
Abstract
Spores of coprophilous fungi (CFS) are widely used in paleoecology as indicators of past herbivore presence, but the processes governing their deposition, retention, and preservation remain insufficiently understood. While most studies rely on sedimentary archives, few have examined how different modern substrates vary in their ability to capture dung fungal spores. This study compares CFS composition and abundance in moss polsters, Tauber-type traps, and dung samples from four ungulate species in European lowland forests, aiming to assess their effectiveness as modern analogs for paleoecological reconstructions. Moss samples exhibited the highest taxonomic diversity of CFS, suggesting they act as passive spore traps, accumulating propagules over extended periods. Dung samples contained the highest concentration of CFS, providing a baseline for understanding fungal community composition in relation to direct spore production sites. Tauber-type traps showed the lowest CFS proportion, highlighting the limited role of long-distance airborne dispersal for most taxa. Despite these differences, statistical analyses revealed weak and non-significant correlations between CFS abundance and environmental variables (coarse woody debris, wetness, openness, ungulate pressure, and forest management intensity), suggesting that factors such as dung decomposition rates and stochastic deposition processes may play a greater role. These findings indicate that CFS assemblages are shaped more by substrate-specific properties and microhabitat variation than by broad environmental gradients. From a paleoecological perspective, the presence of CFS in moss-derived archives may reflect cumulative, long-term deposition rather than contemporaneous herbivore presence. This study emphasizes the need for caution when interpreting CFS signals and supports the integration of multiple modern analogues to refine reconstructions of past herbivore dynamics.
Keywords
Introduction
The presence and activities of herbivorous mammals, particularly large herbivores, play a critical role in shaping ecosystems. Through grazing, browsing, trampling, seed dispersal, and the return of nutrients to the soil, animals influence vegetation structure, plant community composition, and soil dynamics (Forbes et al., 2019; Gill and Beardall, 2001; Heggenes et al., 2017; Kapás et al., 2020; van der Waal et al., 2011). These interactions have cascading effects on ecosystem functioning, biodiversity, and successional processes, underscoring the significance of herbivores as ecological engineers. The impact of herbivores is not limited to modern ecosystems. Still, it extends deep into the past, leaving traces in the fossil record that provide invaluable insights into historical ecosystems and their dynamics.
Coprophilous fungal spores (hereafter CFS) are among the most informative paleoecological proxies for reconstructing the presence of herbivorous mammals. They are produced by fungi that grow on animal dung (Lee et al., 2022; van Asperen et al., 2021). CFS directly reflect herbivore presence and may provide insights into their relative abundance, as their life cycle depends on the availability of dung. As a result, CFS offer a valuable complement to pollen analysis in paleoecological studies. While pollen data primarily inform on vegetation composition and climate, CFS contribute to understanding ecological processes related to herbivore presence, such as grazing pressure or spatial patterns of dung deposition (Gill et al., 2013; Johnson et al., 2015; Mazier et al., 2006).
In the Białowieża Forest, where this study was conducted, wild boar (Sus scrofa) was the most frequently recorded ungulate species during 2012–2014, followed by red deer (Cervus elaphus), European bison (Bison bonasus), roe deer (Capreolus capreolus), and moose (Alces alces), based on multi-year camera trap data (Bubnicki et al., 2019). Although exact population densities are not available for all species, long-term monitoring enables the estimation of their relative abundance, which is essential for interpreting dung fungal spore data in this ecological context.
While most studies on CFS have relied on sedimentary archives, fewer have examined how modern substrates, such as moss polsters, Tauber-type traps, and direct dung samples, vary in their ability to retain and preserve these spores. Understanding these differences is crucial for improving interpretations of the fossil record. Since fungal spores differ in rates of degradation, transport, and retention depending on the substrate, modern comparative studies provide an essential framework for evaluating the reliability of these proxies in both ecological and paleoecological contexts (Sundberg, 2013; van Asperen et al., 2016). By identifying patterns of spore deposition in modern environments, we can refine our ability to reconstruct past herbivore-vegetation interactions.
In recent decades, CFS have gained increasing recognition in paleoecology as reliable indicators of past herbivore populations (Lee et al., 2022; Perrotti et al., 2022; van Asperen et al., 2021). They have been used to trace shifts in herbivore abundance over time (Gill et al., 2009; Robinson et al., 2005), infer grazing impacts on vegetation (Baker et al., 2013; Gill et al., 2013), and even detect extinction events of megafauna during the Late Pleistocene and Holocene (Burney et al., 2003; Gill et al., 2009; Lee et al., 2022). However, despite their utility, the interpretation of these proxies is not without challenges. The deposition and preservation of fungal spores are influenced by a range of environmental factors, including moisture levels, substrate availability, and dispersal mechanisms, as well as taphonomic processes that can alter the spore record (Bradshaw et al., 2003; Raper and Bush, 2009; van Asperen et al., 2016).
The Białowieża Forest (Figure 1), a uniquely preserved lowland temperate forest, offers an exceptional setting for studying the relationship between CFS and herbivore populations. This forest hosts five native ungulate species: red deer (Cervus elaphus), roe deer (Capreolus capreolus), European bison (Bison bonasus), moose (Alces alces), and wild boar (Sus scrofa), in a relatively undisturbed setting; however, due to a recent population collapse (Morelle et al., 2020), wild boar dung was not available for analysis. These species exert varying levels of grazing and browsing pressure across different forest habitats, creating spatial heterogeneity in nutrient deposition and vegetation dynamics (Hardalau et al., 2024; Kupferschmid et al., 2020; Samojlik and Kuijper, 2013). Because different ungulate species interact with their environment in unique ways, including their different diet preferences, their dung may harbor distinct fungal communities, offering potential insight into host-specific fungal assemblages. The diverse habitats of the Białowieża Forest and its long-standing herbivore–vegetation dynamics make it an ideal site for exploring modern analogs relevant to paleoecological reconstructions. Through the analysis of fungal spores in moss polsters, Tauber-type traps, and herbivore dung, we investigate how spore assemblages vary across sample types and environmental contexts. This approach not only aids in interpreting fossil records but also assesses the ecological representativeness of modern spore deposition patterns.
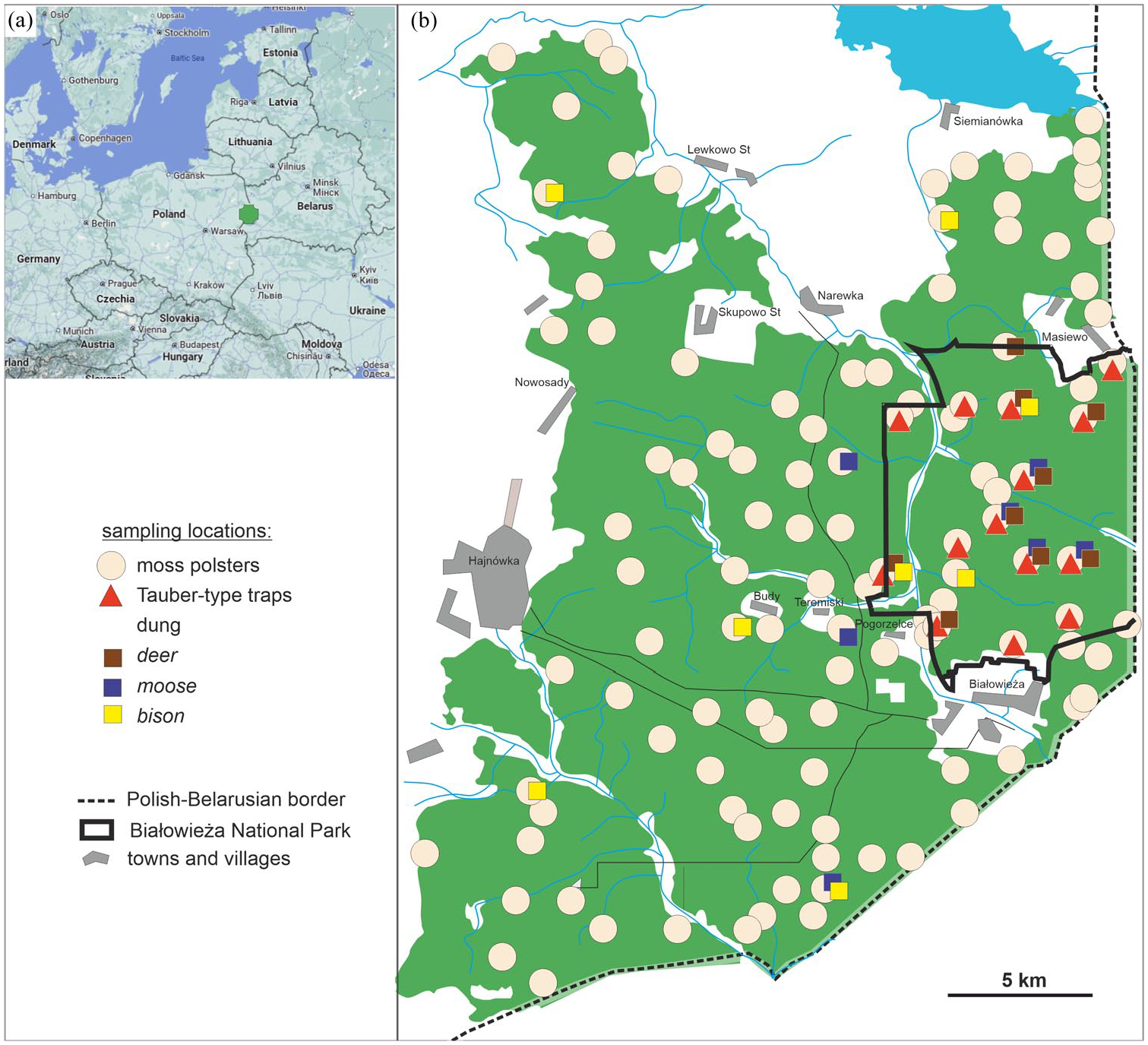
Location of the Białowieża Forest (BF) within Europe (a) and the Polish part of the BF, with the locations of sampling sites: moss polsters, Tauber-type traps, and dung samples from red/roe deer, Eurasian moose, and European bison (b).
This study builds on earlier research conducted within the same project framework (Zimny, 2024a; Zimny et al., 2024, 2025), which explored non-pollen palynomorphs, including fungal spores across different forest substrates and habitats. Here, we focus specifically on CFS and their deposition as recorded in moss polsters, Tauber-type traps, and dung samples. We aim to evaluate the potential of CFS as indicators of herbivorous ungulate presence in the context of European lowland temperate forest. Specifically, we address the following objectives:
Compare the composition and abundance of CFS across three sample types: moss polsters, Tauber-type traps, and dung from four native ungulate species, to assess how effectively each captures fungal assemblages.
Test whether CFS abundance correlates with environmental variables, including habitat wetness, canopy openness, volume of dead wood, ungulate pressure, and intensity of forest management, to identify the main drivers of spore deposition.
Investigate differences in CFS composition among dung samples from different herbivore species to explore potential host-specific fungal associations.
Evaluate the limitations of using CFS as quantitative proxies for herbivore density, with particular attention to dispersal capacity, retention in substrates, and taphonomic biases.
Methods
Study area
The research was conducted in the Polish part of the Białowieża Forest (BF), a temperate broad-leaved and mixed forest ecosystem located in the Central European Lowland (Figure 1a). This forest is characterized by a complex mosaic of ecosystems shaped by varying management histories, including strictly protected areas, nature reserves, and managed forest stands (Jaroszewicz et al., 2019). The diversity of habitats, ranging from riparian forests to dry pine forests, supports high biological diversity and provides a unique setting for studying ecological processes and interactions.
The BF is particularly well-suited for investigating CFS as indicators of herbivore activity due to the presence of five large ungulate species (red deer, roe deer, moose, European bison and wild boar). This ecological variability was a critical factor in selecting sampling locations, ensuring the representation of both managed and unmanaged stands and areas differing by herbivore densities.
The region’s climate, with a mean annual temperature of 6.9°C and a recorded warming trend of 1.27°C over the period 1950–2015 (Boczoń et al., 2018), contributes to the formation of diverse microhabitats within the Białowieża Forest. Substantial amounts of deadwood and varying canopy openness further influence fungal community composition (Zimny et al., 2025), making this forest an ideal setting for ecological studies on coprophilous fungi. Sampling was conducted across a gradient of environmental conditions, including ungulate pressure and forest management intensity, to capture the ecological complexity of this unique lowland forest ecosystem.
