
Abstract
Honey bee (Apis mellifera) workers emerge from the pupae with no circadian rhythms in behavior or brain clock gene expression but show strong rhythms later in life. This postembryonic development of circadian rhythms is reminiscent of that of infants of humans and other primates but contrasts with most insects, which typically emerge from the pupae with strong circadian rhythms. Very little is known about the internal and external factors regulating the ontogeny of circadian rhythms in bees or in other animals. We tested the hypothesis that the environment during early life influences the later expression of circadian rhythms in locomotor activity in young honey bees. We reared newly emerged bees in various social environments, transferred them to individual cages in constant laboratory conditions, and monitored their locomotor activity. We found that the percentage of rhythmic individuals among bees that experienced the colony environment for their first 48 h of adult life was similar to that of older sister foragers, but their rhythms were weaker. Sister bees isolated individually in the laboratory for the same period were significantly less likely to show circadian rhythms in locomotor activity. Bees experiencing the colony environment for only 24 h, or staying for 48 h with 30 same-age sister bees in the laboratory, were similar to bees individually isolated in the laboratory. By contrast, bees that were caged individually or in groups in single- or double-mesh enclosures inside a field colony were as likely to exhibit circadian rhythms as their sisters that were freely moving in the same colony. These findings suggest that the development of the circadian system in young adult honey bees is faster in the colony than in isolation. Direct contact with the queen, workers, or the brood, contact pheromones, and trophallaxis, which are all important means of communication in honey bees, cannot account for the influence of the colony environment, since they were all withheld from the bees in the double-mesh enclosures. Our results suggest that volatile pheromones, the colony microenvironment, or both influence the ontogeny of circadian rhythms in honey bees.
Honey bee workers show a postembryonic development of circadian rhythms (reviewed in Eban-Rothschild and Bloch, 2012; Moore, 2001) that is reminiscent of that of infants of humans and other primates (Rivkees, 2003) but contrasts with most insects that emerge from the pupae with strong circadian rhythms (Fantinou et al., 1998; Fleury et al., 2000; Saunders, 2002). Individually isolated young bees typically do not exhibit circadian rhythms in activity, metabolism, or clock gene expression during the first days after emergence (Bloch and Meshi, 2007; Bloch et al., 2002; Spangler, 1972; Stussi and Harmelin, 1966; Toma et al., 2000). The onset of circadian rhythms in locomotor activity typically occurs during the first 2 weeks of adult life, with significant individual and colony-level variability (e.g., Bloch and Meshi, 2007; Bloch et al., 2002; Toma et al., 2000). The ontogeny of circadian rhythms is endogenous because it occurs under constant conditions, and the rhythms persist (“free-run”) with a period of about 24 h.
Very little is known about the internal and external factors regulating the ontogeny of circadian rhythms in honey bees or in other organisms. It has been suggested that the ontogeny of circadian rhythms in honey bee workers is associated with their age-related transition from nursing to foraging activities (Moore et al., 1998; Toma et al., 2000). In field colonies, young bees care for (“nurse”) the brood and are typically active around the clock with no circadian rhythms; a pattern of activity that may enable them to better care for the developing larvae (Bloch, 2010; Eban-Rothschild and Bloch, 2012; Moore, 2001). Older bees that typically forage for nectar and pollen outside the hive have strong circadian rhythms that are necessary for timing visits to flowers, for time-compensated sun-compass orientation, and for dance language communication (Bloch, 2010). However, manipulating neuroendocrine signals that are known to advance the age of first foraging, and were thus hypothesized to affect circadian rhythms, did not advance the onset of circadian rhythms in locomotor activity for individually isolated bees (e.g., juvenile hormone, octopamine, and cGMP; Ben-Shahar et al., 2003; Bloch and Meshi, 2007; Bloch et al., 2002). There is some evidence suggesting that the ontogeny of circadian rhythms in honey bees is influenced by the social environment. For example, Meshi and Bloch (2007) found that the onset of circadian rhythms in locomotor activity occurred earlier in young bees housed with older foragers outside the colony compared with sister bees housed with a similar number of young bees. However, since activity in this study was monitored for bees in a group, it was impossible to determine whether the foragers’ influence stemmed from accelerating the development of the circadian system of the young bees or from social masking (e.g., the activity of the young bees was stimulated by the foragers activity or heat production). In the current study we tested the influence of different components of the colony environment on the later expression of circadian rhythms in locomotor activity by young bees. The expression of circadian rhythms in locomotor activity and their strength were used as indices for the developmental state of the circadian system. We reared honey bees in various social environments during their first 24-48 h after emergence and later monitored their locomotor activity individually. This experimental design allowed us to measure influences on circadian rhythms that are not masked by the presence of other bees. We found that the ontogeny of circadian rhythms in locomotor activity is affected by the colony environment and that this influence does not require direct contact with other bees or the brood.
