
Abstract
Modern pollen rains have demonstrated that human legacies represent crucial information on the performance of strategies for biological conservation. However, the potential use of pollen rains in aspects related to human disturbance and other cultural processes remains scarcely explored in forest patches of tropical mountain environments. Our study offers an ecological model for stratifying management intensities using moss polster contents in three patches of humid tropical mountain forests from eastern Mexico. Moss polsters were collected at 48 sites between ca. 1600 and 2000 m asl in three Mexican tropical montane cloud forests. Beta diversity, Shannon, and Jaccard indices were calculated for each location. Deciphering palynological data, historical documents, and diversity indicators, we found that the most disturbed patches had the lowest values in the Shannon index (abundance data) and beta diversity (incidence and abundance data) due to intensive agriculture over the last 20 years at least. On the other hand, we found that lower-elevation gradients with a high degree of management intensity and geographical affinity expressed greater similarity according to the Jaccard index for unseen shared species. In contrast, shade-tolerant plants and some hygrophilous trees within the old-growth forest determined the species turnover among higher-elevation gradients in conserved areas. For these reasons, our findings suggest that moss polster contents capture reliable sources of pollen rains around 20 years, as it was once mentioned in the palynological literature. We conclude that pollen rains and management intensity are comparable between ecosystems in long-term approaches and historical legacies.
Introduction
The study of modern pollen rain is essential for interpreting palynological records of Quaternary plants. Different pollination mechanisms, transport, deposition, and preservation of diverse plant species have been studied worldwide (e.g. de Nascimento et al., 2015). Modern pollen studies have also explained different climatic and anthropic parameters, including richness and diversity in distinct altitude gradients, biomes, and ecosystems (Bhattacharya et al., 2011; Correa-Metrio et al., 2011; Domı́nguez-Vázquez et al., 2004; Franco-Gaviria et al., 2018; Islebe et al., 2001). Essential findings from these investigations have identified factors influencing tree cover during the Holocene through relative pollen productivity estimates among different cultural landscapes (Baker et al., 2016). In addition, pollen analysis in moss polsters has shown the influence of some climatic and terrain parameters on vegetation and pollen deposition in mountain ecosystems (Fontana et al., 2023). These approaches reinforce the relevance of pollen analysis and its implications within cultural landscapes and elevational gradients.
One of the most relevant contributions of pollen analysis revolves around the relationship between pollen deposition and modern-day vegetation with elevation, also considering the influence of climate and land use in the diversity patterns of the surrounding vegetation (Schüler et al., 2014). The diversity along an elevational gradient has been essential in Neotropical regions for developing topics related to land-use change, the early development of pre-Hispanic civilizations, and addressing other analytical approaches associated with the environmental history in Mexico during the Holocene (Caballero-Rodríguez et al., 2018; Caballero-Rodríguez et al., 2017; Lozano-García et al., 2007, 2013, 2014). Diversity analysis allows the evaluation of the representativeness of plant communities in mountain landscapes, among other aspects such as evenness, similarity measures, and richness due to anthropic factors (Castro-López et al., 2021; Jantz et al., 2014). As a result, some palynological efforts have focused on patch dynamics and forest trajectories under various levels of human disturbance for understanding neutral stochastic processes (random processes) from the Early to Late- Holocene (Birks, 2019; Jackson and Blois, 2015; Nogué et al., 2018).
However, information on pollen rain from humid tropical mountain ecosystems in eastern Mexico is scarce despite potentially suitable sites for developing these analytical approaches. On the other hand, these palynological tools allow evaluation of some impacts that land use changes exert on the composition, diversity, and richness of plants that are important for the structure and function of these habitats. (e.g. Baker et al., 2016; Castro-López et al., 2021). Therefore, pollen analysis in mountain ecosystems has acquired great relevance in vegetation dynamics in the face of changes in land use, given the inevitable loss of moisture, because they are located in transition zones under anthropic activities with economic and cultural importance (Etienne et al., 2013; Ferro and Morrone, 2014; Mazier et al., 2009).
Our research offers an ecological model to stratify different management intensities that are difficult to distinguish by employing maps that represent changes in land cover as reflected on the pollen spectra and historical records (Bürgi et al., 2017). These subtle, almost imperceptible changes on scales of centuries and millennia may or may not co-occur under different management intensities. They are frequently ecological interactions between plants and some edaphic characteristics such as nutrient dynamics (Bürgi et al., 2017). In addition, these changes in land use or human legacies encompass direct effects on ecosystems, for example, in domesticated landscapes such as the case of Araucaria angustifolia in Brazil, the forests of Theobroma cacao and Inga edulis in the Amazon (Whitney and Cárdenas, 2017). In contrast, human legacies also indirectly influence ecosystems, such as in the case of some disturbances, since they modify ecological processes and the physical environment (Bürgi et al., 2017). These scientific topics are widely represented in the global literature, such as those paleoecological approaches that analyze the persistence of ecosystems under certain anthropogenic disturbances (Birks, 2019; Etienne et al., 2013; Mazier et al., 2009; Mensing et al., 2018; Nogué et al., 2018). Other types of indirect effects evaluated under the long-term approaches have included, for example, seed dispersal within fragmented forest mosaics (Bhagwat et al., 2012), changes in forest biomass due to human disturbance (Jeffers et al., 2015), and the formation of novel landscapes by sociopolitical priorities (Mensing et al., 2018). For these reasons, our research addresses ecological approaches related to spatial heterogeneity and the selection of palynological indicators in the landscape (Bürgi et al., 2017; Chao et al., 2005; Jost, 2007).
In the eastern region of Mexico, tree cover loss has modified the dynamics of ecosystems formed by different patches and fragments (Arroyo-Rodriguez et al., 2013). These approaches assume that species turnover in better-preserved patches will be higher than in those with a more significant loss of forest cover (Arroyo-Rodriguez et al., 2013; Ledo et al., 2009). For the above reasons, our main objective is to study the richness and diversity of pollen rains and other palynological indicators in three study sites along an elevational gradient in the tropical montane cloud forest in eastern Mexico. Our central hypothesis suggests that at higher management intensity, diversity will decrease in patches with greater loss of moisture indicators (Polypodiaceae, among others) and tree cover. Likewise, the palynological similarity between mountain transects will remain in patches and plots with greater geographical, disturbance, and altitudinal affinity. Therefore, we hypothesize that our study sites’ forest patches with lower ecological vulnerability will reflect a higher turnover rate in pollen assemblages (Beta diversity). Furthermore, our results will be essential to developing future research focused on current stochastic processes related to restoration and biological conservation, among other palynological issues (Bush et al., 2021; Correa-Metrio et al., 2011, 2013; Domı́nguez-Vázquez et al., 2004; Figueroa-Rangel et al., 2008; Jacobs, 1982; León-Carreño et al., 2019; Lozano-GarcÍa et al., 2005; Lozano-Garcia and Xelhuantzi-López, 1997).
Materials and methods
Study area
The studied tropical mountain forest is located between 21°24′ and 20°37′ (Figure 1). This area belongs to the Carso Huasteco biogeographical subprovince, located on the slopes of the Gulf of Mexico. It is the northernmost part of the Mexican eastern Neotropics. The best detailed historical records suggest the presence of hunter-gatherer groups (Serranos, Otomíes, and Chichimecas) during the Classic Period ca. 800 A.D. (Carabias-Lillo et al., 1999; Lorenzo-Monterrubio, 2001; Ortiz-Quijano et al., 2015). Within the region, we selected three priority humid tropical mountain gradients for conservation due to the existence of paleoendemic and neoendemic species. In addition, these areas provide hydrological ecosystem services to the adjacent cities and are threatened by global climatic change (Rodríguez-Ramírez et al., 2013). The vegetation in the area is mainly tropical montane cloud forests (TMCF). The tropical montane forest 1 (TM1) site has been recognized as an ecosystem of restricted and relictual distribution, mainly Fagus grandifolia subsp. mexicana (Ortiz-Quijano et al., 2015; Rodríguez-Ramírez et al., 2021). The TM2 and TM3 sites are characterized by characteristic/diagnostic cloud forest vegetation, represented by the genera Liquidambar, Pinus, Quercus, Carya, and Magnolia. Some of these localities have been included in conservation programs for payments, including environmental services (PES) and ecological tourism.