Sampling and environmental variables
The sampling method was described in detail in our previous publications (Zimny, 2024a; Zimny et al., 2024, 2025), where the rationale for this particular method and type of sampling concerning our research objectives is thoroughly explained. Here, we briefly summarize the method, with an emphasis on elements specific to the collection and analysis of CFS.
Full metadata, including sample type and site location, are provided in Supplemental Table S1.
Moss pollsters
In August and September 2021, we collected 114 moss samples across various forest communities in the Polish part of the BF (Figure 1b). Sampling locations were selected based on predictive maps of ungulate presence and density (Bubnicki et al., 2019).
Tauber-type traps
Eighteen Tauber-type traps were established in February 2021, following the European Pollen Monitoring Program (PMP) pollen trapping standards (Hicks et al., 1996). The traps, buried flush with the ground, were protected by mesh covers to prevent small animals from entering and contained a solution of formalin, glycerin, and thymol to preserve the accumulated material and prevent mold growth. All traps were emptied at the end of the flowering season, in October 2021, following standard procedures (Räsänen et al., 2004).
Dung samples
Dung samples (27) were collected in September and October 2021 from 18 sampling sites where excrements were available. These included fresh and dry feces from roe deer Capreolus capreolus and red deer Cervus elaphus (11 samples), Eurasian moose Alces alces (7 samples), and European bison Bison bonasus (9 samples). As roe deer and red deer droppings could not be distinguished in this study, they were grouped under the term “deer.” For clarity, we used simplified terminology in the results section, also for droppings of Eurasian moose as “moose” and those of European bison as “bison.” No wild boar Sus scrofa feces were found, likely due to a significant decline in the wild boar population in the BF since 2015. This decline was driven by the African swine fever (ASF) virus, which has nearly eradicated wild boars in the region (Morelle et al., 2020).
Samples were carefully collected from the center of each dung pile using sterilized syringes to minimize the risk of airborne contamination. Where possible, efforts were made to pool material from multiple individuals of each species. The collected dung was stored in sealed bags during transport to prevent contamination before laboratory processing (Carrión, 2002).
At each site, five environmental variables were recorded to characterize local habitat conditions:
Wetness—ordinal (1–4) field assessment of habitat moisture,
Openness—measured canopy openness in percentages using a Plant Canopy Imager (CID Bio-Science Inc., USA),
Coarse woody debris (CWD)—volume of downed deadwood (m3/ha) estimated with the line-intersect method,
Total ungulate pressure (TUP)—composite index (0–8) reflecting signs of herbivore activity,
Forest management intensity (FMI)—categorical index (0–3) representing human intervention levels.
Detailed scoring criteria and definitions for each variable are provided in Supplemental Table S1.
Laboratory procedures
Sample preparation
All sample types underwent standard palynological preparation to ensure comparability across samples. This included washing with distilled water, 200 μm sieving, centrifugation, and chemical treatments (boiling in 10% KOH, Erdman’s acetolysis for 3 min, mounting in glycerin; Faegri and Iversen, 1989).
Moss samples were weighed, and their volume was recorded in the laboratory. Their weight ranged from 1.6 to 65.5 g (mean: 16 g), and volume from 9 to 90 ml (mean: 32 ml).
The total volume of liquid recovered from the Tauber-type traps ranged from 1000 to 3200 ml.
Dung samples were pre-treated to minimize surface contamination from airborne particles by carefully scraping to obtain the material from the interior. The collected dung subsamples were weighed before processing, ranging from 0.75 to 9.49 g (mean: 3.5 g).
Full metadata, including sample type, volume and/or weight, are provided in Supplemental Table S1.
Microscopic analysis
Prepared slides were analyzed under a light microscope with Nomarski contrast at 400× magnification. For moss and trap samples, a minimum of 700 arboreal pollen grains were counted per sample. For dung samples, a minimum of 250 land plant pollen grains were counted for each sample, along with accompanying fungal NPPs, except for sample MP68, which contained only 128 pollen grains due to the low pollen frequency. The identification of coprophilous fungal spores (CFS) was based on established palynological keys and descriptions provided by van Geel (1976), van Geel (1978), van Geel (1986, 2001), Bakker and van Smeerdijk (1982), van Geel et al. (2003), and van Geel and Aptroot (2006), and was further supported by selected mycological sources, including Li et al. (2023), which aided in the morphological assessment of fungal spore types. The identification process was additionally guided by resources from the Non-Pollen Palynomorph Image Database (NPP-ID; Shumilovskikh et al., 2022). The nomenclature of fungal NPPs follows current general guidelines (Miola, 2012).
Data normalization
CFS abundance was expressed as a percentage of total pollen (TP). While this approach is widely used in paleoecological research (e.g. Davis and Shafer, 2006; Gill et al., 2009; Parker and Williams, 2012), it has well-documented limitations. These include differences in dispersal and depositional pathways between fungal spores and wind-borne pollen, as well as variation in pollen influx across vegetation types (Hicks, 2001; van Asperen et al., 2020). Nevertheless, %TP was applied here to facilitate comparisons across heterogeneous sample types with varying volumes and collection conditions.
Data availability
Full taxonomic resolution of fNPPs is not included here, since these data form the basis of a separate forthcoming publication.
However, the complete dataset for moss samples has been deposited in an institutional repository under embargo and is available upon reasonable request (Zimny, 2024b, https://doi.org/10.58132/IBDFWF).
Full sample-level metadata, including sample weight and/or volume, are presented in Supplemental Table S1.
To ensure transparency, raw counts of total pollen (AP + NAP), CFS, and other fungal non-pollen palynomorphs (fNPPs, excluding CFS) are provided in Supplemental Table S2.
Statistical analyses
To compare the composition and relative abundance of CFS across sample types (moss polsters, Tauber-type traps, and dung), descriptive statistics were calculated, including taxonomic richness, frequency, and mean percentage contribution to the total pollen sum (%TP). The standard error (±1σ) was used to illustrate variability in CFS abundance.
Non-metric multidimensional scaling (NMDS) was employed to visualize patterns in CFS composition and explore their relationship with environmental variables. NMDS ordinations were performed separately for each sample type, using Euclidean distance as the dissimilarity metric and based on CFS taxa’s relative abundance (%TP). Analyses were conducted in PAST version 5.0.2 (Hammer et al., 2021). Environmental variables—coarse woody debris (CWD), wetness, openness, total ungulate pressure (TUP), and forest management intensity (FMI)—were passively fitted onto NMDS ordinations to assess their influence on sample distribution. Model fit was evaluated using the stress value, with lower values indicating a better representation of the data structure.
To test the explanatory power of environmental variables, redundancy analysis (RDA) was also conducted separately for each sample type. The proportion of variance explained by ecological factors was assessed using R² and adjusted R² values. Statistical significance was evaluated via permutation tests (n = 999), with the significance threshold set at p < 0.05.
To assess the potential influence of ecological specificity on ordination results, we repeated the NMDS and RDA analyses using two subsets of the dataset: (1) taxa classified as obligate coprophilous, known to grow exclusively on dung (see Table 1), and (2) all remaining taxa, considered either facultative or ecologically uncertain in their dung specificity (Chmiel, 2006; Kozłowska et al., 2019; Li et al., 2023). Full results of these analyses are presented in the Supplemental Material (Supplemental Tables S3 and S4; Supplemental Figures S1 and S2).
Frequency (% of all samples) and maximum abundance (% TP) of coprophilous fungal spore (CFS) taxa in different sample types.
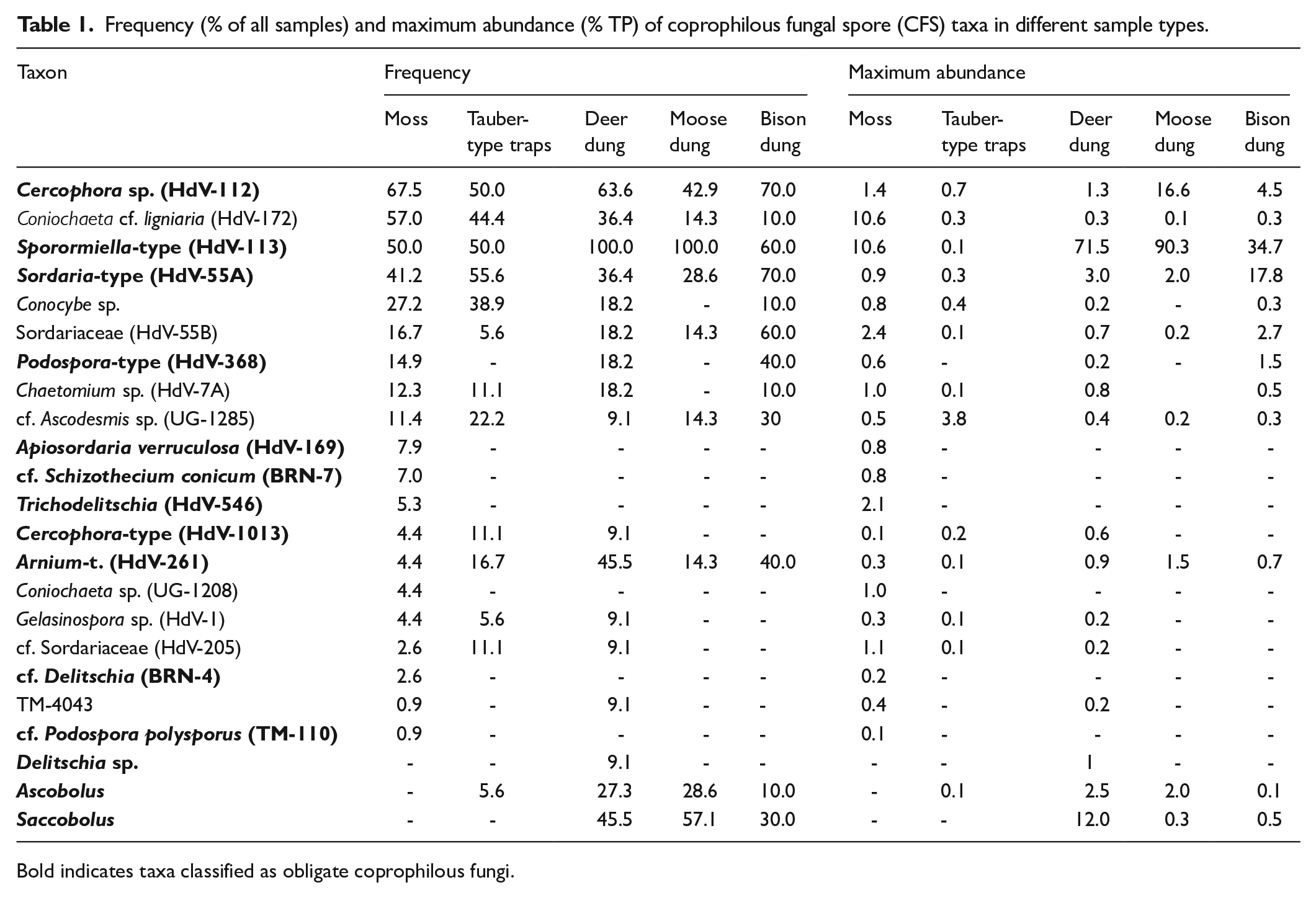
Bold indicates taxa classified as obligate coprophilous fungi.
Results
Comparison of CFS composition across different sample types
Moss pollsters
In moss samples from the Białowieża Forest, coprophilous fungal spores (CFS) accounted for 0%–15.7% of the total pollen sum (TP; Figure 2a), with six samples lacking any CFS. A total of 20 fungal taxa, either exclusively or partially coprophilous, were identified (Table 1, Figure 3). The most frequent taxa were Cercophora sp. (HdV-112) and Coniochaeta cf. ligniaria (HdV-172), detected in over 67% and 57% of the samples, respectively. Sporormiella-type (HdV-113) and Sordaria-type (HdV-55A) were slightly less common, found in more than 40% of the samples, while Conocybe sp. appeared in 27%.
