
Abstract
Some major climatic changes occurred during the Neogene Period, which encompasses the Miocene and Pliocene epochs. The marine flora and fauna either adapted to or became extinct due to changes in climatic conditions and tectonic activity. The Andaman and Nicobar Basin in the northeast Indian Ocean hosts deep-water marine facies with a few shallow-water sequences. The Neogene sediments of the basin archive well-preserved fossil phytoplankton and zooplankton represented by diatoms, radiolarians, silicoflagellates, calcareous nannofossils, including planktonic and benthic foraminifers, etc. The Neogene marine sediments exposed on various islands in the Andaman and Nicobar Group offer a unique and valuable geological archive. Combined with micropalaeontological data obtained from deep-sea sediment cores, these records provide an excellent foundation for comprehensive biostratigraphy and for reconstructing the overall palaeoenvironment and palaeoceanography. The significant events identified by the analysis of microfossils are: the Miocene Climate Optimum (MCO) during the middle Miocene; intensification of the Indian Summer Monsoon (ISM) coupled with upwelling and nutrient-rich conditions during the late Miocene; and low biogenic silica and a cooling event during the early Pliocene. Morphometric analysis of long-ranging calcareous nannofossil species that existed during ~16.4 to ~4.54 Ma reveals that they survived in different environments due to variations in their dimensions. Estimation of radiolarian shell volume reveals a gradual decrease in shell volume with younger age during the late Neogene. Silicoflagellates, another group of phytoplankton, are also useful for estimating relative sea surface palaeotemperature. Datasets generated from fossilised phytoplankton and zooplankton, both from onshore and offshore sequences of the northeast Indian Ocean (Andaman and Nicobar Basin), provide valuable information about the overall Neogene palaeoceanographic scenario.
Keywords
INTRODUCTION
The Neogene Period (23.03-2.58 Ma) witnessed a significant biotic turnover in both marine and terrestrial ecosystems, primarily driven by significant climatic changes. One of the most notable transitions was the shift from a greenhouse climate to an icehouse climate, which fundamentally altered global ecosystems. This biotic turnover also corresponds to the excursion in the global oxygen and carbon isotopes of MI-1 (Bradshaw, 2021). These changes may have occurred due to various factors, namely CO2 concentration in the atmosphere, orbital orientation, oceanic heat transport, and alterations in palaeogeography (Fauquette et al., 2007, and references therein). Climate changes in the Miocene were influenced by oscillations in the magnitudes of Earth’s obliquity and eccentricity. Palaeoceanographic changes in the early and middle Miocene epochs of the Neogene Period were largely influenced by plate tectonics, which ultimately affected Neogene climate and ocean circulation patterns, especially the opening and closing of oceanic gateways (Fauquette et al., 2007; Steinthorsdottir et al., 2021). From the early Miocene onwards, the Indian Ocean connection with the Mediterranean Sea became restricted (Harzhauser et al., 2011). The compartmentalisation of the Tethys Sea into the Mediterranean and Indian Oceans occurred during the early Miocene; however, the exact nature and timing of this dissociation remain poorly understood (Bialik et al., 2019). The reorganisation of water currents on a global scale occurred due to the closure of Meridional mass exchange through the Tethyan Seaway, resulting in upwelling and intensification of the South Asian Monsoon. The late Miocene (8.0-5.33 Ma) has been regarded as a climatically stable period characterised by minor long-term cooling and ice growth episodes (Zachos et al., 2001). The late Miocene Epoch, specifically the Tortonian and Messinian, appears to be a notable period within the Neogene, characterised by the most significant sustained shift from an equitable climate to the modern climatic scenario (Herbert et al., 2016). During the Neogene, the Indian subcontinent, specifically the Andaman and Nicobar Islands, underwent significant tectonic, physiographic, floral, faunal, and oceanographic changes. Some important events, such as the uplift of the Himalayas, the elevation of marine deep-sea sediments in the Andaman-Nicobar Basin, the development of the enormous Bengal Fan, and the disappearance of the Tethys, occurred during the Neogene. The Neogene marine sedimentary rocks of predominantly deep-water origin cover extensive areas across different islands of Andaman and Nicobar Group, and these sediments archive well-preserved fossilised phytoplankton and zooplankton, represented by diatoms, radiolarians, silicoflagellates, calcareous nannofossils, and planktonic as well as benthic foraminifers. Moreover, deep-sea sediment cores drilled as the offshore of the Andaman-Nicobar Basin also provided excellent micropalaeontological data on the Neogene Period. Both the offshore and onshore sediments archive well-preserved marine microfossils those are ideal for dating and correlation (Sharma & Srinivasan, 2007 and the references therein, Chakraborty & Ghosh, 2016, 2021; Chakraborty, Ghosh, McCartney, et al., 2021; Chakraborty, Ghosh, & Saxena, 2021; Chakraborty et al., 2019, 2023; Dey & Ghosh, 2024; Dey et al., 2021, 2022; Roy et al., 2022, 2023, 2025a, 2025b; Saxena & Ghosh, 2024; Saxena et al., 2022). The Messinian sequence and the Tortonian-Messinian transition/boundary in the northeast Indian Ocean have been demarcated from the onshore (Dey & Ghosh, 2024; Saxena & Ghosh, 2024) and offshore (Roy et al., 2025a, 2025b) sequences of the Andaman-Nicobar Basin using multiple microfossil biostratigraphy. In the present study, adaptation and changes in the composition of multiple planktonic assemblages, encompassing various siliceous microfossils, calcareous nannofossils, and planktonic foraminifers, recorded from Neogene sediments of the northeast (NE) Indian Ocean, are discussed in relation to changes in climatic conditions during this period.
GEOLOGICAL AND OCEANOGRAPHIC SETTINGS
The Andaman and Nicobar Islands, situated in the northeastern Indian Ocean, comprise an arcuate chain and are broadly divided into two groups: The Andaman Group and the Nicobar Group. The Andaman Group of Islands is separated from the Nicobar Group by the Ten Degree Channel running along the 10°N latitude (Figure 1A). Due to two major events, that is, the Neogene and prior to Neogene development, the evolution of this Arc System took place (Sharma & Srinivasan, 2007). There is a genetic linkage between the Andaman-Nicobar Arc and the Himalayan chain (Sharma & Srinivasan, 2007 and references therein).