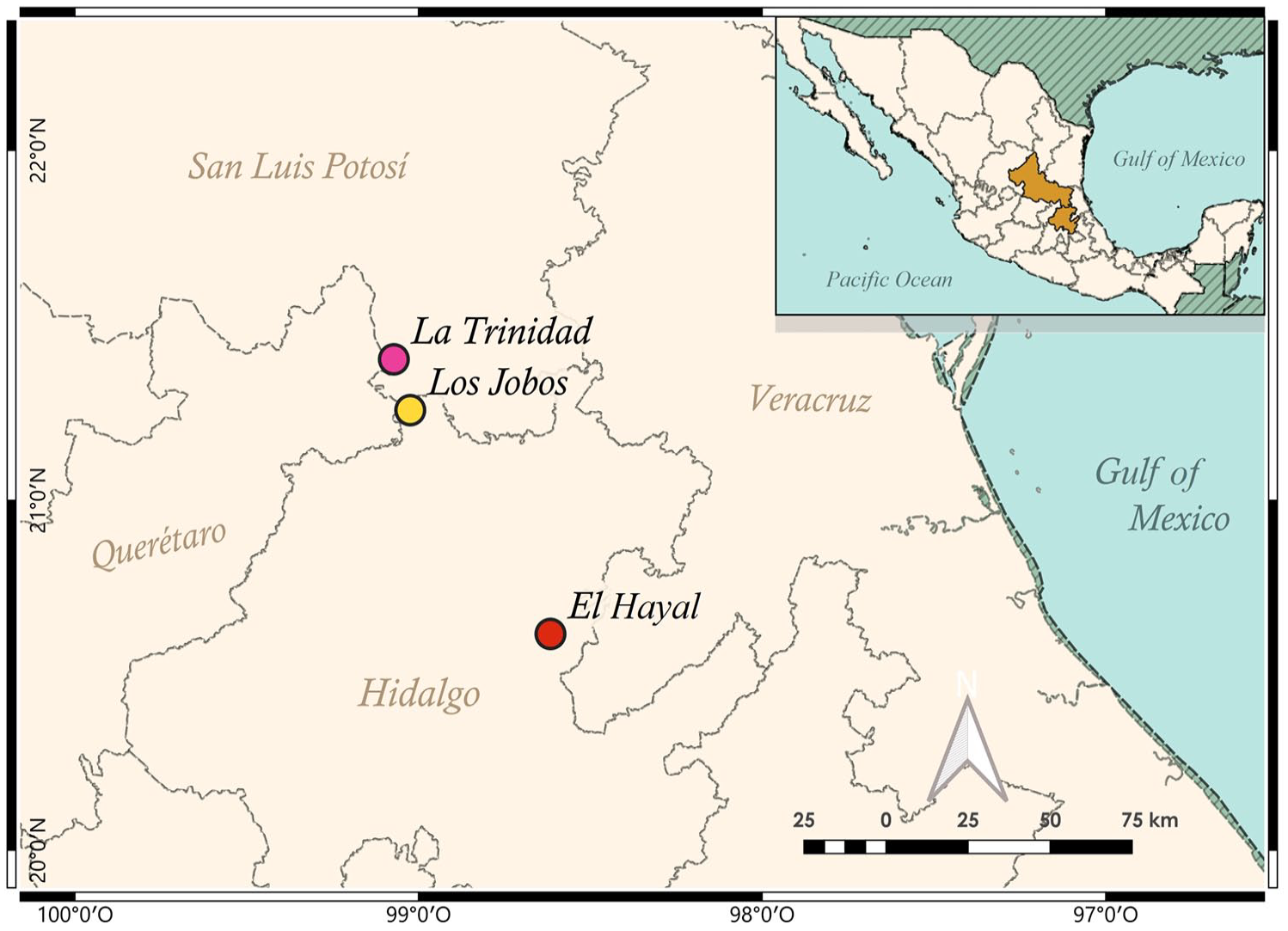
Pollen rain sampling points in three patches of the Mexican TMCF. (TM1) El Hayal, (TM2) La Trinidad, (TM3) Los Jobos.
Each site is characterized by physical elements of the environment and the inclusion of changes in land cover during the last 20 years (Table 1 and Figure 2). TM1 is a communal area with sites under steep slopes between 1000 and 2000 m asl. This area is located under high atmospheric humidity, and its average annual temperature ranges between 12°C and 23°C (Ángeles-Pérez, 2009). The predominant lithological material is Tertiary and Quaternary volcanic rocks, and the soils are mainly Vitric and Humic Andosol (Ángeles-Pérez et al., 2009). The fog regime occurs throughout the year, and the average number of cloudy days is 174, with an annual rainfall of 2047 mm (Ángeles-Pérez et al., 2009). The community promotes the current conservation status for scientific research.
Environmental and physical factors over the last 20 years in humid tropical mountain forests from eastern Mexico.

The tree cover loss and removal of tree canopy based on Global Forest Watch (2023).
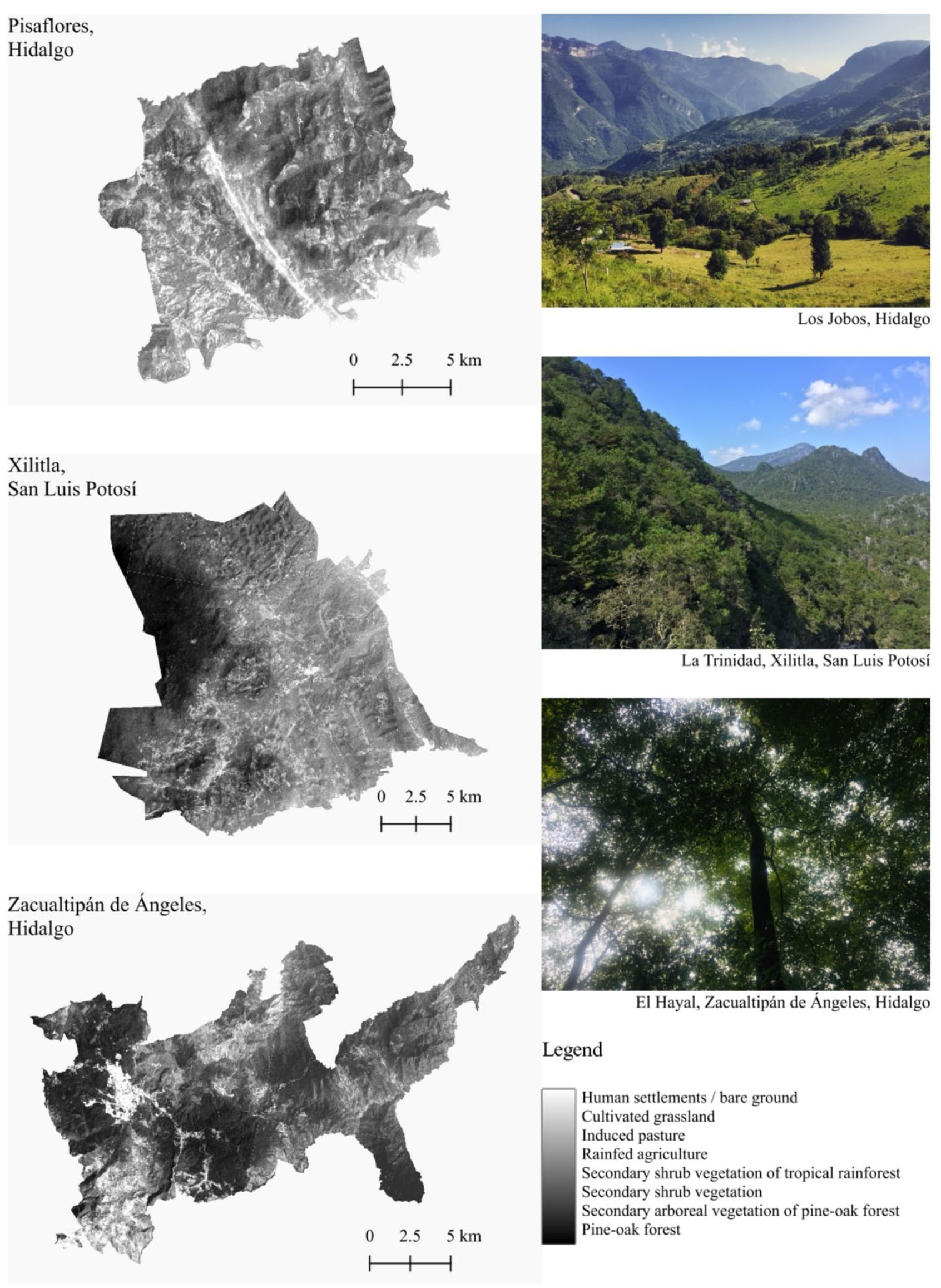
Satellite images for land cover and land use change from eastern Mexico’s montane environments in the three local municipalities sensu INEGI (2023).
