
Abstract
The chicken pineal gland synthesizes and releases melatonin rhythmically in light/dark (LD) cycles, with high melatonin levels during the dark phase, and in constant darkness (DD) for several cycles before it gradually damps to arrhythmicity in DD. Daily administration of norepinephrine (NE) in vivo and in vitro prevents the damping and restores the melatonin rhythm. To investigate the role of the circadian clock on melatonin rhythm damping and of its restoration by NE, the effects of NE administration at different phases of the melatonin cycle revealed a robust rhythm in NE sensitivity in which NE efficacy in increasing melatonin amplitude peaked in late subjective night and early subjective day, suggesting a clock underlying NE sensitivity. However, NE itself had no effect on circadian phase or period of the melatonin rhythms. Transcriptional analyses indicated that even though the rhythm of melatonin output damped to arrhythmicity, messenger RNA (mRNA) encoding clock genes gper2, gper3, gBmal1, gclock, gcry1, and gcry2; enzymes associated with melatonin biosynthesis; and enzymes involved in cyclic nucleotide signaling remained robustly rhythmic. Of these, only gADCY1 (adenylate cyclase 1) and gPDE4D (cAMP-specific 3′,5′-cyclic phosphodiesterase 4D) were affected by NE administration at the mRNA levels, and only ADCY1 was affected at the protein level. The data strongly suggest that damping of the melatonin rhythm in the chick pineal gland occurs at the posttranscriptional level and that a major role of the clock is to regulate pinealocytes’ sensitivity to neuronal input from the brain.
The avian pineal gland is an ideal model for the study of circadian rhythmicity in that it displays the properties of a complete circadian system: 1) it is light sensitive, capable of entraining to environmental light cycles; 2) it expresses an endogenous oscillation; and 3) it rhythmically releases the hormone melatonin as its output in vivo and in vitro (Klein et al., 1997; Natesan et al., 2002). The gland expresses multiple opsin photopigments (Okano and Fukada, 1997), including pinopsin (Okano et al., 1994; Max et al., 1995), melanopsin (Bailey and Cassone, 2005; Chaurasia et al., 2005), and others, depending on the species (Okano and Fukada, 1997). In addition, the flavin-based cryptochrome of birds may itself be photoreceptive (Bailey et al., 2002; Mouritsen and Hore, 2012; Watari et al., 2012). Three enzymes involved in the melatonin biosynthesis pathway are regulated in a circadian pattern at the transcriptional, translational, and posttranslational levels, such that tryptophan hydroxylase (TrH) (Florez et al., 1996; Chong et al., 1998) and arylalkylamine N-acetyl-transferase (AANAT) (Bernard et al., 1997b) are expressed during the night in LD and subjective night in constant darkness (DD), while hydroxyindole-O-methyltransferase (HIOMT) is expressed during the day and subjective day, respectively (Bernard et al., 1993).
The molecular mechanism underlying circadian rhythms is highly conserved among animal phyla, consisting of a transcriptional feedback loop of positive and negative elements (Bell-Pedersen et al., 2005; Buhr and Takahashi, 2013). In vertebrates, the positive elements include the PAS-domain helix-loop-helix transcription factors Bmal1 and clock. Their proteins, BMAL1 and CLOCK, form a heterodimer in the cytoplasm that reenters the nucleus and binds to an E-box sequence (-CACGTG-) in the promotor regions of many genes and activates the expression of the negative elements as well other clock-controlled genes (CCGs). The negative elements, per and cry, form a feedback loop in which the PER and CRY proteins form heterodimers in the cytoplasm and reenter the nucleus to interfere with the activation of the BMAL1/CLOCK dimer. Multiple additional feedback loops, including one with Rev-Erb and RORs, inhibit or activate the expression of Bmal1, respectively. The transcriptional feedback loop of clock genes leads to a rhythmic expression of both clock genes and downstream CCGs, such as Aanat, TrH, and HIOMT, along with many other CCGs. The chicken pineal gland rhythmically expresses most of these clock and melatonin biosynthesis genes in vivo and in vitro (Bailey et al., 2003; Karaganis et al., 2008).
Although it is clear the chick pineal gland contains a photoreceptive circadian oscillator, in the absence of either photic or neural input, the melatonin rhythm damps after 2 to 6 cycles in DD (Takahashi et al., 1980; Robertson and Takahashi, 1988a, 1988b). The reasons for this damping are not clear. Damping may derive from the oscillatory mechanism in each cell declining in amplitude and/or may derive from multiple oscillators within the gland drifting out of phase with one another. At least 2 signals (light and norepinephrine) can reverse the damping of pineal melatonin rhythm. Light has 3 effects on the melatonin rhythm of cultured avian pineal glands: 1) it inhibits melatonin synthesis directly, 2) it increases the amplitude of the rhythm and prevents damping, and 3) it phase shifts the clock mechanism itself within the pineal cells (Zatz et al., 1988). In addition, the sympathetic neurotransmitter norepinephrine (NE), which derives from sympathetic innervation in vivo, inhibits melatonin synthesis and prevents the damping but has no phase shift effect (Cassone and Menaker, 1983; Cassone et al., 1986; Zatz and Mullen, 1988).
The molecular mechanisms by which NE prevents damping are also unknown. NE acts through α2-adrenergic receptor (Deguchi, 1979; Pratt and Takahashi, 1987) and its coupled Gi GTP binding protein, which inhibits adenylate cyclase (ADCY) and leads to lower cAMP levels (Pratt and Takahashi, 1988). When cAMP is low, AANAT, the critical enzyme of melatonin synthesis, is degraded such that melatonin production decreases, indicating a critical role for the cAMP signaling pathway in the regulation of melatonin synthesis (Klein et al., 1997). The present study was undertaken to examine the temporal expression patterns of melatonin release, clock genes, and cAMP signaling pathway-related genes in a microfluidic pineal cell culture model and explored the response to NE administration. The mechanism by which NE increases the amplitude and prevents damping is also addressed.
Materials and Methods
Cell Culture
Pineal glands were collected from embryonic day 21 chick (Gallus gallus domesticus) brain as previously described (Karaganis et al., 2008). Embryonic day 1 eggs were incubated in a GQF 1502 Sportsman incubator (Savannah, GA) at 38 °C, 75% humidity for 20 days. On day 21, 40 embryos were extracted from the eggs and decapitated. The pineal glands were transferred to sterile Dulbecco’s phosphate-buffered saline (PBS) with D-glucose (1.8%) (Sigma-Aldrich, St. Louis, MO) and digested by 0.25% trypsin (10 pineals/mL of trypsin) for 30 min with trituration every 10 min. The debris was removed, and then the dispersed cells were spun down for 10 min at 1000 × g. The pellet was resuspended to 4 pineals/mL (106 cells/mL) in M-199 medium with 10% fetal bovine serum (FBS) (Sigma-Aldrich), 10% chicken serum, and 1% penicillin/streptomycin (Invitrogen, Life Technologies, Grand Island, NY). Then, 30 µL of the cells was loaded into one channel of the 6-channel multiwell microfluidic slide (IBIDI USA, Madison, WI; cat. 80606), or 1 mL of the cells was plated on 6-well plates.
The cells were cultured in a Forma Series II Water Jacket CO2 incubator (Model 3100, Thermo; Life Technologies, Grand Island, NY) and maintained at 36.5 °C with 5% CO2 in constant darkness until 1000 h the next day. They were initially cultured in 12:12 LD cycles (34 µW/cm2 from LED lights) using 1000 h as zeitgeber time 0 (ZT0). When the cells grew to ~80% confluence (after 2-3 light/dark cycles), 2 microfluidic slides with a total of 12 channels of cultured pineal cells were loaded on the superfusion system. The superfusion system consisted of a modified syringe pump (Harvard Apparatus, Holliston, MA), Tygon tubing, the slides, and a customized fraction collector. The pump drove culture medium through the 12 syringes to flow through the microfluidic slides continuously with a flow rate of 100 µL/h/chamber (thus, there was no pulse resulting from medium exchange) where the pineal cells were cultured. Medium containing melatonin secreted from the pineal cells was collected every hour.
Experimental Protocol
Five experiments were conducted. In experiment 1, cells were cultured in LD for 3 days and then placed in DD for 7 days. On day 3 of DD, at CT0, 3 control channels on each slide (n = 6 total) received a single bolus of 100 µL 10 mg/mL ascorbate (Sigma-Aldrich A4034), while the remaining 6 channels received 100 µL 10−6 M norepinephrine bitartrate (Sigma-Aldrich A0937) in 10 mg/mL ascorbate. Samples were collected every hour, and melatonin levels were measured.