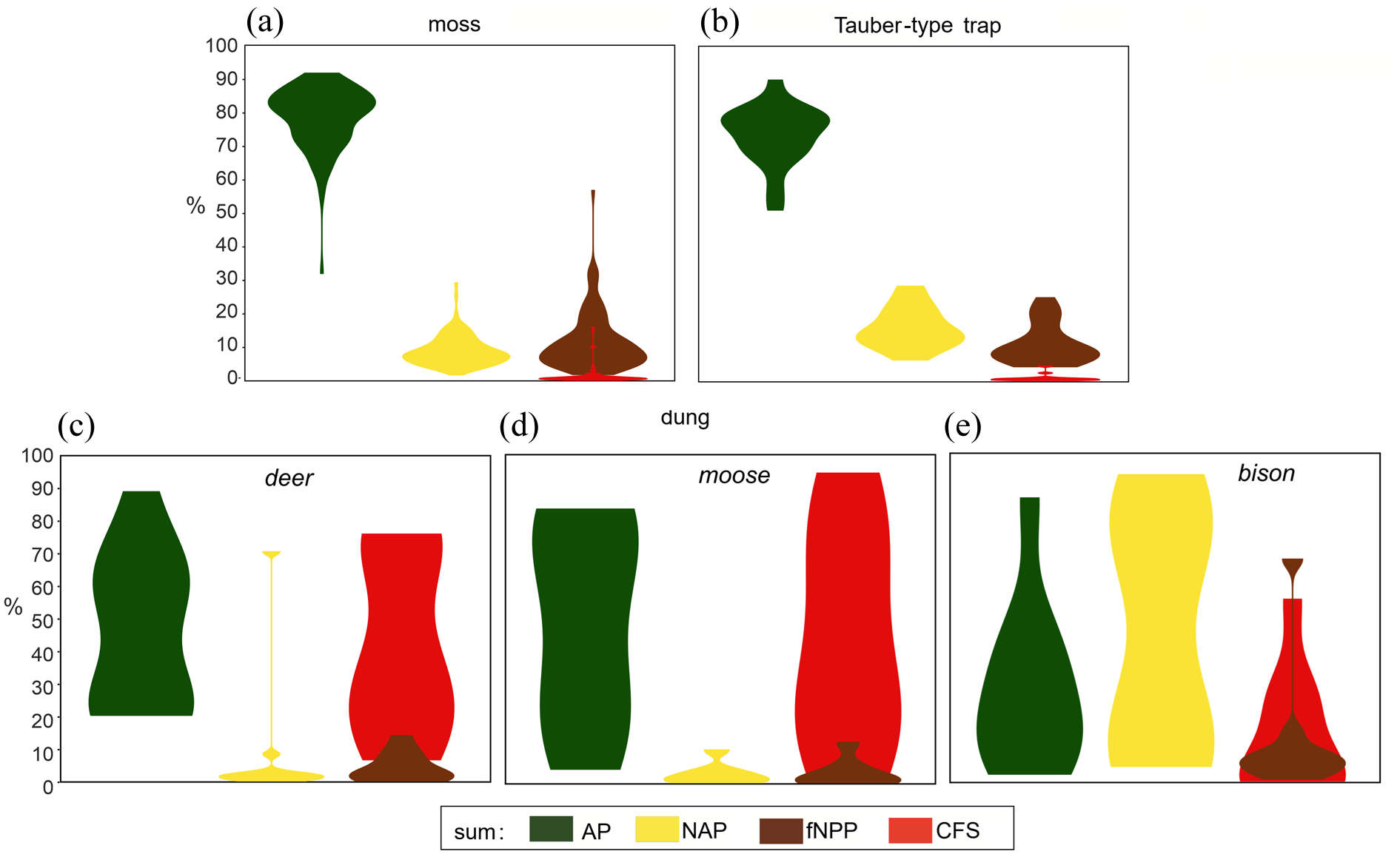
The percentage share of coprophilous fungal spores (CFS) in the total pollen sum (TP) in moss polsters (a), Tauber-type traps (b), and ungulate dung: red/roe deer (c), Eurasian moose (d), European bison (e), presented against the percentage share of AP, NAP, and other fNPPs.
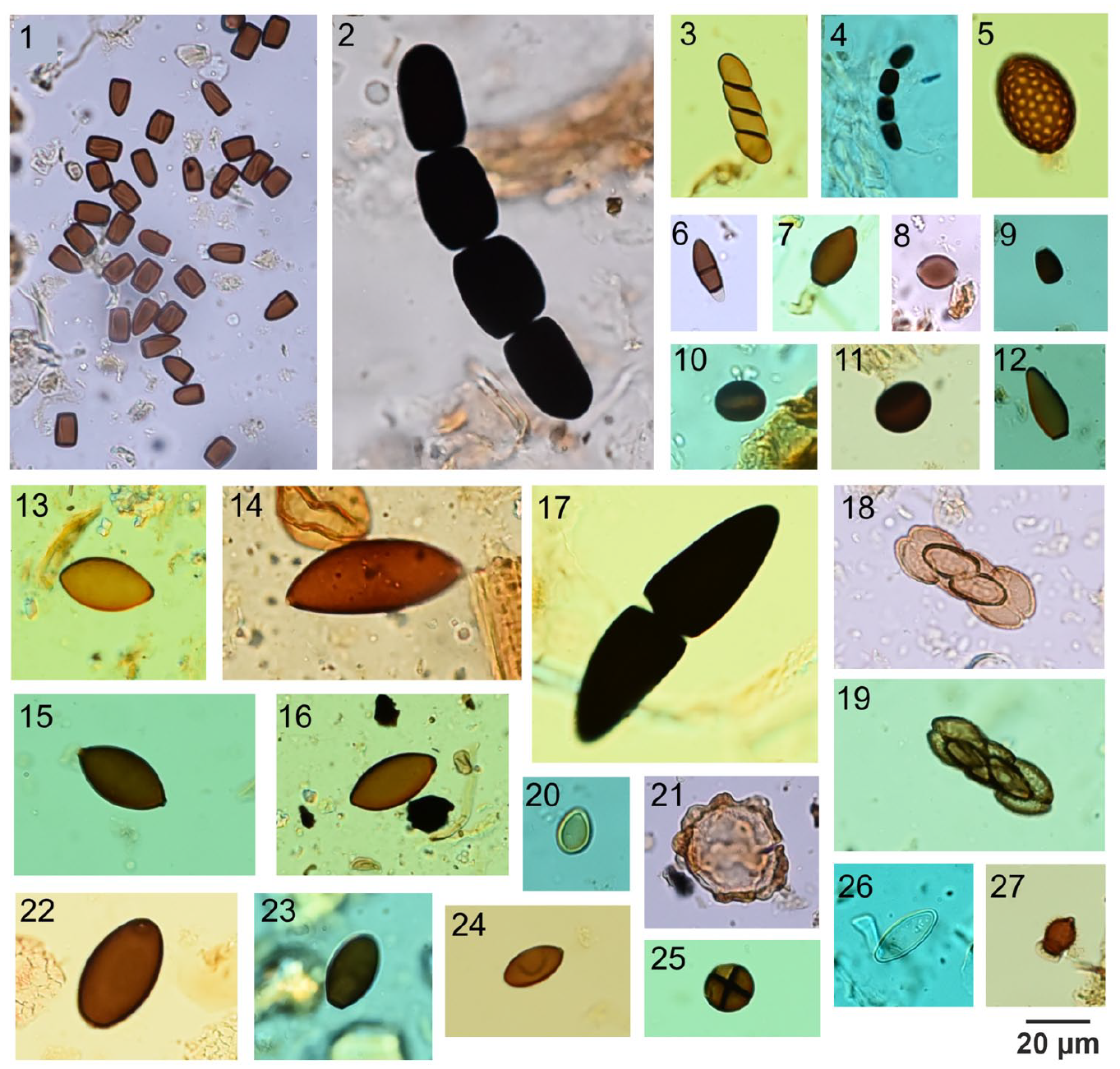
Light microscope photographs of fungal non-pollen palynomorph types identified and analyzed in this study: 1–4. Sporormiella-type (HdV-113); 5. Gelasinospora sp. (HdV-1); 6. Cercophora sp. (HdV-112); 7. Cercophora-t. (HdV-1013); 8. Chaetomium sp. (HdV-7A); 9. Trichodelitschia (HdV-546); 10. Coniochaeta cf. ligniaria (HdV-172); 11. Coniochaeta sp. (UG-1208); 12. cf. Delitschia (BRN-4); 13. Sordaria-type (HdV-55A); 14. Arnium-t. (HdV-261); 15. Sordariaceae (HdV-55B); 16. cf. Sordariaceae (HdV-205); 17. Delitschia sp.; 18–19. Saccobolus; 20. Conocybe sp.; 21. cf. Ascodesmis sp. (UG-1285); 22. Podospora-type (HdV-368); 23. cf. Podospora polysporus (TM-110); 24. cf. Schizothecium conicum (BRN-7); 25. TM-4043; 26. Ascobolus; 27. Apiosordaria verruculosa (HdV-169).
Other taxa, including Sordariaceae (HdV-55B), Podospora-type (HdV-368), Chaetomium sp. (HdV-7A), and cf. Ascodesmis sp. (UG-1285) were present in more than 10% of the samples. Several others, including Apiosordaria verruculosa (HdV-169), cf. Schizothecium conicum (BRN-7), Trichodelitschia (HdV-546), Cercophora-type (HdV-1013), Arnium-type (HdV-261), Coniochaeta sp. (UG-1208), Gelasinospora sp. (HdV-1), cf. Sordariaceae (HdV-205), and cf. Delitschia (BRN-4) occurred infrequently (<10%), with TM-4043 and cf. Podospora polysporus (TM-110) found in single samples.
In terms of abundance, Sporormiella-type (HdV-113) and Coniochaeta cf. ligniaria (HdV-172) exhibited the highest mean percentage contributions to the TP (Figure 4a), indicating their dominant presence in the moss samples. Cercophora sp. (HdV-112) and Sporormiella-type (HdV-113) showed slightly lower values, while the remaining taxa exhibited considerably lower means and more significant variability in occurrence.
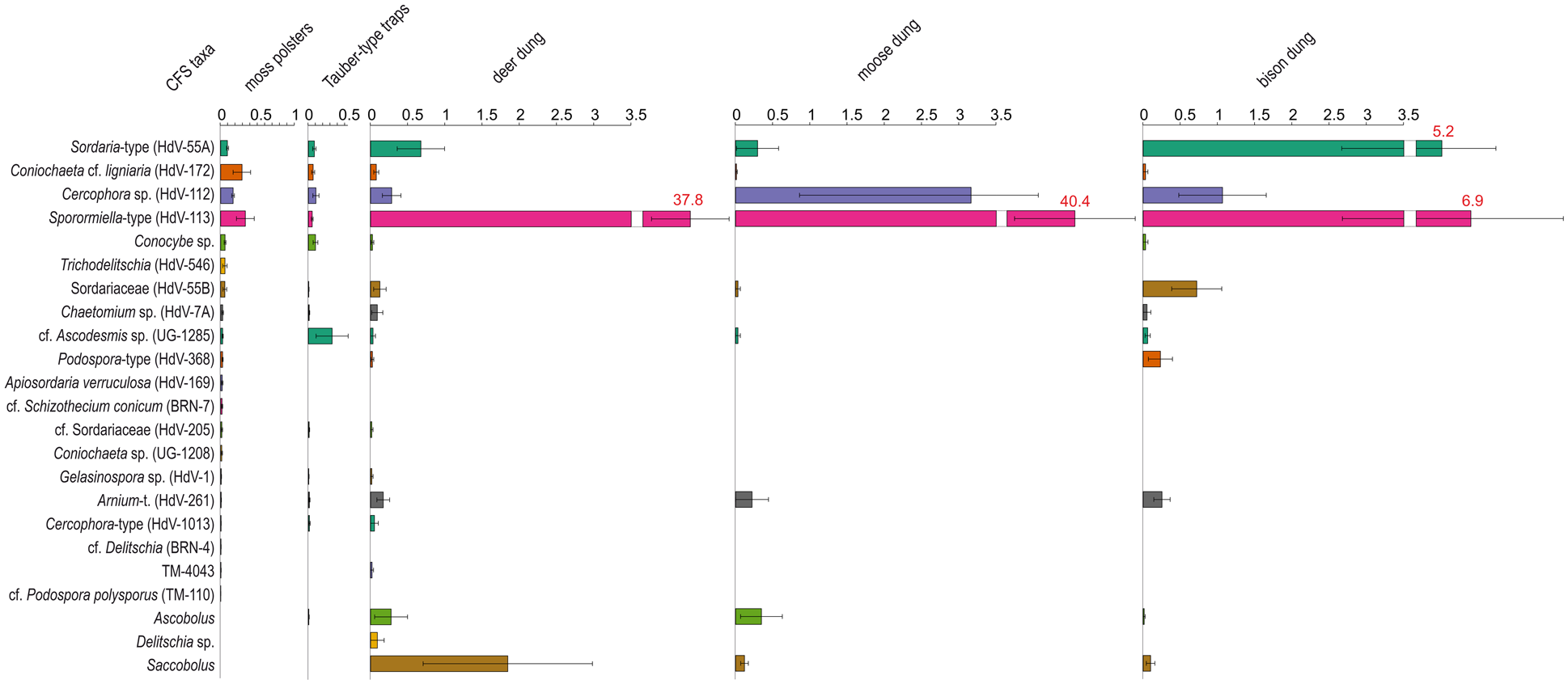
Mean percentage contribution of coprophilous fungal spores (CFS) to the total pollen sum (TP) in (a) 114 moss polsters, (b) 18 Tauber-type traps, and (c) dung samples from red/roe deer (11), Eurasian moose (7) and European bison (9). The bar chart displays the mean percentage values for each taxon, with whiskers representing the standard error (± one sigma). Values marked in red exceeded the plotted scale range.
