOAEL) for selenium are 15 µmol/kg/day and 23 µmol/kg/day, respectively. 28 However, therapeutic, antioxidant and antitumourigenic effects in experimental studies have been consistently associated with supranutritional intakes of Se, several times higher than the dose required to prevent clinical signs of Se deficiency, for anticancerogenic effect it is 100–500 μg Se/kg body weight/day (for rodents).29,30 Results obtained by Rai et al. 31 show that prophylactic administration of the selenium in drinking water (20 µM Se) in a dose of 70 µg Se/rat/day (above 200 µg/kg m.b) for 16 weeks, enhance the activity of superoxide dismutase in liver and serves as a useful protective therapy against damage induced by the hepatotoxin (thioacetamide). On the other hand, also lower doses of the selenium showed the antioxidative activity and alleviated damages in tissues of rats. Administration of selenium in the dose 10 μg Se/kg m.b/day for 60 days prevented alcohol-related injuries to the testis of rats. 32 Sodium selenite administered to rats in their drinking water (0.1 mg Se/l) at a dose of 7 μgSe/kg m.b/day, during a 30-day period, has been shown to be effective in increasing antioxidant enzyme activities 33 and in the dose 10–11.5 μg Se/kg m.b/day for 35 days partially alleviated Cd-induced structural damage in livers of rats. 34 Thus, the dosages of Se in the present experiment were similarly or lower than the dosages used previously in preventive studies because rats received about 19.4 μg Se/kg m.b/day in their drinking water (0.4 Se mg/l) but only in their abstinent period (three days a week).
Doses of magnesium
In the abstinent periods, rats were drinking water containing 100 mg Mg/l, which corresponds with the content of this element in some therapeutic mineral waters. 35 Absorption of Mg from the water is greater than from the food. The drinking of water with high concentration of Mg plays the essential role in the health prevention and is recommended also for persons who abuse alcohol.35–37 Amount of 300–400 mg Mg/person per day (approx. 4.6–6 mg/kg body weight/day) is recommended for human. 38 Three days a week rats were drinking the water with magnesium and so they got an approx. 2 mg Mg/kg of the body weight per day additionally (43–33% the recommended dose of the magnesium).
At the end of the study, rats were anaesthetized by an intramuscular ketaminum injection. Blood samples were collected via tubes after a cardiac puncture and then the animals were killed through the dislocation of cervical circles. All rats were killed during the abstinence period (approximately 72 h from the last day in which they were drinking alcohol).
Livers were removed for biochemical measurement and histological examination. Part of each tissue were cut into pieces and stored at –80°C until used and the remaining part of liver was fixed in 10% formalin solution for histopathological examination. Blood samples were centrifuged (3000 × g, 10 min) to get plasma.
The study was approved by the First Local Ethics Commission for Experiments on Animals in Wroclaw, Poland (No. of the permission – 47/2007).
Histopathological examination
Tissues fixed in the solution of the formalin were accurately rinsed in the water, then processed according to the standard, chemical procedures and embedded in liquid paraffin.
Sunken liver specimens were cut into sections with use of rotary microtome. The samples were transferred to the microscope slides and stained with hematoxylin and eosin (H&E). The histological analyses were performed under light microscope (Olympus BX41), at the enlargement 40, 100 and 400 times.
Estimation of oxidative stress and antioxidative defense
Malondialdehyde (MDA) and the ratio of GSH/GSSG (reduced/oxidized glutathione) were used as markers of oxidative stress in rat livers. The concentration of MDA was measured spectrophotometrically, according to instructions from Oxis Research kit (Bioxytech MDA−586 Assay No 21044). The livers were homogenized in TRIS-HCl buffer (pH = 7.4), containing butylated hydroxytoluene (BHT) as antioxidant. The homogenates (1/9, w/v) were centrifuged at 15000 × g for 10 min at 4°C. The supernatant was used for measurements. Reduced (GSH), oxidized (GSSG) glutathione concentration and GSSG/GSH ratio were determined using kit from Oxis Research (Bioxytech GSH/GSSH-412, No 21040). Liver samples were homogenized in 50 mmol/l potassium phosphate buffer (pH 7.5) containing 1 mmol/l EDTA and 1 mg/ml BSA. The homogenates were divided into two parts. One part was used for the measurement of total glutathione; and the second one was used to determine GSSG concentration. Thiol-scavenging reagent (1-methyl-2-vinylpyridinium trifluoromethanesulfonate, M2VP) was added to eliminate GSH. Next, 5% metaphosforic acid (MPA) was added to the both samples. Homogenates were centrifuged at 1000 × g for 10 min. Deproteinized supernatants were used. Glutathione peroxidase (c-GPx) activity was measured in a homogenate of tissues in cold buffer (50 mM TRIS-HCl, pH 7.5, containing 5 mM EDTA and 1 mM 2-mercaptoethanol), using Oxis Research kit (Bioxytech GPx-340, No 21017). The homogenates (1/9, w/v) were centrifuged at 10,000 × g for 10 min at 4°C. To assay GPx supernatant fluid containing the enzyme was added to a solution containing glutathione, glutathione reductase and NADPH. The enzyme reaction is initiated by adding the substrate, tert-butyl hydroperoxide and the A340 is recorded. Activity of GPx is expressed as mU GPx/mg protein (AGPx = 1 mU/mg protein = 1 nmol oxidized NADPH/min/mg protein). Activity of total SOD was measured using Cayman Chemical Kit (No. 706002). Tissues were homogenized (1/9, w/v) in cold 20 mM HEPES buffer, pH 7.2, containing 1 mM EDTA, 210 mM mannitol and 70 mM sucrose. The homogenates were centrifuged at 1500 × g for 5 min at 4°C and supernatants were used for assay. One unit of SOD is defined as the amount of enzyme needed to exhibit 50% dismutation of the superoxide radical. Activity of catalase (CAT) was measured spectrophotometrically, according to instructions from Cayman Chemical Kit (No. 707002). Livers were homogenized (1/9, w/v) in cold buffer (50 mM potassium phosphate, pH 7.0, containing 1 mM EDTA). The homogenates were centrifuged at 10,000 × g for 15 min at 4°C and supernatants were used for assay. One unit of catalase is defined as the amount of enzyme that will cause the formation of 1 nmol of formaldehyde per 1 min, at 25°C. CAT activity was expressed as U CAT/mg protein.