In experiment 2, cells were treated as in experiment 1 with the exception that beginning on day 3 of DD, cells received a series of 3 boli of 100 µL NE or of ascorbate at CT0 on 3 successive days. Cells were then allowed to survive another 3 days. As above, samples were collected every hour, and melatonin levels were measured.
To ask whether there was a phase-dependent effect of NE on pineal cell melatonin release, in experiment 3, 4 sets of slides were exposed to either NE or ascorbate at 4 different phases in the melatonin cycle: at the beginning of subjective day (CT24/0), mid-subjective day (CT6), early subjective night (CT16), and late subjective night (CT20) 3 times 24 h apart. As above, cells were allowed to incubate for another 3 days while samples were collected. Melatonin was then measured in these samples.
In experiment 4, expression of 40 candidate genes involved in the melatonin rhythm in DD and in the response to NE administration were measured using the Nanostring nCounter system (Nanostring Technologies, Seattle, WA; see below), including clock genes and genes related to the cAMP signaling pathway (Supplemental Table 1; Table 1). These genes were selected from transcriptional profiling studies that identified most likely components of the chick pineal clock as well as sequences known to be involved in monoaminergic signaling (Bailey et al., 2003; Karaganis et al., 2008). NE was administered at CT24 of day 5 (the beginning of day 6 on the superfusion system) in DD, because NE administration created the highest melatonin amplitudes at this time point. The messenger RNA (mRNA) from 3 channels of the control group and 3 channels of the treatment group were collected in 4-h intervals from CT2 of day 8 (the last NE administration cycle) to the end of day 9, a total of 12 time points over the 48 h. The gene expression was measured by Nanostring. Cultured pineal cells on the microfluidic slides were homogenized in Trizol by pipetting for total RNA extraction.
Rhythmicity of genes in DD and the response of genes to norepinephrine (NE).
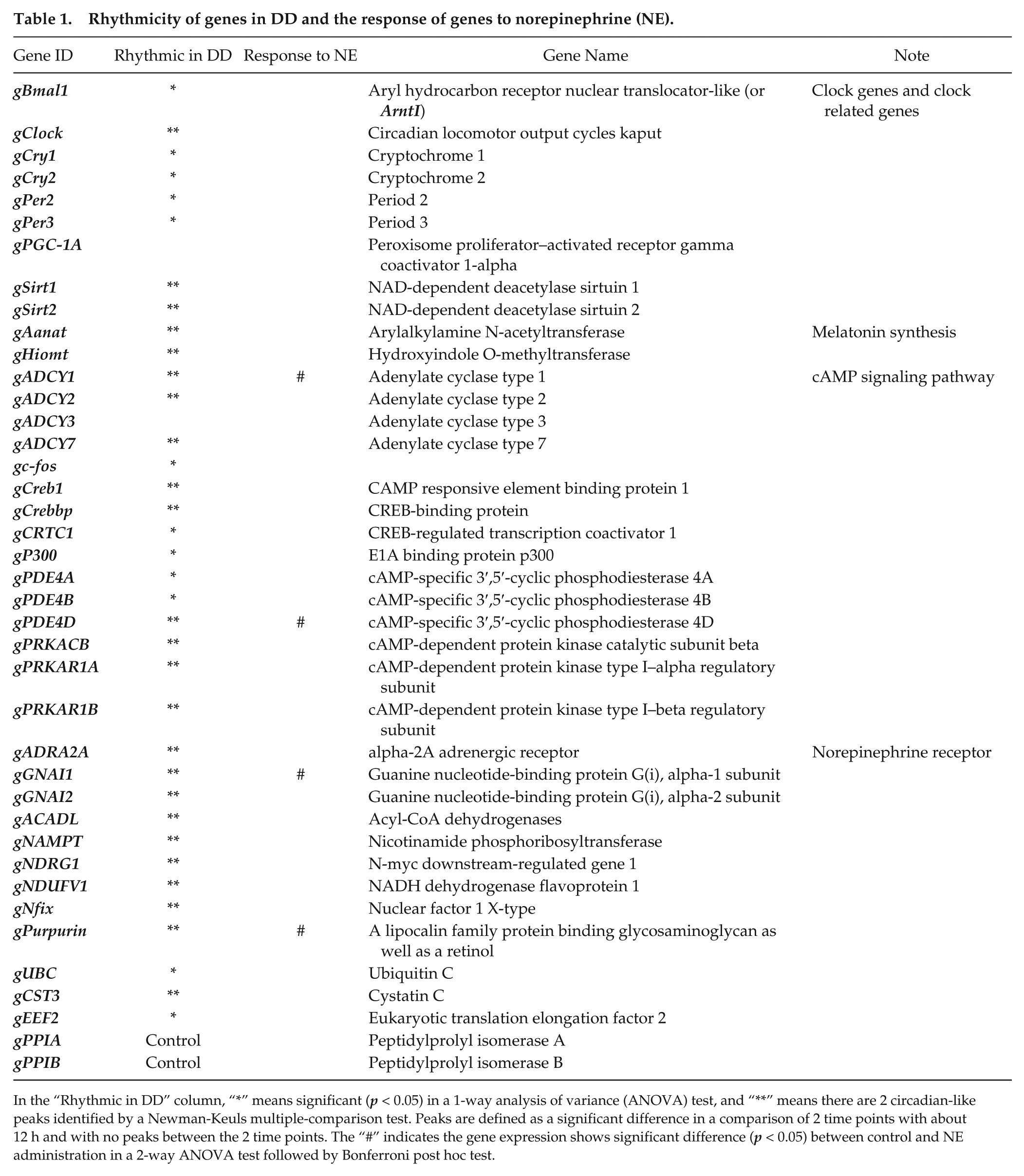
In the “Rhythmic in DD” column, “*” means significant (p < 0.05) in a 1-way analysis of variance (ANOVA) test, and “**” means there are 2 circadian-like peaks identified by a Newman-Keuls multiple-comparison test. Peaks are defined as a significant difference in a comparison of 2 time points with about 12 h and with no peaks between the 2 time points. The “#” indicates the gene expression shows significant difference (p < 0.05) between control and NE administration in a 2-way ANOVA test followed by Bonferroni post hoc test.
In experiment 5, expression of 2 key proteins from the cyclic nucleotide transduction pathway was measured by Western blot analysis. Cells were collected as above from 3 channels as ascorbate control and 3 channels during NE treatment. Protein was extracted as described below and prepared for Western blot analysis for phosphodiesterase 4D (PDE4D), adenylate cyclase I (ADCY1), or peptidyl prolyl isomerase A (PPIA) as a control.
Melatonin Enzyme-Linked Immunosorbent Assay
Melatonin content of culture medium was measured according to Li and Cassone (2015). Monoclonal melatonin antibody (CalBioreagents, San Mateo, CA) was diluted (0.5 µg/mL) in plate coating buffer (pH 7.4, PBS with 5% sucrose) and then loaded as 50-µL antibody solutions per well of the 96-well plates (Thermo Pierce 15041; Life Technologies). The plates were kept at 4 °C overnight (12-20 h). Then the antibody solution was removed and rinsed 4 times with 300 µL washing buffer (pH 7.4, PBS with 0.5% Tween 20) in each well. The plates were blocked with 50 µL Protein-Free (PBS) Blocking Buffer (Thermo Pierce 37516; Life Technologies) and held at 4 °C for 1 to 2 h. After removing of the blocking buffer, the plates were rinsed 2 times with washing buffer and put under vacuum for drying. A stock melatonin (Sigma-Aldrich M-5250) standard of 1 mg/mL was prepared by dissolving 10 mg melatonin in 100 µL dimethyl sulfoxide (DMSO) (Sigma-Aldrich D2650) first and then in 9.9 mL PBS. The stock solution was further diluted with PBS or cell culture medium (M-199, Sigma-Aldrich M7528) with 10% FBS to give 6 individual standards ranging from 50 to 10,000 pg/mL. The standards of the assay covered a wide working range from 100 pg/ml to 10 ng/ml. Then, 25 µL of melatonin calibrators or samples was loaded respectively into each well of the plate, followed by 25 µL Mel-HRP (1:2000 dilution in PBS; CalBioreagents). Then the plate was incubated at 4 °C for 3 h. The liquids were removed, and 50 µL tetramethylbenzidine (TMB) solution (Thermo Scientific 34028; Life Technologies) was added into each well for 15 min after the plates were washed for 4 times with washing buffer. The color reaction was ceased with 50 µL stop solution (50-85-05; KPL, Inc., Gaithersburg, MD). OD readings were recorded by enzyme-linked immunosorbent assay (ELISA) plate reader (iMark; Bio-Rad, Hercules, CA). The sensitivity of the assay was 68 pg/mL in cell culture medium and 26 pg/mL in PBS with as little as 25 µL sample volume. The recovery of melatonin from cell culture medium was 101.0%. The principal cross-reacting compounds were N-acetyl-5-hydroxytryptamine (0.03%) (Sigma-Aldrich A1824) and 5-methoxytryptophol (0.003%). The coefficients of variation (CV) of the assay, within and between runs, ranged between 6.68% and 15.76% in cell culture medium. The mean linearity of a series diluted cell culture medium sample was 105% (CV = 5%), ranging between 98% and 111%, y = 5.5263x + 0.0646, R² = 0.99. Details of this assay are published elsewhere (Li and Cassone, 2015).