Materials and Methods
Bees
We kept honey bee colonies according to standard beekeeping techniques in the Bee Research Facility at the Edmond J. Safra campus of the Hebrew University of Jerusalem, Givat-Ram, Jerusalem, Israel. We marked newly emerged bees with a paint-dot on their thorax within 30 min from emergence and randomly assigned them to one of the experimental conditions (see below and supplementary online material, Suppl. Fig. S1). We provisioned all the bees (besides those freely moving in the colony, see below) with ad libitum sugar syrup (50%, w/w) and pollen. The bees that were placed in the laboratory were kept in a dark environmental chamber (31 ± 1 °C; relative humidity = 55% ± 5%). After 24 or 48 h, during which the bees experienced the experimental social environments, we collected the focal bees and placed each one of them in an individual locomotor activity monitoring cage made of a modified Petri dish (diameter = 90 mm, height = 15 mm) and provisioned with ad libitum sugar syrup (see below). Since the bees that were introduced to field colonies were exposed to daylight twice (during the introduction and the collection from the hive), we also exposed the bees from the other treatments to a similar daylight experience. The exposure to light was less than a minute during the introduction and lasted 5-10 min during the collection.
Locomotor Activity
We placed the monitoring cages with the focal bees in an environmental chamber (29 ± 1 °C; relative humidity = 45% ± 5%), which was illuminated by constant dim red light. Data were collected continuously at a frequency of 1 Hz. We determined circadian rhythmicity based on 4 consecutive days, using a χ2 periodogram analysis (ClockLab circadian analyses software, Actimetrics, USA) for 10-min bins. All the bees besides the foragers were 3-6 days of age during the period for which we analyzed circadian rhythms. For additional details see supplementary online material, Shemesh et al (2007), and Shemesh et al. (2011).
Experiment 1: Influence of Early Experience with the Colony Environment on Circadian Rhythms in Locomotor Activity for Individually Isolated Bees
We assigned newly emerged bees to one of the following treatments: (1) 48 h in a field colony (“Colony 48h”), (2) 24 h in the same field colony (“Colony 24h”), or (3) 48 h in an individual cage in the laboratory (“Isolation 48h”). The individual cages were similar to the monitoring cages (see above). We marked newly emerged bees during 2 consecutive days (for treatments 1 and 2). Between these 2 days, the comb with the pupae was placed in a host field colony. The cages with the individually isolated bees were placed in a dark environmental chamber (see above). We used foragers, which typically exhibit strong circadian rhythms (Eban-Rothschild and Bloch, 2012; Moore, 2001) as a positive control. Foragers were identified as bees returning to the hive with undamaged wings and pollen loads on their hind legs. We repeated the experiment 3 times, each with a different source colony (S73, H1, and H14).
Experiment 2: Influence of Previous Interactions with Same-Age Bees outside the Colony on Circadian Rhythms in Locomotor Activity for Individually Isolated Bees
We assigned newly emerged bees to (1) 48 h in a field colony (“Colony 48h”), (2) 48 h in a wooden cage (11 × 10 × 4.5 cm) with 30 same-age sister bees in the laboratory (“Group 48h”), and (3) 48 h in an individual cage in the laboratory (“Isolation 48h”) (see Experiment 1). We used foragers as a positive control (see above). We repeated this experiment 3 times, each with a different source colony (H11, H6, and H12).
Experiment 3: Influence of Group Caging in Single- and Double-Mesh Enclosures in the Colony on the Later Expression of Circadian Rhythms in Locomotor Activity
We assigned newly emerged bees to (1) 48 h in a field colony (“Colony 48h”), (2) 48 h in a single-mesh enclosure with 30 same-age sister bees inside the same field colony (“SM 48h”), (3) 48 h in a double-mesh enclosure containing 30 same-age sister bees inside the colony (“DM 48h”), and (4) 48 h in a wooden cage with 30 same-age sister bees in the laboratory (“Group 48h”). The enclosures (11 × 10.5 × 2 cm) were made of mesh with 8 holes per inch and were embedded on both sides of an empty comb. For the DM treatment, the enclosure was surrounded by a larger cage (14 × 13.5 × 3 cm) made of mesh (8 holes per inch, same as above) placed 1.5 cm away from the first mesh. Both the SM and DM enclosures prevented the caged bees from interacting with the brood but allowed exposure to the microenvironment (e.g., light, temperature, humidity, and CO2 levels) and the odors of the hive. The DM enclosure, but not the SM, also prevented direct contact with bees outside the enclosure. We placed the frame with the focal bees in the center of the hive, such that the caged bees were flanked by brood-containing honeycombs. We provisioned all cages (inside and outside the colony) with sugar syrup and pollen. We repeated this experiment 3 times, each with a different source colony (HS76, H2, and S85).
Experiment 4: Influence of Caging Bees Individually in Single- or Double-Mesh Enclosures inside the Hive on the Later Expression of Circadian Rhythms
We assigned newly emerged bees to one of the following treatments: (1) 48 h in a field colony (“Colony 48h”), (2) constrained individually to a single-mesh (same as above) enclosure in the same colony during 48 h (“I-SM 48h”), (3) constrained individually to a double-mesh enclosure in the colony during 48 h (“I-DM 48h”; see supplementary online material; Suppl. Fig. S2), and (4) 48 h in an individual cage (7.5 × 2.5 × 2.5 cm) in the laboratory (“Isolation 48h”). The side walls of the individual cages for this treatment were made of transparent glass. The ceiling (2.5 × 2.5) was made of mesh (same as above) allowing improved ventilation. Since the individual glass cages were placed one next to the other, we cannot rule out the possibility that the bees sensed olfactory or vibratory cues from neighboring cages.