Total antioxidant status (TAS) was assayed in serum of rats using Randox Lab kit (NX 2332). Measurements of GSH, GSSG, TAS, GPX, SOD and CAT were performed using plate reader (PowerWave XS, Bio-Tek Instruments). Protein was assayed using a commercial kit (Sigma Diagnostics, procedure No. P5656) according to the method of Lowry et al. 39 Bovine serum albumin was used as standard.
Serum biochemical parameters: liver function, serum lipid parameters and Mg, concentrations
Biochemical parameters of serum were determined in the Biochemical Laboratory of University Hospital (Wroclaw Medical University, Poland) using the Dimension RxLMax auto-analyzer (Siemens Healthcare Diagnostics, USA). Flex Reagent Cartridge (DF43A, DF41A, DF57) of Siemens Healthcare Diagnostics Inc. (Newark, DE, USA) was used for measurement: ALT, AST activities and magnesium concentration, respectively. Serum lipids including: total cholesterol, CHOL (DF27), high-density lipoprotein, HDL (DF47), and triglycerides, TRIG (DF69A), were also assessed using Flex Reagent Cartridges.
The Ritis index was calculated as the quotient of aspartate and alanine aminotransferase activity. 40

Statistical analysis
All data were presented as means ± SD. A statistical comparison between groups of rats was performed using a one-way analysis of the variance (ANOVA) followed by a post hoc tests (NIR, Duncan's). For determination essentiality of differences between the groups of data with heterogeneous variances Cochran-Cox test was used. When the data did not follow the Gaussian distribution, the non-parametric Mann−Whitney test was used. Pearson’s or Spearman’s (for non-parametric data) correlation analysis was conducted for the relationship between measured parameters.
Statistical analysis was performed using STATISTICA 8.0 (for Windows) computer software. Differences were considered statistically significant at p <0.05.
Results
Body weight gain, consumption of fluids, alcohol, magnesium and selenium
Body weights of animals gained during the experiment, consumption of fluids as well as alcohol, magnesium and selenium intake by rats from both control and experimental groups are shown in Table 2.
Administration of alcohol alone (Et) and with magnesium (Et + Mg) caused retardation in the body weight gain in rats, compared with controls. Contrary, the body gains in groups supplemented with selenium and selenium and magnesium simultaneously were comparable with controls. There were no significant differences in alcohol, magnesium and selenium intake with fluids among the experimental groups of rats.
Histopathological examination
Histopathological changes of livers are given in Table 3. Livers of controls showed a normal histological architecture of hepatic lobules. Alcohol treated rats (Et) showed some liver lesions including: degeneration of cytoplasm in hepatocytes with formation of vacuoles, mild micro- and macrocellular fatty changes, mild inflammatory infiltration of mononuclear cells and singles neutrophiles. Sporadic, focal zones of hepatic necrosis were also observed. Magnesium supplementation of rats during their abstinent periods alleviated histopathological changes in liver by diminution of cytoplasmic vacuolation, infiltration of inflammatory cells and disappearance of steatosis and focal hepatocellular necrosis.
Histopathological changes of liver from control (C), alcohol (Et) and alcohol + selenium (Et + Se) or/and magnesium treated rats (Et + Mg, Et + Se + Mg)

The liver sections obtained from animals supplemented with selenium (Et + Se) and selenium plus magnesium simultaneously (Et + Se + Mg) showed normal structural integrity without, cytoplasmic vacuolation, necrosis and fatty degeneration. Passive congestion of the tissue of livers (probably caused by sudden death of rats) were seen equally in both control and alcohol-treated rats.
Markers of oxidative stress and antioxidative defense
Concentration of MDA in livers, reduced and oxidized glutathione (GSH and GSSG) and GSH/GSSG ratio are presented in Table 4.
Concentration of malonylodialdehyde (MDA), reduced and oxidized glutathione (GSH and GSSG) and GSH/GSSG ratio in liver of rats from control (C), treated with ethanol alone (Et) and supplemented with Mg and/or Se (Et + Mg, Et + Se, Et + Se + Mg) groups

Mean (±SD). a, b, c-- significant differences between groups of rats, p < 0.05.
a - Intoxicated with alcohol groups: Et, Et + Mg, Et + Se, Et + Se + Mg significantly different from control group C, p < 0.05.
b-- Groups supplemented with Mg and/or Se (Et + Mg, Et + Se, Et + Se + Mg) significantly different from group intoxicated with alcohol alone Et , p < 0.05.
c-- Et + Mg, Et + Se groups significantly different from group Et + Se + Mg, p < 0.05.