The percentage contribution of individual CFS taxa in the 114 moss samples ranged from 0% to 10.6%. The highest values were recorded for Sporormiella-type (HdV-113) and Coniochaeta cf. ligniaria (HdV-172), reaching 10.6%. In some samples, Sordariaceae (HdV-55B) and Trichodelitschia (HdV-546) exceeded 2%, while Cercophora sp. (HdV-112), cf. Sordariaceae (HdV-205), Coniochaeta sp. (UG-1208), and Chaetomium sp. (HdV-7A) ranged between 1% and 1.37%.
All remaining taxa contributed less than 1% to the TP, including Sordaria-type (HdV-55A), cf. Schizothecium conicum (BRN-7), Apiosordaria verruculosa (HdV-169), Conocybe sp., Podospora-type (HdV-368), cf. Ascodesmis sp. (UG-1285), TM-4043, Gelasinospora sp. (HdV-1), Arnium-type (HdV-261), cf. Delitschia (BRN-4), Cercophora-type (HdV-1013), and cf. Podospora polysporus (TM-110).
Tauber-type traps
In Tauber-type traps, CFS accounted for 0.1%–4.6% of the total pollen sum (TP; Figure 2b). Fourteen CFS taxa were identified (Table 1), with Sordaria-type (HdV-55A) being the most frequent, occurring in over 55% of the samples. Sporormiella-type (HdV-113) and Cercophora sp. (HdV-112) were each found in 50% of the samples, followed by Coniochaeta cf. ligniaria (HdV-172) and Conocybe sp., present in 44% and 39%, respectively. Arnium-type (HdV-261) appeared in 17% of the samples. Less frequent taxa included Cercophora-type (HdV-1013), Chaetomium sp. (HdV-7A), and cf. Sordariaceae (HdV-205), each found twice, while Gelasinospora sp. (HdV-1), Sordariaceae (HdV-55B), and Ascobolus were recorded only once.
In terms of abundance, cf. Ascodesmis sp. (UG-1285) exhibited the highest contribution to the TP, with an average of 3% and a peak value of 3.8% (Figure 4b). Cercophora sp. (HdV-112) and Conocybe sp. followed, with mean values approximately three times lower and maximum values of 0.7% and 0.4%, respectively.
Sordaria-type (HdV-55A), Coniochaeta cf. ligniaria (HdV-172), and Sporormiella-type (HdV-113) had mean contributions below 0.1%, with peak values never exceeding 0.3%. The remaining taxa, including Arnium-type (HdV-261), Cercophora-type (HdV-1013), Chaetomium sp. (HdV-7A), cf. Sordariaceae (HdV-205), Ascobolus, Gelasinospora sp. (HdV-1), and Sordariaceae (HdV-55B) had very low contributions (below 0.015%) and were present in single samples only.
Dung samples
In the 26 dung samples from herbivorous animals (deer, moose and bison), CFS constituted between 0% (sample ID: MP18-4) and 94.3% of the TP (Figure 2c). Their contribution ranged from 7% to 76% in deer, 2.5% to 94% in moose, and 0% to 55% in bison. Taxonomic richness was highest in deer dung with 17 taxa identified, compared to 12 taxa in bison dung, and 9 in moose dung (Table 1).
Sporormiella-type (HdV-113) was the most frequent taxon in cervid (deer and moose) dung, present in all samples. It was also recorded in 60% of bison dung samples (Table 1). In deer dung, Cercophora sp. (HdV-112) was found in over 63% of the samples, followed by Arnium-type (HdV-261) and Saccobolus (both 45%), Sordaria-type (HdV-55A), Coniochaeta cf. ligniaria (HdV-172; >36%), and Ascobolus (>27%; Table 1).
Several taxa, including Sordariaceae (HdV-55B), Podospora-type (HdV-368), Conocybe sp., and Chaetomium sp. (HdV-7A), were found in two samples, while rarer taxa such as cf. Ascodesmis sp. (UG-1285), Cercophora-type (HdV-1013), Gelasinospora (HdV-1), TM-4043, cf. Sordariaceae (HdV-205) and Delitschia sp. appeared only once.
In moose dung, Sporormiella-type (HdV-113) was the most frequent taxon, present in all samples (Table 1). Saccobolus was detected in 57.1% of the cases, while Cercophora sp. (HdV-112) appeared in 42.9%. Ascobolus and Sordaria-type (HdV-55A) were each recorded in 28.6%. Arnium-type (HdV-261), Sordariaceae (HdV-55B), Coniochaeta cf. ligniaria (HdV-172), and cf. Ascodesmis sp. (UG-1285) were identified only once, corresponding to 14.3% of the examined dung.
In bison dung, Cercophora sp. (HdV-112) and Sordaria-type (HdV-55A) were the most frequently detected taxa, each appearing in 70% of the analyzed material (Table 1). Sporormiella-type (HdV-113) and Sordariaceae (HdV-55B) were identified in 60% of the samples. Arnium-type (HdV-261) and Podospora-type (HdV-368) were observed in 40% of the cases, while cf. Ascodesmis sp. (UG-1285) and Saccobolus were each recorded in 30% of dung samples. Coniochaeta cf. ligniaria (HdV-172), Conocybe sp., Chaetomium sp. (HdV-7A), and Ascobolus were rare, all of which were found in only one sample.
In terms of abundance, Sporormiella-type (HdV-113) dominated across all dung samples with a mean contribution of 37.3% to the TP (Figure 4c). Saccobolus followed with 1.84%, while Sordaria-type (HdV-55A) contributed 0.68%. Cercophora sp. (HdV-112) and Ascobolus each averaged 0.28%, and Arnium-type (HdV-261) and Sordariaceae (HdV-55B) contributed 0.17% and 0.13%, respectively. All other taxa remained below 0.1%. Maximum values reached 71.5% for Sporormiella-type (HdV-113), 12% for Saccobolus, 3% for Sordaria-type (HdV-55A), 2.5% for Ascobolus, and 1.3% for Cercophora sp. (HdV-112).
The CFS composition varied notably by dung type, with Sporormiella-type (HdV-113) consistently dominating across all samples, though with significant differences in abundance. In deer dung, it averaged 38% of the TP, occasionally exceeding 71%. Saccobolus was present at lower levels, with a mean of 1.8% and a maximum of 12%, while Sordaria-type (HdV-55A) and Ascobolus remained below 3%.
In moose dung, Sporormiella-type (HdV-113) was even more dominant, averaging 40.4% and occasionally surpassing 90%. Cercophora sp. (HdV-112) had a mean contribution of 3.2%, peaking at 16.6%, while other taxa, including Sordaria-type (HdV-55A) and Arnium-type (HdV-261), remained below 2%.
In bison dung, the assemblage was more balanced, with Sporormiella-type (HdV-113) averaging 6.2%, Sordaria-type (HdV-55A) 4.6%, and Cercophora sp. (HdV-112) 1%; their maxima were 17.8%, 4.5%, and 2.7%, respectively. Podospora-type (HdV-368) remained below 1.5%.
Influence of environmental variables on CFS in different sample types
Moss samples
The NMDS ordination based on Euclidean distance was performed for 108 moss samples to explore the composition of CFS in relation to five environmental variables: CWD, wetness, openness, TUP, and FMI. The stress value of 0.1722 indicated an acceptable fit, with moderate reliability in representing sample relationships in two dimensions.
The NMDS ordination plot (Figure 5) showed that taxa such as Sordariaceae (HdV-55B), Trichodelitschia (HdV-546), and Apiosordaria verruculosa (HdV-169) were positively correlated with the openness vector, suggesting their preference for open habitats. TUP was directed toward the central sample cluster, indicating weak overall structuring by herbivore pressure and was opposite to CWD associated with Arnium-type (HdV-261), implying the relationship of this taxon with habitats rich in deadwood. FMI was positively correlated with Podospora-type (HdV-368) and Chaetomium sp. (HdV-7A), suggesting a possible influence of forest management on the abundance of these taxa. Notably, cf. Podospora polysporus (TM-110) and Coniochaeta sp. (UG-1208) were located away from the main cluster, indicating possible specialization for unique microhabitats not considered in our study.
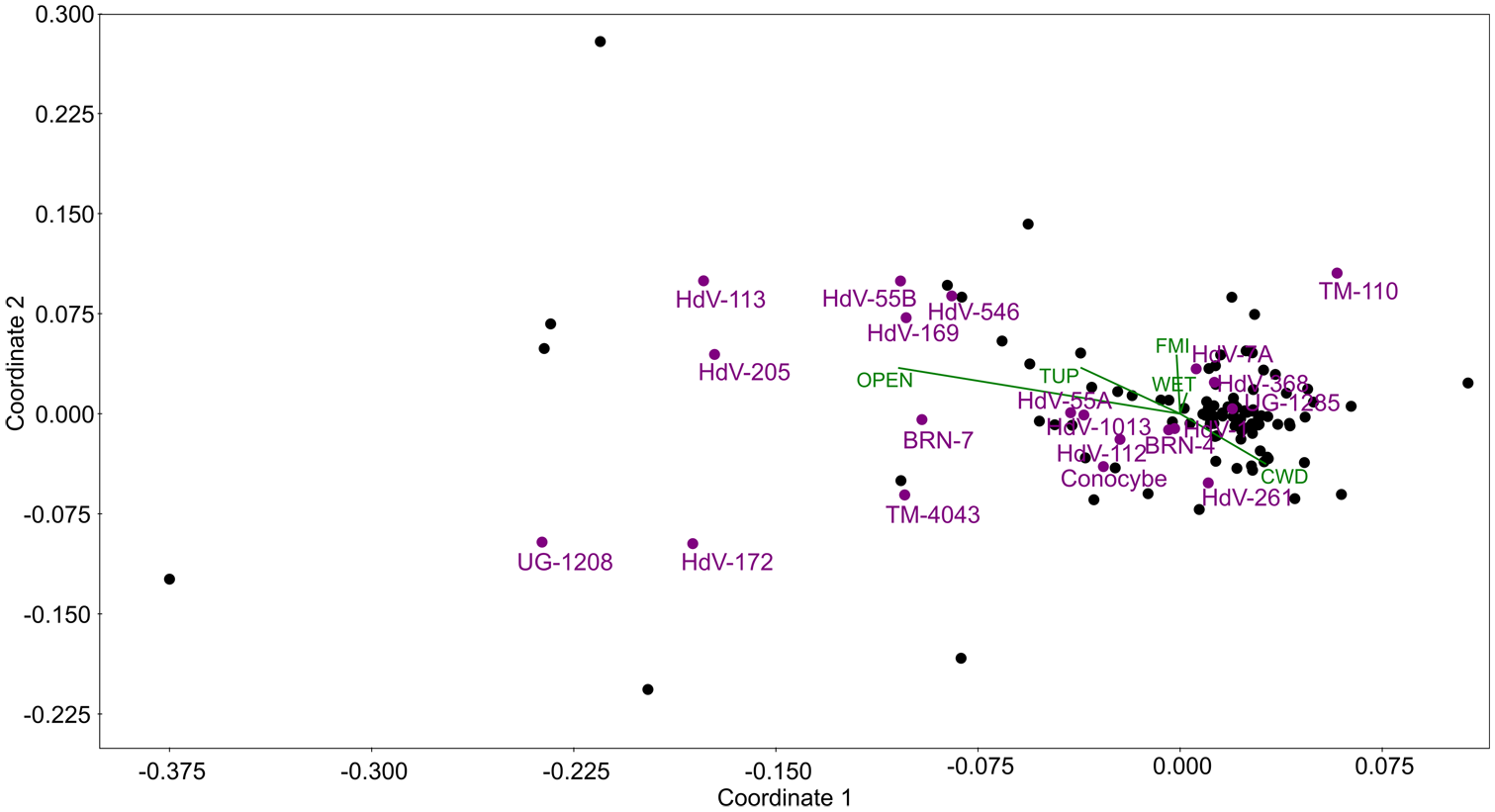
Non-metric multidimensional scaling (NMDS) ordination plot of 108 moss samples (black dots), showing the distribution of 20 coprophilous fungal spore (CFS) types (purple dots and labels) based on their percentage contribution to the total pollen sum (TP), in relation to five environmental variables (green vectors and labels): coarse woody debris (CWD), wetness (WET), openness (OPEN), total ungulate pressure (TUP), and forest management intensity (FMI).