RNA Quantification
Total RNA was extracted by the ZYMO Direct-zol RNA mini Prep kit with DNAase treatment (ZYMO Research, Seattle, WA; cat. R2052). The mRNA expression was measured by gene expression quantification using multiplexed, color-coded probe pairs (NanoString nCounter; Nanostring Technologies) and validated by semi-quantitative polymerase chain reaction (PCR). For NanoString, each of the 40 genes, including clock genes and genes related to cAMP signaling pathway, was targeted for designing and synthesis of probe sets (Supplemental Table S1). Probes sets of 100 bp in length were designed to hybridize specifically to each mRNA target and be identified by the unique color code of each probe (Reis et al., 2011). Samples were subjected to Nanostring nCounter analysis by the University of Kentucky Microarray Core Facility (http://www.research.uky.edu/microarray/). Briefly, for each sample, 10 µL Reporter CodeSet and 10 µL hybridization buffer were mixed with 5 µL RNA (150 ng). Then, 5 µL Capture ProbeSet was added to the mixture, and then the mixture was placed in a 65 °C thermocycler for a 12-h incubation. Once removed from the thermocycler, samples were processed with the nCounter Prep Station (NanoString nCounter; Nanostring Technologies). The data were normalized first by subtracting the average of the negative controls to eliminate background and calibrated by the standard curve from positive controls, and then the data were normalized to the internal control (gPPIA and gPPIB).
For semi-quantitative PCR (qPCR), 1 µg total RNA was digested by the RQ1 DNase I kit (Promega, Madison, WI; cat. M6101) and reverse transcribed to complementary DNA (cDNA) by the qScript cDNA Super Mix kit (Quanta Biosciences, Gaithersburg, MD; cat. 95084). The cDNA from cultured pineal cells and intact pineal glands were used as templates in PCR. Then, 2 µL cDNA (one-fifth of the transcribed total RNA) was mixed with 12.5 µL 2× PCR Master Mix (Promega; cat. M7502), 0.5 µL of each primer (10 µM), and 9.5 µL water to a total of 25 µL. PCR was carried on a Gene Amp PCR system 9700 (Applied Biosystems; Life Technologies). The qPCR was run as follows: 2 min of initial denature at 95 °C was followed by 30 cycles of 50 sec at 95 °C, 40 sec at 50 °C (could be different depending on the annealing temperature of the target gene), and 50 sec at 72°C; then 7 min at 72 °C; and then hold at 4 °C. The primer sequences and data analysis followed previous work in our lab (Karaganis et al., 2008).
Western Blot Analysis and Protein Assay
Total protein of 3 wells from cultured pineal cells was extracted at each point with RIPA buffer (Santa Cruz Biotechnology, Santa Cruz, CA; sc-24948), and the concentration was measured by the Bradford assay (Noble and Bailey, 2009). Western blots were run as previously described (Malloy et al., 2012). Briefly, 5 µg protein sample was run on the SDS-PAGE gel in triplicate (Mini-PROTEIN 3; Bio-Rad) and transferred on a nitrocellulose membrane (Protran BA83; Whatman, Pittsburgh, PA) by Criterion Blotter (Bio-Rad). The membrane was blocked by 5% milk in PBS with 0.5% Tween 20. The antibody of phosphodiesterase 4D (PDE4D), adenylate cyclase I (ADCY1), or peptidyl prolyl isomerase A (PPIA) was applied to the membrane at a 1:1000 dilution. Biotinylated secondary antibody (1:5000 dilution, anti–rabbit IgG) was applied followed with amplification of the avidin/biotinylated enzyme complex and color reaction (VECTASTAIN ABC-AP kit; Vector Laboratories, Burlingame, CA). The PPIA served as an internal control for normalizing the relative expression level of PDE4D and ADCY1. The results were quantified by ImageJ (National Institutes of Health, Bethesda, MD).
Statistics
Melatonin data were subjected to cosinor analysis using linear harmonic regression (CircWave software, Groningen, The Netherlands), as well as analysis of variance (ANOVA) (Karaganis et al., 2008). Nanostring time course data were analyzed by ANOVA followed by the Newman-Keuls multiple-comparison test and CircWave. Gene expression response to NE was analyzed by 2-way ANOVA followed by a Bonferroni multiple-comparison post hoc test. Autocorrelations were calculated by Wessa P software (Wessa, 2015). Melatonin amplitude changes were subjected to a Student t test. ANOVA and t tests were performed using GraphPad Prism (GraphPad Software, La Jolla, CA) and Sigma Stat software (Systat Software, Point Richmond, CA) packages. Western blot results were analyzed by a Student t test.
Results
Melatonin Rhythm of Cultured Chicken Pineal Cells in LD/DD and Effect of NE
Melatonin levels were rhythmic in vitro in both LD and in the first 2 cycles in DD before damping to arrhythmicity (Figures 1-4). Autocorrelation analysis of individual channels for the last 2 days in LD, the first 2 days in DD, and the second 2 days in DD indicated that the melatonin rhythm was robust in the LD cycle and in the first 2 days in DD and then damped out (Figure 1). The combined data sets from 6 channels of the control group and 6 channels with a single NE administration from the perfusion system under LD and DD were measured and profiled (Figure 2). Rhythmicity in LD and DD was analyzed by CircWave and ANOVA. Using these tests, the melatonin outputs in LD displayed a significant circadian rhythm (p < 0.05) and, in composite, in DD did not (p > 0.05), even though time-series analyses on individual channels showed significant rhythmicity in the first 2 days of DD (Figure 1). The single pulse of NE administration suppressed the melatonin production and created a rebound following the trough (Figure 2).