TM2 is a communal area characterized by high mountains with highly abrupt slopes of folded strata and sedimentary rocks of karstic origin. Soils are predominantly Chromic Luvisols and Rendzins to a lesser degree (Reyes-Hernández et al., 2019). The heterogeneity of the relief and the altitude facilitate annual rainfall ranging from 1500 mm to 2500 mm almost yearly (Reyes-Hernández et al., 2019). The current conservation status comes from different initiatives, among which stand out the mountain ecotourism and the cooperation between indigenous mountain peoples living in La Huasteca of San Luis Potosí, Mexico.
TM3 is considered an “ejido” (communal land used for agriculture) located within areas of limestone rock and lutites into the following types of soils: Luvisols and Leptosols (INEGI, 2010). Precipitation usually oscillates between 1100 and 2600 mm per year with a predominantly semi-warm wet climate (INEGI, 2010). Land use is mainly conventional farming and livestock, and the management plans usually vary by altitudinal gradients or optimal areas for rural development. The region’s environmental history mentions that some physiographic characteristics between pine and oak forests, xerophytic vegetation, and tropical forests diversified the hunter-gatherers’ activities 500 years ago (B.P.) (Arroyo-Mosqueda, 2002). In addition, the area was characterized by supporting a type of transregional trade network due to the extraction of silver, lead, and animal production (Arroyo-Mosqueda, 2002).
Sampling of pollen rains and laboratory process
In 2019, we collected moss polsters within 48 plots of 20 × 20 m for spores, pollen, and charcoal particles (Cundill, 1991; Li et al., 2017). We located 16 plots of moss polsters at each forest patch with a difference of 100 m of altitude, between ca. 1600 and 2000 m asl. Due to the steep slopes and a typically closed canopy in the vegetation, four separate plots were placed to cover a forest area of approximately 300 m2 at each altitudinal level (Supplemental Figure 1). Moss polsters were collected in plots under a tree cover greater than 50% using a concave spherical densitometer. In plots located in zones with a tree cover of less than 30%, the moss polsters were collected from tree bark, rocks, and adjacent sites with forest patches and crops. We chose the dimensions of the plots following the studies done in other Neotropical mountain regions (Jantz et al., 2014; Lozano-García et al., 2014).
Samples were packed and labeled, including location, altitude, vegetaton type, sampling type, and conservation status (Table 2). Moss polster contents were placed in distilled water for 4 weeks, then decanted, sieved, and centrifuged (at least three cycles of 3500 rpm for 5 min for 1 week). We used a standard pollen processing (Crausbay and Hotchkiss, 2012) for quantifying ferns (e.g. Lycopodium, Polypodium, Pteris) within the moss polster in our forest patches. Alternatively, we used hydrochloric acid (HCl) and potassium hydroxide (KOH) to remove organic matter and carbonates from the sample. We avoided using hydrofluoric acid (HF) to preserve thin exine pollen grains like members of the CJT group (Cupressus, Juniperus, Taxodium). Subsequently, the obtained residues were acetolyzed, modifying the technique standardized by Faegri and Iversen (1989) with acetic acid (C2H4O2), acetic anhydride (CH3CO)2O, and sulfuric acid (H4SO) to dissolve the cellulosic material and to observe the pollen wall through a light microscope (Ardelean et al., 2020). We used a 400× magnification for pollen grains larger than 40 micrometers and psilate ornamentation in rapid counts. We used 1000× magnification in approximately 40 transects per lamella for smaller pollen grains. Because the study area lacks palynological data, we consulted botanical lists and palynological literature of central and southern Mexico in similar latitudes (Cartujano et al., 2002; Gómez-Noguez et al., 2013; López-Pérez et al., 2011; Monterroso-Rivas, 2009; Montúfar, 1985; Roubik and Moreno, 1991). A list of pollen taxa for each sampling site is shown in Supplemental Table 1.
Pollen rain recollection in TMCFs.
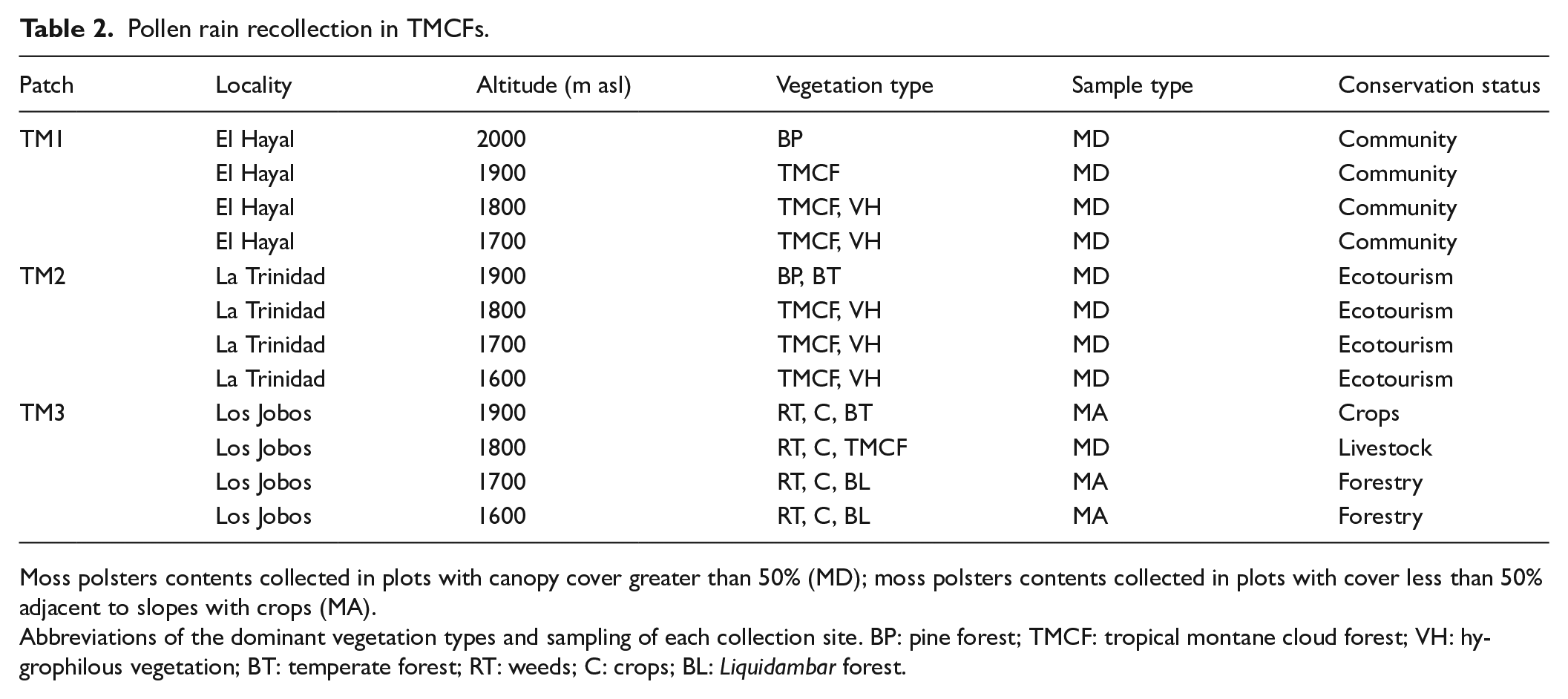
Moss polsters contents collected in plots with canopy cover greater than 50% (MD); moss polsters contents collected in plots with cover less than 50% adjacent to slopes with crops (MA).