The level of hepatic MDA increased significantly (by 24.3%) in ethanol-treated group (Et) in comparison with controls, p < 0.05. Elevations of MDA level in both selenium (Et + Se) alone and of selenium with magnesium-treated (Et + Mg + Se) rats were lower in comparison with ethanol-treated group (by 20.8% and 20.3% respectively; p < 0.05). However, no significant differences in MDA concentrations were observed between the group exposed to alcohol alone (Et) and the group exposed to alcohol with supplementation with magnesium (E + Mg) or between the controls (C) and experimental groups supplemented with selenium (E + Se) and/or magnesium (E + Mg, E + Mg + Se), p > 0.05.
There were no significant differences in the GSH concentration of livers between controls and alcohol-treated groups. However, drinking of alcohol alone (Et) tended to decrease the concentration of reduced glutathione in comparison with the control group and supplemented with Mg and/or Se groups, but only in the case of Et + Mg and Et + Se groups the differences were statistically significant, p > 0.05.
The results show that chronic drinking of alcohol (Et) or drinking of alcohol with magnesium supplementation (Et + Mg) significantly elevated the level of oxidized glutathione in rat livers (by 78.5% and 58% respectively, in comparison with the control group, p > 0.05). Contrary, in groups of rats supplemented with selenium or selenium plus magnesium, hepatic concentrations of GSSG were similar to this one of the controls.
The hepatic concentrations of GSSG were significantly elevated in groups of rats treated with selenium or magnesium separately than in the group supplemented with Mg and Se simultaneously, p > 0.05.
The ratio of GSH and GSSG was significantly lower in Et and Et + Mg groups than in C group, p > 0.05. The GSH/GSSG ratio in rats supplemented with selenium or selenium plus magnesium was substantially higher and it was comparable to the ratio of controls.
The highest GSH/GSSG ratio was found in livers of rats supplemented with magnesium and selenium simultaneously (Et + Mg + Se). The ratio of GSH and GSSG was significantly higher in livers of rats from Et + Mg + Se group in comparison with the rats treated with selenium or magnesium separately, p > 0.05.
Antioxidant enzymes in liver and serum total antioxiant status
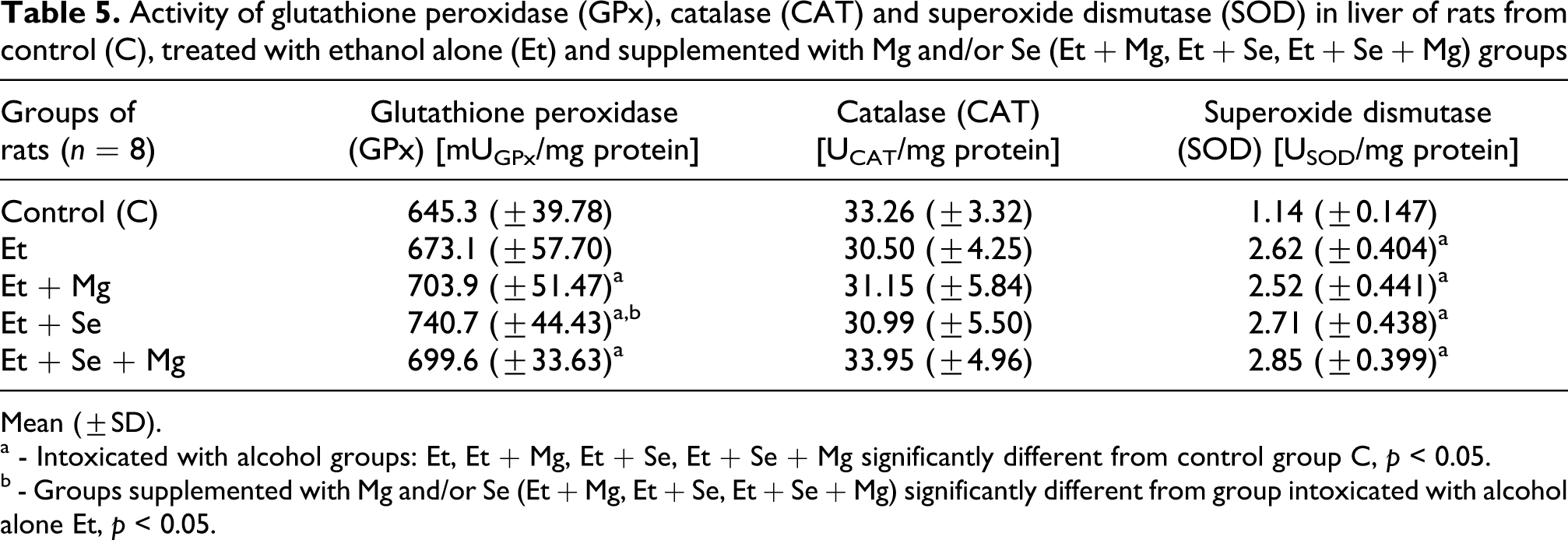
Influence of alcohol intoxication and supplementation with magnesium and/or selenium on hepatic activities of antioxidant enzymes are given in Table 5.