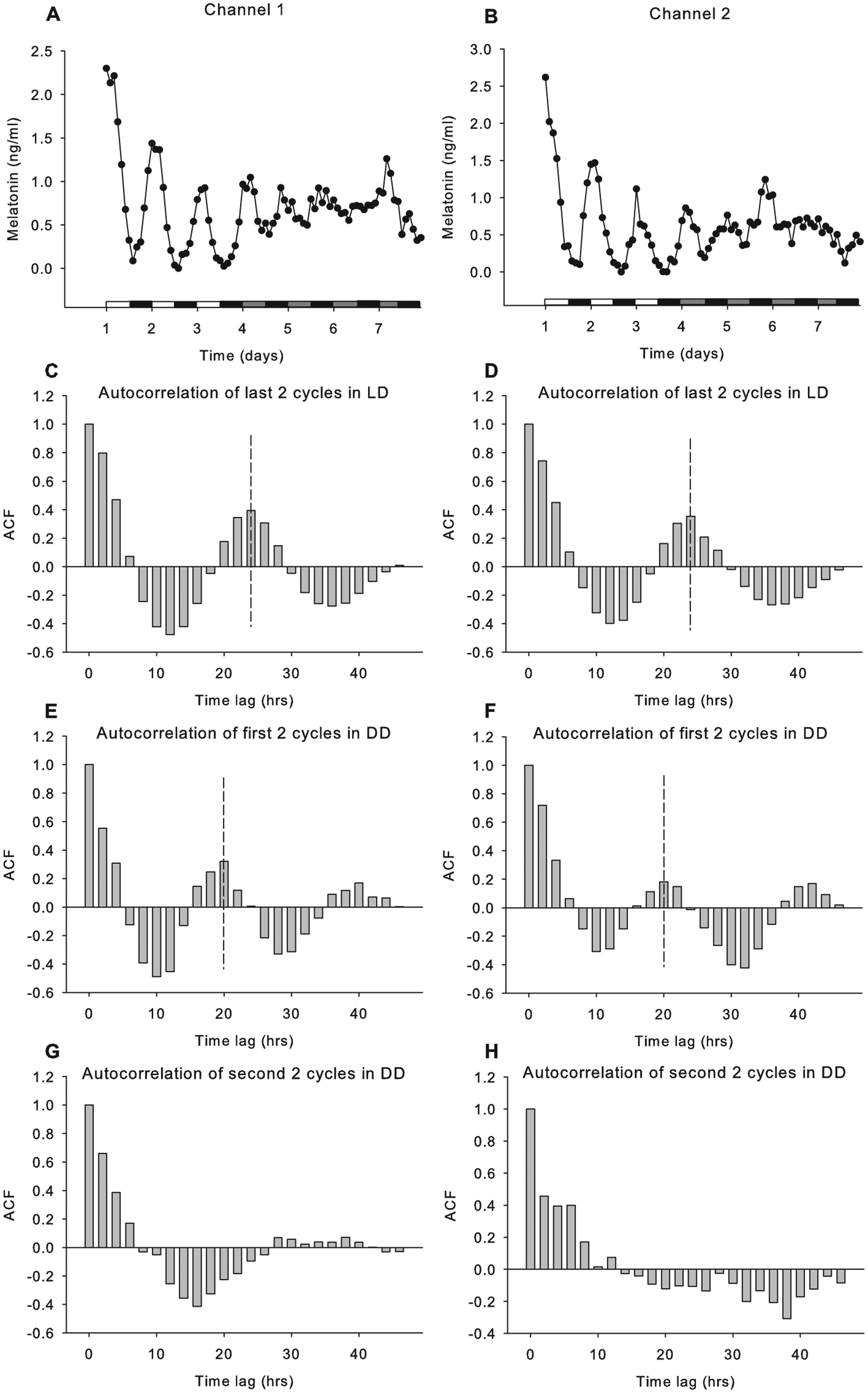
Melatonin rhythms from dispersed pineal cells in 2 individual channels. (A) Melatonin output from single channel 1 from dispersed pineal cells in 3 days LD and 4 days in constant darkness (DD). The x-axis indicates time in days. Day 1 is defined as the day when the perfusion system started (the same definition applies to all the x-axes in the following figures). The white bar on the x-axis indicates daytime with light, the dark bar indicates nighttime in darkness, and the gray bar indicates the subjective daytime in constant darkness. (B) Melatonin output from single channel 2 from dispersed pineal cells in 3 days LD and 4 days in constant darkness (DD). (C-H) The autocorrelation functions corresponding to different phases of the melatonin output rhythms. The time lag (dashed line, indicating the period) of the last 2 cycles in LD is 24 h for both channels, and autocorrelation functions (ACFs) are 0.39 for channel 1 (C) and 0.35 for channel 2 (D); the time lag of the first 2 cycles in DD is 20 h for channel 1 (E) and channel 2 (F) with ACFs of 0.25 and 0.18 for each. The time lag and ACF of the second 2 cycles in DD were not detected (G, H).
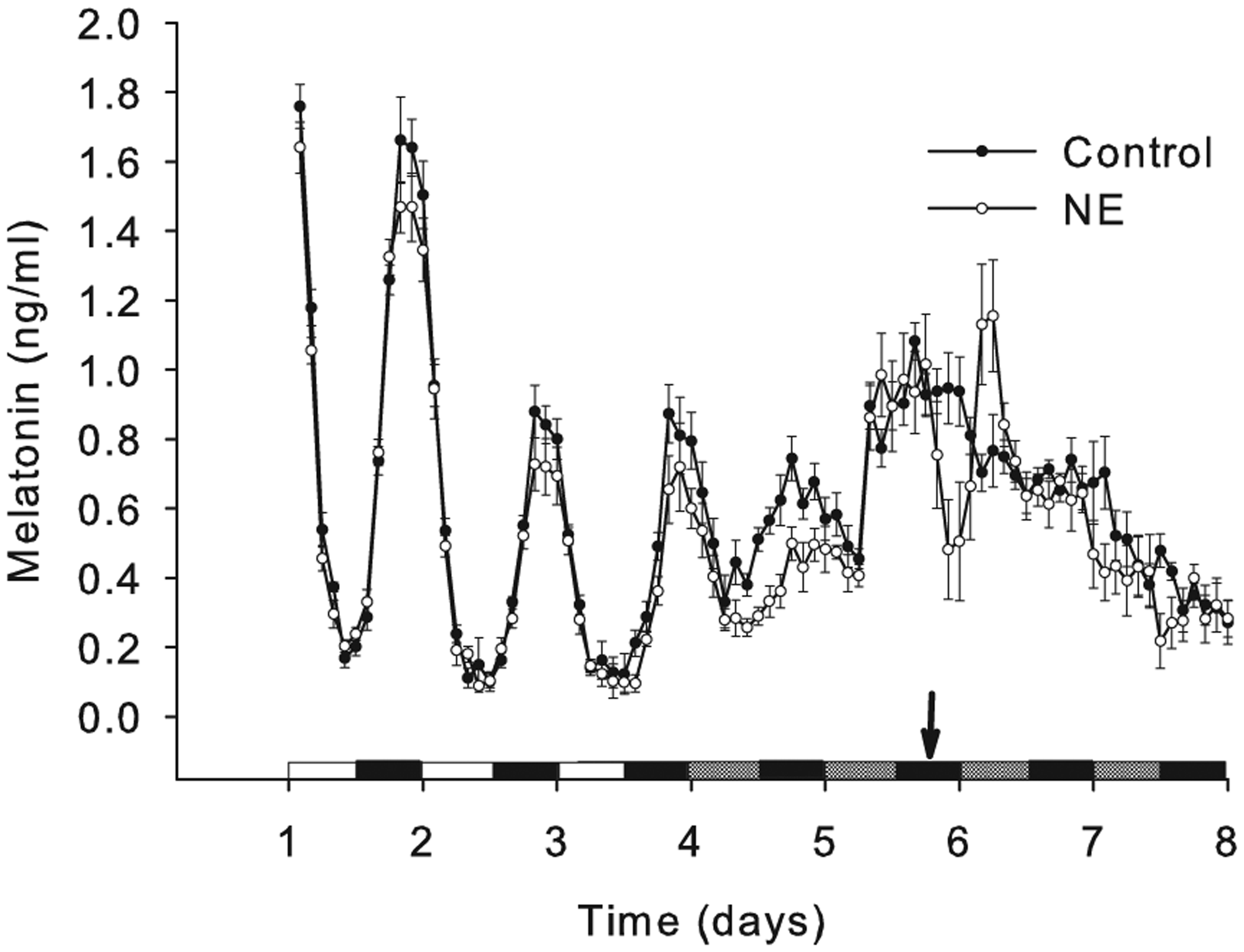
Effects of a single administration of norepinephrine (NE) on melatonin output. Twelve channels of dispersed pineal cells were cultured on a superperfusion system for 3 days under LD and 4 days under DD, and melatonin was measured every 2 h. A single pulse (100 µL of medium with NE [10−6 M] or ascorbate) was administered in the treatment group (white circles) and/or control group (black circles) on the perfusion system at CT18 (arrow) of the second cycle in DD.