Abbreviations of the dominant vegetation types and sampling of each collection site. BP: pine forest; TMCF: tropical montane cloud forest; VH: hygrophilous vegetation; BT: temperate forest; RT: weeds; C: crops; BL: Liquidambar forest.
In each sampled elevation (four plots), we counted at least 300 pollen grains represented as percentages of the total pollen sum. We grouped the obtained percentages into categories, for example, trees, herbs, shrubs, and fire indicators such as charcoal particles performed on the same pollen slides. In addition, we plotted the percentages of spores from the family Polypodiaceae as humidity indicators (Cartujano et al., 2002). Finally, we used the software TilialT: Tilia (v 1.7.16) (Grimm, 2011) to produce pollen diagrams.
Diversity analysis
We analyzed pollen richness and diversity using the taxon and assemblage turnover (the β component). In this analysis, we referred to plant communities only to those pollen assemblages related to recent stochastic processes and different vegetation types (Birks, 2019). To calculate the β diversity or community turnover of each patch under different management intensities, we used the beta diversity of order q, when (q ≠ 1), also known as hierarchical diversity analysis (Arroyo-Rodriguez et al., 2013), using the formula:
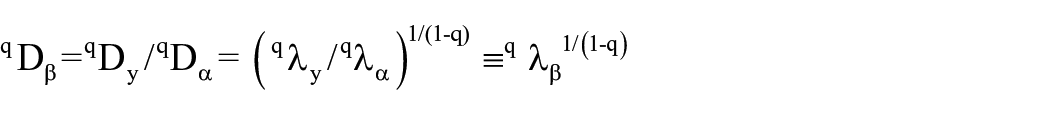
where (qλy / qλα)1/(1-q) corresponds to the sum of the incidences of the species among the total number of samples. Each study site’s alpha, beta, and gamma diversity components were obtained independently using the multiplicative method (Jost, 2007). We constructed incidence matrices for this analysis since this method is insensitive to abundances and compatible with plants that produce different amounts of pollen. Because of these reasons, the value obtained is equivalent only to species richness (Pereyra and Moreno, 2013). In addition, we included a Detrended Correspondence analysis (DCA) to quantify ecological changes from pollen records through R 4.3.1 (R Core Team, 2023).
We also use the traditional Shannon index (Jost, 2007) to measure the diversity of pollen assemblages under different land uses with abundance matrices, following the formula:

where n1 represents the number of individuals of species i, and N the number of individuals of all species, this index has been used to compare the diversity in pollen rains in central Mexico (Castro-López et al., 2021). The value can vary from 0 for communities with a single taxon to 5 with more species (Castro-López et al., 2021). According to the global literature, these values were obtained to corroborate hypotheses under different historical legacies.
To know the degree of similarity between the gradients of our study areas, we employed the Jaccard index based on species abundance data using pollen rains. This approach estimates unseen species (species likely present in a larger homogeneous association sample but missing from the sample data) (Chao et al., 2005). Furthermore, the analysis assumes that two random individuals, one from each of the two samples, belong to any species shared by the two samples and not necessarily to the same shared species (Chao et al., 2005). Therefore, the following formula was used to estimate this index:

where the mathematical expression
Results
Palynological indicators in moss polster contents
A total of 41 species were found in the 48 samples, and we counted an average of 300 pollen grains per sample. The palynological diagram shows each studied area’s abundance and main ecological indicators. We identified some biological indicators, such as trees and grasses, among other elements, including charcoal particles. We gathered and plotted those species recorded with a percentage greater than 5%. We compared these taxa with other moisture and disturbance indicators at different altitude gradients (Figure 3).

Diagram of pollen percentages in three locations of humid tropical montane cloud forests from eastern Mexico.
TM1 (El Hayal)
The patch (TM1) showed pollen abundances higher than 20% in the genera Pinus, Alnus, Quercus, and Magnolia. In elevations of 2000 m asl, the trees are mainly pine forests, while genera such as Tilia and Alnus dominated the lower elevation gradients. These genera can be found in hygrophilous habitats or open and disturbed environments. In contrast, taxa with a percentage less than 20% were Ostrya, Carpinus, Ternstroemia, and Heliocarpus between 1800 and 1700 m asl, while Zanthoxylum and members of the Myrtaceae family between 1900 and 2000 m asl. Elements of the Polypodiaceae family increased between 1700 and 1900 m asl. On the other hand, charcoal particles remained above 1900 m of altitude.
TM2 (La Trinidad)
The patch (TM2) included 20 and 60% values in the genera Pinus, Quercus, and Juniperus. At 1600 meters m asl, genera such as Juglans, Carya, Tilia, and Magnolia dominated the assemblage, while percentages of spores from the Polypodiaceae family increased up to 40%. In contrast, the lowest percentages throughout the pollen assemblage were Ulmus, Clethra, Ostrya, and Magnolia. The values of trees prevailed throughout the altitudinal gradient, contrary to ruderal taxa, while charcoal particles increased at 1800 and 1900 m asl with a decrease located between 1700 and 1600 m asl.
TM3 (Los Jobos)
The patch (TM3) showed the highest values of the Chenopodiaceae-Amaranthaceae (Cheno-Am) group disturbance taxa at 1600 and 1700 m asl. In contrast, the species of Poaceae and Asteraceae families increased respectively at higher altitudes between 1800 and 1900 m asl. The grasses and ruderal taxa percentages were higher than the trees throughout the palynological diagram. The lowest percentages were the genera Quercus, Tilia, Cecropia, Sida, and some elements of Myrtaceae and Moraceae. The charcoal particles as disturbance proxies showed the highest values at all gradients, with the highest at 1700 m asl. The spores of fungi and ferns increased around 20% only from 1700 m asl, respectively.
Diversity and similarity indices
Management intensity was represented by the diversity of pollen rain values in each of the study areas and compared with other taxonomic indicators of disturbance (Table 3). The patches TM1 and TM2 showed the highest values of pollen richness (taxa found in each site) compared to the TM3 forest patch, with 11 pollen taxa (see Table 1 in Supplemental material). Similarly, the beta diversity decreased as management intensity increased. For example, while two communities in the palynological assemblage represented the TM1 patch, the other forest components (TM2 and TM3) obtained lower diversity values. However, the Shannon diversity index showed the highest value of 2.61 at La Trinidad, possibly due to the region’s high incidence of plant genera. Despite this, the lowest diversity value was represented at Los Jobos with a dominant ruderal vegetation type. Also, habitat specialist trees such as the genera Fagus and Magnolia dominated the pollen assemblages in the TM1 and TM2 forest patches.