A. Map of northeast Indian Ocean showing the different islands of Andaman and Nicobar Group and drilling site of NGHP-01-17A marked by red square; B. Map Havelock Island in Ritchie’s Archipelago showing the location of the sections marked by red asterisks; C. Map of Neil Island in Ritchie’s Archipelago showing the location of the sections marked by red asterisks; D. Map of Car Nicobar Island in Nicobar Group of Island showing the location of the sections marked by red asterisks (Modified after Chakraborty et al., 2023).
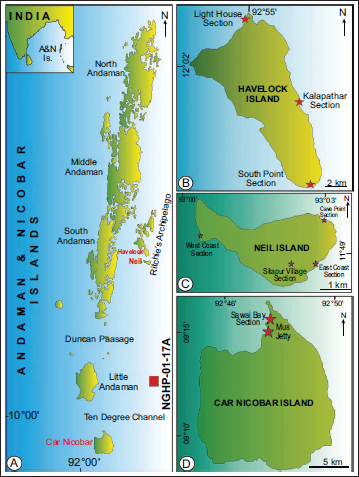
A complex tectonic event accompanied by convergent plate boundaries between the Indian and West Burmese plates was responsible for the formation of the Andaman-Nicobar Basin. The initiation of this basin occurred due to the northward flight of the Indian plate with respect to Asia. Around 59 Ma Ago, a soft collision between the two plates occurred (Klootwijk et al., 1992). According to Singh and Moeremans (2017), the basin was formed by the deposition of 3–3.5 km of Cenozoic sediments that accumulated over the past ~60 Ma.
The Andaman and Nicobar Basin is characterised by thick marine sedimentary deposits ranging from the late Mesozoic to the Quaternary. Two distinct groups of sediments, namely the Andaman Flysch (late Eocene to Oligocene) and the overlying Archipelago Group (Miocene to Pliocene), are present in the basin. The sediments of the Archipelago Group were deposited in an outer-shelf to open-marine settings (Chakraborty & Pal, 2001). In Figure 1A–1D, different islands of the Andaman and Nicobar Group and the drilling site of NGHP-01-17A have been depicted from where the presently analysed datasets were generated.
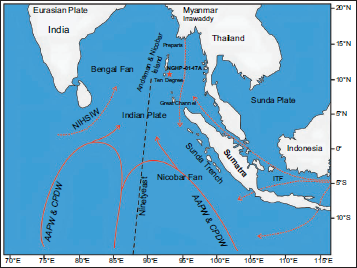
The northeast Indian Ocean is a part of the deep-sea environment of the continental margin (Liu et al., 2021). The sources of intermediate and deep-water masses in the Indian Ocean are the Atlantic and Southern Oceans (Cawthern et al., 2014; Kawagata et al., 2006). The intermediate waters are constituted by the mixture of Antarctic Bottom Water (AABW) and North Atlantic Deep Water (NADW), that is, Circumpolar Deep Water (CPDW) (Kawagata et al., 2006; Vincent, 1974), whereas, the sources of deep waters are northward-flowing cold and substantially oxidised AABW (Figure 2). The influence of low salinity surface water mass and North Indian High Salinity Intermediate Water (NIHSIW) contributes to the upper part (i.e., ~1000 metres) of the water column in the Bay of Bengal (Figure 2). These sources of intermediate and surface water masses in the Bay of Bengal are mainly controlled by the intensification of the summer and winter monsoons (Kawagata et al., 2006; Naqvi et al., 1994; Varkey et al., 1996). Freshwater runoff and the intermixing of water masses originating from the Indian and Pacific Oceans are the sources of deep, intermediate and surface waters in the Andaman Sea (Amiruddin et al., 2011; Ibrahim & Yanagai, 2006).
Part of the northeast Indian Ocean showing ocean circulation pattern (Modified after Cawthern et al., 2014; Roy et al., 2023).
METHODOLOGY
Micropalaeontological datasets generated on multiple siliceous microfossils (radiolarian, diatoms and silicoflagellates) and calcareous nannofossils as well as planktonic foraminifera from different sections (Figure 3) of Havelock Island [(a) Near Kalapathar beach, 12°00′12.51′′ N: 93°00′24.02′′E; (b) South Point, 11°52′ 55.39′′ N: 93°02′ 20.20′′ E and (c) Near Light House, 12°02′57.0012′′ N: 92°57′ 53.712′′ E], Neil Island [(a) Cave Point, 11°50′31.16′′ N: 93°03′53.76′′ E; (b) East Coast, 11°49′29.30′′ N: 93°03′50.94′′ E; (c) Near Sitapur Village, 11°49′24.9′′ N: 93°03′34.3′′ E and (d) West coast near the Natural Bridge, 11°49′48.80′′ N: 93°00′54.50′′ E] of Ritchie’s Archipelago and Car Nicobar Island [(a) Sawai Bay ‘A’, 09°14′ 24.4′′ N: 92°46′ 40.6′′ E and (b) Sawai Bay ‘B’, 09°14′04′′ N: 92°46′34.1′′ E] of Andaman and Nicobar Group of Islands (Chakraborty et al., 2023 and the references therein; Saxena & Ghosh, 2024; Dey & Ghosh, 2024).
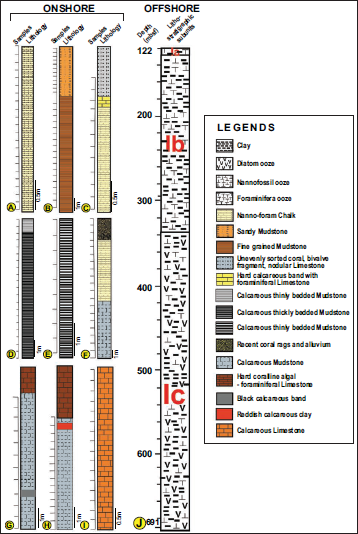
A–C. Lithological features of late early to Middle Miocene sections on Havelock Island: A. Kalapathar Section near beach; B. South Point Section; C. Laccam Point Section near Light House; D–F. Lithological features of late Miocene sections on Neil Island: D. Cave Point Section; E. East Coast Section; F. Sitapur Village Section; G-H. Lithological features of early Pliocene Sections on Car Nicobar Island: G. Sawai Bay ‘A’ Section; H. Sawai Bay ‘B’ Section; I. Lithological features of Plio-Pleistocene Section on the west coast of Neil Neil Island; J. Columnar section of the NGHP-01-17A core showing the lithological characteristics (Modified after Flores et al., 2014).
