
Abstract
Bioerosional structures are well preserved in the Miocene Boka Bil Formation, exposed in the Kaiphundai section, Thangjing Hill Range of Manipur, India. Eleven ichnospecies representing the Trypanites Ichnofacies, viz., Entobia cateniformis, E. ovula, Gastrochaenolites anauchen, G. cluniformis, G. lapidicus, G. orbicularis, G. ornatus, G. torpedo, Maeandropolydora sulcans, Trypanites solitarius and T. weisei, are recorded from the two sections of the Boka Bil Formation. Two distinctive ichnoassemblages, Entobia-dominated and Gastrochaenolites-dominated are recognised. The Entobia-dominated assemblage suggests relatively deeper marine conditions in the deeper part of the back reef or fore reef while the Gastrochaenolites-dominated assemblage points shallower marine conditions, underwater depth of a few metres in the back reef to patch-reef zone of a back-reef lagoon.
Introduction
During the last three decades, the four valid substrate-controlled ichnofacies, namely Glossifungites, Gnathichnus, Teredolites and Trypanites, have received considerable attention as, in addition to their palaeoenvironmental implications, they delineate sequence stratigraphic surfaces. The Trypanites Ichnofacies is characterised by: (a) dominance of deep-tier borings; (b) dominance of sharp-walled, unlined, passively filled, vertical to sub-vertical, cylindrical, simple, tear-vase or U-shaped dwelling structures of suspension feeders or passive predators; (c) presence of branched and multi-chambered borings produced by suspension feeders; (d) low to moderate ichnodiversity; (e) occasional presence of micro borings and (f) high abundance (Buatois & Mangano, 2011). The Trypanites Ichnofacies is typical of cemented, fully lithified substrates, such as sediment-starved hardgrounds, cliffs, beach rocks and reefs, indicating a long period of bioerosion, rocky shore, omission surface, old wave-cut platform and low rate of sedimentation or non-deposition (Bromley & Asgaard, 1993b; Buatois & Mangano, 2011; Frey & Pemberton, 1984; Pemberton et al., 1980, 2001). Rocky shores exposed in the shallow marine environments offer excellent conditions for colonization by marine bioeroding organisms as these marine environments are unique where wave-cut marine erosion platforms are formed (Cachão et al., 2008). Therefore, they can reveal environmental changes and play an essential role in developing palaeo-rocky shores and wave-cut platforms during marine transgressive events (Demircan, 2012).
The Trypanites Ichnofacies ranges in age from the Early Cambrian to the Holocene (Taylor & Wilson, 2003; Wilson, 2007), and in the geological past, this ichnofacies peaked during the Ordovician, Devonian, Jurassic and Tertiary (Wilson, 2007). However, it is concluded here that during the Miocene, the Trypanites Ichnofacies was at its peak, as manifested by publications from different countries (Abdel-Fattah & Assal, 2016; Belaústegui et al., 2012; Bromley & D’Alessandro, 1984; Demircan, 2012; Demircan et al., 2021; Domènech et al., 2001; El-Hedeny & El-Sabbagh, 2018; Farinati & Kulkarni et al., 2007; Johnson et al., 2010; Patel & Shringarpure, 1998; Radwański, 1969, 1970; Radwański et al., 2011; Santos and Mayoral, 2008; Santos et al., 2008, 2010, 2011; Uchman et al., 2018, 2002; Naimi et al., 2021; Zavala, 2002;). In the context of NE India, Mehrotra et al. (2001) recorded a solitary macroboring, Teredolites clavatus, from the Miocene Bhuban Formation of the Aizawl, Mizoram. Many ichnofossils were documented from the Miocene Bhuban succession of the Aizawl, Mizoram (Lokho & Singh, 2013; Rajkonwar et al., 2013, 2014a,b; Tiwari et al., 2011, 2013). Fifteen ichnospecies were recorded by Singh et al. (2010) from the Oligocene-Miocene of Bhuban and Boka Bil formations, Thangjing Hill Range, Manipur. However, no record of the Trypanites Ichnofacies is available from the Miocene Epoch of Manipur, NE India. Because of this, the authors have attempted to record the bioerosional structures of Trypanites Ichnofacies from the Miocene Boka Bil Formation of the Thangjing Hill Range of Manipur.