Palynological indicators under different levels of disturbance: TM1 (community conservation), TM2 (ecotourism), and TM3 (agriculture, livestock, and forest management).

All the values and statistical indices include the pollen abundances of each analyzed patch.
Detrended correspondence analysis (DCA)
We obtained pollen axis length for our three sites to indicate species turnover (Table 4). The highest score was 2.28 SD in forest patch TM1, while the lowest was 1.46 SD and 1.77 SD in sites TM2 and TM3, respectively. However, TM3 was higher than the site TM2, perhaps because of species scores and richness (see Table 2 and Appendix B in the Supplemental material). The relative importance of axis 1 was greater in the three forest patches.
Detrended correspondence analysis results for pollen from the tree forest patches.
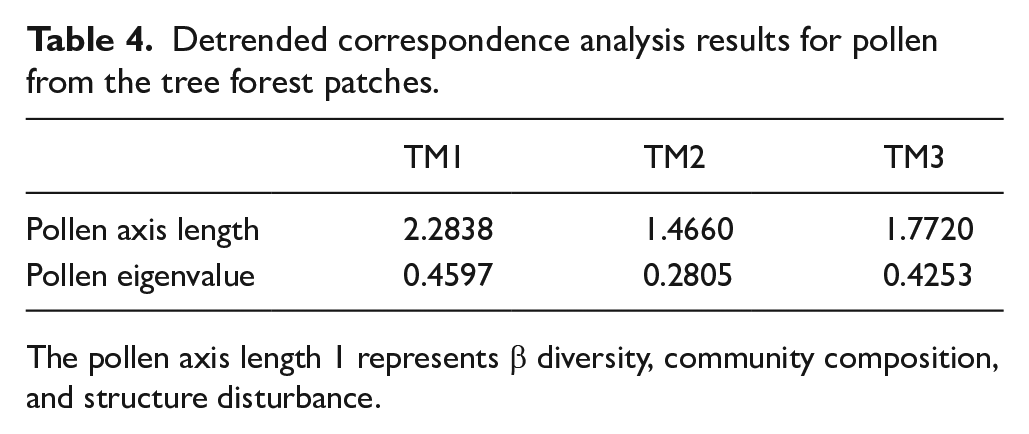
The pollen axis length 1 represents β diversity, community composition, and structure disturbance.
Differences in the open canopy and humidity indicators
The percentage of arboreal pollen was represented as the indicator of the open canopy and compared with the different levels of management intensity (Table 3 and Figure 4 ). In patches TM1 and TM2, disturbance taxa showed low abundances, in contrast to patch TM3 where the percentage of ruderal taxa was highest. The tree cover calculated with the percentage of arboreal pollen in each locality was higher than 50% in TM1 and TM2 concerning fragment TM3 (Los Jobos) because it was dominated mainly by ruderal taxa.
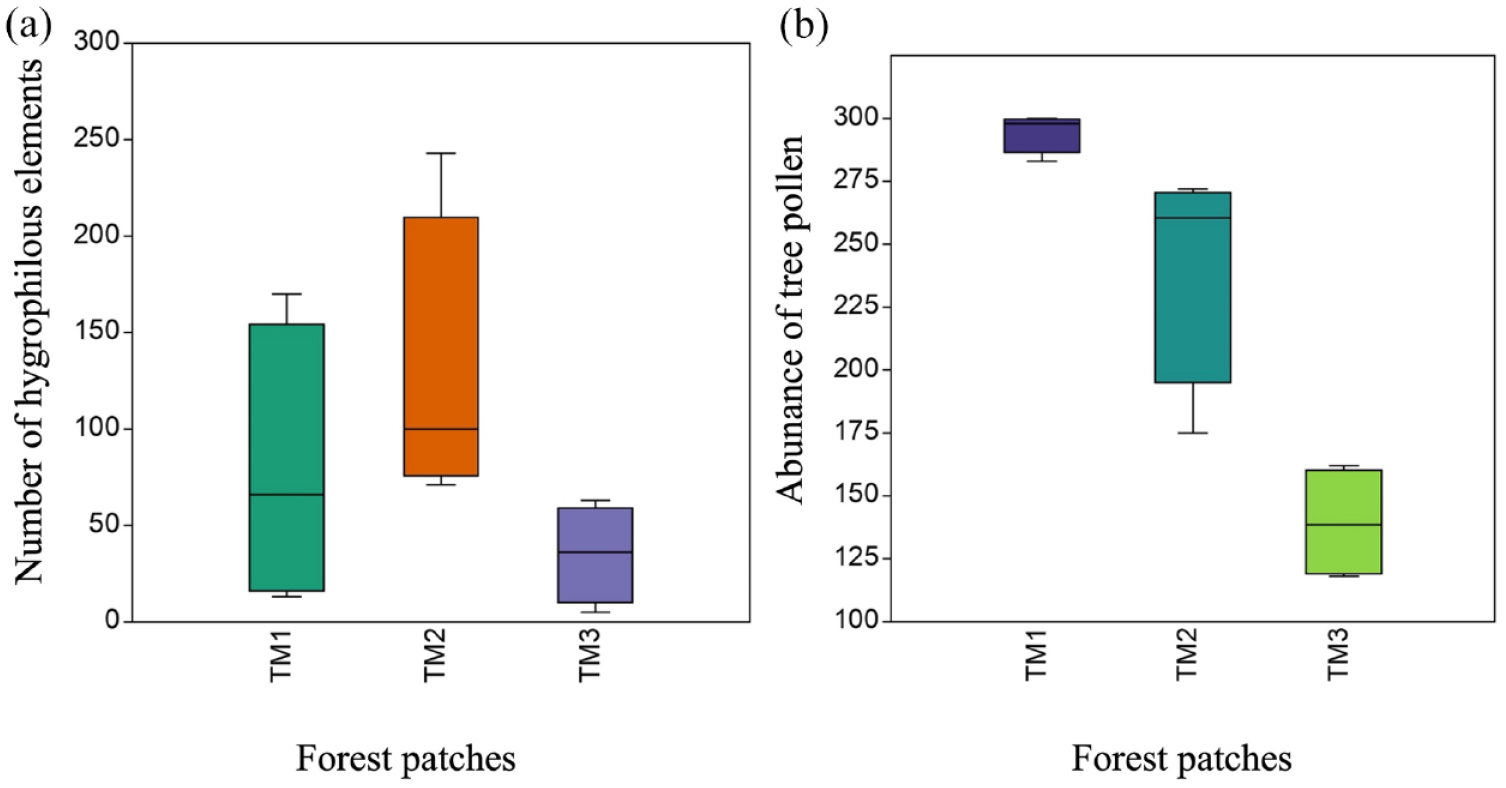
(a) Kruskal-Wallis boxplot among the number of hygrophilous indicators (X2 (1) =4.452, p > 0.05) in the environmental gradients of the patches under analysis. TM1 (El Hayal), TM2 (La Trinidad), TM3 (Los Jobos). (b) Kruskal-Wallis boxplot (X2 (1) =9.846, p < 0.05) of the open canopy represented by the abundance of tree pollen in the environmental gradients in the analyzed forest patches. TM1 (community conservation), TM2 (ecotourism), and TM3 (conventional farming and livestock).
Each patch showed differences in the distribution of values for both humidity indicators (ferns) and tree cover loss (Figure 4). For example, the highest presence of hygrophilous elements ranged between 150 and 200 pollen grains in La Trinidad and El Hayal, while Los Jobos maintained a lower range between 10 and 50 pollen grains. In addition, tree pollen as an indicator of canopy cover had differences among the three patches, with a higher degree of individuals in El Hayal and La Trinidad, in contrast to Los Jobos, with the lowest intervals.
