
Abstract
Organisms have evolved internal biological clocks to regulate their activities based on external environmental cues, such as light, temperature, and food. Environmental disruption of these rhythms, such as caused by constant light or frequent light schedule changes, has been shown to impair development, reduce survival, and increase infection susceptibility and disease progression in numerous organisms. However, the precise role of the biological clock in host-parasite interactions is understudied and has focused on unnatural host-parasite combinations in lab-adapted inbred models. Here, we use the natural interaction between monarch butterflies (Danaus plexippus) and their virulent protozoan parasite, Ophryocystis elektroscirrha, to investigate the effects of constant light and frequent light schedule changes on development, survival, and parasite susceptibility. We show that constant light exposure slows the monarchs’ rate of development but does not increase susceptibility to parasitic infection. Furthermore, frequent schedule changes decrease parasite growth, but have no effect on egg-to-adult survival of infected monarchs. Interestingly, these conditions are usually disruptive to the biological clock, but do not significantly impact the clock of monarch larvae. These unexpected findings show that constant light and frequent schedule changes can uncouple host and parasite performance and highlight how natural relationships are needed to expand our understanding of clocks in host-parasite interactions.
Circadian clocks have a biological periodicity of roughly 24 h and have evolved to allow organisms to synchronize their physiological and behavioral responses with the Earth’s daily rotation (Dunlap, 1999). These rhythms have been found across various species from bacteria to plants and from insects to mammals (Johnson et al., 1996; Reppert and Weaver, 2002; Froy et al., 2003, Tataroglu and Emery, 2014; Cohen and Golden, 2015). All levels of biological organization within an organism, from gene expression to immune function, behavior, and seasonal changes in morphology and development, are subject to circadian regulation (Dunlap, 1999; Denlinger et al., 2017; Scheiermann et al., 2018). The importance of the biological clock has been shown in studies that disrupted the clock. Multiple research studies in mammals and flies have revealed that such disruption can lead to severe biological consequences. For example, flies exposed to jet lag survived only 60 days compared with 80 days for controls (Pittendrigh and Minis, 1972). Circadian disruption also impacts host-parasite infections and infectious disease outcomes. In mice, frequent schedule changes increase proinflammatory cytokine expression and increase mortality when mice are exposed to bacterial endotoxin (Castanon-Cervantes et al., 2010; Adams et al., 2013). Furthermore, disrupting mouse rhythms enhances herpes and influenza A infections (Edgar et al., 2016). Principally, disruption of circadian rhythms causes impairment of the mouse immune response, resulting in increased susceptibility to infections with pathogens such as Salmonella (Bellet et al., 2013). Similarly, flies lacking circadian rhythms in locomotor activity show enhanced sensitivity to Streptococcus pneumoniae and Serratia marcescens infections (Stone et al., 2012).
Although existing studies suggest that disruption of the biological clock reduces health and survival, while increasing pathogenic infections, this common knowledge is heavily biased to a small number of species. Importantly, these species are mostly inbred lab-adapted models. When studies have focused on parasite infection, the studied host-parasite interactions are mostly unnatural, such as thicket rat malaria parasites studied in laboratory mice (Jaffe et al., 1990). Another issue is that most studies have focused on nocturnal animals, which may be poor reflections of diurnal situations. Yan et al. (2020) showed diurnal animals are not simply reversals of nocturnal animals but have distinctly different rhythms and behaviors, thereby highlighting the need for more circadian studies to include diurnal animals. Thus, while the use of traditional models has increased our knowledge in chronobiology, it has been strongly limited to inbred nocturnal animal models. Therefore, our study used outbred diurnal animals and a natural host-parasite relationship to investigate the role of constant light and frequent schedule changes in development, survival, and parasitic infection.
We used Danaus plexippus (monarch butterfly) and its naturally occurring protozoan parasite, Ophryocystis elektroscirrha, to investigate the clock’s role in larval development, survival, and parasite susceptibility. The monarch butterfly is known for its long-distance fall migration in which it travels up to 4500 km from the eastern United States and southeastern Canada to overwinter in central Mexico (Urquhart, 1976). To make this arduous journey, the monarch butterfly uses a time-compensated sun compass to maintain southern flight which is regulated by its biological clock (Reppert et al., 2016; Shlizerman et al., 2016). Monarchs are prone to infection with O. elektroschirrha, which is transmitted when larvae ingest parasite spores on their host plants (milkweeds) (de Roode et al., 2008). The parasite replicates within larval and pupal tissues, and adult monarchs emerge with up to millions of spores on the outside of their bodies (McLaughlin and Myers, 1970). In many instances, adult monarchs do not properly emerge and suffer decreased fecundity, shorter life span, and impeded ability to migrate (Bartel et al., 2011; Altizer et al., 2015; Atlizer and de Roode, 2015). Importantly, studies in adult monarchs reveal that disrupting the adult monarch clock leads to blunted clock gene expression, changes in the timing of adult emergence from the pupa, and altered migration flight patterns (Froy et al., 2003). The established importance of the biological clock driving monarch biology and the occurrence of a natural debilitating parasite make monarchs ideal for studying the role of the biological clock in driving development, survival, and host-parasite interactions.
We reared monarch butterflies from eggs in two different experimental conditions—(1) constant light (or dark) conditions and (2) schedule changing conditions (i.e., a 6-h shifted light-dark schedule every 2 days)—to determine how constant light and frequent schedule changes would influence monarch development, survival, and parasite susceptibility. We used constant conditions to address any potential masking effects and then a common jet lag schedule to desynchronize the clock. We found that these conditions can impact the host and parasite in different ways. Contrary to the more common observations of circadian disruption in vertebrates, we reveal here that constant light and frequent schedule changes do not increase susceptibility to O. elektroscirrha infection. Interestingly, we observed that parasites in monarchs in constant darkness had increased growth without impacting survival and monarchs with frequent schedule changes had a decrease in parasite growth. This study highlights the need for circadian host-parasite studies to include natural host-parasite relationships and non-lab-adapted outbred animals.