The aims of the present study are threefold: (a) to record the bioerosional structures of the Boka Bil Formation, (b) to analyse and interpret the relative abundance of ichnospecies and (c) to interpret the paleoenvironment of the Miocene sediments of the section.
Materials and Methods
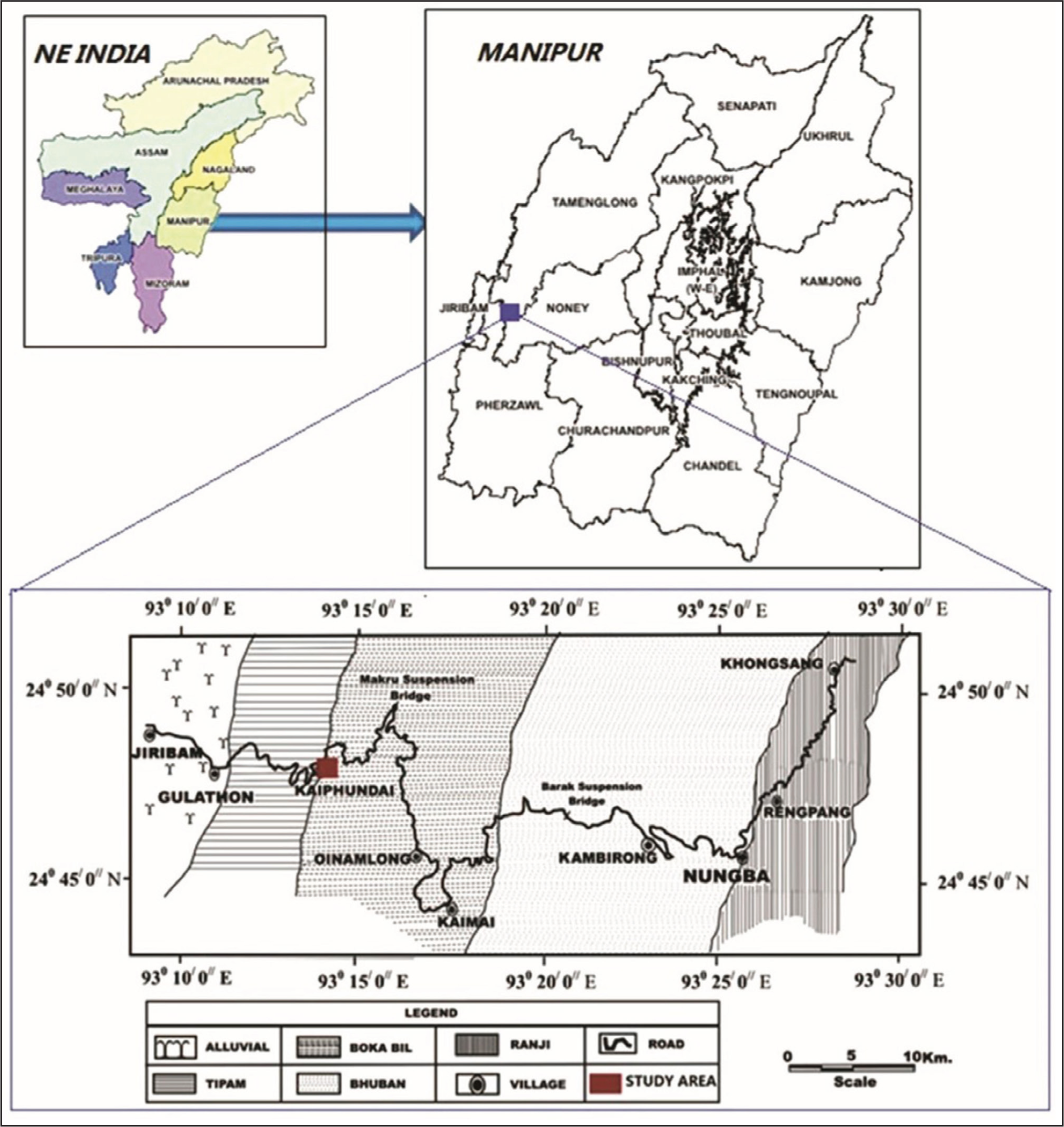
Field investigations included examining and collecting borings from the upper unit of the Miocene Boka Bil Formation. Boring material was collected from the two outcrops near Kaiphundai village, Thangjing Hill Range of Manipur (Figure 1). Outcrop no.1 (24° 47ʹ42ʺ N-93° 13ʹ50ʺ E) of the Kaiphundai village (Figure 2a). In this outcrop, fossiliferous muddy shale, clayey sandstone and hard carbonates rocks are associated with non-fossiliferous massive and pinkish ferruginous sandstone. Outcrop no. 2 (24°47ʹ 38ʺ N–93°13ʹ54ʺE) of the Kaiphundai village (Figure 2b). Some tens of metres northeast of outcrop no. 1, outcrop no. 2 comprise mainly fossiliferous poorly lithified carbonate rocks associated with non-fossiliferous bedded sandstone and siltstone.