Species turnover in elevational gradients and differential preservation
The Jaccard similarity index for unseen shared species threw three main groups regarding beta diversity (Figure 5). The first one was clustered by two elevations (1900 and 1800 m asl) in TM3 (Los Jobos) and the lower altitudinal transects (1800 and 1700 m asl) in La Trinidad. The most disturbed transects united the second group in Los Jobos (1700 and 1600 m asl). In contrast, the third group was clustered by all the transects in the TM1 patch (El Hayal) and two elevations in La Trinidad (1900 and 1600 m. asl). The analysis allowed the division of two main groups: a) the most disturbed but with geographic and environmental affinities (TM1 and TM2), and b) the most distant forest patches with higher conservation status.
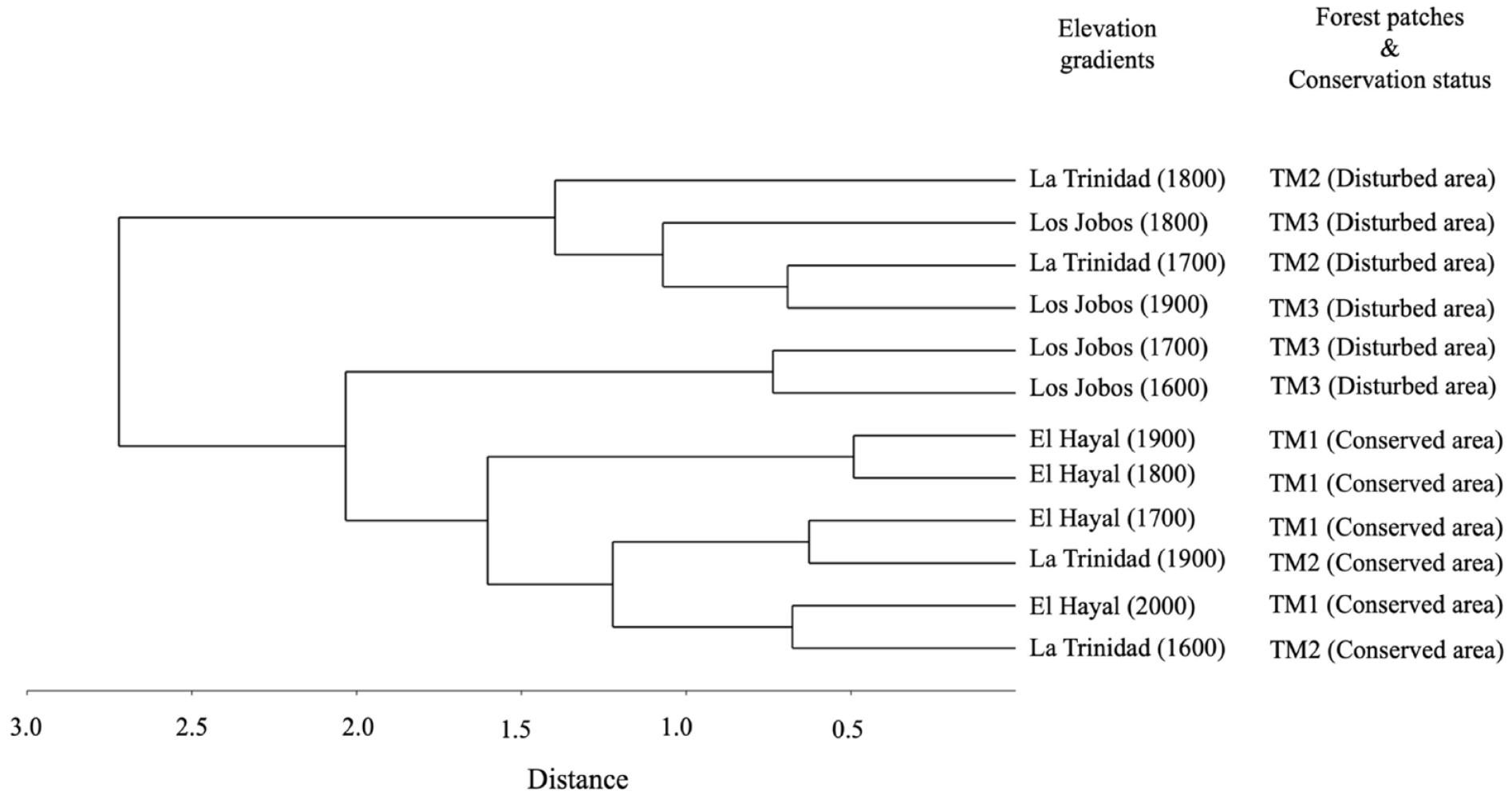
Dendrogram of similarity (Jaccard index for unseen shared species) and the new statistical approach of Chao et al. (2005). The gradients are expressed as m asl. and the patches are represented in the following order TM1 (El Hayal), TM2 (La Trinidad), TM3 (Los Jobos).
Discussion
Management intensity: Changes in structure and composition within forest patches
Currently, remote sensing offers the possibility of describing some structural and functional aspects of ecological systems for decision-making in conservation (Mihai et al., 2017) through the mapping of the biotic and abiotic components of ecosystems (land-cover, land-use, ecological disruption, and plant phenology). In the Neotropics, human impacts involving forest clearance, cultivation of Zea mays, and slash-and-burn activities have been documented elsewhere in Mexico and South America. There is little or no tree regeneration today because of sheep and deer grazing. If areas are fenced, the field layer and tree seedlings grow, but the ground layer’s rare bryophytes decline because of the field-layer expansion. The annual deforestation percentages estimated in the three study areas were higher than the tropical montane forest deforestation rate worldwide (−1.1) (Doumenge et al., 1995). However, the results were lower than reported in other regions of Mexico. Deforestation of TMCF has been as high as −1.95 in the western zone of the Trans-Mexican Volcanic Belt (RuIz et al., 2016), −3.84 in the Sierra Madre Oriental (Leija-Loredo et al., 2011), and −8.5 in the mountainous areas in the southeast of the state of Chiapas (Navarrete et al., 2010). In the Sierra Alta of Hidalgo, high deforestation values show an upsetting trend for the conservation of TMCF in Mexico. The above results indicate that losses in vegetation cover since 1976 were due to the expansion of agricultural activities in the region, with the establishment of paddocks for livestock being the most predominant reason, followed by an increase in rainfed agriculture and finally, with a minuscule proportion and in very localized areas, the growth of irrigated agriculture.
Recent detailed studies show that modern palynological richness and diversity (see Birks et al., 2016 for the distinction between richness and diversity in a palynological context) are, in part, a reflection of contemporary floristic α-richness and diversity (Felde et al., 2016), and, in part, a reflection of landscape mosaic structure (spatial β-diversity) (Matthias et al., 2015; Meltsov et al., 2013). Spatial β-diversity of pollen assemblages can be estimated by diversity partitioning for a particular time (Birks et al., 2016; Blarquez et al., 2014; Felde et al., 2016). Compositional turnover (temporal β-diversity) in fossil assemblages is commonly quantified as a rate-of-change (Birks, 2012) or turnover (Birks, 2006) metric. For example, the diversity values in our three sites showed a spatial difference due to the environmental characteristics in which the moss was collected (e.g. open canopy). However, the significance of these patches will also depend on compositional change (temporal β-diversity). In principle, historical records and satellite data demonstrated that forest patches TM2 and TM3 had the highest percentages of tree cover loss. This fact would have implications for understanding whether beta diversity has changed over time, for example, by estimating biomass through pollen accumulation rates in response to human activity (Seppä et al., 2009). On the other hand, forest clearance and agriculture on fertile soils increase α- and β-diversity, and biomass decreases as the complex mosaic landscapes develop (e.g. Peglar, 1993). In our model, these facts would corroborate whether high values like the Shannon index at site TM2 or DCA score at site TM3 were related to nutrient cycling or soil changes due to agriculture.