Roy et al. (2022, 2023, 2025a, 2025b) generated extensive high-resolution micropalaeontological data on diatoms, silicoflagellates, radiolarians and calcareous nannofossils from the offshore of the Andaman and Nicobar Basin drilling site NGHP-01-17A of NGHP Expedition 1. The drilling site is located east of Little Andaman Island near the Andaman back-arc basin (10° 45′ 11′′N, 93° 6′ 44′′E; Figures 1A and 3).
To understand the adaptation and successful survival of the phytoplankton (calcareous nannofossils) and zooplankton (radiolarians) under varying climatic conditions in the northeast Indian Ocean, ranging in age from approximately 16.4 Ma (Miocene) to ~4.19 Ma (Pliocene), Ghosh et al. (2025) employed various mathematical parameters. Morphometric analysis of some calcareous nannofossil species and the estimation of shell volumes of some radiolarian species were considered in this study.
To evaluate the influence of Neogene climate fluctuations on the planktonic community, the previously mentioned datasets were analysed.
NEOGENE PLANKTON ASSEMBLAGES FROM THE ONSHORE AND OFFSHORE OF THE ANDAMAN AND NICOBAR BASIN
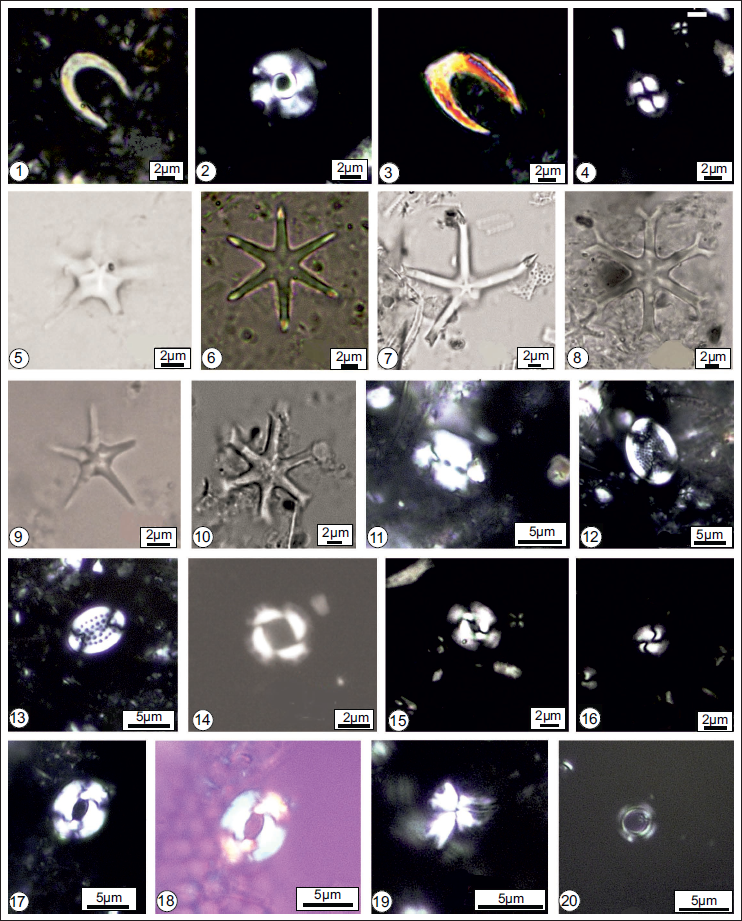
A wide array of microfossils, represented by the remains of phytoplankton and zooplankton, from both onshore and offshore sequences of the Andaman and Nicobar Basin, indicates that the sediments range in age from the middle Miocene to the Plio-Pleistocene, based on index/marker species. Light microscopic images of some significant taxa of diatoms (Plate 1, Figures 1–28), calcareous nannofossils (Plate 2, Figures 1–20), radiolarians and silicoflagellates (Plate 3, Figures 1–24), along with SEM images of planktonic foraminifera and silicoflagellates (Plate 4, Figures 1–9), are illustrated. The figured slides (BSIP Slide Nos. 16776, 16780-16781, 16784-16789, 16781-16793, 16800, 16803-16805) and SEM stubs (BSIP Sample Nos. 9746/9-19) are housed at Birbal Sahni Institute of Palaeosciences, Lucknow.
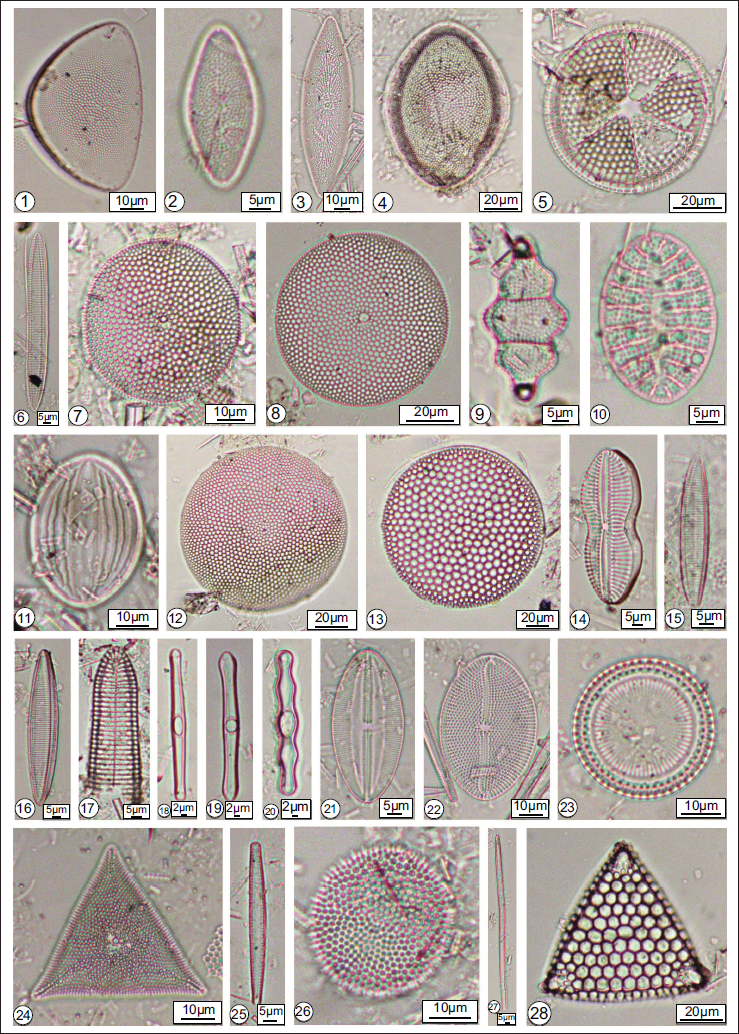
Some significant Neogene diatoms from the onshore and offshore sequences of Andaman and Nicobar Basin: 1. Actinocyclus cuneiformis; 2. Actinocyclus ellipticus; 3. Actinocyclus ellipticus f. Lanceolatus; 4. Actinocyclus ellipticus var. javanica; 5. Actinoptychussenarius; 6. Alveus marinus; 7. Azpeitia nodulifera; 8. Azpeitia nodulifera f. Cyclopus; 9. Biddulphia tridens, 10. Campyloneis grevellei; 11. Cocconeis pellucid; 12. Coscinodiscus asteromphalus; 13. Coscinodiscus radiates; 14. Diploneis crabro; 15. Fragilariopsis miocenica; 16. Fragilariopsis reinholdii; 17. Gephyria media; 18. Grammatophora marina; 19. Grammatophora oceanic; 20. Grammatophora undulate; 21. Lyrella spectabilis; 22. Mastogloia splendid; 23. P. sulcata; 24. Pseudotriceratium punctatum; 25. Thalassionema nitzschoides; 26. Thalassiosira grunowii; 27. T. longissima; 28. Triceratium favus.