For caging bees individually inside the hive, we constructed a horizontal wooden separation in the center of an empty honeycomb frame (without a comb) that divided the frame into 2 similar compartments (10 × 3 × 43.4). In each compartment we placed a row of 11 mesh cages (7.5 × 2.5 × 2.5 cm), with a gap of 1.5 cm between each pair of adjacent cages (Suppl. Fig. S2). For the DM treatment, we covered the lower part of the frame with an additional mesh placed 1.5 cm away, after we introduced the bees to the cages. In each trial we introduced 33 bees into 3 host colonies—11 bees for each of the treatments. All the focal bees were sisters obtained from the same source colony. We placed the frames with the cages in the center of the hive, such that they were surrounded by brood-containing honeycomb frames (Suppl. Fig. S2b). We provisioned all cages (inside and outside the hive) with ad libitum sugar syrup and pollen. We repeated this experiment 2 times, each with a different
source colony (H2b and H11b).
Results
Experiment 1: Influence of Early Experience with the Colony Environment on Circadian Rhythms in Locomotor Activity for Individually Isolated Bees
Foragers typically had strong circadian rhythms (Figs. 1d and 2) and the highest level of locomotor activity (Suppl. Fig. S3). Young bees that were individually isolated for their first 2 days after emergence were less likely to exhibit circadian rhythms compared with their sister bees that experienced the colony environment during the same period (Figs. 1a, c, and 2a). Bees that experienced the colony environment for only 24 h were less likely to exhibit circadian rhythms in locomotor activity compared with their sisters who stayed in the same colony for 48 h (Figs. 1b and 2a). Age cannot account for this finding because circadian rhythms were determined when the bees from the 2 groups were 3-6 days of age. The strength of circadian rhythms (power) tended to be higher for the individually isolated bees than for the bees that experienced the colony environment (this comparison includes only bees with statistically significant rhythms; Fig. 2b). The activity level was higher for the individually isolated bees compared with the bees that experienced the hive environment for 24 h in all 3 repetitions and compared with those that experienced the colony environment for 48 h in only 1 repetition (Suppl. Fig. S3). The free-running period (FRP) was similar for bees experiencing the different social environments (Suppl. Table S1). Taken together, these results suggest that early experience with the colony environment has a strong influence on the later expression of circadian rhythms in young honey bees and that these influences vary with the duration of experience.