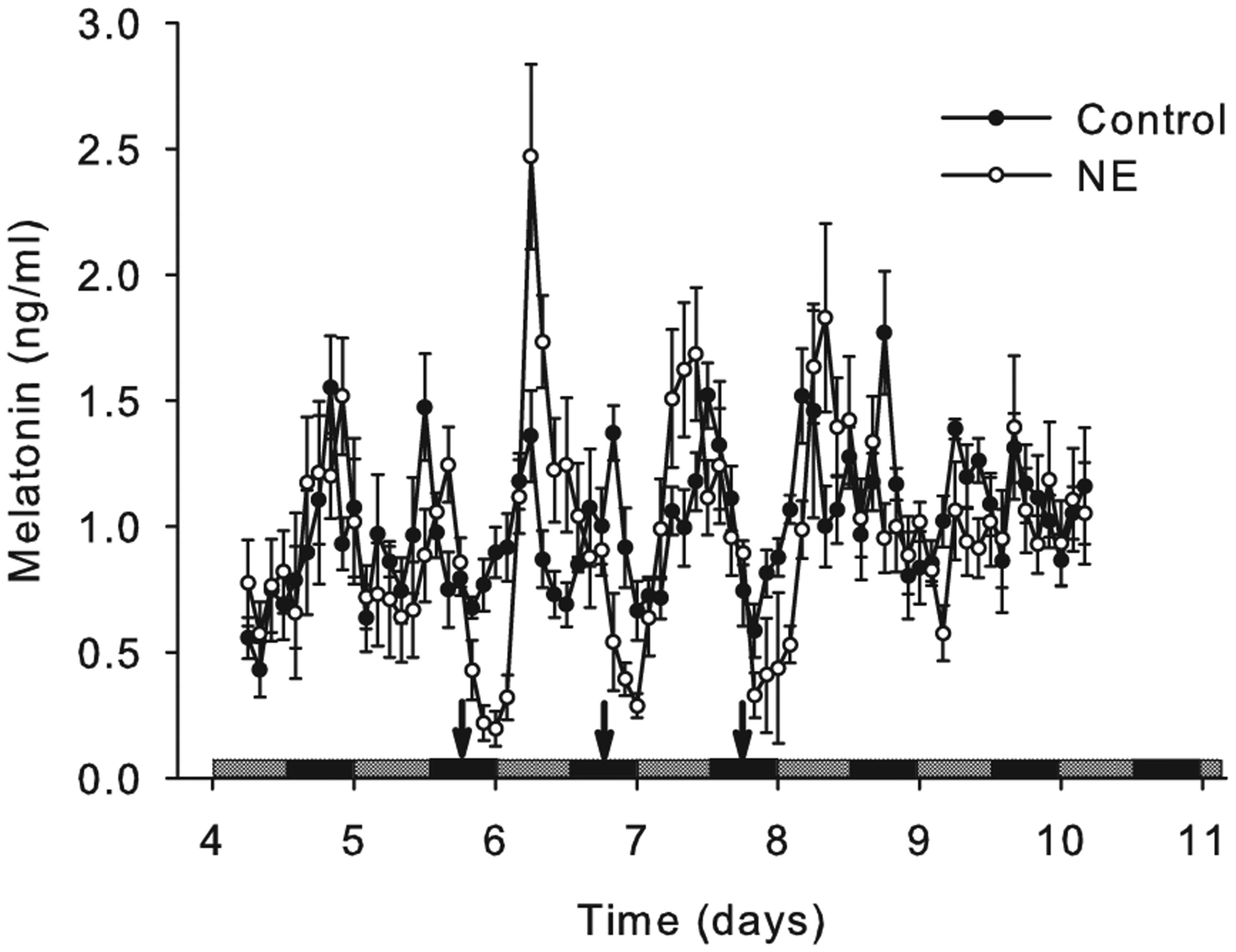
Effects of 3 consecutive norepinephrine (NE) administrations on melatonin output. Dispersed pineal cells were cultured in the superperfusion system for 3 days under LD and 7 days under DD. The plotting starts from CT6 in day 4 under DD. At CT18 of days 5 to 7 under DD, 100 µL of medium with NE (1 µM) or ascorbate was administered (the 3 arrows) to each channel of the treatment group (white circles) or control group (black circles).
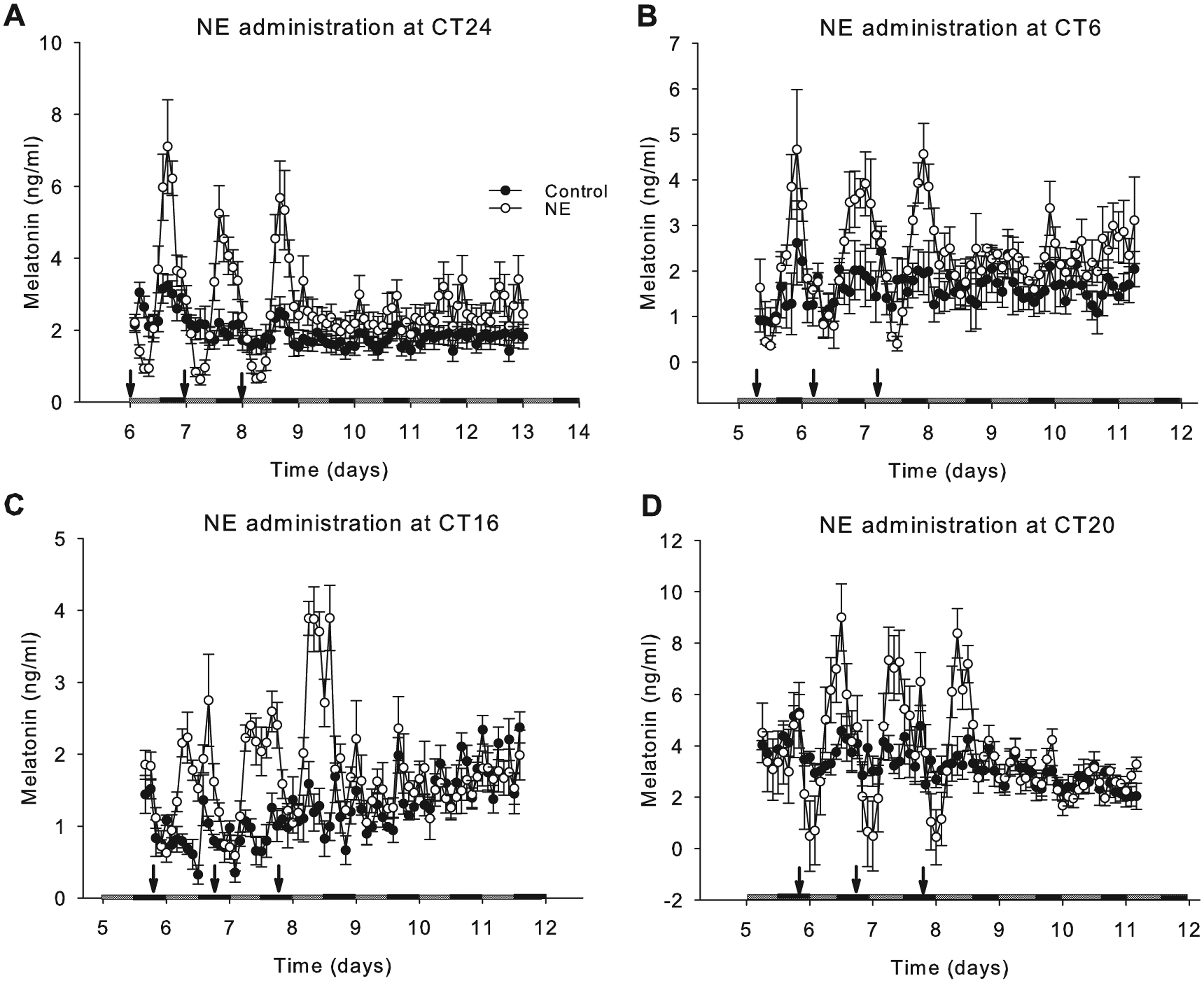
Effects of norepinephrine (NE) on melatonin output at different phases of the melatonin cycle. Dispersed pineal cells were cultured on the superperfusion system for 3 days in LD and 9 cycles in DD. The melatonin outputs were measured and plotted. The plotting starts from the day with the first NE administration in DD. At CT24 (A), CT6 (B), CT16 (C), and CT20 (D) of days 5, 6, and 7 under DD, 100 µL of medium with NE (1 µM) or ascorbate was administered (the 3 arrows) to each channel of the treatment group (white circles) and control group (black circles).
Daily Administration of NE Recovered the Melatonin Rhythm of Cultured Chicken Pineal Cells
To test whether rhythmic administration of NE could increase the amplitude of the melatonin rhythm, NE was administered for 3 cycles in DD. The melatonin output in the NE administration group was rhythmic (p < 0.05) and in the control group was not significantly rhythmic (p > 0.05). The period of melatonin rhythm in the NE administration group was 24.4 ± 0.55 h, while the period in the control group was not significant or not circadian (Figure 3).
Recovery Effect of NE Administration on the Melatonin Rhythm of Cultured Chicken Pineal Cells Was Phase Dependent
Although the melatonin output became arrhythmic in DD, it was possible that the clock system was still working. To test whether the effect of NE administration was phase dependent, NE administration was performed at 4 different phases relative to predicted phases from the LD data: the trough (CT6), 4 hours before the peak (CT16), at the peak (CT20), and 4 hours after the peak of melatonin (CT24) (Figure 4A-D). The amplitude of the recovered melatonin rhythm following NE administration at different phases was analyzed by cosinor analysis and compared statistically (Figure 5). The amplitudes stimulated by NE administration at different phases were significantly different from each other (p < 0.05 by t test) and showed a time-dependent pattern in which amplitude recovery was greatest during the late subjective night (Figure 5).