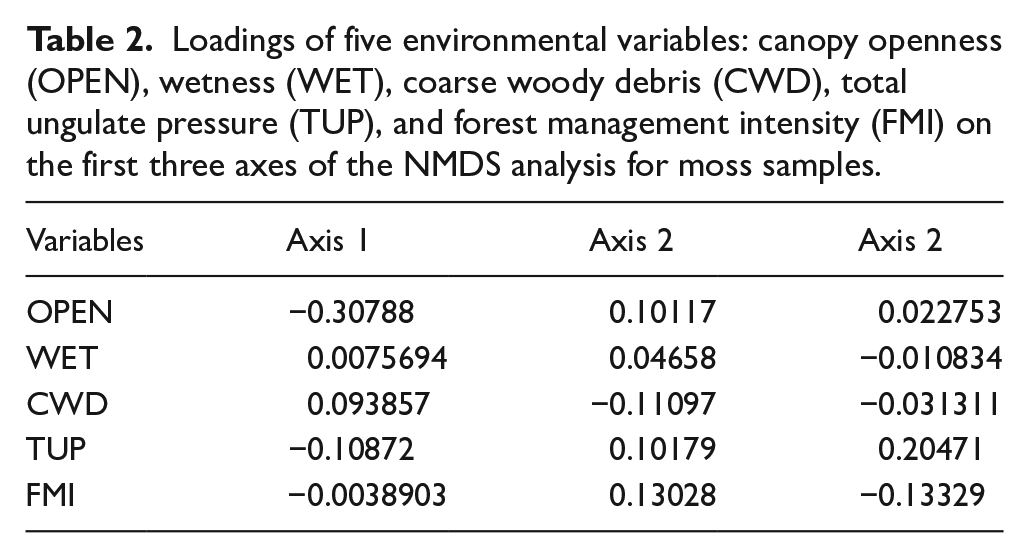
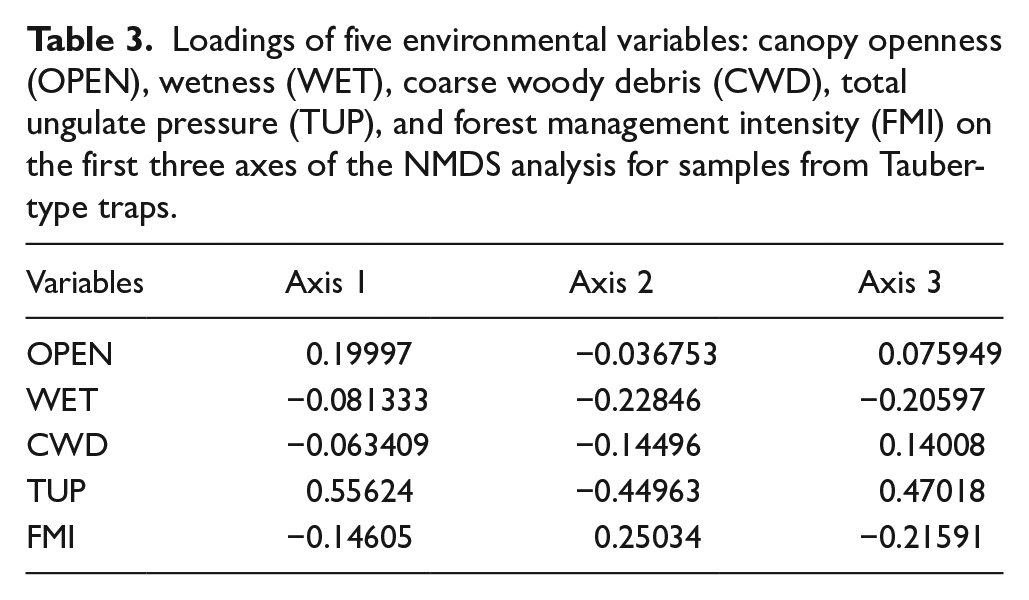
Table 2 presents the loadings of environmental variables on the first three NMDS axes. Among them, openness and TUP had the strongest influence on sample distribution, whereas wetness had the weakest.
Loadings of five environmental variables: canopy openness (OPEN), wetness (WET), coarse woody debris (CWD), total ungulate pressure (TUP), and forest management intensity (FMI) on the first three axes of the NMDS analysis for moss samples.
The redundancy analysis (RDA) indicated low explanatory power of environmental variables: R² = 0.072 (7.2%), R² adj = 0.0269 (2.7%), with p = 0.137 and F = 1.59. The lack of statistical significance suggests that most variance in CFS composition is not explained by the measured variables.
To evaluate whether CFS assemblages may be structured differently when restricted to obligate dung fungi, we conducted separate NMDS and RDA analyses for taxa classified as obligate and non-obligate (facultative or ecologically uncertain) coprophilous spore types. For obligate CFS, the RDA explained slightly more variance than in the full dataset (R² = 0.0968; R² adj = 0.0472), although the model was not statistically significant (F = 1.95, p = 0.113). In contrast, the facultative group showed substantially lower explanatory power (R² adj = 0.0042; p = 0.355). The NMDS ordinations revealed broadly similar compositional patterns across both groups, with only minor shifts in taxa positions. These results suggest that while restricting analyses to obligate coprophiles may slightly improve the model fit, it does not fundamentally alter the ecological patterns observed. Supplemental figures and tables (Supplemental Figures S1 and S2, Supplemental Tables S3 and S4) provide visualizations and full statistics of both subsets.
Tauber-type traps
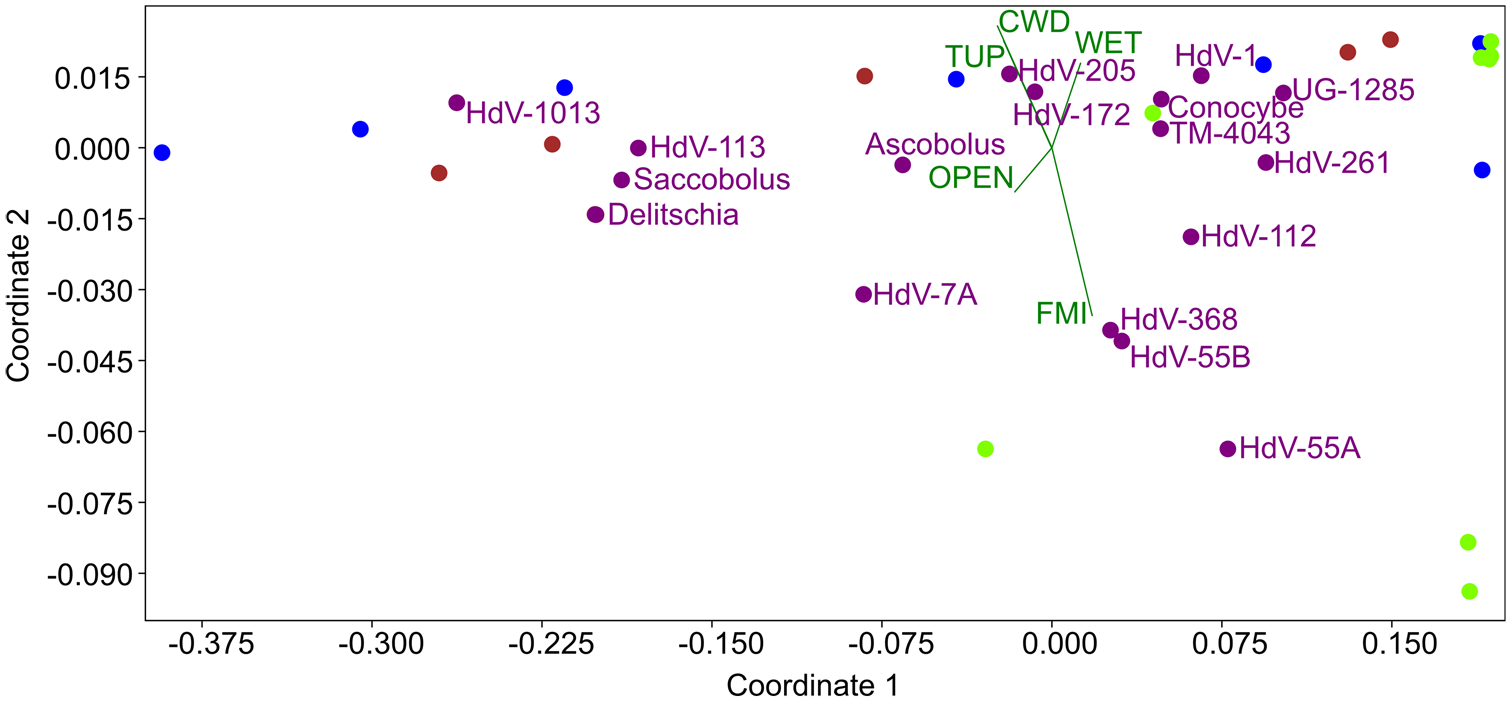
NMDS analysis of 18 Tauber-type trap samples yielded a stress value of 0.1042, indicating a good fit. Most CFS taxa clustered in the left part of the ordination plot (Figure 6), suggesting similar environmental preferences. The oppositely directed TUP and FMI vectors strongly differentiated the samples. Taxa, such as Cercophora-t. (HdV-1013) and Coniochaeta sp. (UG-1285) were positioned apart from the main cluster, potentially preferring habitats with lower ungulate pressure. Other environmental variables, such as CWD, wetness, and openness, exerted minimal influence on sample distribution, as indicated by their short vector lengths.