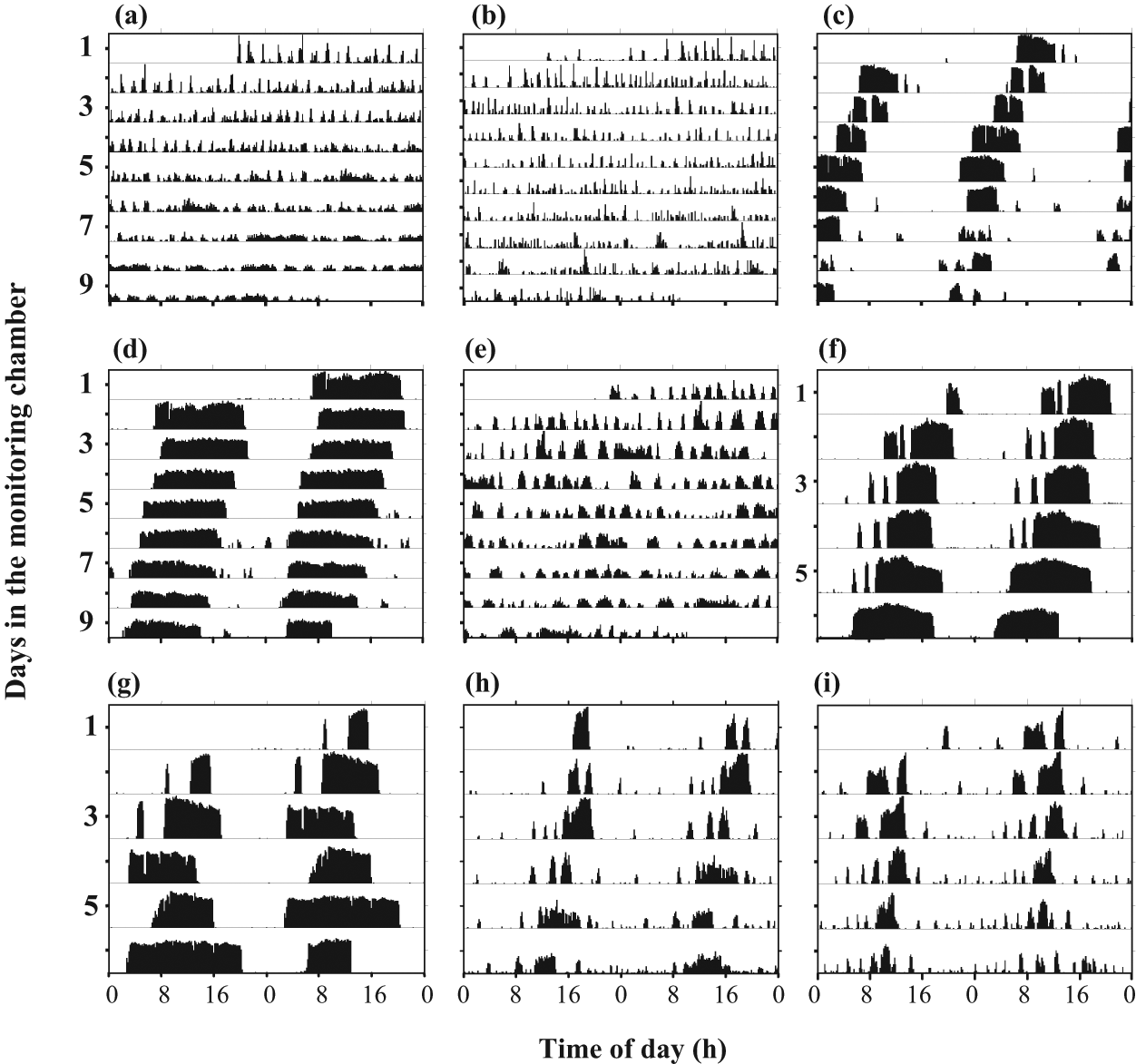
Representative double-plot actograms. The y-axis depicts days in the monitoring chamber. The height of the bars within each day corresponds to the level of locomotor activity in a 10-min bin. (a) A 3-day-old bee that spent its first 48 h individually isolated in the laboratory. (b) A 2-day-old bee that experienced the colony environment for the first 24 h post pupal eclosion. (c) A 3-day-old bee that experienced the colony environment for the first 48 h post eclosion. (d) A forager (~3 weeks old) collected from a field colony. (e) A 3-day-old bee that was caged in the laboratory with 30 additional same-age bees for the first 48 h post eclosion. (f) A 3-day-old bee confined with 30 additional same-age bees to a single-mesh enclosure inside a field colony for the first 48h post eclosion. (g) Same as (f), but in a double-mesh enclosure. (h) Same as (f), but in an individual mesh-enclosure (i) Same as (g), but in an individual mesh-enclosure. The bees in c, d, and f-i show significant circadian rhythms in locomotor activity from day 1. Bees in a, b, and e do not show circadian rhythms in locomotor activity.
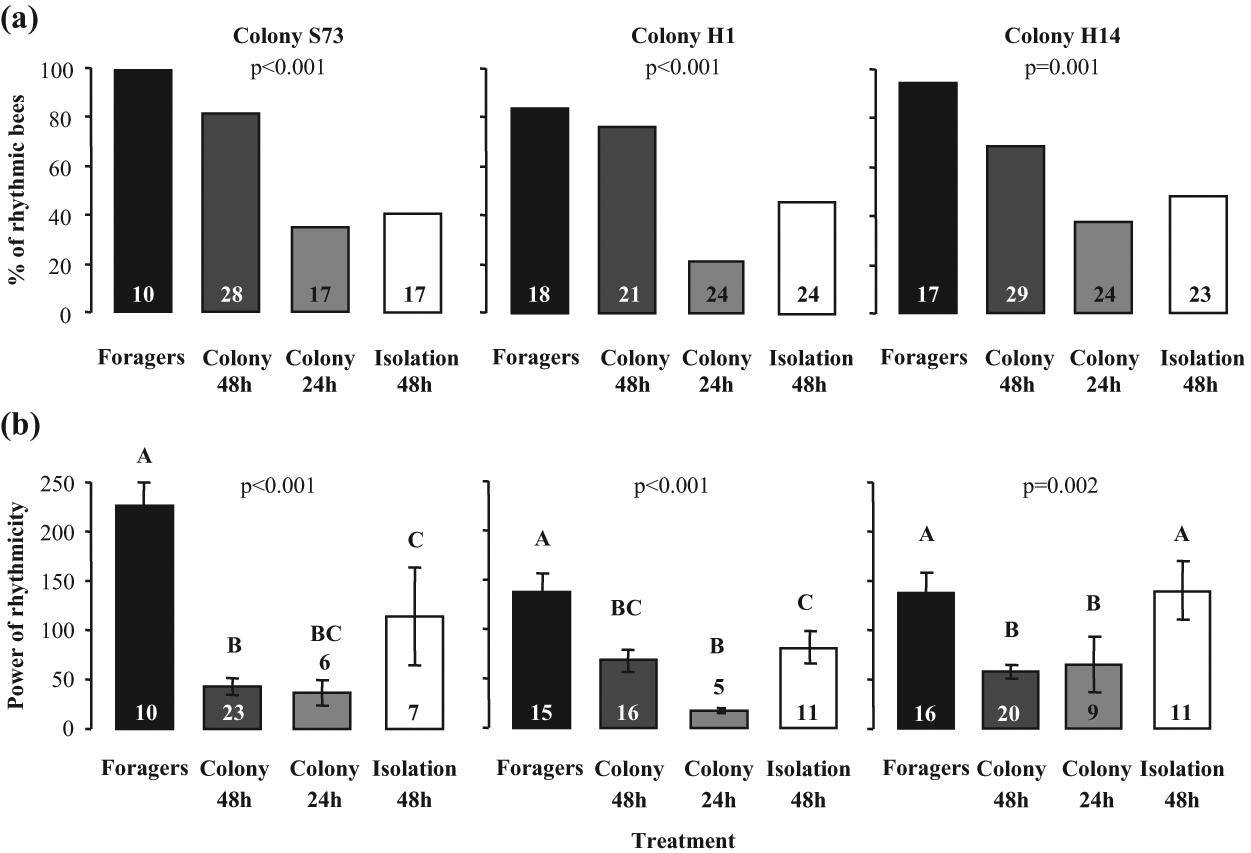
The influence of early experience with the colony environment on the later expression of circadian rhythms in locomotor activity. (a) The percentage of bees with significant circadian rhythms in locomotor activity (Pearson chi-square tests). (b) The power (mean ± SE) of circadian rhythms in locomotor activity for bees with statistically significant rhythms. Groups with different capital letters differ significantly (1-way ANOVA test, LCD post hoc test, p < 0.05). “Foragers” = bees of unknown age collected at the hive entrance with pollen loads in their corbiculates. “Colony 48h” = 3-day-old bees that were freely-moving in a field colony during their first 48 h post eclosion. “Colony 24h”= 3-day-old bees that were freely-moving in a field colony during their first 24 h post eclosion. “Isolation 48h” = 3-day-old bees that were individually isolated in the laboratory from emergence. For additional details see Materials and Methods. The analyses are based on 4 consecutive days, when the young bees were 3-6 days of age. The plots represent results from 3 different trials, each with bees from a different source colony. Numbers inside or above bars depict sample size.