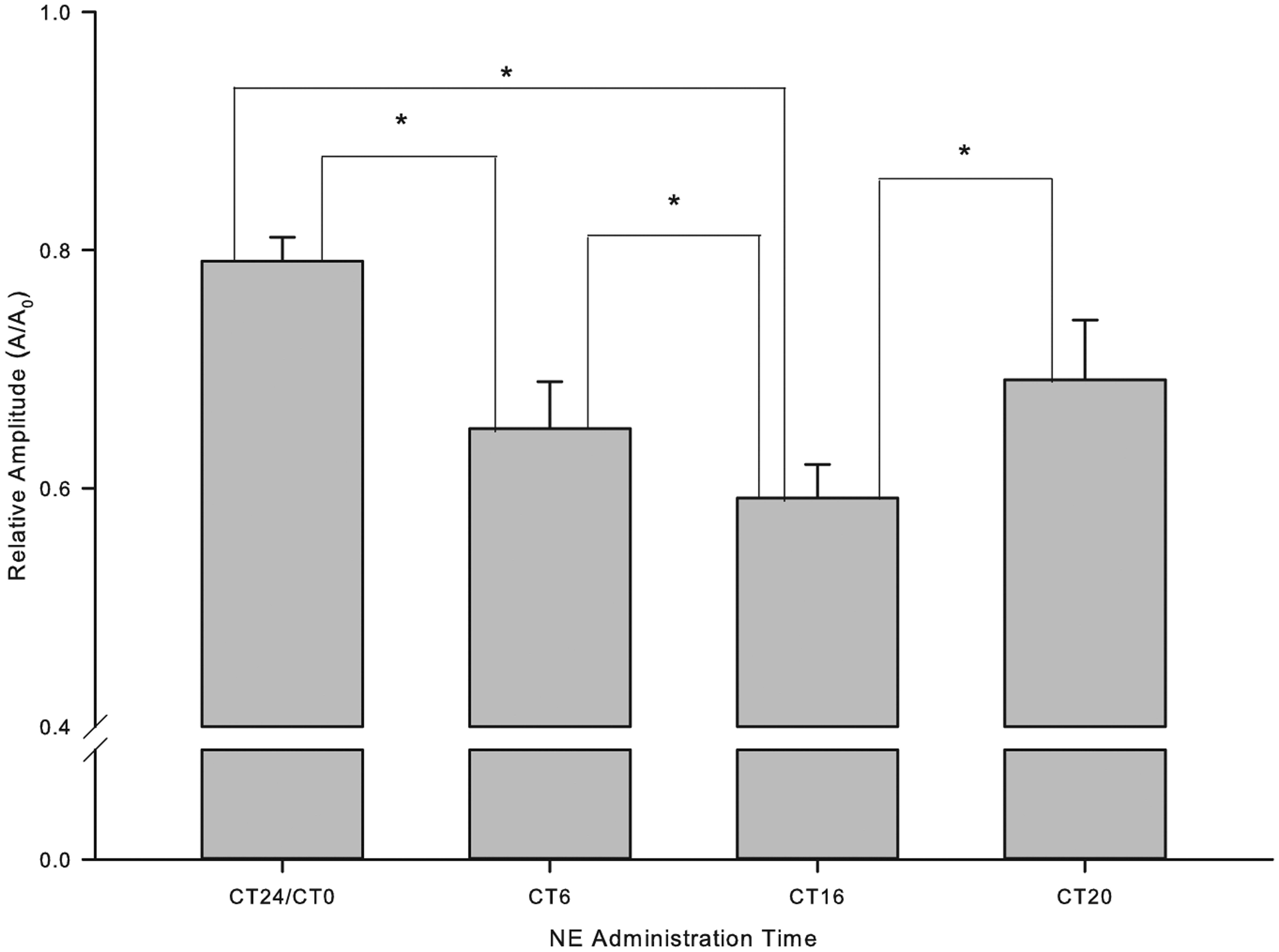
Administration of norepinephrine (NE) to dispersed pineal cells increased amplitude of the melatonin rhythm differentially depending on the phase of administration. NE was more efficacious in restoring amplitude in the late subjective night (CT20) and early subjective day (CT0/24) than at other circadian phases. The amplitude of the recovered melatonin rhythms with NE administration at different phase was analyzed by cosinor analysis and compared by the relative amplitude. The amplitudes (A) created by NE administration at each time point (CT24/0, CT6, CT16, and CT20) were normalized by the averaged concentration of the melatonin during the 3 cycles (A0) and plotted as relative amplitude (A/A0). The relative amplitudes at each time point were compared to each other by a t test. The differences between each group were significant (p < 0.05) as labeled (*) and displayed a time-dependent pattern.
Clock Genes Were Still Rhythmic in DD
Most of the genes on the array, including clock genes, were still rhythmic in DD through 1-way ANOVA followed with the Newman-Keuls multiple-comparison test (Table 1 and Figure 6; p < 0.05). However, only gAanat, gHiomt, gADCY1, and gADCY3 passed CircWave analysis (p < 0.05). As examples, the 4 clock genes gBmal1 (Figure 6A), gClock (Figure 6B), gCry1 (Figure 6C), and gPer3 (Figure 6D) exhibited high-amplitude rhythms, similar in amplitude to previous studies by this laboratory in vitro (Karaganis et al. 2008). The 2 genes on the melatonin synthesis pathway, gAanat (Figure 6E) and gHiomt (Figure 6F), together with gADCY1 (Figure 6G) and gPDE4D (Figure 6H), were also rhythmic.
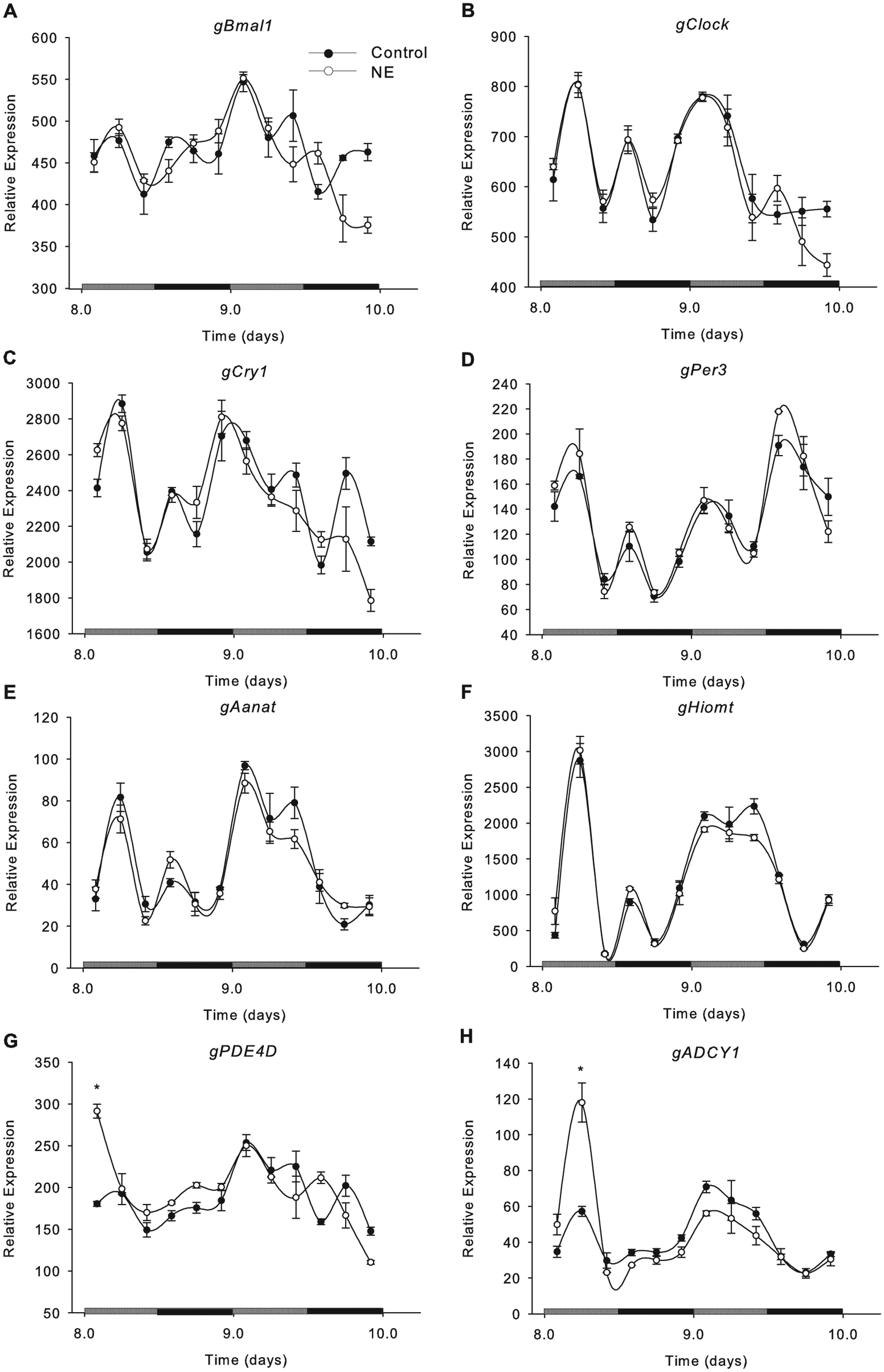
Gene expression of related genes. Circadian patterns of messenger RNA (mRNA) encoding clock proteins (A-D), melatonin synthesis enzymes (E, F), and cyclic nucleotide metabolism (G, H) persisted in dispersed pineal cells held in DD. Of these, only gPDE4D and gADCY1 were affected by exogenous norepinephrine (NE) administration. The relative mRNA expression of each gene in the control group (black circles, n = 3) and NE administration group (white circles, n = 3) was plotted on a timeline for 2 days. The NE administration significantly (p < 0.05) affected the gene expression of gPDE4D (G) and gADCY1 (H) in the treatment group (*).