Forest patch (TM1)
All diversity indices in the forest patch (TM1) were the highest concerning the other study areas. The DCA value at the TM1 site (2.2 SD) was similar to the reported for this Neotropical region (2.6 SD). According to Correa-Metrio et al. (2014), this value found in southeastern Mexico reflects slower environmental changes due to the life cycle of trees. In addition, the values coincided with the high percentage of tree pollen compared to ruderal taxa at this site. For example, in our palynological assemblage, the incidences and high percentage of high pollen producers such as Pinus and Quercus were higher than disturbance species. This difference has been studied in other cultural landscapes worldwide, where the relationship between higher and lower producers increases considerably in conserved areas (Baker et al., 2016). Moreover, in the forest patch (TM1), the incidence of cloud forest trees such as Fagus, Magnolia, and Clethra was not shared with the most disturbed fragment. According to Global Forest Watch, this fact means that diversity values in the TM1 forest patch remain highest due to its conservation status, as seen with the tree cover loss. In contrast, the number of diagnostic incidences for secondary forest trees (Heliocarpus and Zanthoxylum) (Guariguata and Ostertag, 2001) and herbaceous plants in the TM3 fragment, such as Poaceae and Asteraceae dominated the herbaceous component of our palynological diagram. These results have suggested in the global literature that deforestation, the fallow period reduction, and a decrease in the extent of old-growth forests lead to the loss of richness, diversity, and resilience of ecosystems in the landscape (Jakovac et al., 2015). Furthermore, a crucial factor in interpreting our results is the high percentage of anemophilous pollen in patch TM1 (El Hayal) compared to patches TM2 and TM3. This is possibly due to silvicultural practices for pine regeneration in the face of disturbances, also known as (the seed-tree method) in adjacent areas (Figueroa-Navarro et al., 2010). This practice is favorable in sites where humidity prevails most of the year by fostering a lot of tree and shrub species of little or no commercial value (Figueroa-Navarro et al., 2010).
Forest patch (TM2)
The beta diversity, Shannon, and richness values in the TM2 forest patch were like the TM1 fragment. The patches with the highest conservation status showed more tree genera incidences and richness from old-growth forests. For example, the TM1 and TM2 forest patches presented a similar richness of arboreal taxa. This fact means that the two estimators (Shannon Index and Beta Diversity) would facilitate explaining changes between the richness and diversity of different vegetation types and in sites with several degrees of conservation and environmental disturbance. In addition, identifying habitat-specialist trees and the spores of some hygrophilous elements were determinants to characterize the sites with the highest degree of humidity, even though we did not find a significant difference in the number of spores among the three forest patches. In contrast, in the case of arboreal pollen, our results showed statistical significance in the environmental characterization for each of the different management intensities. This occurred in the forest patches TM1 and TM2, where the extent of old-growth forest is similar concerning the richness of native trees and the number of hygrophilous elements.
Forest patch (TM3)
In the forest patch TM3, some components, such as ruderal taxa and charcoal particles, encompass the greatest disturbance indicators throughout the palynological diagram. These data suggest that the low percentages of tree pollen in the TM3 patch and the increase in disturbance species adapted to greater amounts of light erode the beta diversity values in this case. Similar studies have shown that the homogenization of beta diversity may be related to the loss of shade-tolerant plants due to forest clearance (Arroyo-Rodriguez et al., 2013). Historical documents reveal a long history of disturbance during the last 500 years due to mining and agriculture-related activities, livestock, and illegal logging in this region (Arroyo-Mosqueda, 2002; Carabias-Lillo et al., 1999). In addition, relevant studies of the Late-Holocene in Mexico have shown the dominance of disturbance indicators of the Amaranthaceae family and the Chenopodiaceae group in habitats under similar anthropogenic conditions (Caballero et al., 2006). These results would explain the highest content of ruderal taxa, the lowest incidence of native trees, and the decrease in hygrophilous elements in the pollen rains from Los Jobos.
Other factors that could influence our pollen assemblages include canopy structure and light limitation, such as in some regions of Poland (Baker et al., 2016). For example, Baker et al. (2016) mention that the Poaceae flowering is expected to be higher in full light conditions where agriculture prevails, such as in the case of our most disturbed fragment. In other words, this means that the reduction of our diversity indices and the tree cover loss reflect the behavior of our palynological results. For these reasons, the diversity values of pollen rains for the three forest patches can corroborate the management intensities that co-occur, according to Global Forest Watch.
Even though our DCA scores were lower than those reported in southeastern Mexico ∼ 2.6 SD, 2.92 SD, 3.1 SD ∼ (see Correa-Metrio et al., 2014; Franco-Gaviria et al., 2018), our results suggest the great vulnerability of these transitional ecosystems (TMCF), particularly in steep mountains from eastern Mexico. Other approaches, such as Hill’s numbers, fail because they do not find a consistent relationship between palynological richness/diversity and plot vegetation richness/diversity due to differences in the open canopy, pollen source area, or variations in flower structure (Gosling et al., 2018). According to Gosling et al. (2018), palynological richness exceeds vegetation richness, while palynological diversity is lower than vegetation diversity. For these reasons, our study suggests that it is more appropriate to compare values based on Detrended Correspondence Analysis (DCA).
Species turnover in elevational gradients and differential preservation
According to the Jaccard analysis, two transects showed similarities among forest patches: a) the lower-elevation transects with a high degree of management intensity and geographical affinity, and b) some higher-elevation transects in conserved areas determined by shade-tolerant plants and some hygrophilous trees within the old-growth forest. These diversity patterns suggest that the similarity among all forest patches was related to management intensity, the degree of humidity of the study areas, and certain terrain parameters such as soils. For example, the percentages of anemophilous and arboreal pollen are significant in the conserved areas and at higher altitudes corresponding to TM1 and TM2 fragments. The charcoal particles and some herbaceous elements (disturbance indicators) are in a higher percentage concerning the patch (TM3) in our palynological diagram. In this context, previous archaeopalynological studies have exposed how the influence of some climatic, edaphic, and biotic factors can contribute to differential pollen preservation (Hevly, 1981). According to Hevly (1981), some anemophilous tree species, such as Pinus and Juniperus can increase their pollen proportions as altitude increases and the number of disturbances decreases.
Another essential factor in our study was the presence of shade-tolerant tree species and hygrophilous plants such as the genus Fagus, Magnolia, and Clethra in the forest patch (TM1). For example, Ángeles-Pérez et al. (2009) have shown that regeneration of shade-tolerant species such as Fagus grandifolia subsp. mexicana can survive and establish under limiting light conditions. However, this regeneration mechanism depends on a persistent reservoir of slow-growing seedlings, which can be found in the herbaceous and shrub strata (Ángeles-Pérez et al., 2009).
In contrast, a shared environmental history determined the similarity among lower-elevation transects with a high degree of management intensity and geographical affinity. For example, some elements of the Poaceae family, herbs, and tree species of the Cupressus, Juniperus, and Taxodium groups were shared more between these sites. According to historical data, since the beginning of the 16th century, the region known as Sierra Gorda Hidalguense promoted activities such as agriculture and livestock as main secondary economic sources (Arroyo-Mosqueda, 2002). Fontana et al. (2023) found that the relationship between Poaceae and their occurrence in plant communities is predominantly good in sites close to forest edges within an elevation gradient. This could explain the similarity among altitude gradients of the TM2 and TM3 patches due to some surrounding historical processes. For these reasons, the most relevant factors controlling vegetation distribution are identified in elevation (canopy structure), management intensity, and the degree of humidity.
Although the TM3 patch presented differences in the moss collection due to the quality, quantity, and type of sampling, the relationship between arboreal and non-arboreal pollen remained in the pollen diagram. Furthermore, despite the abovementioned sampling, the pollen preserve of patch TM3 recorded the main vegetation types in the area considerably with their respective abundances. Consequently, an important factor was modifying the chemical acetolysis technique since this method did not include HF. Therefore, it positively influenced the preservation of fungal spores in the three patches analyzed.