Activity of glutathione peroxidase (GPx), catalase (CAT) and superoxide dismutase (SOD) in liver of rats from control (C), treated with ethanol alone (Et) and supplemented with Mg and/or Se (Et + Mg, Et + Se, Et + Se + Mg) groups
Mean (±SD).
a - Intoxicated with alcohol groups: Et, Et + Mg, Et + Se, Et + Se + Mg significantly different from control group C, p < 0.05.
b - Groups supplemented with Mg and/or Se (Et + Mg, Et + Se, Et + Se + Mg) significantly different from group intoxicated with alcohol alone Et, p < 0.05.
The hepatic activities of SOD significantly increased as a result of alcohol administration alone as well as together with magnesium and/or selenium. Two days after the last consumption of the alcohol, activities of SOD in livers of rats from alcohol-treated groups (Et, Et + Mg, Et + Se, Et + Se + Mg) were about twice as high as in livers from control rats (C), p < 0.05. There were no significant differences in SOD activities in livers among the other experimental groups of rats.
The hepatic activity of glutathione peroxidise in comparison with the same of control group tended to increase due to drinking of alcohol during experiment. In the case of groups supplemented with magnesium, selenium and magnesium and selenium simultaneously (Et + Mg, Et + Se, Et + Se + Mg) these differences were statistically significant and amounted to 4.3%, 14.8% and 8.4% respectively, p > 0.05. The highest activity of glutathione peroxidise was observed in rats intoxicated with alcohol and supplemented with selenium (Et + Se), but this elevation was statistically significant only when compared with the rats treated with alcohol alone (Et).
Two days after the last consumption of the alcohol there were no significant differences in the hepatic catalase activities between control and alcohol-treated groups.
Total antioxidant status
TAS is a very important parameter because it indicates a serum’s ability to cope with oxidative stress. The results of measuring the levels of TAS in serum of rats are shown in Figure 1.

Influence of alcohol intoxication alone and with magnesium and/or selenium supplementation on total antioxidant status (mmol/l) in serum of rats. a, b, c, d - significant differences between groups of rats, p < 0.05; a - Intoxicated with alcohol groups: Et, Et + Mg, Et + Se, Et + Se + Mg significantly different from control group C, p < 0.05. b - Groups supplemented with Mg and/or Se (Et + Mg, Et + Se, Et + Se + Mg) significantly different from group intoxicated with alcohol alone Et , p < 0.05. c - Et + Mg, Et + Se groups significantly different from group Et + Se + Mg, p < 0.05. d - Et + Mg significantly different from Et + Se, p < 0.05.
A significant decline (by 27%) of the TAS was observed only in rats treated with alcohol alone (Et) in comparison with the control group, p < 0.05.
Supplementation by magnesium and/or selenium (Et + Mg, Et + Se, Et + Se + Mg) elevated total antioxidant status in serum of rats intoxicated with alcohol. The rats from Et + Mg, Et + Se and Et + Se + Mg groups exhibited a significantly higher level of TAS than rats from Et group, by 26%, 73% and 39% respectively, p < 0.05. The highest TAS level was observed in the animals administrated with alcohol and supplemented with selenium alone (Et + Se), p < 0.05.
Alanine aminotransferase, aspartate aminotransferase activities and De Ritis index
The alterations of the liver function under the chronic drinking of alcohol and the administration of Se and Mg were estimated by determining the activities of ALT, AST in serum of rats and assessment of the De Ritis index. Results are shown in Table 6(mean ± SD).
Activity of glutathione peroxidase (GPx), catalase (CAT) and superoxide dismutase (SOD) in liver of rats from control (C), treated with ethanol alone (Et) and supplemented with Mg and/or Se (Et + Mg, Et + Se, Et + Se + Mg) groups

Mean (±SD).
a, b, c, d - significant differences between groups of rats, p < 0.05.
a - Intoxicated with alcohol groups: Et, Et + Mg, Et + Se, Et + Se + Mg significantly different from control group C, p < 0.05.
b - Groups supplemented with Mg and/or Se (Et + Mg, Et + Se, Et + Se + Mg) significantly different from group intoxicated with alcohol alone Et , p < 0.05.
c - Et + Mg, Et + Se groups significantly different from group Et + Se + Mg, p < 0.05.
d - Et + Mg significantly different from Et + Se, p < 0.05.
Concentration of total cholesterol in serum significantly decreased in Et and Et + Mg groups when compared with the control animals (C), by 12.4% and 16% respectively (p < 0.05). This elevation was statistically significant only in two groups of animals: one treated with alcohol alone (Et) and second one supplemented with magnesium and selenium simultaneously (Et + Se + Mg), by 40% and 31% respectively, p < 0.05. There were no significant differences in AST activities in serum among the other groups of rats. Exposure to alcohol substantially increased ALT activity in serum of rats supplemented with Se or Se plus Mg in comparison to controls, by 28% and 39% respectively, p < 0.05 and in comparison to rats supplemented by magnesium alone by 27% and 37% (p < 0.005) respectively. The value of the De Ritis index increased as a result of alcohol administration in Et and Et + Mg groups in comparison with the control group, by 17% and 20% respectively, p < 0.05. Contrary, administration of the Se and Se + Mg in drinking water (during of abstinence periods) decreased De Ritis index to control value. In rats exposed to alcohol and supplemented with selenium (Et + Se) or selenium and magnesium (Et + Se + Mg) simultaneously De Ritis index was significantly lower in comparison with the group treated with alcohol alone (Et) by 20.4% and 19%, respectively and with group magnesium supplemented (Et + Mg) alone by 22% and 21%, respectively, p < 0.05.