Methods
Animals and Housing
Two experiments were conducted to investigate circadian disruption effects on host-parasite interactions. Monarch larvae were the outbred offspring of monarchs originally collected from St. Marks, Florida. In our study, we used n = 6-10 for the constant conditions experiment and n = 19-23 for the frequent schedule change experiment. Eggs were laid on Asclepias incarnata plants. These plants were grown in the Emory University green house.
Constant Conditions Experiment
Eggs were placed in one of three conditions: light-dark (LD) controls (0700 h to 2100 h), constant light (LL), or constant darkness (DD). Monarchs remained in these conditions for their entire life cycle. All monarchs were housed individually in clear 16-ounce solo cups with the temperature maintained at 25 °C in three identical circadian light- and humidity-controlled incubators with cold-cathode compact florescent light bulbs that produce white light (with roughly 400 lumens experienced by individual monarchs) (Tritech Research, Inc). A digital thermometer was placed inside each incubator to monitor internal temperature. No significant temperature changes in lights-on or lights-off were recorded. Temperature only changed by ±1 °C. Larvae were fed ad libitum with A. incarnata leaf cuttings.
Schedule Change Experiment
Eggs were placed in one of two conditions: controls had a normal 14 h of light and 10 h of darkness. The schedule change (SC) group had their lights turned on 6 h earlier every 2 days until the pupal stage (~12 days following egg hatching) because O. elektroscirrha infection occurs during early larval development. After larvae reached the pupal stage, they returned to lights-on at 0700 h and lights-off at 2100 h. All larvae were housed individually on whole A. incarnata plants with clear plastic tubes (5-inch diameter × 22.5-inch height) and net coverings in two identical temperature- and humidity-controlled walk-in climate chambers at 25 °C, with TL835 fluorescent white light tubes (with roughly 2000 lumens experienced by individual monarchs). For each experiment, pupae were glued to the lids of new solo cups with hot glue, and each monarch was allowed to emerge in the solo cups. Monarchs were then placed in glassine envelopes and maintained in their respective conditions for the remainder of each experiment. Adults were not fed and life span was analyzed on starvation, in accordance with previous studies on this host-parasite system. Importantly, previous work has shown that life span under these conditions is correlated with life span under more natural conditions (de Roode et al., 2009).
Infection and Parasite Growth
For both experiments, a single parasite clone (E42-2) of O. elektroscirrha was used. On reaching the second instar, monarch larvae were inoculated with 10 parasite spores. To do this, a leaf disk was placed on a moist filter paper in a 10-cm-diameter Petri dish and the 10 parasite spores were placed on the leaf disk using a drawn-out glass capillary tube. Larvae were placed in the Petri dishes and kept there until they had completely consumed their leaf disks, and therefore all 10 spores. To determine the parasite infection status of adult monarchs, we conducted spore counts on all infected monarchs (constant conditions: LD n = 10, LL n = 10, and DD n = 6; schedule change conditions: controls n = 21 and SC n = 22). After adult death, the bodies were vortexed on a Vortex Genie in 5 mL of water for 5 min each to release the spores from the monarch’s body. The spores were counted on a KOVA glasstic slide. Four chambers per sample were counted and averaged.
RNA Extraction and Gene Expression
Constant Conditions RNA Extraction
Larvae were placed in LD (14 h of light and 10 h of darkness), DD, or LL. Cohorts of 4 to 6 larvae, third and fourth instars, were collected at 4-h intervals during a 24-h period. All samples were frozen in liquid nitrogen upon collection and placed at −80 °C until RNA extraction.
Schedule Change RNA Extraction
Larvae were placed in 14 h of light and 10 h of darkness with lights-on at 0700 h and lights-off at 2100 h for 7 days. After 7 days, cohorts of 6 larvae, third and fourth instars, were collected at 4-h intervals during a 24-h period. On day 8, larvae had a single schedule change where the lights came on 6 h earlier (i.e., lights-on at 0100 h and lights-off at 1500 h). After 24 h in this schedule, cohorts of 6 larvae, third and fourth instars, were collected in 4-h intervals during a 48-h period. All samples were frozen in liquid nitrogen upon collection and frozen at −80 °C until RNA extraction.
Quantitative Polymerase Chain Reaction
RNA was extracted from larval heads using a TRIzol protocol or RNeasy mini kit (Qiagen). Reverse transcription was conducted with high-capacity complementary DNA (cDNA) reverse transcription kit (Thermo Fisher). Housekeeping gene RP49 (Pan et al., 2015) and genes of interest cry2, tim, and per were analyzed. SSO advanced universal SYBR green (Biorad) was used to run quantitative polymerase chain reaction (qPCR). Primer efficiency was tested for near 100% for each primer set. Primer sequences were as follows: Ribosomal Protein 49 forward “CCG GAA GGT GTT AGT CCA CAA C”; Ribosomal Protein 49 reverse “CGG CGC AGT ACT TCC TAT TCT G”; Cry2 forward “GTT CAG GAA GGG TCT CAG ATT AC; Cry 2 reverse “GGC GAA CCA TGG GTC TAT AAT”; Tim forward “AGC CAC TTC GTA AAG GAA ATC A”; Tim reverse “TCC TGC AAC TCG CAC ATA AG”; Per forward “CCT GGC CAT CTT CAT CGT TAA T”; Per reverse “CAG AGG TCT CAA TCT GGG TTT C.” Individual reactions were used to quantify each RNA level in a given cDNA sample, and the average Ct from duplicated reactions within the same run was used for quantification. The data for each gene at a given time point were normalized to Rp49 as an internal control and normalized to the mean of 1 sample within a set for statistics.
Statistics
Most of the statistical tests were conducted with JMP software (JMP 15, SAS Institute Inc., Cary, NC). Larval development time, pupal development time, and adult life span were assessed using full factorial analysis of variance (ANOVA). Spore counts were assessed using one-way ANOVA or Student t test. Group differences were considered significant at p ≤ 0.05. RAIN (Thaben and Westermark, 2014), a software package for R, which uses nonparametric methods, was used to analyze qPCR data and determine rhythmicity of the messenger RNA (mRNA) expression.