Even though our research did not capture information on spores of coprophilous fungi indicators of livestock and grazing, such as Glomus and Sporormiella (Mensing et al., 2018), our study was focused on specific aspects of taphonomic importance. Multidisciplinary mountain studies in Western Europe have verified the importance of current knowledge of pollen rains to reconstruct similar plant communities considering different historical sources (Mazier et al., 2009). These investigations recognize that aspects related to the edaphic characteristics of the sites and the type of paleoecological source are essential for interpreting statistical data. Such is the case of our study sites that, according to the Jaccard dendrogram corrected by Chao et al. (2005), we found that there is an edaphic difference between the acid-type pH in the gradients that make up the TM1 fragment concerning an alkaline-type pH in the TM2 and TM3 fragments.
Specific terrain parameters, such as soils, influence the differential preservation of pollen and spores. The high percentage and preservation potential of palynomorphs in the TM1 fragment of our palynological diagram can frequently be characterized by acid soils and humic profiles. For example, in an archeological context, especially in acid and humic soils without biological attack by earthworms, the incorporation and stratification of pollen and spores in the soil profiles are favored by humic colloids (Scaife, 2014). In this way, the downward movement of pollen and spores in such profiles is slow. Therefore, their release from those colloidal particles depends on various pedological conditions, favoring larger grains (Scaife, 2014). On the other hand, changes in land use that favor soil acidity can increase the pollen frequencies of recent profiles more than of older ones and soils with low microbial activity (Dimbleby, 1957; Scaife, 2014).
In this paleoecological context, calculating the absolute frequencies of the palynological spectrum is crucial for interpreting archeological data and those that need other approaches, such as peat bogs or lakes (Scaife, 2014). For these reasons, our study suggests complementary chemical tests in moss polsters to address aspects of taphonomic bias in mountain environments. Under these approaches, new lines of research can play a relevant role in the forest trajectories that have occurred in the last 500 years, mainly due to aspects related to natural disturbances and those of anthropogenic origin. Finally, our data yield the first palynological diagrams of the region in the same way that they can interpret some ecological conditions in each study site.
Implications for biological conservation
Patchy forest cover within a landscape mosaic represents a source of inspiration to guide the conservation and forest management of terrestrial ecosystems (Baker et al., 2016). According to Baker et al. (2016), specific paleoecological approaches highlight the importance of some natural forces to promote tree recruitment and regeneration of more light-demanding species into open forest gaps. In addition, under these conditions of disturbance in the landscape, it is possible to evaluate the synergistic effects between human activities and natural disturbances on the resilience of ecosystems (Bhagwat et al., 2012). In this context, validating specific disturbance hypotheses of past vegetation and continuous forest cover with a closed canopy is crucial in biological conservation (Baker et al., 2016).
Any attempt to reduce the extinction risk and, ultimately, biodiversity loss requires a better understanding of what the threats are, where they are, what were the baseline conditions, and how to minimize the risks (Chape et al., 2005; Forest et al., 2007; Joppa et al., 2016; Willis et al., 2007). Such constructs are highly relevant in management and restoration studies (e.g. Froyd and Willis, 2008; Jackson, 2012; Jackson and Hobbs, 2009; Landres et al., 1999; Wiens et al., 2012; Wingard et al., 2017) as conservation managers increasingly recognize that establishing realistic management goals requires knowledge of the range of natural variability not only of the vegetation but also of climate and disturbance regimes (Froyd and Willis, 2008). The “range of natural variability” concept can be directly linked to Quaternary ecological dynamics. The basic idea behind the concept is that “resilient ecosystems will be maintained if land management activities operate within the range of conditions that would be expected under the natural disturbance regime” (Froyd and Willis, 2008).
Our forest patches represent different environmental histories; this is the case of the broadly conserved TM1 site. However, sites TM2 and TM3 could present disturbance regimes that shifted over time. It is essential to understand whether conventional agriculture at the TM3 site exceeded the range of natural variability or if some agroforestry practices at the TM2 site favored ecosystem resilience. Legacies’ positive and negative effects must be considered to develop adequate management plans that adapt to these ranges of natural variability, on the understanding that both the age and the type of disturbance play a crucial role in the resilience of the TMCF.
In eastern Mexico, pollen analysis represents an opportunity for selecting essential biological indicators in the environmental history of tropical mountain ecosystems. This is possible due to the spatial heterogeneity and the continuous transformation of the landscape during the last centuries. Furthermore, these promising research areas can reveal the history of ancient agroforestry systems that persist until the present, also known as Tének milpa fields in La Huasteca Potosina (Heindorf et al., 2019). For these reasons, we suggest including these approaches in strengthening current palynological research from this Neotropical region (Jiménez-Zamora et al., 2023).
Conclusions
Our palynological results showed the relationship of the main mechanisms in the loss of tree cover for the three patches of humid tropical mountain forests: (a) management intensity, (b) land use changes due to sociopolitical priorities, and (c) human legacies. In addition, our results based on incidence and abundance matrices shared similar values; for example, we found that beta diversity (incidence and abundance data) and Shannon diversity index (abundance data) decreased due to intensive agriculture. On the other hand, this study found some biological, environmental, and edaphic components of the ecosystems and particular pollen assemblages in sites under different land uses and separated between 0 and 152 km away. These facts suggest that in some aspects of biological conservation, moss polster contents represent similar sources of historical information for around 20 years. Likewise, the standardized acetolysis technique can be essential in environmental studies without prior palynological information. This must be considered to preserve the greatest amount of pollen under certain differential preservation conditions and several disturbances. Furthermore, this approach is a methodological advantage because it produces reliable pollen percentages in moss samples at different points.
Our results represent the first pollen analysis in eastern Mexico’s humid tropical montane cloud forests and show the trajectories of three forest patches and some elements related to disturbances in our palynological diagrams. For instance, we explained how applying similarity analysis, such as the Jaccard index, could carefully provide related aspects of taphonomic bias. The proposal discussed and developed in the present study could be used in paleoecological studies with approaches in differential preservation and historical legacies. Birks (2019) recognizes that biological communities should be interpreted by their ecological attributes and the entire abiotic environment in both biomes and ecosystems. Jackson and Blois (2015) emphasize the importance of biological communities as a link between phylogenetic biogeography and modern ecology, especially for Quaternary studies. For these reasons, we consider including historical archives, palynological indicators of management intensity, and implementing some aspects of taphonomic bias crucial within a patch dynamics approach and other current topics such as conservation biology.
Supplemental Material
sj-pdf-1-hol-10.1177_09596836231219471 – Supplemental material for Modern pollen rain in humid tropical mountain forests from eastern Mexico: A patch-dynamics approach
Supplemental material, sj-pdf-1-hol-10.1177_09596836231219471 for Modern pollen rain in humid tropical mountain forests from eastern Mexico: A patch-dynamics approach by Tonatiuh Jiménez-Zamora, David Espinosa, Leopoldo Galicia, Irán Rivera-González and Isolda Luna-Vega in The Holocene
Footnotes
Acknowledgements
We acknowledge Ernesto Chanes Rodríguez, Othón Alcántara, Julio César Ramírez, Anaid Mora Villa, Andrea Martínez Reséndiz, and Auda Garcia for helping us in the field. A special acknowledgment to La Trinidad Cabins for their hospitality in the Mexican cloud montane forest of the Huasteca Potosina.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Three anonymous referees made valuable suggestions in the manuscript. This scientific paper is part of the requirements for obtaining a Doctoral degree at the Posgrado en Ciencias Biológicas, UNAM. DGAPA PAPIIT IN2223218 financed this project. LTJZ received funds from CONAHCYT No. 748412 for his doctoral research.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.