Serum lipid levels
Mean (±SD) of serum lipids including: CHOL, HDL and TRIG in all groups of rats are presented in Figure 2.
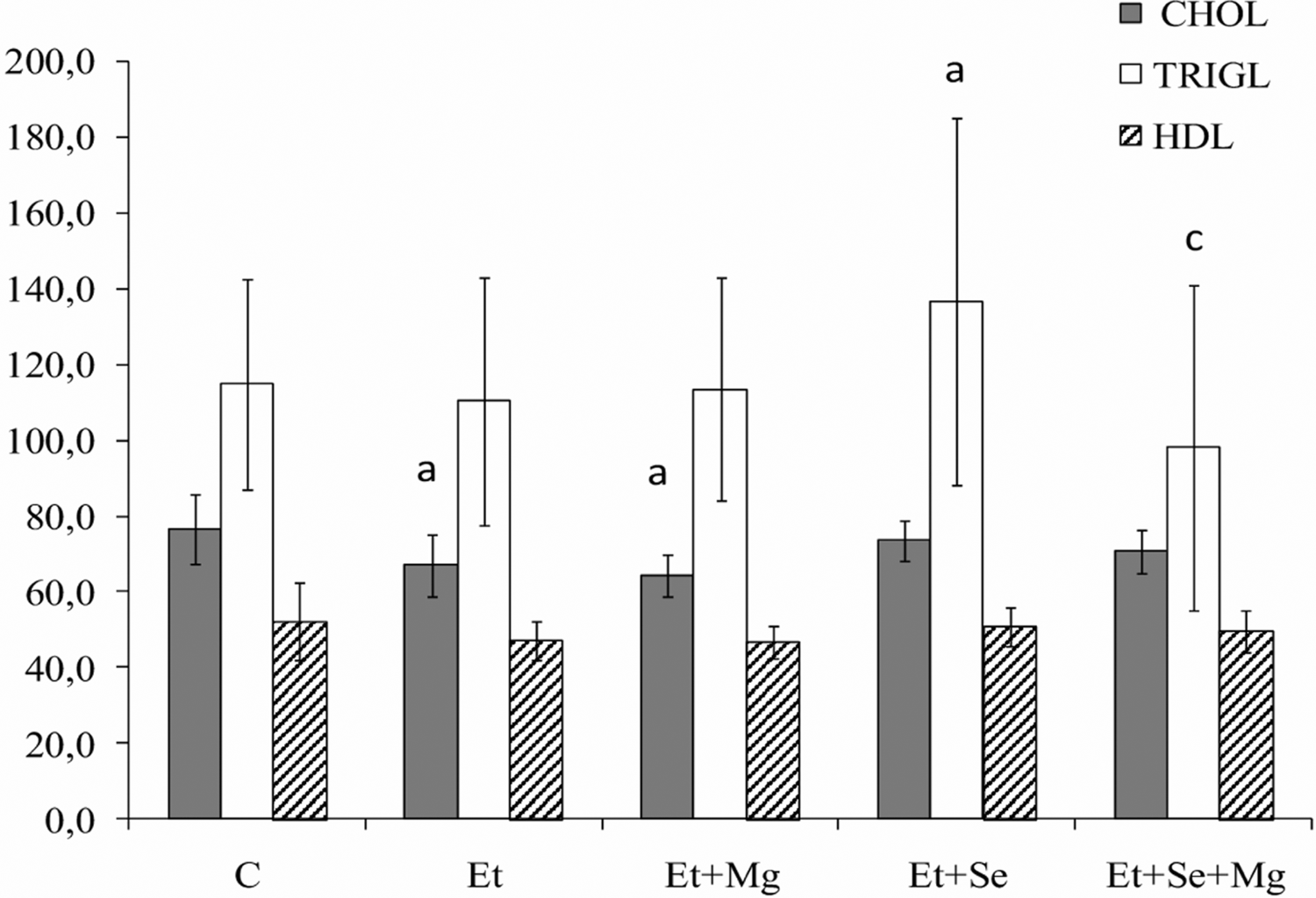
Influence of alcohol intoxication alone and with magnesium and/or selenium supplementation on lipid levels (mg/dl) in serum of rats. a - Intoxicated with alcohol groups: Et, Et + Mg, Et + Se, Et + Se + Mg significantly different from control group C, p < 0.05. c - Et + Se + Mg group significantly different from groups Et + Mg, and Et + Se, p < 0.05.
Concentration of total cholesterol in serum significantly decreased in Et and Et + Mg groups when compared with the control animals (C), by 12.4% and 16% respectively (p < 0.05). In the groups supplemented with selenium and selenium plus magnesium simultaneously, concentration of total cholesterol in serum was similar to the control group. In rats exposed to alcohol and supplemented with Se alone the concentration of triglycerides was significantly higher in comparison with the group supplemented with selenium plus magnesium. There were no significant differences in concentration of triglycerides among the other groups of rats.