Experiment 2: Influence of Previous Interactions with Same-Age Bees outside the Colony on Circadian Rhythms in Locomotor Activity for Individually Isolated Bees
As in Experiment 1, most of the foragers and the young bees that experienced the colony environment during their first 48 h after emergence showed significant circadian rhythms in locomotor activity (Fig. 3a). However, foragers had stronger rhythms and higher levels of activity compared with their younger sisters (Figs. 3b and S4). Young bees that were isolated individually were less likely to show circadian rhythms in locomotor activity compared with their sisters that experienced the colony environment during this period (Fig. 3a). Young bees that were placed in groups outside the colony during their first 48 h after emergence (“Group 48h”) were similar overall to those isolated individually (Figs. 1e and 3). In the repetitions with bees from colonies H11 and H12, there were no differences in the FRP for the bees from the different treatments, whereas foragers had a longer FRP in the third repetition (Colony H6; Suppl. Table S2). The level of activity was similar for the 3 groups of young bees (Suppl. Fig. S4). These results suggest that social contact with same-age bees outside the natural context of the colony does not have the same influence on the development of circadian rhythms as the entire colony environment.
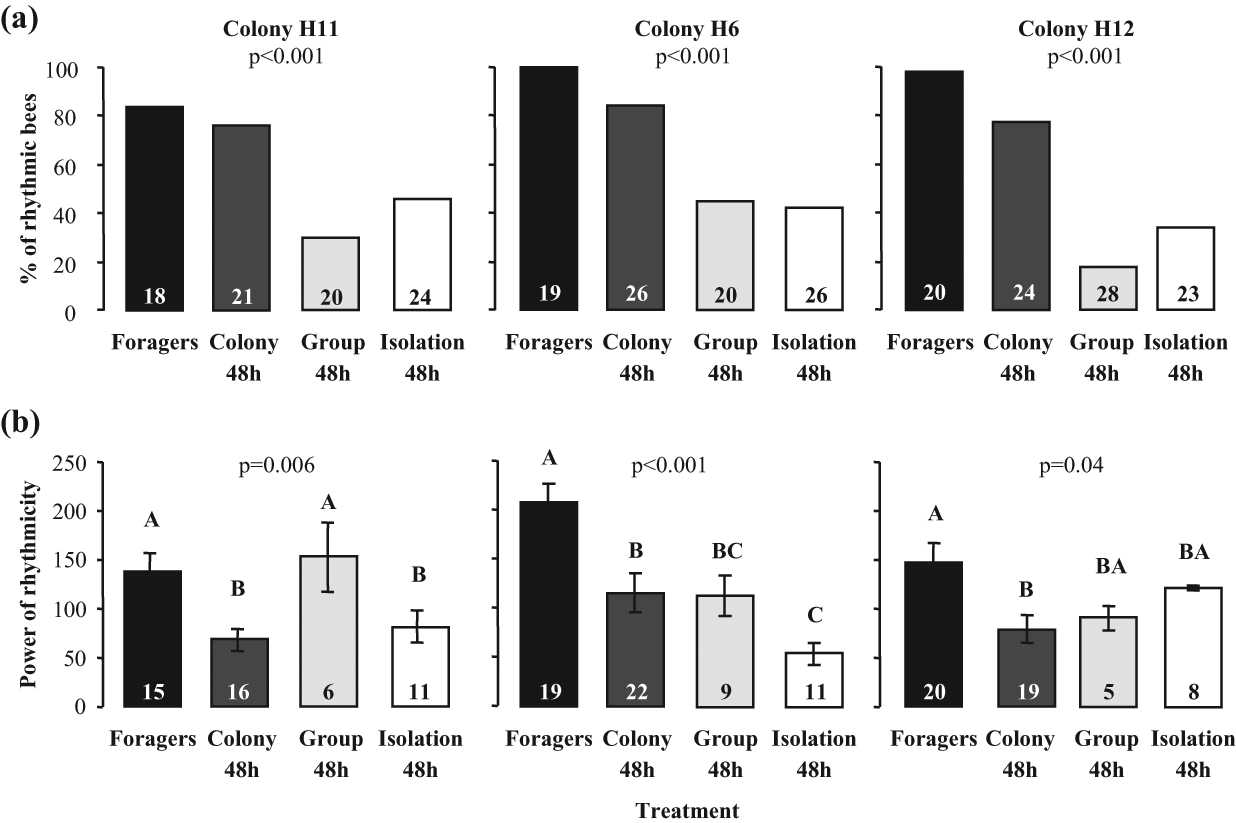
The influence of previous interactions with same-age bees in the laboratory on the later expression of circadian rhythms in locomotor activity. (a) The percentage of bees with significant circadian rhythms in locomotor activity. (b) The power of circadian rhythms in locomotor activity for rhythmic bees. “Group 48h” = 3-day-old bees that were caged with 30 same-age bees in the laboratory during their first 48h post eclosion. Other details as in Figure 3.