Results
The Larval Clock Is Rhythmic in Constant Light
Constant light has been shown to negatively impact host clock rhythms by causing dampened or arrhythmic clock gene expression and behavioral changes in rodents (Ohta et al., 2006; Mizutani et al., 2017; Jing et al., 2020). If constant light disrupts the monarch’s clock, we should observe effects on clock gene expression. We, thus, used quantitative real-time polymerase chain reaction (qPCR) to examine the temporal expression of 2 key clock genes cryotochrome2 (cry2) and period (per) in third instar (larvae) heads at 4-h intervals in a 14-h:10-h LD cycle, during DD and during LL.
Contrary to the expectations, monarch larvae appear to maintain rhythmic gene expression in all conditions. Larval cry2 and per RNA expression levels exhibited a daily rhythm in light-dark with peak levels at zeitgeber time (German for “time giver”; ZT) 20 and low levels at ZT 8 (p < 0.0001; Figure 1). Cry2 (p = 0.0003) and per (p < 0.0001) rhythms persisted in constant darkness; however, the peaks occurred at circadian time (CT) 12 and 20 with the lowest expression at CT 8 (Figure 1). We found that the expression of cry2 and per (p < 0.0001) in constant light were rhythmic but slightly altered with each other. Cry2 RNA expression levels peaked at CT 16 and per levels peaked at CT 20. The finding that constant light and darkness conditions maintained rhythmicity of clock genes suggests that circadian clock regulation in the larval stage is less prone to disruption than in the adult based on previous studies.
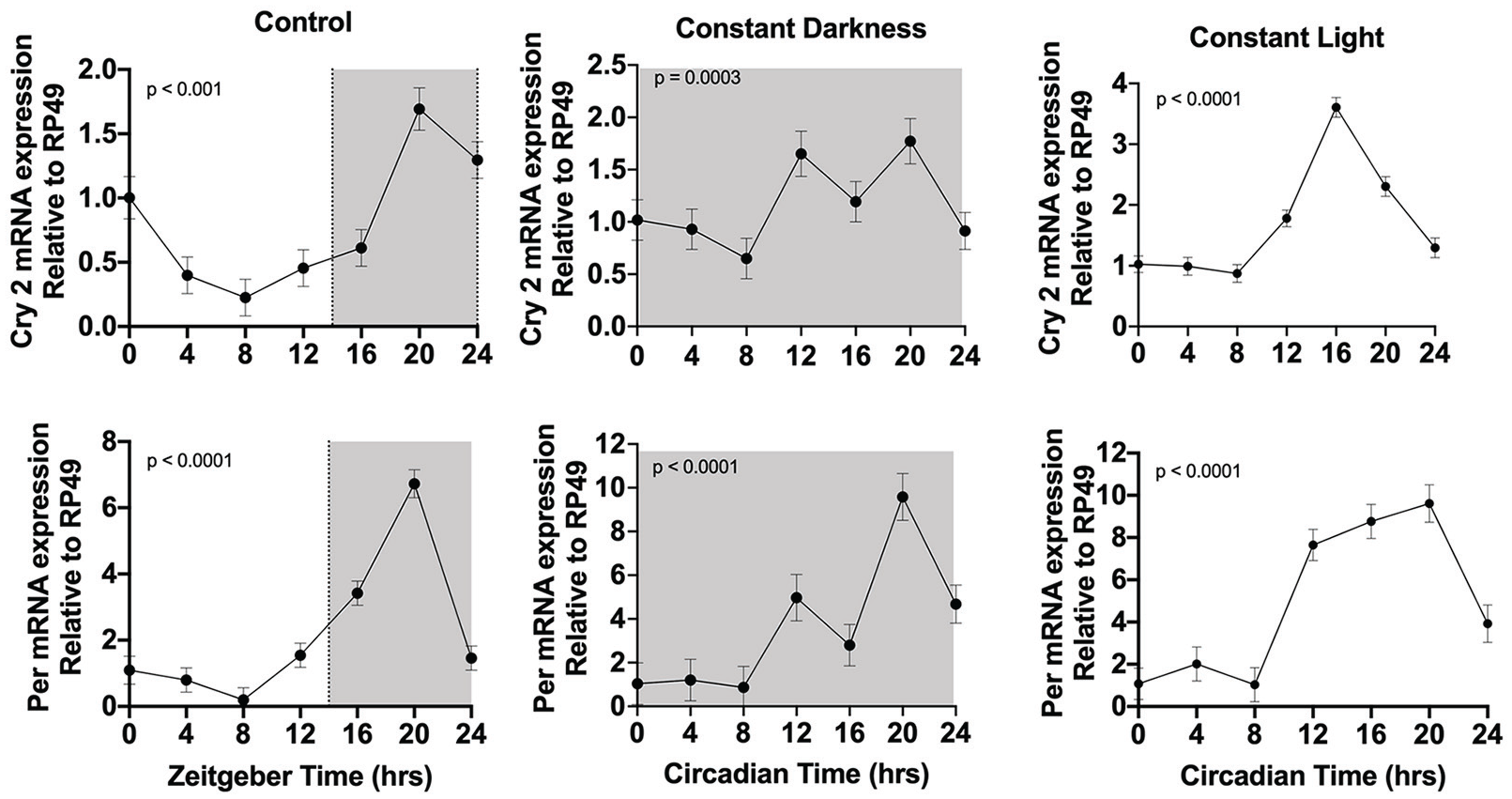
Clock gene expression in monarch larvae in control (light-dark: LD) and constant conditions (dark-dark: DD; light-light: LL). Clock gene RNA expression was measured in larval heads. Heads were collected at 4-h intervals for 24 h in LD, DD, and LL. RNA levels were quantitated by qPCR. White is lights-on and shaded is lights-off. The p-value was determined by RAIN, with p < 0.05 indicating rhythmic expression. Each value is mean ± SEM. Abbreviations: qPCR = quantitative polymerase chain reaction; SEM = standard error of the mean.