Some stratigraphically and paleoecologically important Neogene calcareous nannofossils from the onshore and offshore sequences of Andaman and Nicobar Basin: 1. Amaurolithus delicatus; 2. Calcidiscus macintyrei; 3. Ceratolithus cristatus; 4. Coccolithus pelagicus; 5. Discoaster berggrenii; 6. Discoasterbrouweri; 7. Discoaster hamatus; 8. Discoaster variabilis; 9. Discoasterquinqueramus; 10. Discoaster surculus; 11. Helicosphaera carteri; 12. Pontosphaera discopora; 13. Pontosphaera multipora; 14. Pseudoemiliania lacunosa; 15. Reticulofenestra haqii; 16. Reticulofenestra minuta; 17–18. Reticulofenestra pseudoumbilicus; 19. Spenolithus moriformis; 20. Umbilicosphaera rotula.
Some significant Neogene radiolarians and silicoflagellates from the onshore and offshore sequences of the Andaman and Nicobar Basin. Radiolaria 1–19: 1. Acrobotrys cribosa; 2. Acrobotrys disolenia; 3. Acrobotrys tritubus; 4. Anthocyrtidium zanguebaricum; 5. Botryocyrtis scutum; 6. Botryostrobus auritus-australis; 7. Cornutella profunda; 8-9. Diartus hughesi; 10. Didymocyrtis antepenultima; 11. Didymocyrtis avita; 12. Didymocyrtis penultima; 13. Lithopera bacca; 14. Phormostichoartus doliolum; 15. Siphocampe lineata; 16. Siphocampe lineata; 17. Stylodictya tenuispina; 18. Stylodictya validispina; 19. Tetrapyle octacantha. Silicoflagellates 20–24: 20. Bachmannocena elliptica; 21. Dictyocha calida; 22. Dictyocha extensa; 23. Dictyocha fibula; 24. Stephanocha speculum (5-sided).
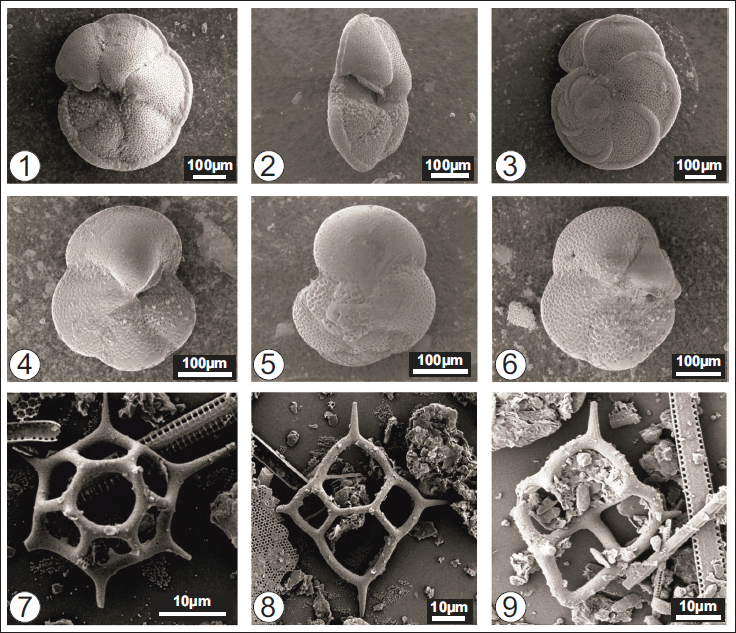
SEM images of stratigraphically important Neogene planktonic foraminifers and silicoflagellates from the onshore and offshore sequences of the Andaman and Nicobar Basin. Planktonic foraminifera 1–6: 1–3. Globorotalia menardii; 4–6. Globorotalia margaritae. Silcoflagettae 7–9: 7. Stephanocha speculum (6-sided); 8. Dictyocha fibula; 9. Dictyocha extensa.
Burdigalian-Langhian plankton assemblages from Havelock Island
Road cutting section near Kalapathar beach
The commonly occurring diatom taxa recorded from this section near Kalapathar beach (12°00′12.51′′ N: 93°00′24.02′′E) are Actinocyclus ingens, Cestodiscus peplum, Coscinodiscus radiatus, Grammatophora angulosa, G. longissima, Paralia sulcata, Rossiella paleacea, Thalassionema nitzschioides, Thalassiothrix longissima, Xanthiopyxis oblonga, etc. Owing to the presence of index diatom taxon C. peplum from the section near Kalapathar beach (12°00′12.51′′ N: 93°00′24.02′′E) of Havelock Island, Chakraborty et al. (2019) and Saxena et al. (2021) assigned it to the C. peplum Zone of Barron (1985a), ranging in age from late early to early middle Miocene. The age was later calibrated by Lazarus et al. (2014) to 16.4-14.1 Ma. The commonly occurring diatom taxa are listed in Supplementary Table 1.
Owing to the presence of the Base (B) of index radiolarian taxon Calocycletta costata and absence of Dorcadospyris alata, Dey et al. (2022) assigned this section to the C. costata Zone, that is, RN4 Zone of Sanfilippo and Nigrini (1998). The significant radiolarian species reported from this section are provided in Supplementary Table 1.