Experiment 3: Influence of Group Caging in Single- and Double-Mesh Enclosures in the Colony on the Later Expression of Circadian Rhythms
The bees that moved freely in the colony and those confined to a single- or double-mesh enclosure were similarly likely to show circadian rhythms and had a similar power of rhythmicity (Figs. 1f, 1g, 4, and S5a). By contrast, as in Experiment 2, bees that were caged outside the colony with a group of 30 bees were less likely to exhibit circadian rhythms compared with their sisters that experienced the colony environment (Fig. 4; the p value was 0.06 in the trial with colony S85). There were no significant differences in the strength of circadian rhythms between the experimental groups, but in the repetitions with colonies HS76 and H2, there was a nonsignificant trend toward lower power for the laboratory-caged bees (Suppl. Fig. S5a). Similarly, there were no consistent differences in activity level or FRP between the experimental groups (Suppl. Fig. S5b and Suppl. Suppl. Table S3). Since the double-mesh prevented direct contact with the brood, the queen, and bees moving freely in the hive, our results indicate that these factors cannot account for the influence of the colony environment on the later expression of circadian rhythms in locomotor activity.
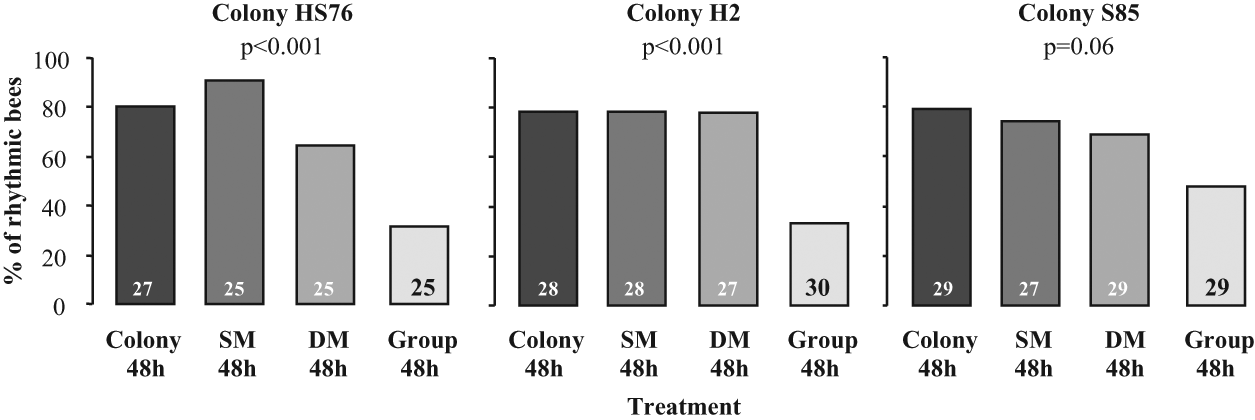
The influence of group caging in single- or double-mesh enclosures in the colony on the later expression of circadian rhythms in locomotor activity. The plots depict the percentage of bees with significant circadian rhythms in locomotor activity. “SM 48h” = 3-day-old bees that were caged with 30 same-age bees in a single-mesh enclosure in the colony during their first 48 h post eclosion. “DM 48h” = same as previous, but caged in a double-mesh enclosure. Other details as in Figures 3 and 4.
Experiment 4: Influence of Caging Bees Individually in Single- or Double-Mesh Enclosures inside the Hive on the Later Expression of Circadian Rhythms
To examine the possible influences of social interactions inside the mesh cages, we performed an additional experiment in which newly emerged bees were isolated individually in single- or double-mesh cages inside the hive. As in Experiments 1 and 2, bees isolated individually in the laboratory were less likely to show circadian rhythms compared with their same age sister bees that experienced the colony environment (Fig. 5a). This includes the repetition with colony H11b in which the percentage of rhythmic bees was unusually high in comparison with bees from the 7 other source colonies used in this study. In this trial circadian rhythms were also weaker (lower power) for the individually isolated bees compared with the bees from the 3 other experimental groups (Fig. 5b). The bees that were confined to a single- or double-mesh individual enclosure, or were freely moving in the hive, were similarly likely to exhibit circadian rhythms and had a similar power of rhythmicity (Fig. 5). There were no differences in FRP between the experimental groups, but the level of activity tended to be higher for the bees is the SM and DM enclosures (Suppl. Table S4 and Suppl. Fig. S6). These results strengthen our findings that direct contact with other bees is not needed for the influence of the colony environment on the development of circadian rhythms in locomotor activity.