Larval Development Is Influenced By Constant Conditions
Since we observed that constant light does not cause desynchronization as anticipated, but larvae maintained their rhythms, we investigated whether constant light would have any physiological effects on monarch development, survival, and parasite susceptibility. Therefore, we placed monarch eggs in LD, LL, or DD and allowed them to develop under these conditions. We compared the rate of development of infected and uninfected monarchs from larva to pupation (larval development time) and from pupa to adult eclosion (pupal development time). We observed that rearing monarchs in constant conditions resulted in longer larval development times of 1.7 days (95% confidence interval [CI]: 1.4-2) for uninfected larvae and 1.2 days (95% CI: 1-1.4) for infected larvae (Figure 2a). Although there was a clear effect on larval development time, all larvae survived to adulthood (Figure 2b). In addition, we observed that pupal development time was longer by 2.7 days (95% CI: 2.5-2.9) in LL for uninfected pupae and by 2.2 days (95% CI: 2-2.4) for infected pupae (Figure 2c). This suggests that the lighting conditions have a greater effect on the rate of development than O. elektroscirrha infection. Indeed, infection with O. elektroscirrha had a minimal role in monarch development, a finding consistent with previous studies (de Roode et al., 2007).
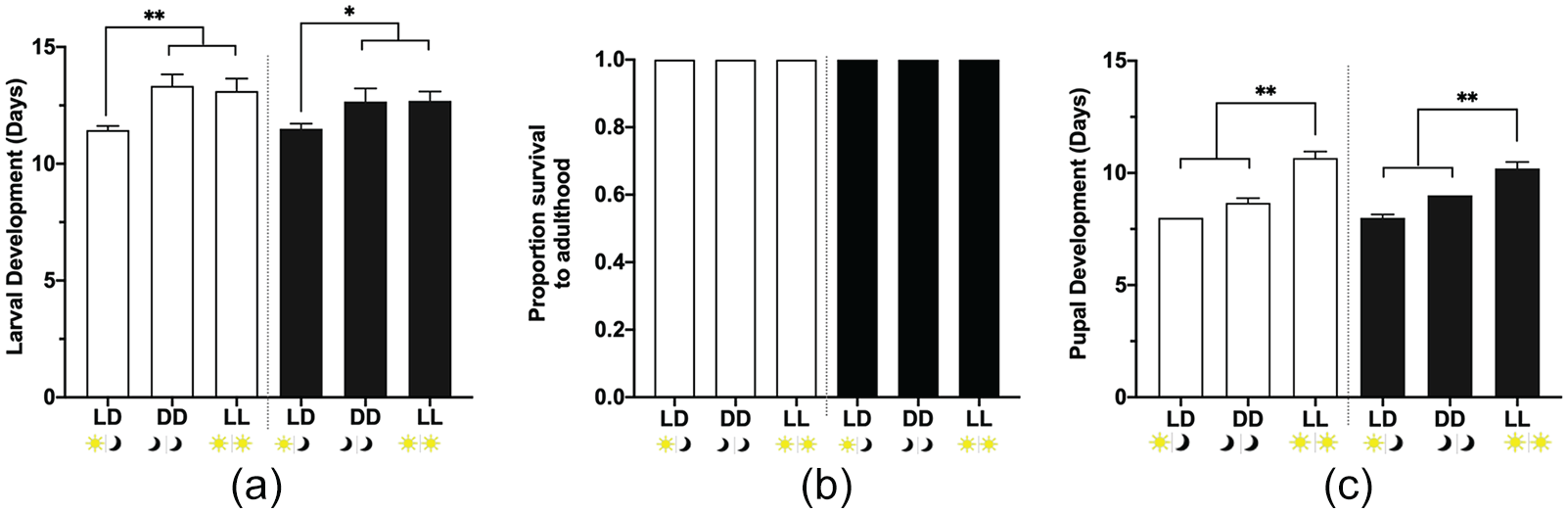
Larval development in control (light-dark: LD) and constant conditions (dark-dark: DD; light-light: LL). (a) Larval development time (larva to pupa). Full factorial ANOVA: parasite treatment p = 0.3128, light condition p = 0.0003, and parasite treatment × light condition p = 0.6730. (b) Survival to adulthood. (c) Pupal development time (pupa to adult). Full factorial ANOVA: parasite treatment p = 0.8059, light condition p < 0.0001, and parasite treatment × light condition p = 0.2093. Open bars are uninfected larvae: LD n =9, DD n = 6, and LL n = 9; dark bars are infected larvae: LD n = 10, LL n = 10, and DD n = 6. Values are mean + SEM. *p < 0.05, **p < 0.01. Abbreviations: ANOVA = analysis of variance; SEM = standard error of the mean.
Parasite Growth Is Affected, Not Adult Life Span
After we observed that there was a clear effect of light conditions on larval and pupal development, we investigated adult life span (under starvation conditions; see materials and methods) of infected and uninfected monarchs, as well as parasite growth (spore load) in infected monarchs. We observed that uninfected adult monarchs in constant light had a shorter life span of 1.8 days (95% CI: 1.5-2.1) compared with light-dark controls (Figure 3a). This suggest that constant light impacts the longevity of adult monarchs when they are uninfected. However, infected adults showed no differences in life span (Figure 3a). Furthermore, we observed no difference in parasite growth between light-dark controls and constant light (Figure 3b). Instead, our results showed an increase of 53% (95% CI: 52%-54%) in parasite growth when larvae are reared in constant dark conditions, not in constant light conditions (Figure 3b). Although there was an increase in parasite spores in constant darkness, it did not result in a significant decrease in adult life span compared with light-dark controls.