This section is characterised by the dominance of cosmopolitan calcareous nannofossil taxa, reflecting a well-developed open-marine environment. Based on calcareous nannofossils, Chakraborty, Ghosh, and Saxena (2021) assigned this section to the Helicosphaera ampliaperta Zone, that is, NN4 Zone of Martini (1971), corresponding to the CNM6-CNM7 zones of Backman et al. (2012). For the frequently occurring calcareous nannofossil species, please see Supplementary Table 1.
South Point Section
The South Point Section of Havelock Island (11°52′ 55.39′′ N: 93°02′ 20.20′′ E) is also assignable to the C. peplum Zone of Barron (1985a) based on the presence of the diatom species C. peplum (Saxena et al., 2021).
Based on radiolarian events, the lower part of this section was assigned to the C. costata Zone, that is, RN4 Zone of Sanfilippo and Nigrini (1998) and the upper part was designated (Dey et al., 2022) as RN5 Zone, that is, D. alata Zone of Sanfilippo and Nigrini (1998).
Based on calcareous nannfossil events, that is, Top common (Tc) of Discoaster deflandrei recorded from the base of the South Point Section, it was assigned to NN4 Zone of Martini (1971), corresponding to CNM6 Zone of Backman et al. (2012) by Chakraborty, Ghosh, and Saxena (2021).
Occurrence of significant species of diatoms, radiolarians and calcareous nannofossils South Point Section are provided in Supplementary Table 1.
Section near the Light House
Diatom assemblage recorded from this section (12°02′57.0012′′ N: 92°57′ 53.712′′ E) near Light House of the Havelock is dominated by Actinocyclus senarius, Aulacodiscus patulus, Biddulphia tridens, Coscinodisus marginatus, R. paleacea, etc. (Saxena et al., 2021).
Using radiolarian biostratigraphy, Dey et al. (2022) assigned this section to the RN5 Zone, that is, the D. alata Zone of Sanfilippo and Nigrini (1998), owing to the presence of Base (B) of D. alata.
Eighteen species of calcareous nannofossils belonging to nine genera were reported by Chakraborty, Ghosh, and Saxena (2021) from this section near the Light House of Havelock Island. The calcareous nannofossil assemblage is assignable to the NN4 Zone of Martini (1971) and is comparable to the CNM6 Zone of Backman et al. (2012).
A list of the biostratigraphically significant diatom, radiolaria and calcareous nannofossil taxa is given in Supplementary Table 1.
Tortonian-Messinian plankton assemblages from Neil Island
Cave Point Section
The diatom assemblage recorded from the Cave Point Section (11°50′31.16′′ N: 93°03′53.76′′ E) of Neil Island is represented by 82 diatom taxa, and amongst these, 29 taxa are planktonic, and 53 taxa are benthic (Chakraborty & Ghosh, 2016). The significant taxa include Actinocyclus ellipticus, Azpeitia nodulifera, T. nitzschioides, etc.
The base of the section is defined by the radiolarian events - Top (T) of Diartus hughesi, and the secondary datum is demarcated by the Base (B) of Didymocyrtis penultima (Chakraborty & Ghosh, 2015, 2016) and Stichocorys delmontensis. Owing to the presence of the above-mentioned radiolarian events, this section was assigned to the RN8 and RN9 zones of Sanfilippo and Nigrini (1998) by Chakraborty and Ghosh (2015, 2016).
The calcareous nannofossil assemblage of this section is dominated by Helicosphaera carteri, small reticulofenestrids (i.e., Reticulofenestra haqii and R. minuta) and Spenolithus moriformis, with commonly occurring species Coccolithus leptoporus, Discoaster quinqueramus, Helicosphaera sellii, Spenolithus abies and Umbilicosphaera rotula. This section is assignable to D. quinqueramus Zone (NN11) of Martini (1971). It can be correlated to CNM16 of Backman et al. (2012) owing to the presence of marker taxon D. quinqueramus (Chakraborty, Ghosh, & Saxena, 2021).
Significant taxa of diatoms, radiolarians and calcareous nannofossils recorded from the Cave Point Section are included in Supplementary Table 1.
East Coast Section
Assemblages of diatoms, radiolarians, and calcareous nannofossils are similar to those recorded from the East Coast Section of Neil Island (11°49′29.30′′ N: 93°03′50.94′′ E). Amongst the diatoms, A. ellipticus, A. nodulifera, C. radiatus, T. nitzschioides, and T. longissima are the most dominant forms.
The radiolarian events are marked by the Base (B) and top (T) of S. delmontensis, the Base (B) of S. peregina and D. penultima alongside the top (T) of D. hughesi (Chakraborty et al., 2023). Based on these events, this section is assigned to the RN8 and RN9 zones of Sanfilippo and Nigrini (1998).
In having the presence of index calcareous nannofossil species Discoaster quinqueramus throughout this section, it is also assignable to NN11 Zone of Martini (1971) that corresponds to CNM16 Zone of Backman et al. (2012). Reticulofenestra minuta and R. haqii are the most dominant species, and Sphenolithus moriformis is the subdominant species (Chakraborty, Ghosh, & Saxena, 2021).
Significant taxa of diatoms, radiolarians and calcareous nannofossils recorded from the East Coast Section are listed in Supplementary Table 1.
Section near Sitapur Village
The section near Sitapur Village (11°49′24.9′′ N: 93°03′34.3′′ E) was assigned by Chakraborty et al. (2023) to Nitzschia miocenica Zone of diatoms designated by Barron (1985b). According to Barron (1985b), the Base (B) and Top (T) of Fragilariopsis miocenica (= N. miocenica) are at 7.39 Ma and 5.7 Ma, respectively.
The radiolarian event is marked by the evolutionary transition (ET) from Didymocyrtis antepenultima to D. penultima, and the base of this section is assignable to the RN8 Zone of Sanfilippo and Nigrini (1998). The ET of S. delmontensis to Stichocorys peregrina has also been recognised in this section (Chakraborty et al., 2023). Accordingly, the upper part of the section can be assigned to the RN9 Zone of Sanfilippo and Nigrini (1998).
The same samples of this section also yielded well-preserved silicoflagellates (Saxena & Ghosh, 2024). The silicoflagellate assemblage is assignable to the Dictyocha extensa Partial Range Zone (McCartney et al., 1995; Roy et al., 2023).
Based on the occurrence of index calcareous nannofossils, namely Amaurolithus primus, D. quinqueramus, D. berggrenii in this section, Chakraborty et al. (2023) assigned it to NN11a-NN11b subzones of Martini and Müller (1986), corresponding to CNM16-CNM20 zones of Backman et al. (2012).