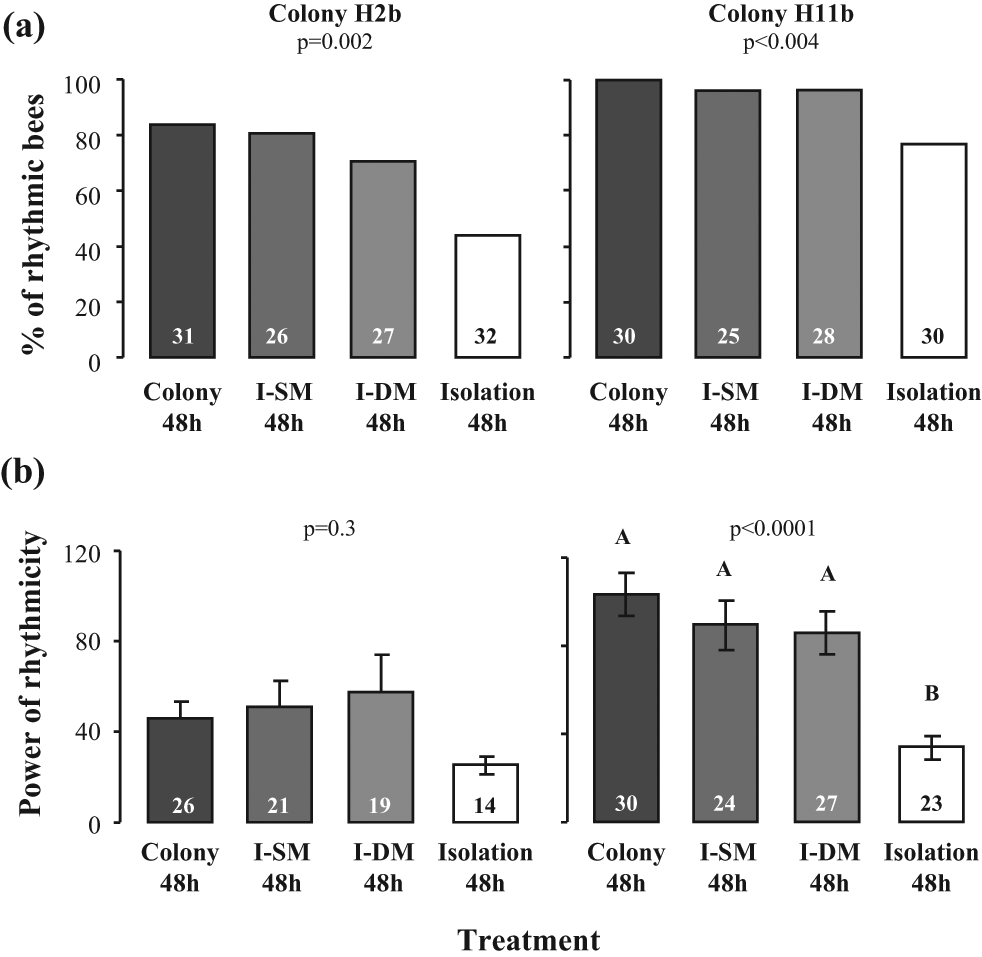
The influence of caging bees individually in single- or double-mesh enclosures inside the hive on the later expression of circadian rhythms in locomotor activity. (a) The percentage of bees with significant circadian rhythms in locomotor activity. (b) The power of circadian rhythms in locomotor activity for rhythmic bees. “I-SM 48h” = 3-day-old bees that were individually caged in a single-mesh enclosure in the colony during their first 48h post eclosion. “I-DM 48h” = same as previous, but caged in a double-mesh enclosure. Other details as in Fig. 3.
Discussion
Our study shows that early experience of the colony environment has profound influence on the later expression of circadian rhythms in locomotor activity in young honey bees. In experiments with bees from 11 different source colonies, young bees that experienced the colony environment for the first 48 h after emergence were more likely to show circadian rhythms in locomotor activity compared with their same-age sister bees that were removed from the colony for the same period. The young bees that experienced the colony environment for 48 h had, nevertheless, weaker circadian rhythms than their older sister foragers, suggesting that their circadian system was not yet fully matured. Twenty-four hours in the colony or 48 h in the laboratory with a group of same-age sister bees was not sufficient for obtaining a similarly high percentage of bees showing circadian rhythms in locomotor activity. The influence of the colony environment was apparent even in bees that were in the hive but caged individually or with 30 other bees in double-mesh enclosures. These in-hive caged bees had no direct contact with the queen, the brood, or other worker bees but nevertheless were as likely to exhibit circadian rhythms as their sisters freely moving in the colony. Therefore, tactile signals, contact pheromones, and trophallaxis (i.e., the transfer of food or other fluids among conspecifics), which are all important means of communication in honey bees, cannot account for the influence of the colony environment on the development of circadian rhythms. Our results suggest that volatile pheromones, the colony microenvironment, or both influence the development of circadian rhythms in locomotor activity in young honey bees.
Young bees developing in a colony typically attend the brood around-the-clock, and therefore their activity profile does not inform the observer on the developmental state of their circadian system (Bloch, 2010). Thus, the ontogeny of circadian rhythms has been commonly studied in bees that were isolated individually in the laboratory shortly after emergence. Under these conditions, worker bees typically show circadian rhythms in locomotor activity for the first time only when they are around 1 or 2 weeks of age (Eban-Rothschild and Bloch, 2012; Moore, 2001). By contrast, we show that most 3-day-old bees that experienced the colony environment during their first 48 h showed significant circadian rhythms in locomotor activity when removed to the laboratory. Nevertheless, these bees still had weaker circadian rhythms compared with their sister foragers (typically older than 3 weeks of age). Furthermore, the propensity to show circadian rhythms was lower for bees that experienced the colony environment for only 24 h. These results suggest that the circadian system of 3-day-old bees is not yet fully matured. Studies on brain clock gene expression patterns provide another line of evidence for the ontogeny of the circadian system in bees in free-foraging colonies. Brain period mRNA levels show weaker oscillations and overall lower levels in 1-day-old bees relative to older bees (Bloch et al., 2004). Furthermore, the amplitude of Period and Cryptochrome transcript oscillation is larger for foragers than for other groups of younger bees that are not nurses (Shemesh et al., 2007; Shemesh et al., 2010). Assuming that the expression of locomotor activity rhythms in the laboratory informs us about the functioning of the circadian clock, our findings suggest that the circadian system of bees is developing after their emergence from the pupae. The pace of development is influenced by the environment experienced by the bee and differs between the colony and the laboratory.