Serum concentration of HDL and ratio total cholesterol/HDL were not affected by alcohol administration alone and with magnesium and/or selenium (Figure 3).

Influence of alcohol intoxication alone and with magnesium and/or selenium supplementation on total cholesterol/HDL index.
Serum magnesium concentration
Concentration of magnesium in serum of rats is presented in Figure 4.

Influence of alcohol intoxication alone and with magnesium and/or selenium supplementation on concentration of magnesium in serum (mg/dl). a - Intoxicated with alcohol groups: Et, Et+Mg, Et+Se, Et+Se+Mg significantly different from control group C, p < 0,05. b - Groups supplemented with Mg and/or Se (Et+Mg, Et+Se, Et+Se+Mg) significantly different from group intoxicated with alcohol alone Et, p < 0,05.
Chronic drinking of alcohol (Et group) caused slight but significant decrease of serum concentration of magnesium in comparison with the control group (by 10%, p <0.05). The serum concentration of Mg in animals supplement with Mg and Mg + Se was similar to the control group. In rats supplemented with selenium alone (Et + Se) magnesium content in serum was significantly higher than in Et group.
Correlation analysis
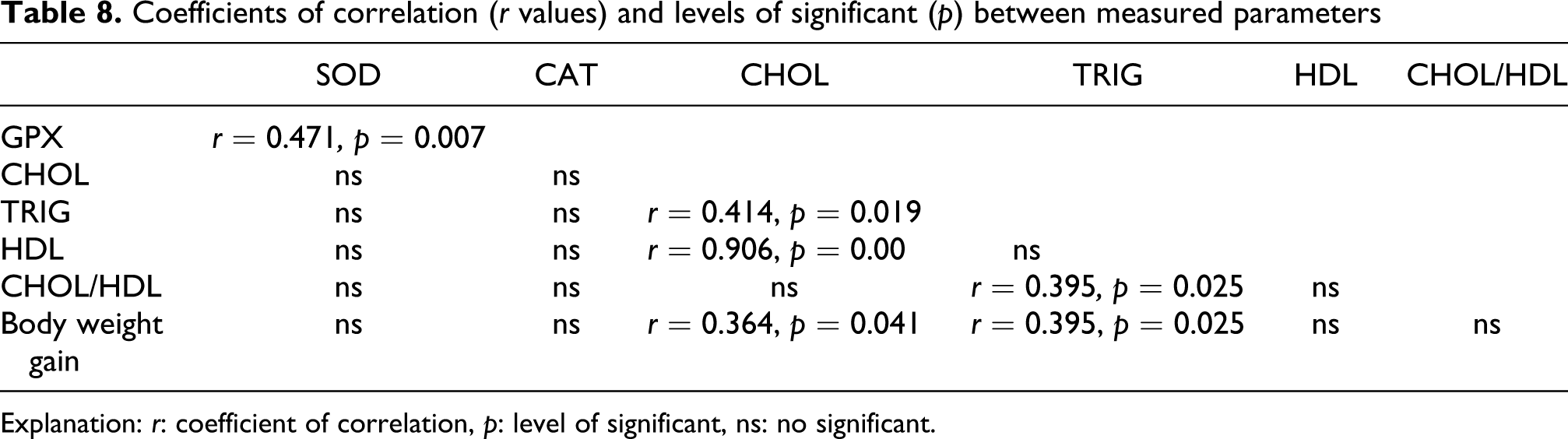
Correlation coefficients (r values) and levels of significance (p) between measured parameters are shown in Tables 7 and 8. Correlation analysis identified a strong relationship between increase of the De Ritis index (which was used to assess the extent of liver damage) and the depletion of TAS in serum (r = –0.495, p = 0.00158) and decline of rats' body weight gain (r = –0.414, p = 0.012). Moreover, negative correlations were observed between the TAS and the hepatic concentration of MDA (r = –0.336, p = 0.0392) and positive relationships between the TAS and the hepatic content of reduced glutathione (r = 0.485, p = 0.00363), GSH/GSSG ratio (r = 0.457, p = 0.0065) also the glutathione peroxidise activity (r = 0.334, p = 0.040). The elevation of AST activity in the serum correlated with the rise of ALT (r = 0.726, p = 0.0000) and De Ritis index (r = 0.396, p = 0.0136) and also with the fall in serum concentration of magnesium (r = –0.339, p = 0.0403). Concentration of MDA in liver was observed to be positively correlated with hepatic oxidized glutathione (r = 0.444, p = 0.01) and negatively with GSH/GSSG ratio (r = –0.453, p = 0.008) and serum concentration of total cholesterol (r = –0.416, p = 0.018).
Coefficients of correlation (r values) and levels of significant (p) between measured parameters

Explanation: r: coefficient of correlation, p: level of significant, ns: no significant.
Coefficients of correlation (r values) and levels of significant (p) between measured parameters
Explanation: r: coefficient of correlation, p: level of significant, ns: no significant.
Serum total cholesterol concentration was found to be positively correlated with triglycerides (r = 0.414, p = 0.019), HDL (r = 0.906, p = 0.000000) and body weight gain of rats (r = 0.364, p = 0.041). Positive correlations were observed between the serum content of triglycerides and body weight gain of rats (r = 0.395, p = 0.025) and total cholesterol/HDL index (r = 0.425, p = 0.015). Hepatic CAT activity and the GSH/GSSG ratio was found to show a significant, positive correlation (r = 0.342, p = 0.0479). Moreover, strong positive correlation was found between the activity of superoxide dismutase and glutathione peroxidase in liver (r = 0.471, p = 0.007).