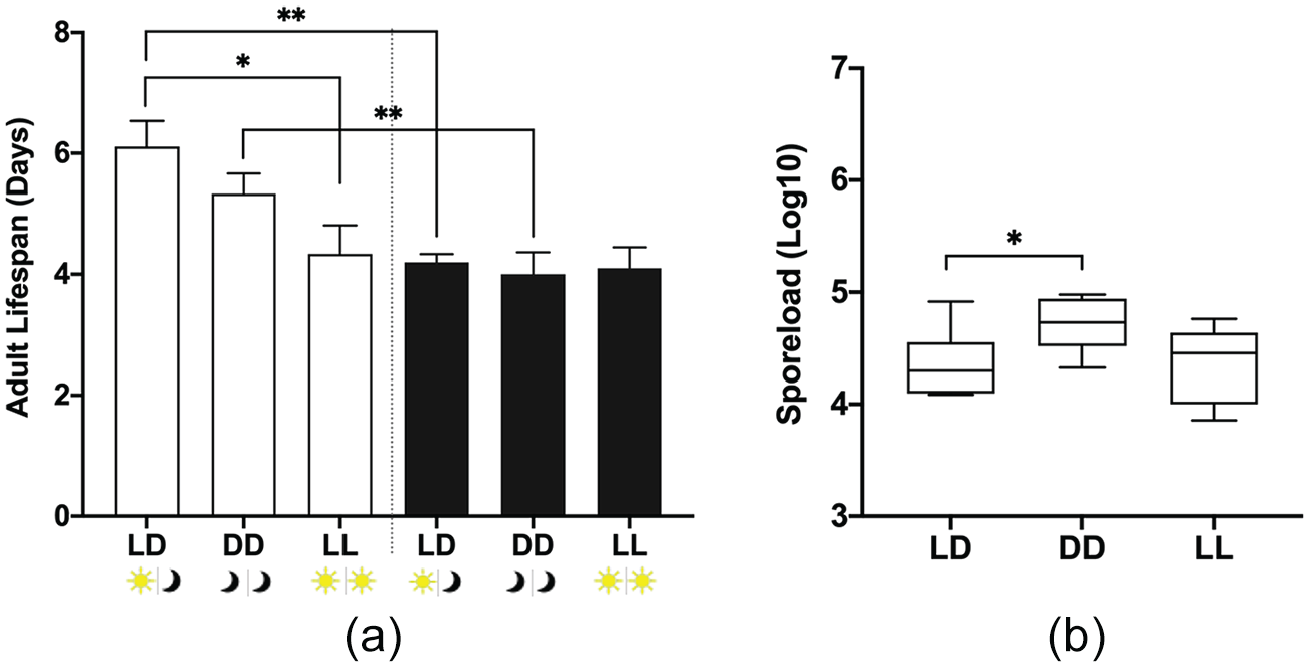
The life span of adult monarchs and parasite growth in control (light-dark: LD) and constant conditions (dark-dark: DD; light-light: LL). (a) Life span of adult monarchs after emerging from their chrysalis. Open bars are uninfected adults: LD n = 9, DD n = 6, and LL n = 9; dark bars are infected adults: LD n = 10, LL n = 10, and DD n = 6. Each value is mean + SEM. Full factorial ANOVA: parasite treatment p = 0.0004, light condition p = 0.0308, and parasite treatment × light condition p = 0.0560. (b) Parasite growth was measured by counting the number of spores on the abdomens of infected monarchs after death: p = 0.0311. *p < 0.05, **p < 0.01. Abbreviations: ANOVA = analysis of variance; SEM = standard error of the mean.
Schedule Changes Cause the Monarch Larval Clock to Be Out of Synchronization With the Light-Dark Cycle
As with subjecting animals to constant light, exposing animals to multiple schedule changes can lead to circadian disruption and reduce an animal’s ability to survive (Castanon-Cervantes et al., 2010). If multiple schedule changes disrupt the monarch’s clock, we should observe effects on core clock gene expression. Therefore, we collected larval heads at 4-h intervals for 24 h. We observed that before the schedule change, core clock genes cry2 (p < 0.0001), per (p < 0.0001), and timeless (tim; p = 0.0006) all peak at ZT 20 (Figure 4) as observed in the above constant conditions experiment. We then exposed larvae to a single 6-h phase advance (i.e., lights came on 6 h earlier) and observed that 1 day after the schedule change the core genes were not synchronized (Figure 4) with the new lighting schedule, with cry2 and tim peaking at ZT 4 and 20, and with per peaking at ZT 20. We observed that on day 2, clock gene expression remained unsynchronized (Figure 4) with the lighting environment. This suggests that the larvae were unable to adjust to the new schedule after 2 days and that the larval clock is difficult to shift.