In the field, sedimentological characteristics of these beds were observed. The geometry, thickness, lateral extent, packing, sorting, lower and upper contacts and taxonomic composition of the beds were investigated. The observed and collected borings from the outcrops were photographed and identified to ichnogenera and ichnospecies levels based on morphological characters. The relative abundance and distribution of were also documented. All the studied specimens occur as natural casts. A few raw cut sections of the specimens were also photographed for morphological analysis of some specific ichnogenera. Relative abundance data of bioeroders were made after proper research in the field and laboratory observations. All the collected specimens are housed in the Earth Sciences Department, Manipur University, under reference numbers ESD/MU/IF/B/0717 to 0781.
Location and Geological Map of the Kaiphundai Section, Thangjing Hill Range, Manipur, NE India (after Singh et al., 2010).
General Stratigraphy and Sedimentology
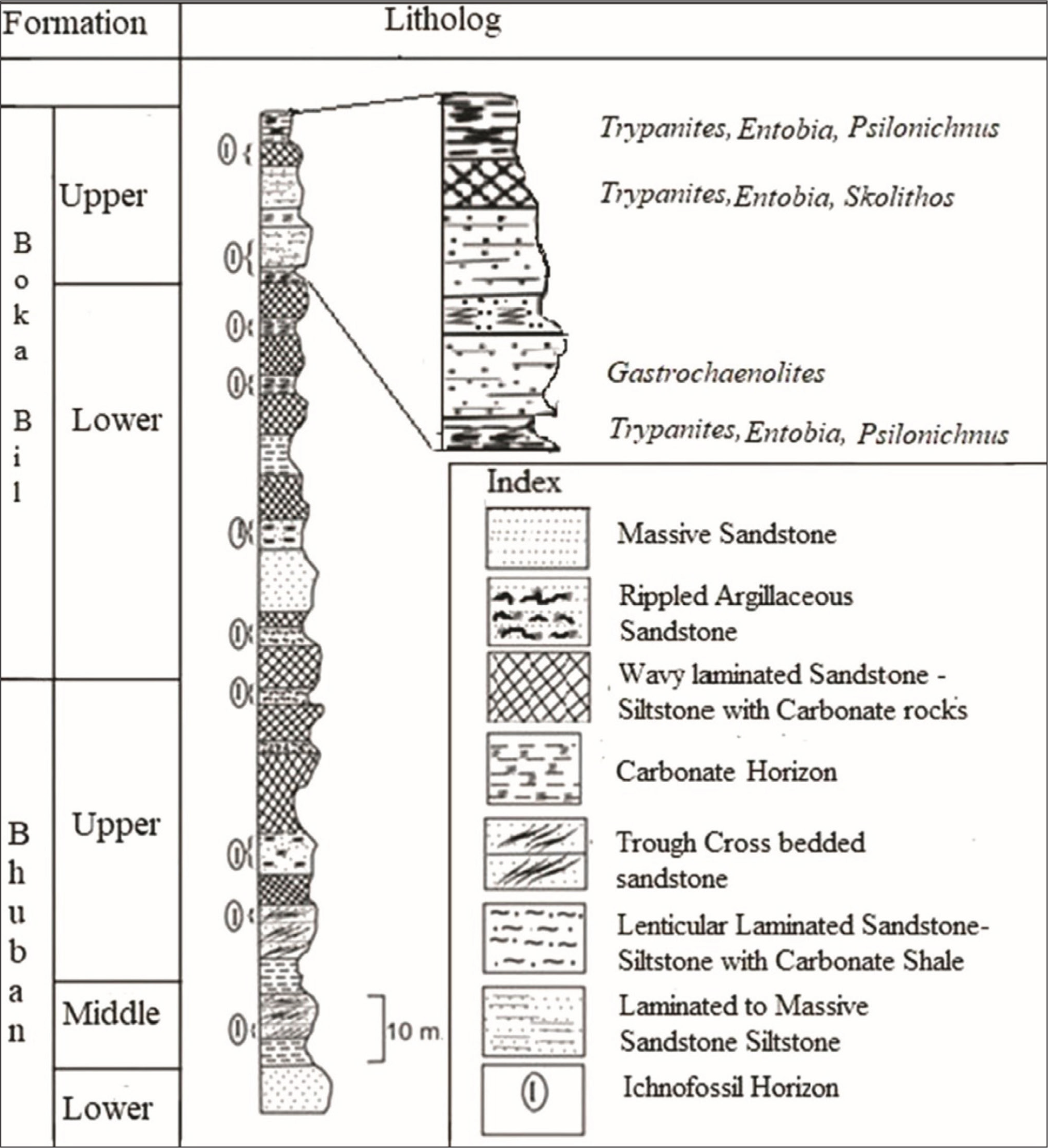
The Paleogene-Neogene lithostratigraphic succession in the Thangjing Hill Rage of Manipur has been grouped into the Barail, the Surma and the Tipam groups. The Miocene sediments of the Surma Group are unconformably underlain by the Barail Group and overlain by the Tipam Group (Table 1). It has been divided into two formations, Bhuban and Boka Bil (Figure 3). The Bhuban Formation is divided into Lower, Middle and Upper units. The Boka Bil Formation is divided into Lower and Upper units (Figure 3). The entire sedimentary column of the Bhuban and Boka Bil formations is a repetitive succession of arenaceous and argillaceous rocks. Lithologically, it mainly consists of sandstone, siltstone, shale, mudstone and a few pockets of shaly limestone and calcareous sandstone. The hard carbonate rocks are locally exposed in the upper unit of the Boka Bil Formation (Figure 2a).
Stratigraphic Succession of the Thangjing Hill Range (Modified after Kachhara et al., 2000).

(a) Field Photograph of Kaiphundai First Outcrop, Showing Inclined Strata of Coral and Carbonate Facies Associated with Massive, Unfossiliferous Pinkish Ferruginous Sandstone. Scale Bar: 50 cm. (b) Field Outcrop Photograph of the Kaiphundai Second Outcrop, Showing Incline Strata of Marly Sandstone Associated with Fossiliferous Light Grey and Dark Grey Shale and Fine-Grained Massive Sandstone. Scale Bar: 50 cm. (After Singh et al., 2010).