PDE4D and ADCY1 Respond to NE in a Time-Dependent Pattern
NE administration did not alter the expression of most genes in the Nanostring array, including clock genes (Table 1). The exceptions were gADCY1, gPDE4D, gGNAI1, and gPurpurin (2-way ANOVA followed with a Bonferroni post hoc test, p < 0.05). Since gPDE4D and gADCY1 are important for the cAMP signaling pathway, tightly related to melatonin production in the pineal gland, 2 protein products of these genes were selected for further analysis: gPDE4D and gADCY1 (Figure 6G,H): gPDE4D mRNA was upregulated 2 h after NE administration, while gADCY1 mRNA was upregulated 6 h after NE administration. After that, there was no difference between the treatment and control groups for both genes. To investigate the response at the protein level, ADCY1 and PDE4D proteins were measured by Western blot. The results indicate the protein level of ADCY1 is significantly downregulated 6 h after NE administration and upregulated 18 h after NE administration (t test, p < 0.05), which matches the trough and the peak of melatonin rhythm (Figure 7). However, the protein expression of PDE4D did not display any significant change (t test, p > 0.05).
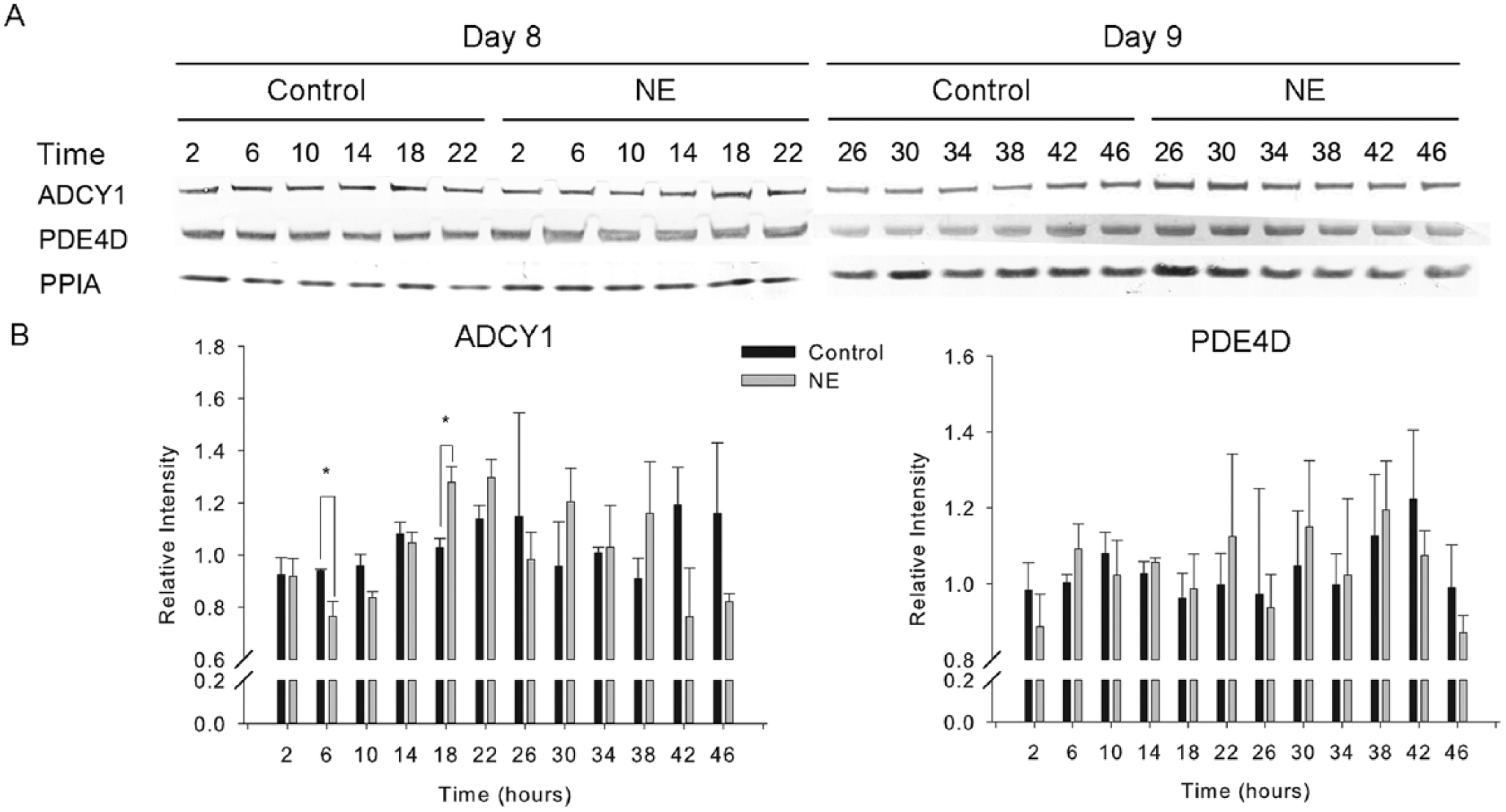
Western blot analysis of ADCY1 and PDE4D in the presence or absence of norepinephrine (NE). Pineal cells were cultured in 6-well plates at concentration of 2 × 105cells/cm2 under LD cycles. After 3 days, the cells were transferred to constant darkness for 2 days, and either NE or blank medium was administered at CT24 of day 5 in DD for 3 cycles, similar to the situation in the superperfusion system. For each time point, there were 3 replicates in the control and NE administration groups. The protein samples were collected 2 h after NE administration with a 4-h interval for 2 days. (A) Protein expression of ADCY1 and PDE4D was measured by Western blot. The numbers indicated the time (hours) after NE administration. (B) The Western blot data were quantified by ImageJ. The expression of ADCY1 and PED4D was normalized by PPIA and plotted, and the data were analyzed by paired t test at each time point. The paired t test showed a significant difference between control and NE administration groups at 6 h (*) and 18 h (**) in ADCY1 expression (p < 0.05). There was no significant difference between control and NE administration groups of PDE4D.
Discussion
The avian circadian system consists of multiple circadian pacemakers and photoreceptors whose mutual interactions are critical for self-sustained biological rhythms in behavior, physiology, and biochemistry (Cassone and Menaker, 1984; Gwinner and Brandstatter, 2001). The circadian pacemakers include the pineal gland, the hypothalamic suprachiasmatic nuclei (SCN), and the ocular retinae. Each of these components expresses circadian patterns of physiological and metabolic activity in LD and DD, which damp after several cycles in DD (Woller and Gonze, 2013). The mechanisms by which these pacemakers damp are not known. The 2 most plausible explanations are the desynchronization among individual oscillatory cells and/or a decrease in the amplitudes of the oscillatory mechanisms themselves in all cells. In the mammalian SCN, where bioluminescent and fluorescent reporter constructs reveal individual SCN cells’ gene expression, damping appears to be the result of desynchronization of reporter gene output among self-sustained oscillators in the cell population (Yamazaki et al., 2000; Ukai et al., 2007; Webb et al., 2012).