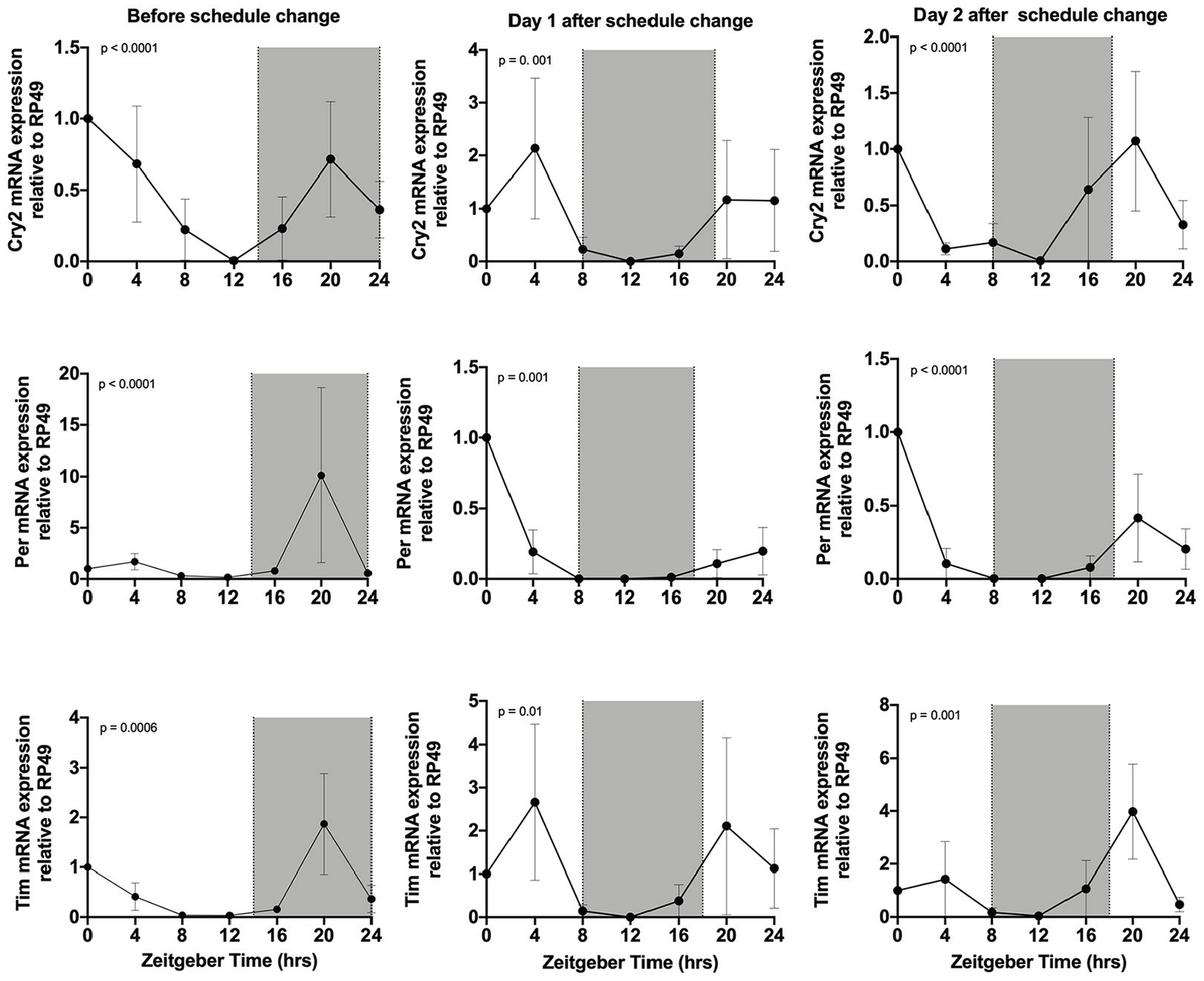
Clock gene expression after a single schedule change. Clock gene RNA expression in larval heads. Samples were collected at 4-h intervals for 24 h during a regular schedule of lights-on at ZT 0 (0700 h) and lights-off at ZT 14 (2100 h) (before schedule change) and after a 6-h schedule change lights-on at ZT 18 (0100 h) and lights-off at ZT 8 (1500 h); day 1 and day 2 after schedule change are the same schedule; no schedule change occurred in between. RNA levels were quantified by qPCR. White is lights-on and shaded is lights-off. The p-value was determined by RAIN, with p < 0.05 indicating rhythmic expression. Each value is mean ± SEM. Abbreviations: ZT = zeitgeber time; qPCR = quantitative polymerase chain reaction; SEM = standard error of the mean.
Schedule Changes and Larval Development
In many instances, circadian disruption caused by frequent schedule changes can lead to increased disease progression (Filipski et al., 2003, 2009; Scheer et al., 2009). As reported above in the constant conditions experiment, we observed longer development times and decreased adult life span in uninfected monarchs in constant light. We decided to further investigate how the common use of schedule changes would impact development, survival, and O. elektroscirrha infection. Therefore, we quantified larval and pupal development time in uninfected and infected monarchs. We observed that infected monarchs reared under a schedule change took 0.6 days (95% CI: 0.4-0.8) less to develop than infected controls (Figure 5a). We compared larval survival for controls and schedule change monarchs and observed similar survival rates. Schedule changes did not decrease monarch larval and pupal survival to adulthood (Figure 5b). Moreover, we observed no differences in pupal development time for controls and schedule change in uninfected and infected monarchs (Figure 5c). The impact of schedule changes on larval and pupal development time was small compared with the effects observed in constant light conditions.
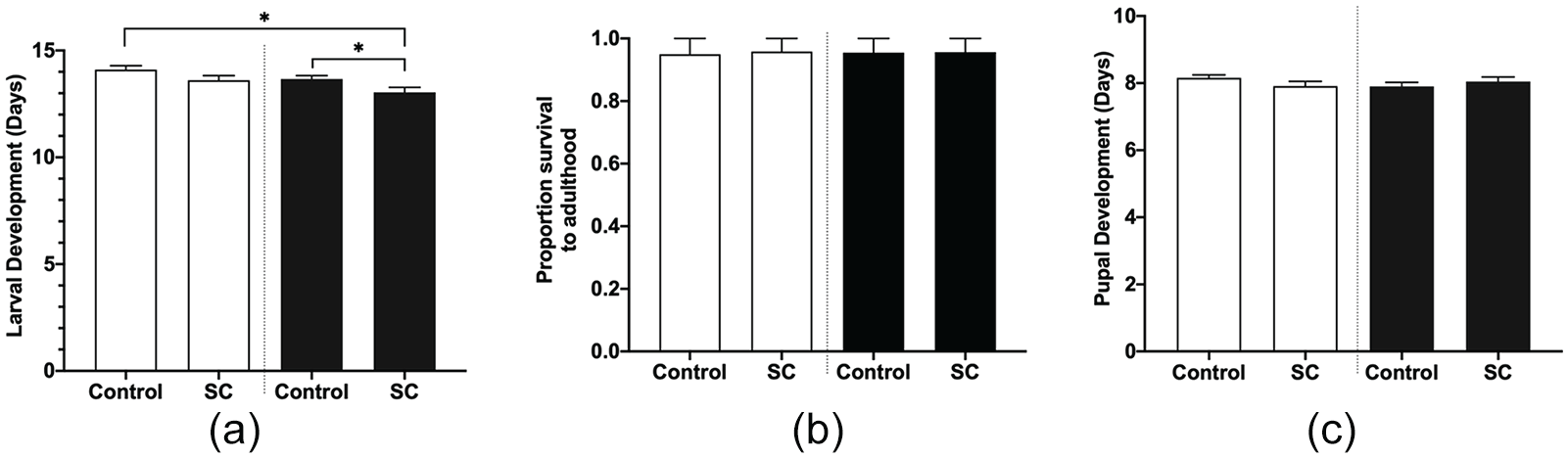
Effects of control and schedule change (SC) light conditions on larval and pupal development time. (a) Larval development time. Full factorial ANOVA: parasite treatment p = 0.0159, light condition p = 0.0074, and parasite treatment × light condition p = 0.7601. (b) Survival to adulthood. (c) Pupal development time. Full factorial ANOVA: parasite treatment p = 0.6327, light condition p = 0.6801, and parasite treatment × light condition p = 0.1294. Open bars are uninfected monarchs: controls n = 19 and SC n = 23; dark bars are infected monarchs: controls n = 21 and SC n = 22. Values are mean + SEM. Abbreviations: ANOVA = analysis of variance; SEM = standard error of the mean.