Generalized Lithocolumn of the Bhuban and Boka Bil Formations of the Thangjing Hill Range Showing Distributions of Ichnofossils (Modified after Singh et al., 2010).
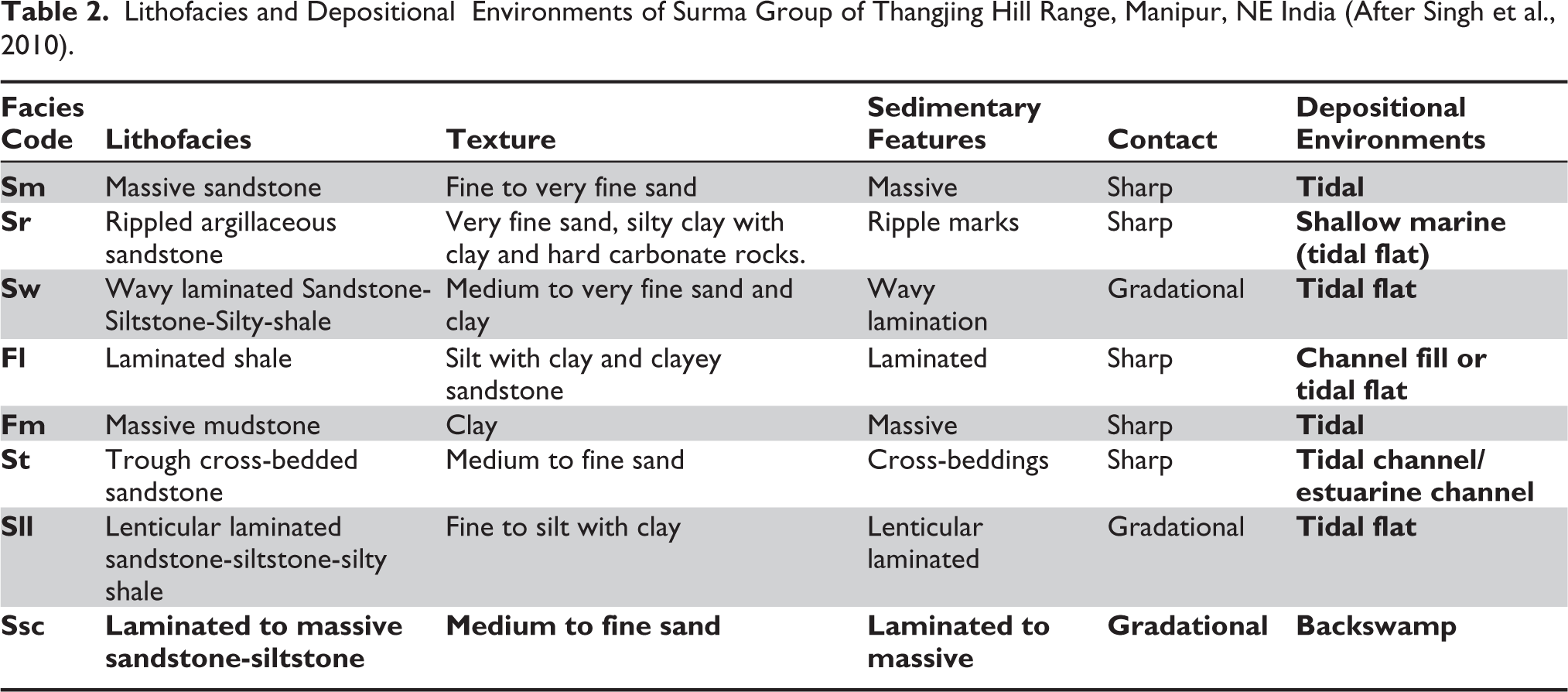
The Miocene molasses sediments of the Surma Group exposed in the Thangjing Hill Range of Manipur are characterised by several lithofacies containing many ichnofossils. Total of eight lithofacies: massive sandstone (Sm), rippled argillaceous sandstone (Sr), wavy laminated sandstone-siltstone-silty shale (Sw), laminated shale (Fl), massive mudstone (Fm), trough cross-bedded sandstone (St), lenticular laminated sandstone-siltstone-silty shale (Sll) and laminated to massive sandstone-siltstone (Ssc) are identified and described based on the scheme suggested by Miall (1978, 1990), Rust (1978), Reading (1996), Allen (1970), Reineck and Singh (1980), Walker (1984) and Stow (1986). The description of these lithofacies and their essential characteristics are shown in Table 2. The Bhuban and Boka Bil Formations sediments were deposited under fluctuating sea levels and moderate to strong energy conditions in lower intertidal to subtidal environments based on sedimentological attributes and ichnoassemblages (Singh et al., 2010).