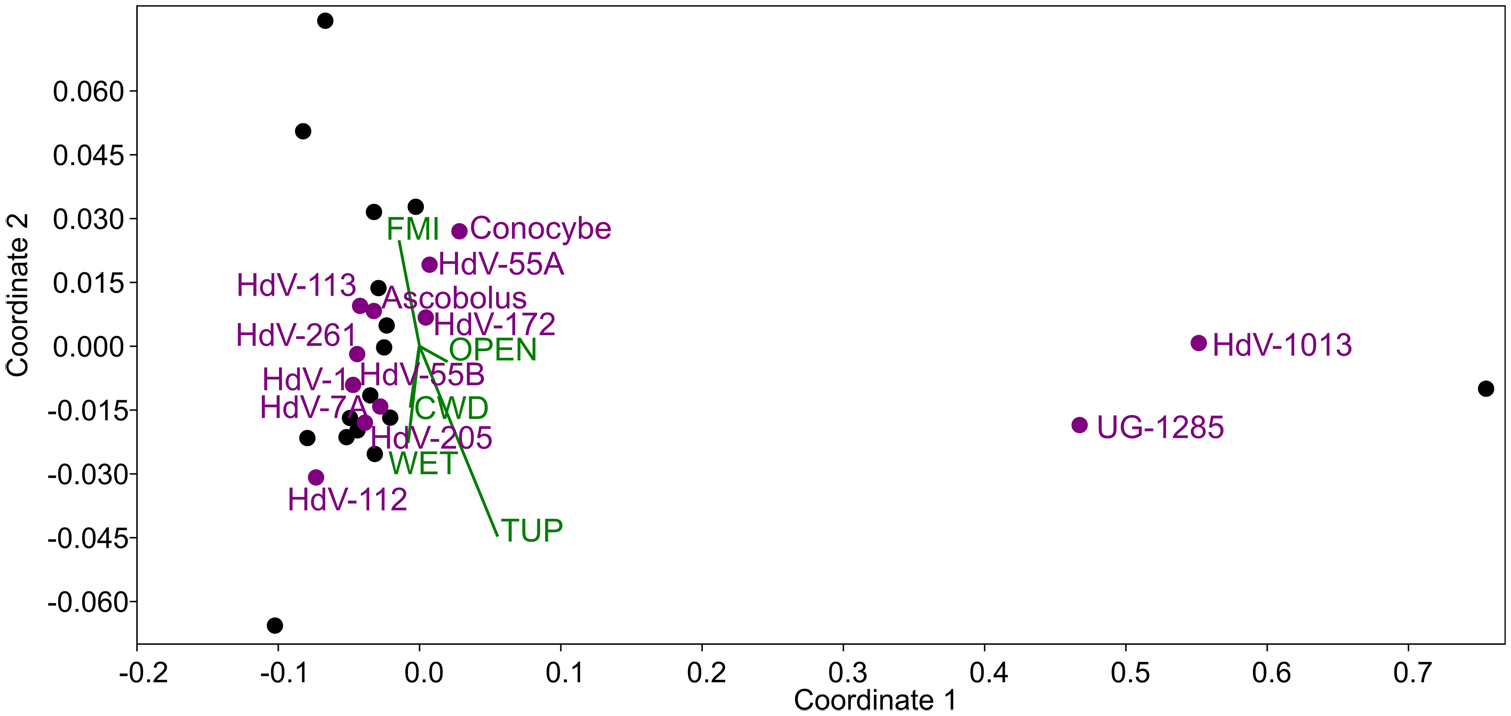
NMDS ordination plot of 18 samples from Tauber-type traps (black dots), showing the distribution of 13 coprophilous fungal spore (CFS) types (purple dots) in relation to five environmental variables (green vectors and labels): coarse woody debris (CWD), wetness (WET), canopy openness (OPEN), total ungulate pressure (TUP), and forest management intensity (FMI).
Table 3 confirms that TUP had the most significant influence (along all three axes considered), while FMI showed weaker effects. Other vectors had minor roles in shaping sample distribution.
Loadings of five environmental variables: canopy openness (OPEN), wetness (WET), coarse woody debris (CWD), total ungulate pressure (TUP), and forest management intensity (FMI) on the first three axes of the NMDS analysis for samples from Tauber-type traps.
RDA explained 42.58% of the total variance in CFS composition (R² = 0.4258), with R² adj = 0.1865, but the model was not statistically significant (p = 0.129, F = 1.78). While TUP appeared influential, the overall model suggests moderate structuring with notable unexplained variability.
Dung samples
NMDS ordination of 27 dung samples (red/roe deer—11, moose—7, bison—9) yielded a stress value of 0.0056, indicating an excellent fit. Samples clustered distinctly by host species (Figure 7): bison dung (green dots) generally occupied the right side of the ordination space, moose dung (blue dots) was positioned more centrally, while deer dung (brown dots) was more scattered across the left side. However, the environmental vectors (CWD, wetness, openness, TUP, FMI) were relatively short, indicating only subtle influences on the distribution of samples and taxa.
NMDS ordination plot of dung samples from red/roe deer (brown dots), moose (blue dots), and European bison (green dots), showing the distribution of 17 coprophilous fungal spore (CFS) types (purple dots) in relation to five environmental variables (green vectors and labels): coarse woody debris (CWD), wetness (WET), canopy openness (OPEN), total ungulate pressure (TUP), and forest management intensity (FMI).
Among the environmental factors, wetness and TUP showed the strongest associations with Axis 3 (loadings: wetness = 0.389, TUP = 0.318), while CWD had a notable effect along Axes 1 (−0.295) and 2 (0.331; Table 4). FMI showed a moderate negative influence on Axis 2 (−0.446), suggesting it may contribute to sample variation along this gradient. Openness exhibited relatively weak loadings on all axes.
Loadings of five environmental variables canopy openness (OPEN), wetness (WET), coarse woody debris (CWD), total ungulate pressure (TUP), and forest management intensity (FMI) on the first three axes of the NMDS analysis for dung samples.

Interpretation of taxon-specific distributions remains tentative due to the limited strength of environmental vectors. For instance, Gelasinospora sp. (HdV-1) and Conocybe sp. were positioned near the CWD and TUP vectors, suggesting a possible association with decomposing woody material and herbivore activity, although the short vector lengths caution against overinterpretation. Sordaria-type (HdV-55A) and Sordariaceae (HdV-55B) aligned more closely with FMI, while Podospora-type (HdV-368) and Chaetomium sp. (HdV-7A) were closer to openness. However, the shortness of vectors limits confidence in these associations.
The RDA model explains 26.63% of the variance (R² = 0.2663), with R² adj = 0.0916. Despite modest explanatory power, the model was not statistically significant (p = 0.196, F = 1.52). This indicates a weak influence of measured environmental variables on CFS composition in dung samples, suggesting that differences between the samples were due to the unmeasured factors or host-specific dung characteristics.
A comparative analysis across moss, Tauber-type traps, and dung samples revealed consistent patterns in how environmental gradients influence CFS assemblages. Openness and TUP emerged as the most influential variables, particularly in moss samples and Tauber-type traps, whereas in dung samples, CFS assemblages appeared to be more strongly shaped by host-specific conditions and habitat wetness. However, redundancy analysis (RDA) across all sample types showed low and statistically non-significant explanatory power, indicating that CFS composition may be influenced by unmeasured microhabitat factors or stochastic processes.
The NMDS ordination revealed substantial overlap among the three studied sample types (moss polsters, Tauber-type traps and ungulate dung), indicating a high degree of compositional similarity in CFS assemblages (Figure 8). Dung samples exhibited the most significant variation in CFS composition, with points broadly dispersed across the ordination space. In contrast, samples from moss and Tauber-type traps were more tightly clustered, predominantly in the left section of the plot. The stress value of 0.1768 indicates an acceptable representation of the data in the reduced-dimensional space.
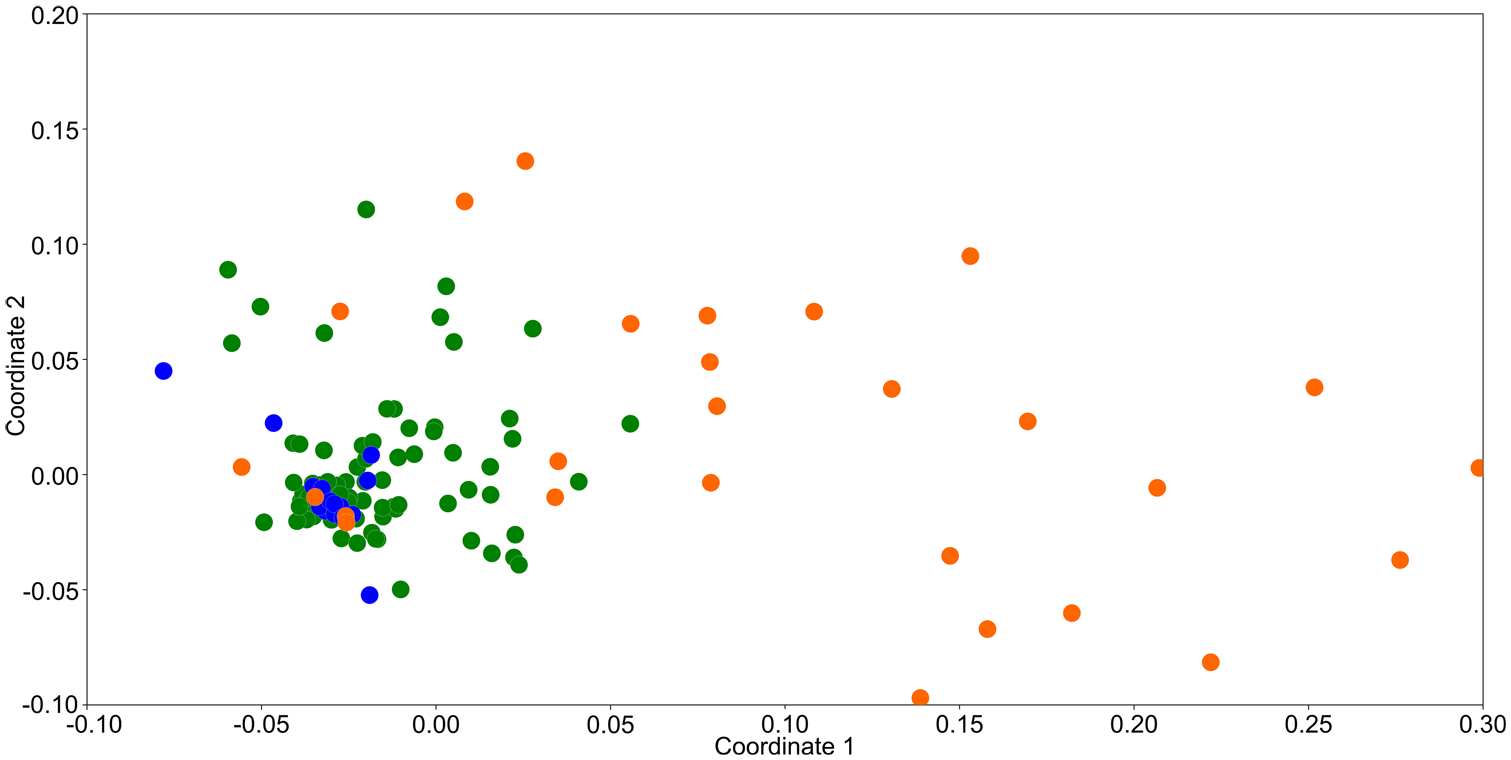
NMDS ordination plot showing the distribution of 158 samples from three types: dung (orange dots), moss polsters (green dots), and Tauber-type traps (blue dots), based on the composition of coprophilous fungal spores (CFS).
Discussion
Composition and abundance of coprophilous fungal spores in different sample types
The analysis of CFS composition and abundance in different sample types revealed significant differences between moss samples, Tauber-type traps, and dung samples. Mosses exhibited the highest taxonomic diversity, suggesting that they function as passive spore traps, effectively accumulating coprophilous fungal spores primarily from local sources and, to a lesser extent, from airborne deposition over extended periods. Similar retention mechanisms have been demonstrated for pollen in moss carpets, which capture and preserve airborne material over time (Räsänen et al., 2004). Compared to dung samples, which were dominated by Sporormiella-type (HdV-113), Podospora-type (HdV-368), and Sordaria-type (HdV-55A), moss samples contained a wider range of coprophilous fungi taxa, including those that are not strictly dung-dependent but also occur on other decomposing organic matter, such as Chaetomium sp. (HdV-7A) and Gelasinospora sp. (HdV-1), as well as taxa potentially associated with wood decomposition, like Coniochaeta cf. ligniaria (HdV-172; Krug et al., 2004).
However, the variability in the abundance of CFS within moss samples was much higher than in other types of sampling, ranging from 0% to 15.7% TP. The absence of CFS in six moss samples likely reflects local variation in dung availability, possibly influenced by herbivore activity, movement patterns, and differences in moss-specific retention capacity (Sundberg, 2013). The low proportion of CFS in Tauber-type traps, ranging from 0.1% to 4.6% TP, indicates that the airborne dispersal of these fungi is likely limited. Unlike many wind-dispersed fungal taxa, coprophilous fungi do not typically produce lightweight, airborne spores capable of long-distance transport, as their dispersal relies on ballistic ejection mechanisms and substrate-dependent spread. This means that their presence in moss samples likely results from proximity to dung deposits rather than long-distance transport (van Asperen et al., 2016; van Asperen et al., 2022).
Additionally, local water activity and microtopographic position may influence the lateral movement and accumulation of fungal spores, especially in moss carpets and open trap surfaces. Although these effects are best documented for pollen transport (e.g. Lisitsyna et al., 2012), similar taphonomic processes may operate for fungal spores, particularly those sensitive to moisture and substrate structure (van Asperen et al., 2022).