Parasite Growth Is Affected by Frequent Schedule Changes in the Host
We further investigated the effect of schedule changes on adult life span and parasite growth. We found a clear effect of parasite treatment (p < 0.0001) but not of light condition (p = 0.3849; Figure 6a), with infected monarchs living 1.2 days (95% CI: 1-1.4) shorter than uninfected monarchs under both light conditions. Again, this suggests that frequent schedule changes do not have a negative impact on the host’s ability to survive the parasitic infection. Interestingly, however, monarchs in the schedule change group experienced a decrease in parasite growth by 43% (95% CI: 42-44; p = 0.0101; Figure 6b), suggesting that although there was no severe effect on the host, there was some effect of the circadian disruption on the parasite. This suggest that the parasite may have its own biological clock, or it may be responding to a specific protein in the monarch that may have been altered but does not impact development.
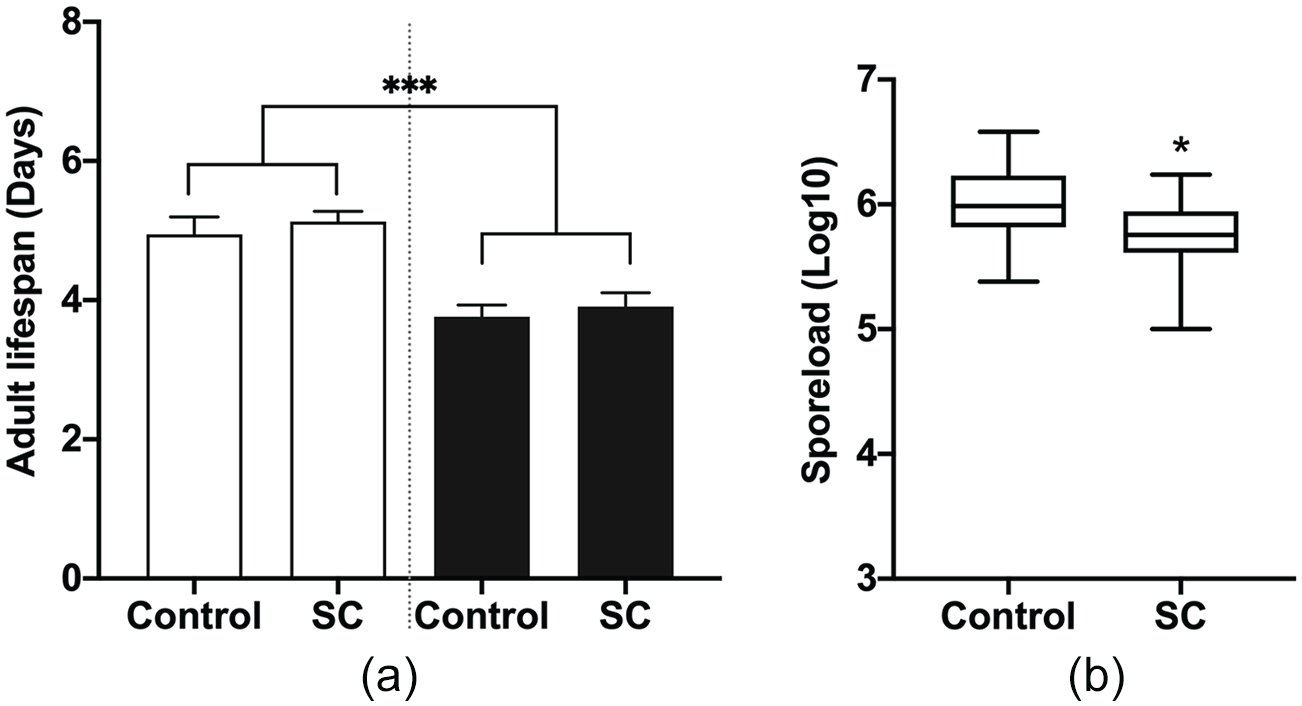
The effect of multiple schedule changes (SC) on adult life span and parasite growth. (a) The number of days monarchs survived as an adult under normal light conditions (control) or after multiple SC. Open bars are uninfected monarchs: controls n = 19 and SC n = 23; dark bars are infected monarchs: controls n = 21 and SC n = 22. Each value is mean + SEM. Full factorial ANOVA: parasite treatment p < 0.0001, light condition p = 0. 3849, and parasite treatment × light condition p = 0.9246. (b) Parasite growth, as measured by the number of parasite spores on infected adult abdomens after death: p = 0.0101. ***p < 0.001, *p < 0.05. Abbreviations: ANOVA = analysis of variance; SEM = standard error of the mean
Discussion
Our work shows that the study of outbred, diurnal animals with a natural host-parasite system can yield results contrary to lab-adapted inbred models. We show here that placing monarchs in constant light or dark conditions for their entire life does not result in arrhythmicity or dampened rhythms of core clock genes. Instead, larvae remained rhythmic in per, cry2, and tim gene expression, with a slight difference occurring in constant light with per peaking at CT 12 and 20 and cry2 peaking at CT 16 compared with controls where both per and cry2 peaked at ZT 20. However, external desynchronization was observed in frequent schedule changes, with per, cry2, and tim peaking during the light phase instead of the dark phase after a schedule change. Yet the adverse health consequences that are often associated with frequent schedule changes were not observed in our study. Although monarchs took 2-3 days longer to develop in constant light conditions, it did not increase susceptibility to the parasite infection. These findings oppose the status quo that constant conditions increase host susceptibility to infection. Interestingly, different types of environmental circadian disruption of the host had opposing effects on O. elektroscirrha growth. Constant light, commonly used to disrupt the clock in a variety of model systems, had no effect on the parasite, but constant darkness, which is not usually qualified as disruptive, increased parasite growth. In contrast, frequent schedule changes actually decreased parasite growth.