Lithofacies and Depositional Environments of Surma Group of Thangjing Hill Range, Manipur, NE India (After Singh et al., 2010).
Description of Lithofacies
Massive Sandstone Facies (Sm)
This lithofacies commonly occurs in the middle and lower units of the Bhuban and Boka Bil formations exposed in the Sebirong and Oinamlong sections, respectively. The light grey fine grain massive sandstone well represents it. At places, it consists of reddish brown ferruginous massive sandstone containing Thalassinoides isp. (Singh et al., 2010), indicating shallow marine deposition environment.
Ripple Argillaceous Sandstone Facies (Sr)
It is characterised by ripple marks in the yellowish-white clayey sandstone and hard carbonate rocks. It occurs in the lower and upper unit of the Boka Bil Formation exposed in the Oinamlong and Kaiphundai sections. Maeandropolydora sulcans, Trypanites weisei and Entobia ovula bioerosion are associated with this facies in the Kaiphundai section and Psilonichnus upsilon in the Oinamlong section (Singh et al., 2010), suggesting backshore marginal shallow marine environment (Frey et al., 1984).
Wavy Laminated Sandstone-Siltstone-Silty Shale Facies (Sw)
It occurs in the Upper unit of Boka Bil Formation exposed in the Kaiphundai section. It is characterised by wavy laminated to thinly bedded fine-grained, light grey to buff sandstone, siltstone, carbonate shale associated with Entobia ovula, Trypanites bioerosion and Skolithos ichnofossils (Singh et al., 2010), indicating a shallow marine sub-tidal depositional environment.
Laminated Shale Facies (Fl)
It is characterised by the presence of alternate sub-facies of thinly laminated yellowish grey and light grey silty shale facies, which occurs in the lower unit of the Boka Bil Formation, exposed in the Oinamlong and Kaiphundai sections. Gyrochorte comosa, Helminthoida isp., Lockeia and Phycodes isp. are associated with this facies in the Oinamlong section (Singh et al., 2010), suggesting a shallow marine sub-tidal depositional environment with moderate to high energy conditions.
Massive Mudstone Facies (Fm)
Massive mudstone facies is characterised by the presence of massive light grey carbonate mudstone. It is well-exposed in the lower and upper unit of the Boka Bil Formation, in the Kaiphundai and Oinamlong sections. It consists of Gastrochaenolites lapidicus, G. torpedo, G. anauchen, G. cluniformis, G. orbicularis and G. ornatus in the Kaiphundai section, suggesting a shallow marine environment whereas, in the Oinamlong section, it is associated with Rutichnus, Planolites, Palaeophycus, Fucusopsis, Gyrochorte and Ophiomorpha isp. (Singh et al., 2010) suggesting fluctuation in the sea level and energy condition.
Trough Cross-bedded Sandstone Facies (St)
It is characterised by the presence of trough cross-bedding in the brownish to light grey moderate to fine-grained sandstone and exposed in the middle unit of the Bhuban Formation in Kambirong and Sebirong sections. It consists of Planolites, Ophiomorpha and Diplocraterion isp. (Singh et al., 2010) in the two sections, indicating a shallow marine sub-tidal depositional environment.