What in the colony environment accounts for the accelerated development relative to the laboratory? Given the evidence that social contact and food exchange with older bees are necessary for the proper development of young bees (Naiem et al., 1999), one can hypothesize that in their absence the development of the circadian system is compromised. Our findings, however, are not consistent with this hypothesis. Bees placed individually in a double-mesh enclosure in the hive, and thus prevented from direct contact and food exchange with other bees, were similarly likely to exhibit circadian rhythms as bees freely moving in the colony. The bees caged in DM enclosures could sense volatile odors, including pheromones, and these may have influenced the development of their circadian rhythms. Chemical communication is a major channel of communication in honey bees, and pheromones modulate many aspects of their life (Le Conte and Hefetz, 2008; Wilson, 1971). An additional component of the colony that could have influenced the ontogeny of circadian rhythms is the hive microenvironment. Studies on the environmental influences on the ontogeny of circadian rhythms commonly focus on light (e.g., Barrett and Page, 1989; Kaneko and Cahill, 2005; Page, 1991). Although light might have some influence on the development of circadian rhythms in honey bees (Bloch et al., 2001), it is not likely to account for our findings because all the bees in our study were similarly exposed to it. Light is also not likely to be important for the development of circadian rhythms in field colonies in which young bees typically stay in the dark cavity of the nest. Other environmental factors, such as temperature, humidity, and CO2 concentration, which may have differed between the colony and the laboratory, cannot be excluded without additional studies. There is indeed evidence that the rearing temperature during larval or pupal stages has lasting effects on behavior and physiology during the adult stage (Becher et al., 2009; Groh et al., 2004; Tautz et al., 2003). However, we do not know of clear evidence that in honey bees the ambient temperature influences developmental processes during the adult stage.
Our findings extend previous evidence for the pivotal influence of the colony environment on the circadian system of honey bees (Eban-Rothschild and Bloch, 2012). Importantly, our results indicate that different factors in the colony environment influence the expression of circadian rhythms in nurses and the development of the circadian system in young bees. Direct contact with the brood is necessary for the modulation of circadian rhythms in nurse bees (Shemesh et al., 2010) but not for the ontogeny of circadian rhythms in young bees.
Our findings are somewhat counterintuitive because an undeveloped or underdeveloped circadian system could have theoretically benefited young bees that typically tend the brood around the clock. Furthermore, the presence of foragers typically postpones the expression of physiological and behavioral traits associated with foragers in young bees (Robinson, 1992; Bloch, 2010). Our current findings, however, are consistent with the findings of Shemesh et al. (2007) and Meshi and Bloch (2007) that showed an earlier manifestation of circadian rhythms in young bees that were housed inside an observation hive or in the laboratory in small groups with foragers (respectively), compared with young bees housed only with same-age bees outside the hive. Thus, the accelerated development of the circadian system in bees in a colony relative to isolation appears to be independent of the environmental regulation of age-related division of labor. Rather it seems to indicate that young bees develop better in the colony than in isolation in the laboratory.
This interpretation of our findings is consistent with a growing body of evidence showing that the colony environment influences the normal development of young adult honey bees. For example, the volume of the mushroom bodies was reduced in bees that were isolated during their first 8 days compared with normally developing bees (Maleszka et al., 2009). Bees that experienced the colony environment during days 2-6 of adult life also showed improved learning and memory performance relative to sister bees that were individually isolated during the same period (Ichikawa and Sasaki, 2003). Similar to our findings, the influence of the colony environment was evident also in bees that were prevented from direct contact with other bees by a double-mesh enclosure (Ichikawa and Sasaki, 2003). In a broader perspective, these findings for bees fit well with studies showing that social isolation has strong effects on the development of brain and behavior in both vertebrates and invertebrates (e.g., Fone and Porkess, 2008; Hall, 1998; Harlow, 1965; Heisenberg et al., 1995; Scotto Lomassese et al., 2000; Technau, 2007).
Our findings suggest that factors in the colony environment affect the development of the circadian system in young honey bees and that these factors are different from those regulating the switch of nurse bees between activity with and without circadian rhythms. It is still unclear whether the observed differences in the expression of circadian rhythms stem from influences on the development of the circadian network or its coupling to the motor centers controlling locomotor activity. Additional studies are needed also to elucidate the specific environmental factors and the sensory modalities involved in mediating the influence of the colony environment on the development of the circadian system in young adult bees.
Footnotes
Acknowledgements
The authors thank Rafi Nir, Taryn Harpaz, and Yafit Brenner for assistance with the bees. This work was supported by grants from the US-Israel Binational Science Foundation (BSF, 2007-465), the Israel Science Foundation (ISF, grant 606/02), and the German Israel Foundation (GIF, Contract Number I-822-73.1/2004) to G.B.
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.