Significant taxa of diatoms, radiolarians, silicoflagellates and calcareous nannofossils recorded from the Sitapur Village Section are summarised in Supplementary Table 1.
In addition to the above-mentioned silicified microfossils and calcareous nannofossils, the planktonic foraminiferal assemblage of this section comprises the species belonging to the genera Dentoglobigerina, Globigerinoides, Globoconella, Globorotalia, Globorotaloides, Globoturborotalita, Neogloboquadrina, Orbulina, Sphaeroidinella and Trilobatus (Dey & Ghosh, 2024). The presence of index taxa Globorotalia menardii and Globorotalia margaritae signifies the Tortonian-Messinian transition. Following the planktonic foraminiferal zonation of Berggren et al. (1995), the base of this section of Neil Island is assignable to M13b (Globorotalia pleistotumida/ G. lenguaensis Concurrent Range Zone), and the samples above 50 cm from the base to the top can be assigned to M14 (Globigerinoides extremus Partial Range Zone) zones.
Thus, the age interpretation of the Sitapur Village Section on Neil Island, based on calcareous nannofossil biostratigraphy, is also consistent with the diatom, radiolarian, silicoflagellate and planktonic foraminiferal biostratigraphic records. Based on the index calcareous nannofossils, the age for the base of the section has been calibrated as younger than 8.3 Ma but older than 7.39 Ma, and the top as younger than 5.53 Ma but older than 5.4 Ma, following the age connotation of Backman et al. (2012) and Raffi et al. (2020). Raffi et al. (2020) defined the Tortonian-Messinian boundary at 7.25 Ma. Obviously, the Tortonian-Messinian transition/boundary exists within this section of Neil Island.
Tortonian-Messinian-Gelasian plankton assemblages from the offshore of the Andaman and Nicobar Basin
Tortonian diatoms and silicoflagellates, Tortonian-Messinian radiolarians and Tortonian-Gelasian calcareous nannofossils have been recorded by Roy et al. (2022, 2023, 2025a, 2025b) from the sediment core of NGHP-01-17A (10° 45′ 11′′N, 93° 6′ 44′′ E). The entire recovered core drilled during NGHP Expedition-01 was designated as a single unit, that is, Unit I; however, based on changes in carbonate and biogenic silica content, it was further subdivided into Ia, Ib, and Ic (Collett et al., 2012).
Roy et al. (2022) proposed three diatom bio-zonations from subunit Ic. The diatom bio-zonations are summarised in Supplementary Table 2. Roy et al. (2023) analysed the samples from subunit Ic of the core (684.09–369.62 mbsf) for silicoflagellate content and the taxa belonging to the genera Bachmannocena, Dictyocha, Distephanopsis, Paramesocena, and Stephanocha. For the first time, from the Tortonian of the northeast Indian Ocean, they proposed two Partial Range zones, namely Dictyocha varia and D. extensa and one Subzone, that is, Bachmannocena elliptica. In their study, the cumulative abundance of all taxa belonging to the genera Dictyocha and Stephanocha revealed overwhelming dominance of Dictyocha over Stephanocha.
From the Tortonian-Messinian sequence (684.09-220.37 mbsf) of the sediment core, Roy et al. (2025a) recognised four radiolarian events or zones based on index radiolarian species. The recognised radiolarian events are provided in Supplementary Table 2.
In a recent study from the NGHP-01-17A core, Roy et al. (2025b) recorded. Tortonian to Gelasian calcareous nannofossil assemblage assignable to NN9-NN19 zones of Martini (1971), corresponding to CNM13-CNPL7 zones of Backman et al. (2012), based on marker/index taxa. The significant calcareous nannofossil events that were recognised by Roy et al. (2025b) are provided in Supplementary Table 2.
Zanclean plankton assemblages from Car Nicobar
Sawai Bay ‘B’ Section
Chakraborty, Ghosh, McCartney, et al. (2021) and Chakraborty, Ghosh, and Saxena (2021) recorded siliceous microfossils (diatoms, radiolarians and silicoflagellates) and calcareous nannofossils from Sawai Bay ‘B’ Section (09°14′04′′ N: 92°46′34.1′′ E) on Car Nicobar Island. The section has been dated to the Zanclean (Chakraborty, Ghosh, McCartney, et al., 2021; Chakraborty, Ghosh, & Saxena, 2021) based on index calcareous nannofossils.
Moderately preserved diatom taxa in the Sawai Bay ‘B’ are represented by the taxa belonging to the genera Actinocyclu, Azpeitia, Coscinodiscus, Grammatophora, Triceratium, etc. and amongst these Triceratium favus is the most dominant species. Chakraborty, Ghosh, McCartney, et al. (2021) commented that, owing to low diversity, scanty and sporadic presence of diatoms, it was difficult to correlate the diatom assemblage with any standard diatom bio-zone.
The commonly occurring radiolarian taxa in this section are: Didymocyrtis avita, Euchitonia spp., Siphocampe lineata, S. peregrina, Semantis spp., Stylochlamydium sp., etc. (Chakraborty, Ghosh, McCartney, et al., 2021). Presence of S. peregrina throughout the section is indicative of S. peregrina Interval Zone, that is, the RN9 Zone of Sanfilippo and Nigrini (1998). The occurrence of D. avita in the samples reconfirms its placement in the RN9 Zone (Kamikuri et al., 2009).
In the Sawai Bay ‘B’ Section, the most common species of silicoflagellates is Dictyocha varia, and the other sporadically present taxa are D. extensa, B. elliptica, and Dictyocha cf. fibula subsp. fibula and Dictyocha fibula subsp. mutabilis (Chakraborty, Ghosh, McCartney, et al., 2021). Sporadic presence of silicoflagellates in this section prevented its assignment to any silicoflagellate biostratigraphic zone.
Sawai Bay ‘A’ Section
This section lacks any siliceous microfossils. Based on calcareous nannofossils, the Sawai Bay ‘A’ Section is assignable to NN12-NN13 zones of Martini (1971), corresponding to CNPL1-CNPL2 zones of Backman et al. (2012), owing to the occurrence of C. armatus and absence of C. cristatus (Chakraborty, Ghosh, McCartney, et al., 2021; Chakraborty, Ghosh, & Saxena, 2021; Chakraborty et al., 2023). The presence of Ceratolithus acutus in all samples indicates a Zanclean age. Based on the calcareous nannofossil index, the base of the section was dated to ~5.33 Ma, and the upper part to ~5.08 Ma.