Laminated to Massive Sandstone-Siltstone-Silty Shale Facies (Ssc)
This facies is represented by laminated to massive, reddish brown to greyish white, fine-grained sandstone, siltstone and carbonate shale. It occurs in the upper unit of the Boka Bil Formation exposed in the Kaiphundai section. It is marked by the presence of Gastrochaenolites, Entobia and Trypanites bioerosion suggesting a shallow marine environment.
Lenticular Laminated Sandstone-Siltstone-Silty Shale Facies (Sll)
It is characterised by the presence of lenticular structure within the thinly bedded sandstone, siltstone and carbonate shale. It occurs in the upper unit of the Boka Bil Formation, exposed in the Kaiphundai and Barak Waterfall sections. The Kaiphundai section consists of Gastrochaenolites, Trypanites and Entobia bioerosion. In contrast, in the Barak Waterfall section, this facies is associated with Planolites beverleyensis, Palaeophycus tubularis, Fucusopsis, Gyrochorte comosa, Ophiomorpha isp. (Singh et al., 2010), suggesting a shallow marine sub-tidal depositional environment with moderate to high energy conditions.
Laminated to Massive Sandstone-Siltstone Facies (Ssc)
This facies is defined based on the presence of laminated to massive, reddish brown to greyish white, medium to fine-grained sandstone and siltstone. It occurs in the upper unit of the Boka Bil Formation exposed in the Kaiphundai section. It consists of Palaeophycus isp. (Singh et al., 2010), suggesting a back swamp environment.
Systematics
Ethological categorisation of macroborings by Gibert et al. (1998); Buatois et al. (2016); categories of architectural designs in trace fossils by Buatois et al. (2017) and ichnofamilies classification by Wisshak et al. (2019) are used in the present article. As the hard substrate macroboring assemblages of the Paleogene-Neogene resemble very closely to those of the Holocene (Taylor & Wilson, 2003) and the present 11 ichnospecies are from the Miocene, a few occurrences of these ichnospecies from the Paleogene-Neogene and the Quaternary horizons from various countries are mentioned with appropriate places.
Ichnofamily Entobiaidae Wisshak et al. (2019)
Ichnogenus Entobia Bronn (1837)
Entobia cateniformis Bromley and D’Alessandro (1984)
(Plate 1(1))
(1) Photographs of the Network Galleries of Entobia Cateniformis (Ec) Showing a Dense Boxwork (Boxes); (2) Photograph of Entobia in the Reefal Carbonates Facies, Showing Intensive Borings of Entobia Ovula (Eo) and ‘Worm’ Borings Trypanites Weisei (Tw) from Kaiphundai Second Outcrop. Scale Bar: 8 mm.

Material: Two specimens (ESD/MU/IF/B: 0745-0746).
Description: Boring system consists of camerate network galleries characterised by narrow and elongated chambers. The shape of the chambers is subrounded to subrectangular, long axes with an average length of about three-fourths of mm and short axes, approximately half of the long axis. Short, well-developed apertural canals rarely occur. The chambers are connected by poorly defined canals or fused without intercameral canals.
Remarks: Entobia is categorised as ‘camerate box work boring’ concerning architectural designs of trace fossils produced by sponges (Buatois et al., 2017). Ethologically, Entobia is classified as domichnion (Buatois et al., 2016; Gibert et al., 1998). Entobia ranges in age from the Devonian to the Holocene (Bromley, 1970, 2004; Bromley & D’Alessandro, 1984, 1989; Buatois et al., 2016; Fursich et al., 1994; Mikuláš, 1992; Taylor & Wilson, 2003; Wilson, 2007). Entobia cateniformis is known from the Early Pleistocene Calcarenite di Gravina Formation, Italy (Bromley & D’Alessandro, 1984); the Early Miocene Chhasra Formation of Kachchh, India (Kulkarni et al., 2007); the Middle Miocene of Ukraine (Radwański et al., 2011); the Miocene rocky shore of Concepción Formation, México (Pineda-Salgado et al., 2015); the Miocene reefs of the northwest Red Sea, Egypt (Abdel-Fattah & Assal, 2016) and the Early Eocene of Kachchh Basin, India (Gurav & Kulkarni, 2018).