It is important to note that the Tauber-type traps were sampled only during a single annual cycle (2021), which may limit the generalizability of the results due to potential interannual and seasonal variation in spore deposition. While no published datasets exist for seasonal changes in coprophilous fungal spore (CFS) deposition in forest pollen traps, previous research has demonstrated that pollen accumulation in Tauber-type traps can vary substantially across seasons and years, depending on vegetation dynamics and environmental conditions (Pardoe et al., 2010). Although CFS dispersal mechanisms differ from those of pollen, such findings underscore the importance of multi-year sampling when aiming to capture temporal patterns. Experimental studies of CFS dynamics have shown seasonal peaks in spore abundance, with some taxa becoming more prominent during colder periods or under specific habitat conditions (van Asperen et al., 2020). This temporal variability highlights the need for cautious interpretation of short-term datasets and reinforces the value of complementary sampling strategies. Despite the limited temporal scope of our trap dataset, its comparison with contemporaneously collected moss and dung samples allowed us to assess how different sampling approaches integrate fungal signals over time and space.
In contrast, dung samples exhibited the highest concentration of CFS, reaching 94.3% TP, confirming that this substrate is a potential source of CFS in paleoecological deposits (Gill et al., 2013). The absence of CFS in one bison dung sample (M18-4) is likely explained by the early stage of dung decomposition at the time of collection, as the sample was taken from a fresh bison deposit before the development of visible fungal fruiting bodies. Although all samples were processed using an identical palynological protocol, it is worth noting that this standard method may preferentially preserve thick-walled spores (e.g. Sporormiella) while underrepresenting more fragile taxa such as Ascobolus (Richardson, 2001; van Asperen, 2017; van Asperen et al., 2016). This may affect overall spore counts, but does not account for the complete absence observed in this particular case. Moreover, the number of counted fungal spores and pollen grains varied substantially across sample types, particularly in dung samples, where overall spore abundance was often low. To enable comparisons across diverse substrates with different volumes and sedimentation contexts, we relied on percentage values relative to the total pollen sum (%TP). However, proportional estimates based on low total counts are subject to increased statistical uncertainty. Following Maher (1972), even minor differences in %TP may reflect stochastic variation rather than ecologically meaningful patterns when absolute numbers are small. We therefore caution against over-interpreting low %TP values, especially when based on fewer than five counted spores per sample. While we were unable to calculate concentrations (e.g. influx) due to the absence of exotic marker grains in dung samples, we provide all raw count data and sample-level metadata in Supplemental Tables S1 and S2 to ensure transparency and enable further scrutiny or recalculation using alternative metrics.
Differences in fungal composition between dung samples suggest that coprophilous fungi may also be influenced by microenvironmental conditions associated with different herbivore species, including variations in dung moisture, microbial composition, and nutrient content (Perrotti and van Asperen, 2019). In deer dung, Sporormiella-type (HdV-113) was dominant, consistent with previous studies identifying it as a strong indicator of large herbivore presence (van Geel et al., 2011). However, our findings confirm that Sporormiella is also abundant in the dung of extant medium-sized herbivores, such as deer, which highlights the importance of ecological context when interpreting its presence in paleoecological records, especially concerning megafaunal decline (cf. Gill et al., 2009). Bison dung contained a higher proportion of Sordaria-type (HdV-55A) and Cercophora sp. (HdV-112), which may reflect differences in the dung microbiota, moisture content, and nutrient composition—factors known to influence fungal community structure in herbivore dung (Perrotti and van Asperen, 2019).
The observed variation in fungal diversity across dung types suggests that herbivore-specific differences in digestive processes, diet composition, and dung decomposition rates may shape CFS assemblages. One of the key challenges in analyzing CFS lies in their preservation across substrates and microenvironments. Not all coprophilous fungi are equally resistant to taphonomic degradation, and preservation may differ substantially between mosses, dung, and sedimentary archives (van Asperen et al., 2016). In mosses, spore retention and degradation are influenced by moisture levels and oxidation conditions, while in dung, microbial succession and rapid decomposition may selectively preserve more robust taxa.
These differences highlight the importance of considering both ecological and taphonomic factors when interpreting CFS records. In mosses, spores may accumulate gradually from both direct dung contact and passive airborne deposition, meaning their presence does not necessarily indicate recent or local herbivore activity. Rather, it may reflect an integrated signal shaped by environmental heterogeneity over time. Understanding these processes is essential for improving the reliability of CFS as indicators of herbivore activity in both modern and paleoecological contexts.
Although large herbivores such as European bison and red deer often favor open habitats for foraging, they also make regular use of forested environments for shelter and resting. As shown by Kuijper et al. (2014) and Bubnicki et al. (2019), considerable dung accumulation can occur even in closed-canopy forests, particularly in structurally diverse stands. This behavioral flexibility likely contributes to the presence of coprophilous fungal spores in forest-based samples.
Our interpretation aligns with findings by van Asperen et al. (2020), who used pollen traps and grazing experiments to explore how CFS are incorporated into surface archives. Their study emphasized the influence of animal behavior and environmental filtering on spore deposition and cautioned against overinterpreting low CFS abundances. The inclusion of three contrasting substrates in our study (moss, traps, and dung) reinforces the idea that CFS records are highly localized and shaped by both ecological dynamics and methodological factors.
The influence of environmental variables on the composition of coprophilous fungal spores in samples
The variability in the composition of CFS across different sample types was not well explained by the selected environmental variables: coarse woody debris (CWD), habitat wetness, canopy openness, total ungulate pressure (TUP), and forest management intensity (FMI). The lack of strong relationships suggests that these factors are not the primary drivers shaping CFS assemblages. Previous studies indicate that spore deposition is influenced by multiple ecological and environmental factors, including the type and composition of fecal material (Blackford and Innes, 2006), microclimatic conditions (van Asperen et al., 2016), and seasonal variations (Raper and Bush, 2009).
While NMDS analysis suggested some differentiation in fungal composition based on ungulate pressure and openness of the habitat, the results from RDA did not confirm the significance of these relationships. The explanatory power of the model was relatively low across all sample types, suggesting that additional factors, such as dung decomposition rates, soil properties, or microbial interactions, may play a more dominant role (Perrotti and van Asperen, 2019), but they were not considered in our study.
In moss samples, CFS composition appeared highly heterogeneous, with no strong correlation with local environmental conditions. The lack of significant relationships may reflect the passive nature of mosses as spore traps, accumulating fungal spores from various sources over time rather than serving as direct indicators of local herbivore activity (Sundberg, 2013). This could explain why variables such as openness or TUP were not strongly associated with CFS composition in moss samples.
To evaluate whether the inclusion of facultative or ecologically ambiguous taxa might obscure underlying ecological patterns, we repeated the NMDS and RDA analyses using only obligate dung fungi. The resulting ordination structures and environmental associations were similar to those observed in the full dataset. While the RDA for obligate taxa explained marginally more variance, it remained statistically non-significant. These findings suggest that the broader taxonomic scope did not introduce bias into the ordination results and that facultative taxa did not obscure key ecological signals.
In Tauber-type traps, the proportion of CFS was lower compared to moss and dung samples, and environmental variables explained only a minor fraction of the variance. The relatively low abundance of CFS in air traps suggests that wind dispersal is not a major factor in the distribution of these fungi, reinforcing the idea that deposition is primarily linked to direct substrate contact rather than long-range atmospheric transport (Krug et al., 2004).
Dung samples exhibited the strongest link to herbivore presence, yet even here, the impact of environmental variables was not straightforward. While some taxa, such as Sporormiella-type (HdV-113), were more abundant in areas with high ungulate pressure, the overall CFS composition showed a high degree of variability. This could be attributed to differences in dung decomposition rates, moisture content, and nutrient availability, which can shape the fungal community independent of broader environmental factors (Blackford and Innes, 2006; Perrotti and van Asperen, 2019).
An additional source of variability may be the relatively low number of CFS counted in many samples. As highlighted by van Asperen et al. (2020) and confirmed by Perrotti et al. (2022), dung fungal spore concentrations are often low, which increases uncertainty in relative abundance measures and may obscure ecological trends. In our study, spore counts were not standardized using exotic markers, and percentage values were calculated relative to total pollen, potentially amplifying random variation, especially for rare taxa. This methodological limitation could partially explain the weak associations observed between CFS composition and environmental variables, even in dung samples.
Stochastic factors, such as sudden shifts in wind direction, precipitation events, or fluctuations in humidity, may further obscure direct relationships between fungal spores and environmental variables (Aylor, 2003; Jackson and Bayliss, 2011). These unpredictable influences highlight the complexity of spore deposition processes and suggest that a multi-proxy approach, including soil chemistry, plant community analysis, and microbial composition, may be necessary to better understand the ecological drivers shaping CFS assemblages.
The role of mosses as passive spore traps and interpretational limitations
The observed patterns of CFS presence in moss samples suggest that moss polsters act as passive spore traps, accumulating CFS over time rather than directly reflecting recent herbivore activity. Mosses can retain spores through both direct deposition from dung sources and passive airborne accumulation, although the extent of the latter remains uncertain (Gjerde et al., 2020; Sundberg, 2013). Unlike sedimentary environments, where fungal spores may be embedded in stratified deposits, mosses function as open systems, continuously exposed to environmental influences that may affect fungal spore retention and composition.
Most of the coprophilous fungi detected in this study, such as Sporormiella, Podospora, Ascobolus, Saccobolus, and Sordaria, rely on active ballistic spore discharge, dispersing their spores over short distances to colonize fresh dung (Bell, 2005; Krug et al., 2004; van Asperen et al., 2022). Given these dispersal mechanisms, the presence of these taxa in moss samples suggests that fungal spores were primarily acquired through local deposition rather than long-distance transport. This aligns with previous studies indicating that CFS are rarely dispersed over long distances due to their relatively heavy, substrate-dependent dispersal strategy (van Asperen et al., 2016; van Asperen et al., 2022). The short-distance dispersal of CFS makes them, in our opinion, a reliable indicator of the presence of large herbivores in the direct vicinity of the sampling site, stressing their relevance for paleoecological analyses.
On the other hand, some taxa, such as Chaetomium sp. (HdV-7A) and Gelasinospora sp. (HdV-1), are not strictly coprophilous but frequently occur also on other decomposing organic matter (Richardson, 2001; Simões Calaça et al., 2020). Chaetomium sp. is predominantly saprobic and commonly found on plant debris and soil, while some species also inhabit dung, suggesting a broader ecological role of taxa belonging to this genus (Simões Calaça et al., 2020). While Chaetomium species are not highly specialized for a single substrate, their presence in dung highlights potential interactions with other coprophilous fungi and organic decomposition processes. Similarly, Gelasinospora species have been frequently isolated from dung samples, but their ability to colonize other organic substrates, including soil, suggests that their spores may be introduced into moss samples from both dung and decaying plant material (Piasai and Manoch, 2009). The capacity of these fungi to exploit multiple substrates complicates their interpretation as direct indicators of herbivore presence and emphasizes the need for a cautious approach in paleoecological reconstructions.
Another limitation in interpreting fungal spore assemblages in moss samples is the potential for differential spore retention and degradation. While mosses are effective at accumulating spores, they do not preserve them indefinitely. Exposure to fluctuating moisture levels, microbial degradation, and UV radiation can lead to the selective loss of fungal spores, favoring those with more resilient wall structures. Similar taphonomic processes have been documented in sedimentary environments, where spore preservation is strongly influenced by moisture, microbial activity, and oxidation (Carrión, 2002), and are likely to affect moss-derived archives in comparable ways. Additionally, the hydrophobic properties of certain spores, such as those of Sporormiella, may enhance their adherence to moss surfaces, potentially biasing the representation of coprophilous fungi in moss samples compared to their actual abundance in the environment (Raper and Bush, 2009).
Another consideration is the uncertain temporal resolution of fungal spore accumulation in mosses. Unlike direct dung sampling, which provides an immediate snapshot of fungal composition or Tauber-type traps collecting spores for the whole vegetative season, moss-derived spore assemblages may integrate fungal propagules over unknown extended periods. The degree to which mosses accumulate and retain fungal spores remains unknown and likely varies with environmental conditions, including rainfall frequency, wind exposure, and moss species characteristics. This temporal uncertainty complicates the use of moss-derived CFS as direct indicators of recent herbivore presence, emphasizing the need for cautious interpretation in both ecological and paleoecological studies (Perrotti and van Asperen, 2019).