In the case of the chick pineal gland, pinealocytes synthesize and release melatonin such that melatonin levels are high in the dark phase of night and low during the day (Natesan et al., 2002). In vivo, when chicks are placed in DD, the melatonin rhythm persists for at least 10 days without a decrease in amplitude (Cassone and Menaker, 1983; Bernard et al., 1997a). When the sympathetic innervation to the pineal gland, which secretes NE during the day in LD and subjective day in DD (Cassone et al. 1986), is interrupted, pineal melatonin rhythms persist in LD but damp in DD (Cassone and Menaker, 1983). In vitro, pineal glands or dispersed cells express high-amplitude melatonin rhythms in LD (Takahashi et al., 1980; Robertson and Takahashi, 1988a, 1988b), which damp in DD (Takahashi et al., 1980; Robertson and Takahashi, 1988a, 1988b; Zatz and Mullen, 1988). In both in vivo and in vitro, administration of NE or NE agonists decreases melatonin output immediately, which in turn causes a rebound of melatonin titers (Cassone and Menaker, 1983; Zatz and Mullen, 1988).
In the present study, microfluidic cultures of pineal cells exhibited high-amplitude daily rhythms of melatonin release in an LD cycle, which persisted for 2 to 3 cycles in DD (Figures 1 and 2). However, after 2 days in DD, the amplitude of the melatonin rhythm declined, damping to arrhythmicity (Figures 1-4), suggesting that the clock mechanism itself had damped. However, clock gene expression remained robustly rhythmic with appropriate phase relationships such that negative element gPer3 was expressed 180° out of phase with positive element gBmal1 (Table 1; Figure 6A-D). Furthermore, at the mRNA level, both gAanat and gHiomt remained rhythmic, even as their collective output damped to arrhythmicity (Table 1; Figure 6E,F).
Previous studies have shown gAanat mRNA and protein levels in chicken pineal to be rhythmic in both LD and DD (Bernard et al., 1997a; Bernard et al., 1997b) and that melatonin production is parallel to the AANAT activity (Binkley et al., 1973). However, in these studies, the experiments were performed for only 1 or 2 cycles in DD before the melatonin rhythm damped. Damping of the melatonin rhythm cannot be due to flow through culture conditions, since high-amplitude rhythms can be reestablished with LD (Robertson and Takahashi, 1988a, 1988b) or norepinephrine (NE; Figures 2-4). A single administration of NE to the pineal cultures immediately decreases melatonin output in the cultures and results in a rapid rebound of melatonin 8 to 12 h later (Figure 2). This effect is amplified when NE is repeatedly administered 3 times 24 h apart, but under no circumstances could we identify a phase-dependent change in the phase or period of the subsequent rhythm (Figure 3). These observations confirm the view that while NE affects melatonin amplitude and reverses damping, it has no effect on phase-shifting the clock itself (Zatz and Mullen, 1988). This idea is punctuated by the fact that NE administration also has no effect on clock gene expression, at least at the mRNA level (Figure 6A-D). Interestingly, however, the amplitude of the rebound was phase dependent (Figure 4), in which the relative amplitude of the rebound was greater when NE was administered in the late subjective night (CT20-24) than it was during the day or early subjective night (CT6-16). This suggests that while NE has no effect on the pineal clock, the pineal clock affects the pineal’s sensitivity to NE. It is interesting to note that the phase at which NE has least effect (CT6) is the time when NE is released in vivo (Cassone et al., 1986).
What is the mechanism by which the pineal clock alters sensitivity to NE? At the mRNA level, circadian patterns in norepinephrine receptors, G-protein components, and mediators of cyclic nucleotide signaling persisted in DD, presumably under the regulation of the pineal circadian clock (Table 1; Figure 6G,H). Furthermore, NE administration selectively altered the expression of genes associated with cAMP metabolism: gPDE4D, gGNAI2, and gADCY1.
At the posttranscriptional level, the most important factor concerning melatonin production appears to be cAMP signaling pathway. In birds, NE activates α2-adrenoreceptors, which couple with a Gi GTP binding protein, inhibiting adenylate cyclase activity (Deguchi, 1979; Voisin and Collin, 1986; Pratt and Takahashi, 1987, 1988). When cAMP is low, AANAT is degraded so that melatonin production decreases. cAMP analogs stimulate melatonin production but do not have any effect on the cycles following their administration (Zatz and Mullen, 1988). In the rat, the promoter region of the Aanat gene contains one perfect cAMP response element (CRE) site, one CRE-like sequence, an inverted CCAAT box, and an activating protein 1 (AP-1) site (Baler et al., 1997; Baler et al., 1999; Burke et al., 1999), capable of binding the cAMP/PKA-dependent phosphorylated cAMP response element-binding protein (P-CREB) (Chen and Baler, 2000). In the chicken, a CRE-like sequence has also been identified in the promoter area of the gAanat gene (Haque et al., 2011). The CRE-like sequence can upregulate the transcription of gAanat in chicken retina cells, but its function in the pineal gland is still unclear. Besides ADCY, cAMP levels can also be regulated by phosphodiesterase (PDE) in rat pineal gland (Kim et al., 2007). In this study, the results revealed that the mRNA of gPDE4D and gADCY1 respond to NE administration sequentially: the mRNA of gPDE4D was upregulated 2 h after NE administration and then dropped back, while the ADCY1 mRNA was upregulated 6 h after NE administration and then dropped back.
Based on this evidence, we hypothesized that NE upregulates gPDE4D, which degrades the cAMP, leading to the trough of melatonin production, and subsequently NE upregulates gADCY1, which synthesizes the cAMP, creating the rebound of melatonin production. However, the Western blots only identified the downregulation of gADCY1 6 h after NE administration and the upregulation of gADCY1 18 h after NE administration, which was consistent with the melatonin profile after NE administration. No effect of NE could be found on gPDE4D protein content. It is possible gPDE4B activity is affected, however. Nikaido and Takahashi (1989) have shown that chick pineal cells rhythmically released cAMP in phase with the melatonin rhythm. The cAMP rhythm damped in DD, consistent with the rhythm of melatonin (Nikaido and Takahashi, 1989). It is reasonable to hypothesize that gADCY1 is key to damping and NE-induced recovery of the melatonin rhythm. Our results indicate that the melatonin rhythm damps in parallel with the damping of the gADCY1 rhythm at the protein level, even when gAanat mRNA is still rhythmic. It is possible that NE recovers the melatonin rhythm through creating a rebound in gADCY1.
Our results further revealed that the amplitude of the melatonin rebound created by NE in DD was phase dependent. The phase-dependent effects may result from rhythmic clock gene expression in DD, although direct, causative evidence of this is wanting. The NE receptor gADRA2A and the receptor subunit gGNAI1 and gGNAI2 are all rhythmic in vitro, indicating that they are under the control of an internal clock. Although the clock does not directly maintain the melatonin output of the pineal cells, it still functions in gating the response of pineal cells to the NE input. The role of clock genes in pineal physiology is still not fully understood in birds or mammals. For example, clock genes in the rat pineal gland are rhythmically expressed, but melatonin production is not rhythmic in vitro at all and increases only in response to NE stimulation through the cAMP pathway (Fukuhara and Tosini, 2008; Bailey et al., 2009). Our results show that clock genes in the pineal gland, as well as other peripheral tissues, might serve by gating the tissue’s response to received signals, indicating a common mechanism in which the clock genes do not directly control physiological outputs but gate the response to external stimulation. The details of how clock genes generate the overall rhythmic gene expression in the chicken still need to be determined.
In summary, our results demonstrate that damping of the melatonin rhythm in cultured pineal cells occurs at the posttranscriptional level, despite rhythmic expression of clock genes, gAanat and gHiomt. The expression of gADCY1 at the protein level was likely the main factor in the damping of the melatonin rhythm in DD as well as the recovery of the melatonin rhythm by NE. Although the clock genes did not directly control the melatonin production in DD, they continued to gate the response of melatonin production to NE.
Footnotes
Acknowledgements
We wish to express appreciation to Jiffin Paulose for advice and suggestions, to Clifford Harpole and John Wright for revisions of the paper, and to Ruochun Zhao and Angelina Cheng for assistance in experiments. This research was supported by the University of Kentucky, NIH P01 NS39546, and HHMI “Sustaining Excellence” Science Education grant 52008116 to VMC.
Conflict of Interest Statement
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.