Piacenzian – Gelasian plankton assemblage from Neil Island
West coast near the Natural Bridge
From this section on Niel Island (11°49′48.80′′ N: 93°00′54.50′′ E), the Piacenzian–Gelasian foraminiferal taxa belonging to the genera Dentoglobigerina, Globigerina, Globigerinoides, Globoconella, Globorotalia, Globorotaloides, Globoturborotalita, Neogloboquadrina, Orbulina, and Trilobatus have been recorded (Dey et al., 2021). Based on index species, namely Dentoglobigerina altispira, Globorotalia miocenica and Globoturborotalita woodi, the section has been dated as Piacenzian–Gelasian and is comparable to N20/N21 zones of Kennett and Srinivasan (1983) and PL4 to PL6 zones of Berggren (1995). Dey et al. (2021) also suggested the Plio-Pleistocene boundary in this section of Neil Island.
Morphometric analysis on calcareous nannofossils and estimation of the shell volumes of radiolarians from the Neogene sediments of Andaman and Nicobar Basin
Size variations of calcareous nannofossils are controlled by various factors, namely sea surface temperature, monsoonal variation, productivity/nutrient availability and sea level fluctuations (Young, 1998). To assess these features, Ghosh et al. (2025) carried out morphometric analysis on selected species of calcareous nannofossils, namely, Calcidiscus leptoporus, Coccolithus pelagicus, H. carteri, R. haqii, R. minuta, Sphenolithus abies and U. rotula that continued their existence during the middle Miocene (~16.4 Ma) to early Pliocene (~4.54M). Changes in their dimensions were noted at different time intervals to analyse the size variations of each calcareous nannofossil species across time slices.
The shell volume of radiolarians plays an important role in their adaptation and diversity. Shell volume estimation of the six species of radiolarians was conducted by Ghosh et al. (2025). They documented their presence from the Toronian (~9.46 Ma) to the Messinian (~5.33 Ma) in offshore sediments from the NGHP-01-17A core. Only five selected Nassellarian radiolarian species, that is, Artostrobus annulatus, Cornutella profunda, Phormostichoartus doliolum, Siphocampe lineate, and S. delmontensis, and one spumellarian species, S. delmontensis, continued their presence from Tortonian to Messinian. Using the mathematical formula proposed by Lazarus et al. (2009), the shell volumes were measured by microscopic examination. For the hollow spheres, the silica volume was measured using two different formulae (Ghosh et al., 2025).
NEOGENE CLIMATE AND RESPONSE OF PLANKTON
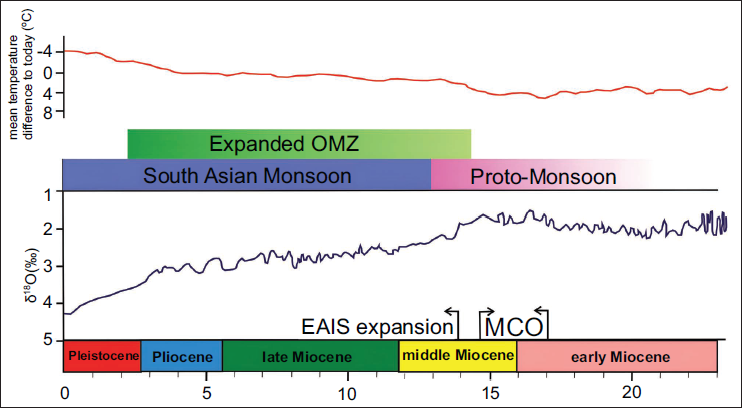
During the Neogene Period, within the Miocene Epoch, an exceptionally warm climatic phase known as the Miocene Climatic Optimum (MCO) occurred. This global phenomenon occurred during the middle Miocene, approximately between 17 and 14.7 million years ago (Herold et al., 2008; Holbourn et al., 2005). The substantial increase in the global δ13C value estimated during the early Miocene is ~19 to 16 Ma (Wright et al., 1992). The middle Miocene was ~6°C warmer than present-day at middle latitudes (Herold et al., 2008). According to Woodruff and Savin (1989), the Indian Ocean was also a likely source of high δ13C water in the early and middle Miocene. A progressively cooler climate persisted after the MCO event. An isotope record, based on 13C/12C ratios of organic constituents, reveals that a global cooling trend occurred in the late Miocene, with a drop of ~3.1 to 4.3°C in marine water temperature in the photic zone (Schoell et al., 1994). Based on a distinctive response to astronomical forcing, Westerhold et al. (2020) opined that in the early Tortonian, global temperatures were ~4 to 11°C higher than present-day, and during the late Tortonian, global temperatures were almost 3°C warmer than present-day (Figure 4). Stratigraphical, geophysical, and geochemical data collected from IODP Expedition 359 in the Maldives archipelago (Betzler et al., 2017) indicate that the South Asian Monsoon intensified during the late Miocene (Tortonian), coinciding with the expansion of the Oxygen Minimum Zone (OMZ) (Figure 4). Three significant events of global significance in the Pliocene, namely (a) the formation of Arctic ice sheets, (b) the emergence of a land bridge between South America and North America, and (c) the reorganisation of Indonesian Throughflow (ITF). According to Auer et al. (2019), these are the key elements of global thermohaline circulation. All these palaeoceanographic and tectonic events are responsible for the climatic shifts during the Neogene Period. The fossilised phytoplankton and zooplankton recorded from the Andaman-Nicobar Basin in the northeast Indian Ocean reveal how plankton communities adapted and evolved in response to different climatic events during the Neogene.
The South Asian Monsoon intensification event during the late Miocene and Oxygen Minimum Zone (OMZ) based on the Oxygen isotopic record (EAIS East Antarctic Ice Sheet Expansion and MCO: Miocene Climate Optimum) and mean global temperatures during the Neogene Period for the low and mid-latitudes (modified after Betzler et al., 2017; Westerhold et al., 2020).