Discussion
Both animals and human studies show that alcohol-induced oxidative stress is further intensified by malnutrition found very often in alcoholics, which resulted in antioxidant deficiency and this plays a key role in liver damage.16,41,42 As expected, chronic intoxication of rats with ethanol caused liver damage, as indicated by changes in the histological picture and a significant increase in the activity of AST and ALT in serum and growth in the De Ritis ratio. Hepatocyte damage increases permeability of cell membranes or causes their complete destruction. The damaged liver tissue releases enzymes, including AST and ALT, and the measurement of their activity in serum is used to assess damage of that organ. 43
A characteristic feature of alcoholic liver disease is a distinguished predominance of AST over ALT. Abstinence periods are often characterised by elevated AST values and normal ALT values. Such enzyme pattern in alcohol-induced liver damage is a result of release of mitochondrial AST, whose half-life period in blood is significantly longer than in the case of cytoplasmic transaminases. Dimensionless De Ritis ratio (AST/ALT) is considered to be a useful diagnostic element in detection of alcoholic liver damage. An increase in De Ritis ratio to the value close to 2 may already suggest alcoholic liver damage, while >2 is an almost certain criterion of such damage.43,44
Histopathological examination confirmed the results obtained from measurements of the activity of liver enzymes. Chronic alcohol consumption by rats caused changes in hepatic parenchyma and the hepatocyte structure. Microscopic examination revealed small to relatively abundant areas of microvesicular and macrovesicular steatosis of hepatocytes, small infiltrations of mononuclear inflammatory cells, cytoplasmic vacuolation and sporadically occurring focal necrosis of hepatocytes. Administration of magnesium in drinking water significantly decreased the activity of both AST and ALT in serum. Despite the still elevated De Ritis ratio, observations of histopathological changes demonstrated inhibitory effect of magnesium on liver damage. Hepatocytes of rats supplemented with magnesium showed only areas with cytoplasmic vacuolation, without steatosis, infiltrations of inflammatory cells, or areas with focal necrosis. However, drinking of water enriched with selenium or both Se and Mg turned out to be much more effective in preventing degenerative liver changes. The De Ritis ratio in the Et + Se and Et + Se + Mg groups was similar to the control group, the histological picture of liver slices showed no degenerative changes seen in animals drinking only alcohol (Et) or supplemented only with magnesium (Et + Mg), the parenchymal architecture of that organ was normal and similar to the architecture observed in control animals. High concentration of MDA, elevated even during the abstinence, and lower ratio of GSH/GSSG in the livers of animals drinking alcohol only as compared with the control group confirm the widely accepted view of the significant role of oxidative stress in alcoholic liver damage. Likewise, Pemberton et al. 41 in their studies observed that in the group of people with alcoholic liver disease, the MDA level was significantly higher than in healthy subjects.
Administration of magnesium, selenium and selenium together with magnesium significantly enhanced antioxidant defences in rats, as indicated by a significant increase of the TAS in serum, GPX activity in the liver and the GSH/GSSG ratio. Both TAS and GPX activity were highest in rats supplemented with selenium alone. Similar results were obtained by Sivaram et al., 16 who conducted an experiment on guinea pigs. In the group receiving ethanol together with selenium (at a dose of 0.5 mg Se/kg of body weight) and ascorbic acid, GPx activity was significantly greater and the amount of the formed MDA was significantly smaller than the values observed in guinea pigs drinking alcohol alone.
The results indicate that administration of selenium, selenium together with magnesium and Mg alone, but to a lower degree, increases the antioxidant status in the organisms of rats, contributing to mitigation of the effects of oxidative stress and inhibition of the development of degenerative liver changes. Increased activity of SOD in the livers of animals from all groups drinking alcohol in comparison with the control group indicates increased production of superoxide anion radical (O2·–) even during the period of abstinence.
Superoxide dismutase neutralizes O2·– for less toxic compounds (e.g. H2O2), which are then removed by catalase or glutathione peroxidise. 45 However, in the case of animals drinking alcohol only, no significant increases of the activities of those enzymes were observed, therefore that line of antioxidant defence seemed to be not efficient enough to prevent damage. In contrast, in the case of rats supplemented with selenium, selenium plus magnesium, or, to less degree, with magnesium only, the observed increment of the glutathione peroxidase activity strengthened and improved antioxidant defence, significantly decreasing oxidative stress and damage to hepatic parenchyma.