Circadian disruption has been shown to have adverse health effects in mammals and Drosophila. Exposing mice to constant light during early development disrupts the developing biological clock and desynchronizes the neurons in the master clock in the brain (Ohta et al., 2006). Other studies revealed that constant light decreased the development of mouse immune tolerance (Mizutani et al., 2017) and that circadian disruption decreased survival (Castanon-Cervantes et al., 2010). In Drosophila, similar results were observed: when flies were exposed to chronic circadian disruption, they had a decrease in longevity (Boomgarden et al., 2019). Similarly, our constant light conditions resulted in decreased survival in uninfected monarchs and longer development times. This could be due to an overexposure to light affecting survival and development in uninfected monarchs, rather than disruption since we observed a small effect on clock gene expression in constant light. Yet, interestingly, the larvae maintained their rhythms in constant light. The robustness of their rhythms suggests that the monarch clock during early development is harder to phase shift and therefore may help larvae to better handle disruptive conditions overall. In addition, monarch larvae were fed live plants. It is possible that the plant’s clock or nutrients played a role in regulating the monarch larval clock, thus resulting in maintained rhythms in constant light. However, further research on plant and insect larval clock interactions is needed. Moreover, there is still much to study and understand about the monarch’s clock and its role in other biological processes and whether O. elektroscirrha has its own clock. Reppert (2007) highlights the monarch clock as an ancestral clock which is a hybrid between the mammalian and Drosophila clocks. The monarch biological clock possesses duplicate cryptochrome genes with 2 different functions. It contains a Drosophila-like cry1 which functions primarily as a blue-light photoreceptor and a mammalian-like cry2 that functions as a potent transcriptional repressor (Song et al., 2007; Zhu et al., 2008). Previous studies in monarchs have shown that genetic manipulation of the core clock genes such as clk and cry leads to time-of-day effects on eclosion, with adults emerging from their pupae throughout the day instead of during the early morning (Markert et al., 2016; Zhang et al., 2017). However, these studies revealed no evidence to suggest that a disrupted clock in monarchs leads to negative effects on health.
Constant conditions and frequent schedule changes had varying effects on the host-parasite interaction. Constant light increased monarch larval and pupal development time without impacting parasite growth. Yet under constant darkness we observed increased parasite growth. Constant darkness improved the parasite’s ability to replicate without significantly impacting the adult life span. On the contrary, frequent schedule changes decreased parasite growth, but did not improve adult life span. This suggests a potential decoupling of parasite growth and virulence, indicating that constant light and frequent schedule changes affect monarch tolerance of infection (Lefevre et al., 2011). Even though there were no great differences in the rate of development after multiple schedule changes, it is possible that at the molecular level changes occurred, which led to the decrease in parasite growth. O’Donnell et al. (2013) used opposing light-dark schedules to cause a mismatch between host and parasite. They transferred Plasmodium parasites from mice in a standard light-dark condition (i.e., lights-on at 0000 h and lights-off at 1200 h) to mice with a reversed light-dark schedule. This revealed that Plasmodium prefers to be synchronized with its host and that when it cannot achieve that, it significantly impacts its ability to infect the host (O’Donnell et al., 2013).
We focused on the larval stage because larvae are the only monarch life stages that can be infected by O. elektroscirrha, and thus serve as the starting point for the parasite life cycle. A study conducted by Niepoth et al. (2018) similarly investigated circadian gene expression—as well as feeding behavior—in monarch larvae. Interestingly, however, that study also investigated adults and found that clock gene expression differs between larvae and the adults. The adults expressed per and tim in equal quantities, and the larvae expressed tim more highly than per (Niepoth et al., 2018). That work and ours suggest that further investigation of the differences between larval clock development and the adult stage is needed. Similarly, our study leads to the need for further understanding of the parasite. It is not yet clear whether the parasite has its own rhythms or whether it is controlled by the host’s rhythms. Research conducted in Trypanosoma brucei (the causative agent of African sleeping sickness) (Rijo-Ferreira et al., 2017) and Plasmodium (the causative agent of malaria) (O’Donnell et al., 2011; Smith et al., 2020) revealed that both these parasites have intrinsic clocks. Thus, given the phylogenetic relatedness between Plasmodium and O. elektroscirrha—both are apicomplexan parasites (Carreno et al., 1999)—it is possible that O. elektroscirrha has its own biological clock as well. Ongoing genomic studies will enable us to determine whether this is the case, opening up new avenues to investigate the relative importance of host and parasite genes in mediating circadian disruption on the host-parasite interaction.
Footnotes
Acknowledgements
We thank Erik Edwards for maintaining plants, Joselyne Chavez for help maintaining monarchs, and the de Roode Lab for early morning and late night sample collections.
This research was funded by the National Science Foundation grant IOS-1557724 to J.C.d.R. and IRACDA Fellowships in Research and Science Teaching (NIH K12 GM 000680).
Conflict Of Interest Statement
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.