The Burdigalian-Langhian assemblages of diatoms, radiolarians, and calcareous nannofossils recorded from Havelock Island are dominated by warm-water taxa (Chakraborty et al., 2019, 2023). The age of the sections, Burdigalian to Langhian on Havelock Island derived from calcareous nannofossils, radiolarians, and diatoms, is also coeval with the MCO. The calcareous nannofossil species that persisted from Burdigalian to Zanclean also adapted to the extreme event of MCO through size variation (Ghosh et al., 2025). Diatoms and radiolarians recorded from Tortonian (both from the onshore, i.e., Neil Island and offshore, i.e., NGHP-17A) of the Andaman and Nicobar Basin (Chakraborty & Ghosh, 2016; Roy et al., 2022, 2025a) indicate extensive ocean upwelling. The presence of cold-water diatom taxa, namely Actinoptychus senarius and Coscinodiscus marginatus, along with the dominance of T. nitzschioides, indicates monsoon intensification and ocean upwelling during the late Miocene. Planktonic diatoms were more abundant than benthic diatoms, and planktonic blooms were obviously due to increased nutrient-rich conditions. Chakraborty, Ghosh, and Saxena (2021) and Roy et al. (2025b) estimated that sedimentation rates during the Tortonian were very high, and an increase in terrigenous input into the Andaman Sea during the Tortonian is a plausible factor. High nutrient-rich conditions are also evident from the Tortonian calcareous nannofossil assemblages from the onshore (Chakraborty, Ghosh, & Saxena, 2021) and offshore (Roy et al., 2025b) of the Andaman and Nicobar Basin. Using the ratio of silicoflagellate genera Dictyocha and Stephanocha, the overall sea surface palaeotemperature was determined, and Roy et al. (2023) opined that an overall warm condition prevailed during 9.86-7.39 Ma. This trend of nutrient-rich ocean upwelling conditions continued till the Messinian, and that is supported by the diatom, radiolarian, silicoflagellate, calcareous nannofossil and planktonic foraminiferal assemblages recorded from the Tortonian-Messinian Section of Neil Island (Chakraborty et al., 2023; Dey & Ghosh, 2024; Saxena & Ghosh, 2024). This can be linked to the intensification of the Indian Summer Monsoon and biogenic bloom during the Tortonian-Messinian. Abundance and diversity of siliceous microfossils dramatically decreased in the Zanclean sediments of Car Nicobar Island in comparison to the Tortonian and Messinian. Possible dissolution of the siliceous microfossils was envisaged by Chakraborty, Ghosh, McCartney, et al. (2021). The sedimentation rate also substantially decreased during the Zanclean, as revealed by the study of calcareous nannofossils from both onshore (Chakraborty, Ghosh, & Saxena, 2021) and offshore (Roy et al., 2025b) sequences of the Andaman-Nicobar Basin. Evidence of early Pliocene low primary productivity and low bSi accumulation from the Zanclean of Car Nicobar Island was suggested by Chakraborty and Ghosh (2021), possibly linked to the impact of the ITF constriction (Chakraborty & Ghosh, 2021).
The Plio-Pleistocene planktonic foraminiferal assemblage recorded by Dey et al. (2021) from the West Coast Section is dominated by Globigerina bulloides and Neogloboquadrina pachyderma, which is indicative of intense upwelling. The warm-water species Neogloboquadrina dutertrei is also recognised as an upwelling responder and inhabits regions of high productivity. During the late Pliocene to early Pleistocene, the preponderance of N. pachyderma implies a cooling event. However, the two taxa, N. dutertrei and N. pachyderma, alternated in dominance during the late Pliocene to early Pleistocene. This may be due to alternation between relatively cooler and warmer climatic conditions (Dey et al., 2021). Planktonic foraminiferal evidence indicates that the initiation of cooler climatic conditions and an upwelling event occurred during the late Pliocene and intensified in the early Pleistocene.
Morphometric analysis of calcareous nannofossil species reveals that some species that persisted from Burdigalian to Zanclean (~16.4 Ma to ~4.54 Ma) adapted to the extreme event of MCO and also successfully survived during the Tortonian-Messinian upwelling condition through size variation (Ghosh et al., 2025). The shell volume estimation of radiolarians using mathematical tools by Ghosh et al. (2025) revealed that there was a gradual decrease in radiolarian shell volume during ~9.46 Ma to ~4.19 Ma, which suggests competition for dissolved silica, a shared nutrient for both radiolarians and diatoms, resulted in biological co-evolution between radiolarians and marine diatoms and became more pronounced in the Neogene.
CONCLUSIONS
The foregoing analysis clearly indicates that both phytoplankton (diatoms, calcareous nannofossils and silicoflagellates) and zooplankton (radiolarians and planktonic foraminifera) exhibited distinct and coordinated significant climatic fluctuations. The major climatic events, namely the MCO during the Burdigalian-Langhian, ocean upwelling and intensification of the summer monsoon during the Tortonian-Messinian, low primary productivity during the Zanclean, and progressively cooler climatic conditions during the Piacenzian–Gelasian, have been interpreted based on the different plankton assemblages. The biogenic bloom and upwelling conditions during the late Miocene are also supported by the high sedimentation rate and low primary productivity during the Plio-Pleistocene may be linked to the sharp decline in sedimentation rate alongside the constriction of the ITF. Low primary productivity during the Plio-Pleistocene is evident from the paucity or complete absence of siliceous microfossils (diatoms, radiolarians and silicoflagellates) both in the onshore and offshore sediments. Most of the climatic events recognised from the studies of Andaman and Nicobar Basin, located in the northeast Indian Ocean, are correlatable to the global events that occurred during the Neogene Period.
Footnotes
Acknowledgements
The authors are indebted to the Director, Birbal Sahni Institute of Palaeosciences, Lucknow, India, for his kind permission (Reference: BSIP/RDCC/Publication no. 54/2025-26) to carry out this work and for providing the necessary laboratory facilities. A.K.G. is grateful to CSIR for the award of Emeritus Scientist [CSIR-HRDG No. 21(1143)/22/EMR-II]. L.R. is grateful to the Head, Department of Earth Sciences, Indian Institute of Technology Bombay, Powai, Mumbai – 400076, Maharashtra, for the award of Post-Doctoral Fellowship. L.R. is also thankful to DST for the DST-INSPIRE Faculty Fellowship (IFA24-EAS 133) and to the Head, Department of Earth Sciences, Indian Institute of Engineering Science and Technology (IIEST), Shibpur. Thanks are due to Dr (Ms) Stuti Saxena and Dr (Mrs) Rikee Dey for their help during the field visit to some of the islands of the Andaman and Nicobar Group. A.C. is thankful to the Department of Geology, Faculty of Science, University Malaya. Sincere thanks are due to the two anonymous reviewers for their valuable comments and especially to the Guest Editor, Prof. Ashutosh K. Singh for his invitation to submit the article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This work was carried out with the support of CSIR Emeritus Scientist Scheme [CSIR-HRDG No. 21(1143)/22/EMR-II], Post-Doctoral Fellowship of IIT Bombay, DST-INSPIRE Faculty Fellowship (IFA24-EAS 133).
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.