The above studies suggest that alcohol, causing increased generation of free radicals, also mobilises the antioxidant defence system. However, such defence often proves to be insufficient, which leads to oxidative damage of lipid membranes and other cellular structures. Also, Lutnicki et al. 46 observed the significant increase of the activities of antioxidant enzymes (GPX and SOD) after rats were exposed to alcohol, and then the fall in the activity of GPX and intensification of oxidative processes in liver. The level of MDA increased and was the highest at the moment of the lowest activity of GPX, with elapsed time after the alcohol application. Our results show that supplementation with selenium and/or magnesium activates antioxidant enzymes, intensifies antioxidative processes and strengthens the defence, contributing to mitigation of the oxidative stress in the liver and reduction of alcohol-induced damage. Administration of selenium or selenium together with magnesium turned out to be more effective in intensification of antioxidant defences and inhibition of the development of liver disease than supplementation with magnesium alone. However, Hans et al., 14 who for 6 weeks supplemented rats with magnesium in drinking water at a concentration higher to the one used in our study (900 mg/l), observed significant impact of that element on increased activity of glutathione transferase and SOD in the liver as well as concentrations of vitamin C and the SH groups in serum, and consequently, a lowering of the alloxan-induced oxidative stress in the livers of rats with diabetes. The results also indicate that high doses of the magnesium (higher than used in our experiment) can enhance antioxidative defence in the liver and similarly as the selenium cause the normalization of the function of this organ.
Selenium alone had a stronger effect on the increasing of antioxidant status (TAS) in serum and GPx activity than combined supplementation of selenium and magnesium. On the other hand, the higher GSH/GSSG ratio in the group supplemented by selenium plus magnesium simultaneously indicates that reducing of oxidative damage of this group was more pronounced than the group receiving selenium only. Despite this, administration of both selenium alone and selenium plus magnesium decreased concentration of products of lipid peroxidation (MDA) to a comparable extent.
Such result may point to an independent effect of selenium and magnesium in inhibition of alcohol-induced degenerative liver changes. Lack of a clear synergetic effect between magnesium and selenium can also be explained by assuming that the selenium dose used in the study was sufficient to inhibit alcohol-induced malfunctioning of the liver.
We observed that drinking of alcohol at a young age is associated with less gain of weight. The lowest weight gain were observed in the groups of animals drinking alcohol alone (Et) or drinking alcohol and supplemented with magnesium alone (Et + Mg).The results obtained by other authors47,48 also confirm that chronic alcohol intake inhibits normal development of the organism and promotes malnutrition. This phenomenon occurs not only in experimental animals but is also common in people using alcohol. Alcohol consumption causes a shortage of important nutrients, including the compounds involved in oxidative defence of the body, e.g. retinol, α-tocopherol, vitamin C and minerals such as Zn, Se, Mg and Cu.8,19 Supplementation with selenium or selenium together with magnesium resulted in the weight gain of the animals drinking alcohol similar to the weight gain in control animals.
Alcohol consumption affects lipid metabolism; however, this effect is complex and the results obtained by different authors are inconsistent and include an increase in HDL, a drop in total cholesterol and LDL20,21 and an increase in the LDL cholesterol and triglycerides.22,23 The effect of alcohol on lipid metabolism depends on many factors, such as the type of consumed alcoholic beverage, drinking pattern, duration of alcohol abuse, age and the eating habits.20–23 In our study, chronic alcohol consumption by rats resulted in a decrease in total cholesterol in the serum of animals drinking alcohol only (Et) and supplemented with magnesium alone (Et + Mg) and the absence of changes in the concentration of HDL and triglycerides as well as the ratio of total cholesterol to HDL. The administration of selenium alone as well as selenium together with magnesium raised the total cholesterol levels to the control value. In addition, the group supplemented with selenium alone showed a significant increase in the concentration of triglycerides. The concentration of both total cholesterol and triglycerides correlated positively with an increase in animal body weight so it seems that a decrease in the concentration of total cholesterol was mainly associated with worse nutritional status of the animals in the groups Et and Et + Mg compared to the animals additionally supplemented with selenium.
Although selenium typically demonstrates hypolipemic action, 49 in large doses it can have the opposite effect and raise the concentration of lipids in serum. Bleys et al. 50 in studies on a representative US population observed a strong correlation between high selenium concentration in serum and high concentration of total cholesterol, HDL, triglycerides and apolipoproteins: apo A and apo B; therefore, according to the authors the supplements containing selenium should be screened for their impact on lipid metabolism.
Also Nolan et al., 48 in the study of rats with fatty liver, which developed as a result of consumption of a high-fat diet together with alcohol, observed a significantly higher fat content in the livers of animals that were additionally supplemented with a high dose of selenium (0.09 mg/kg body weight/day) than in the group receiving only a high-fat diet and alcohol. In our studies, the administration of magnesium together with selenium reduced concentration of triglycerides to the control value.
Supplementation of Se or selenium plus magnesium simultaneously significantly enhances antioxidant defences and is more effective against alcohol-induced oxidative stress, disturbance of liver function and cholesterol metabolism than the supplementation of magnesium alone. Moreover, administration of Se plus Mg simultaneously normalizes all measured parameters of lipids in serum of rats. Lack of a clear synergetic, therapeutic effect on liver between magnesium and selenium can be explained by assuming that the selenium dose used in the study was sufficient to inhibit alcohol-induced malfunctioning of this tissue.
Present results confirm the significant role of the oxidative stress in the development of alcohol-induced changes in the liver and the effectiveness of supplementation of antioxidants such as Mg and Se, and especially the selenium in the appeasement of these damages. Co-administration of the selenium and the magnesium normalizes disturbances of weight gain and lipids metabolism in serum. The supplementation with these nutrients can be put-upon in the prophylaxis of the liver disease at alcoholics and persons abusing of alcohol.
Footnotes
This study was supported by the fund of statutory research of Wroclaw Medical University. Project No: ST-31.