Despite these challenges, the ability of mosses to accumulate CFS from multiple sources over time makes them valuable for detecting general trends in herbivore activity. However, to improve the temporal and ecological resolution of such records, future research should integrate complementary sampling approaches, such as direct dung sampling, sediment core analysis, and molecular methods, to validate interpretations derived from moss-based CFS assemblages.
This need for methodological triangulation is underscored by the work of Davies (2019) and Davies et al. (2022), who demonstrate that in peatland environments, CFS abundances are better interpreted as indicators of ecologically impactful grazing pressure than direct proxies for herbivore biomass. Their findings emphasize the importance of considering local vegetation sensitivity, site-specific taphonomic filters, and the limitations of individual proxies. These insights align with our conclusion that moss-based CFS assemblages reflect not only local herbivore presence, but also environmental filtering mechanisms that influence spore retention and composition.
While our study focuses on wild herbivores in forest ecosystems, the taphonomic patterns described here may also inform interpretations of CFS assemblages in agro-pastoral contexts. The morphological similarity between deer pellets and those of sheep or goats, and between bison and cattle dung, suggests potential analogies in depositional processes. Nevertheless, differences in land-use intensity and animal behavior under domestication should be carefully accounted for when applying these results to archeological or paleoecological reconstructions.
Ecological and paleoecological significance of coprophilous fungal spores
From a paleoecological perspective, our findings emphasize the importance of cautious interpretation when using CFS as indicators of past herbivore activity. The potential for spore retention and deposition from multiple sources over time means that paleoecological reconstructions based solely on fungal spores in moss-derived deposits may overestimate the continuous presence of herbivores (Baker et al., 2013). Previous studies on sedimentary fungal records have shown that long-term spore accumulation can distort signals of past ecological conditions, particularly when deposition processes are influenced by environmental variability (Baker et al., 2013; Carrión, 2002). It must be considered that in our study, we sampled mostly terricolous mosses, which grow in environments more intensively used by herbivores, while paleoecological deposits are mostly derived from peat mosses growing in wet environments inaccessible or unfavorable to large herbivores. Therefore, the Tauber-type traps collecting CFS due to the long-distance dispersal are probably the closest proxies of paleoecological sampling.
The results of the NMDS ordination conducted on all sample types (moss polsters, Tauber-type traps, and dung) further support this conclusion. All three sample sets showed a high degree of compositional similarity but differed in the level of variation in CFS composition, which was the highest in dung samples, lower in moss polsters, and the lowest in Tauber-type traps. This gradation likely reflects the nature of each sampling method. Dung provides a direct substrate for fungal development, whereas moss polsters act as passive collectors, influenced by both airborne spores and occasional contact with dung. Tauber-type traps, designed to capture only airborne material, recorded the narrowest spectrum of CFS. Notably, in the NMDS ordination space, Tauber-type trap samples clustered within the distribution of moss polsters, and both were encompassed by the broader scatter of dung samples. This spatial arrangement suggests that both moss polsters and Tauber-type traps reflect herbivore dung availability, though to a limited extent and by different mechanisms. In our opinion, the results from the Tauber-type traps reflect this relationship in a manner more comparable to that found in paleoecological peat samples than do the moss polsters. This is because well-watered peat bogs, which produce sedimentary records, are rarely used by some large herbivores, such as the European bison, which tend to prefer fresh and moist deciduous forests and forest glades (Daleszczyk et al., 2007; Krasińska and Krasiński, 2007). Consequently, the CFS signal in peat sediments, like in the Tauber-type traps, is shaped primarily by airborne input rather than direct contact with dung.
Additionally, the taphonomic properties of CFS may introduce biases in paleoecological records. Spore degradation, selective preservation, and hydrophobicity of certain fungal taxa can all influence their detectability in sediment archives. For instance, spores of Sporormiella and Podospora possess thick cell walls, making them more resistant to decay than spores of more fragile taxa (van Asperen et al., 2016). In moss-derived deposits, the open structure of moss carpets may further influence spore retention, as spores may accumulate from both local and regional sources, potentially leading to mixed signals regarding past herbivore activity (Cundill, 1991).
If spores can be deposited both locally and through passive atmospheric accumulation, their presence in fossil records does not necessarily indicate constant herbivore activity in a given area. Furthermore, differences in spore transport and retention may lead to spatial variation in fungal assemblages, complicating direct correlations between fungal spore records and the past abundance of large herbivores. Consequently, multi-proxy approaches incorporating sediment core analyses, dung deposits, and plant macrofossils are necessary to improve the reliability of herbivore reconstructions (Gill et al., 2013; van Asperen et al., 2016).
Our data suggest that the presence of CFS in modern forest samples does not always directly reflect the density or ecological impact of large herbivores. In the case of the European bison (Bison bonasus), this may be related to its dietary flexibility and ability to cope with suboptimal habitats. Modern ecological studies have shown that European bison forage across a range of habitat types, selecting patches not only based on forage quality but also biomass availability, especially in nutrient-rich, wet forest environments (Kowalczyk et al., 2024). These findings support the view that European bison are not forest specialists, but rather habitat generalists with adaptive foraging strategies. This perspective resonates with paleoecological interpretations of fossil bison. For example, van Asperen and Kahlke (2017) demonstrated that Bison menneri from the Early Pleistocene type site of Untermassfeld exhibited a mixed-feeding strategy, exploiting both browse and more abrasive grasses depending on seasonal availability. Taken together, these observations suggest that even in modern closed forest systems, the occurrence of bison may not always produce a strong or consistent palynological signal of CFS. Their use of suboptimal habitats, variable dung deposition patterns, and flexible dietary ecology could all contribute to low detectability in spore assemblages despite physical presence.
Despite these limitations, CFS remain valuable indicators of herbivore presence when interpreted in the appropriate ecological context. To improve the reliability of CFS as paleoecological proxies, their interpretation must account for microhabitat variability, spore transport dynamics, and taphonomic processes shaping their preservation.
Conclusions
This study provides a comprehensive comparison of coprophilous fungal spore (CFS) assemblages in moss polsters, Tauber-type traps, and herbivore dung, offering novel insights into their deposition dynamics, substrate specificity, and their utility as indicators in both ecological and paleoecological contexts.
Moss polsters exhibited the highest taxonomic diversity of CFS, supporting their role as effective passive accumulators of spores from local and potentially regional sources. In contrast, dung samples yielded the highest concentrations of CFS, reflecting direct production at the spore source and offering a reference point for assessing CFS diversity and abundance. Tauber-type traps consistently recorded the lowest CFS values, underscoring the limited role of long-distance wind dispersal for most dung fungi and emphasizing the importance of local substrate contact in spore deposition.
Despite clear compositional differences between sample types, multivariate analyses revealed no strong or statistically significant correlations between CFS assemblages and environmental variables such as coarse woody debris, habitat wetness, canopy openness, ungulate pressure, and forest management intensity. These findings suggest that stochastic deposition processes, dung decomposition rates, and microhabitat-specific factors, rather than broad environmental gradients, play a dominant role in shaping modern CFS distributions.
From an ecological perspective, our findings highlight the heterogeneity and substrate-dependence of CFS occurrence. While dung samples were dominated by typical obligate dung fungi (Sporormiella-type (HdV-113), Podospora-type (HdV-368), and Sordaria-type (HdV-55A)), moss samples incorporated a broader range of taxa, including facultative or ecologically ambiguous forms (e.g. Chaetomium sp. (HdV-7A), Gelasinospora sp. (HdV-1)), indicating temporal integration and exposure to diverse ecological inputs. This underlines the complexity of interpreting modern CFS records and the need to consider both ecological traits of fungi and taphonomic pathways affecting their deposition and preservation.
From a paleoecological perspective, these results emphasize the importance of cautious interpretation when using moss-based fungal spore records to infer past herbivore presence. While stratified peat cores composed of mosses can provide temporally resolved archives, the surface moss samples analyzed in this study do not capture a real-time snapshot of fungal deposition but rather reflect the gradual accumulation of spores over an undetermined period. Consequently, the presence of CFS in such modern moss samples reflects an integrated signal rather than a precise temporal resolution. Additionally, taphonomic factors such as spore degradation, selective preservation, and taxon-specific dispersal mechanisms may bias fungal assemblages in both modern and fossil samples. However, this temporal integration may also be beneficial in a paleoecological context, as it helps to smooth out short-term fluctuations and provides a more robust signal of long-term herbivore activity patterns.
Further research is needed to refine the use of CFS as ecological and paleoecological indicators. Key questions include the duration of spore retention in moss carpets and how this influences their reliability in paleoecological reconstructions; the role of microhabitat factors in spore abundance and preservation; and whether molecular methods could enhance taxonomic resolution and improve identification of fungal spores in both modern and fossil material. Comparative studies across different ecosystems would also clarify how CFS deposition patterns vary with climate and vegetation. Addressing these challenges will enhance the reliability of CFS-based reconstructions and deepen our understanding of herbivore-ecosystem interactions over time.
Supplemental Material
sj-docx-1-hol-10.1177_09596836251366202 – Supplemental material for Following the spores: insights from mosses, pollen traps, and dung into coprophilous fungal spore deposition
Supplemental material, sj-docx-1-hol-10.1177_09596836251366202 for Following the spores: insights from mosses, pollen traps, and dung into coprophilous fungal spore deposition by Marcelina Zimny and Bogdan Jaroszewicz in The Holocene
Supplemental Material
sj-docx-2-hol-10.1177_09596836251366202 – Supplemental material for Following the spores: insights from mosses, pollen traps, and dung into coprophilous fungal spore deposition
Supplemental material, sj-docx-2-hol-10.1177_09596836251366202 for Following the spores: insights from mosses, pollen traps, and dung into coprophilous fungal spore deposition by Marcelina Zimny and Bogdan Jaroszewicz in The Holocene
Supplemental Material
sj-docx-3-hol-10.1177_09596836251366202 – Supplemental material for Following the spores: insights from mosses, pollen traps, and dung into coprophilous fungal spore deposition
Supplemental material, sj-docx-3-hol-10.1177_09596836251366202 for Following the spores: insights from mosses, pollen traps, and dung into coprophilous fungal spore deposition by Marcelina Zimny and Bogdan Jaroszewicz in The Holocene
Supplemental Material
sj-docx-4-hol-10.1177_09596836251366202 – Supplemental material for Following the spores: insights from mosses, pollen traps, and dung into coprophilous fungal spore deposition
Supplemental material, sj-docx-4-hol-10.1177_09596836251366202 for Following the spores: insights from mosses, pollen traps, and dung into coprophilous fungal spore deposition by Marcelina Zimny and Bogdan Jaroszewicz in The Holocene
Supplemental Material
sj-docx-5-hol-10.1177_09596836251366202 – Supplemental material for Following the spores: insights from mosses, pollen traps, and dung into coprophilous fungal spore deposition
Supplemental material, sj-docx-5-hol-10.1177_09596836251366202 for Following the spores: insights from mosses, pollen traps, and dung into coprophilous fungal spore deposition by Marcelina Zimny and Bogdan Jaroszewicz in The Holocene
Supplemental Material
sj-xlsx-6-hol-10.1177_09596836251366202 – Supplemental material for Following the spores: insights from mosses, pollen traps, and dung into coprophilous fungal spore deposition
Supplemental material, sj-xlsx-6-hol-10.1177_09596836251366202 for Following the spores: insights from mosses, pollen traps, and dung into coprophilous fungal spore deposition by Marcelina Zimny and Bogdan Jaroszewicz in The Holocene
Supplemental Material
sj-xlsx-7-hol-10.1177_09596836251366202 – Supplemental material for Following the spores: insights from mosses, pollen traps, and dung into coprophilous fungal spore deposition
Supplemental material, sj-xlsx-7-hol-10.1177_09596836251366202 for Following the spores: insights from mosses, pollen traps, and dung into coprophilous fungal spore deposition by Marcelina Zimny and Bogdan Jaroszewicz in The Holocene
Footnotes
Acknowledgements
We are grateful to the reviewers for their thoughtful and constructive feedback, which significantly improved the clarity and quality of the manuscript. We also thank Grzegorz Zimny for his valuable assistance in fieldwork and laboratory processing.
Author contributions
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was financed by the National Science Centre, Poland (research project no. 2020/37/B/NZ8/02803).
Ethical considerations
Not applicable. This study did not involve human participants, human data, or animal experimentation requiring ethical approval.
Data availability
Data will be made available on request.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.