Horizon and Locality: Carbonate sandy sandstone of Sr facies, upper unit of the Boka Bil Formation, exposed in Kaiphundai section (outcrop no. 2), Thangjing Hill Range, Manipur, NE India.
Entobia ovula Bromley and D’Alessandro (1984)
Material: Three specimens (ESD/MU/IF/B: 0741-0743).
Description: Casts of borings box work galleries comprise crowded ovoid to globular chambers characterised by light, almost straight tunnels, about 0.5 mm in diameter. The chambers are aligned and contact each other directly or indirectly via short intercameral canals. The fusion of two or more chambers was observed. Apertural openings are not well preserved.
Remark: Entobia ovula is known from the Miocene rocky shore in southern Iberia (Santos et al., 2008); the Miocene rocky shore in south Spain (Santos et al., 2011); the Miocene Reefs of the northwest Red Sea, Egypt (Abdel-Fattah & Assal, 2016); the Middle Miocene of the northern Western Desert of Egypt (El-Hedeny & El-Sabbagh, 2018); the early Eocene of Kachchh Basin, India (Gurav & Kulkarni, 2018) and the late Pleistocene of Red Sea Coast, Saudi Arabia (Demircan et al., 2021).
Horizon and Locality: Carbonate sandy sandstone of Sr facies, Carbonate silty shale of Ssc and Sll facies, upper unit of the Boka Bil Formation, exposed in Kaiphundai section (outcrop no. 2), Thangjing Hill Range, Manipur, NE India.
Ichnofamily Gastrochaenolitidae Wisshak et al. (2019)
Ichnogenus Gastrochaenolites Leymerie (1842)
Gastrochaenolites anauchen Wilson and Palmer (1998)
(Plate 2(3))
Material: Three specimens (ESD/MU/IF/B: 0725 - 0727).
Description: Borings with smooth, elongated and circular cross-section throughout length; expand gradually below the aperture, with a maximum diameter about three-fourths of the depth; bases are rounded; no distinguishable neck. The longitudinal axes of the borings are mostly perpendicular to the substrate surfaces, but a few have low angles inclination < 30° (Plate 2(3)). Aperture diameters range from 2 to 6 mm, and longitudinal depths range from 8 to 14 mm.
Remarks: Gastrochaenolites is categorised as ‘clavate-shaped boring’ with regard to architectural designs of trace fossils (Buatois et al., 2017). Ethologically, Gastrochaenolites is classified as domichnion (Buatois et al., 2016; Gibert et al., 1998). Gastrochaenolites ranges in age from the Ordovician to the Holocene (Buatois et al., 2016; Ekdale & Bromley, 2001; Kelly & Bromley, 1984; Wilson, 2007; Wilson & Palmer, 1998; Taylor & Wilson, 2003). Gastrochaenolites anauchen is reported from the rocky shore deposit of Pennsylvania (Johnson, 1988, 1992) and the early Pennsylvanian, Arkansas, USA (Wilson & Palmer, 1998)
Horizon and Locality: Carbonate mudstone of Fm facies and carbonate shale of Sll facies, upper unit of the Boka Bil Formation exposed at Kaiphundai section (Outcrop no. 1), Thangjing Hill Range, Manipur, NE India
Gastrochaenolites cluniformis Kelly and Bromley (1984)
(Plate 2(4))
Material: Two specimens (ESD/MU/IF/B: 0730 - 0731).
Description: Smooth, unlined, clavate-shaped borings, straight to slightly curved, bilobate at the base and circular to ovate at the aperture. The maximum diameter is located near the centre of the main chamber, which has one principal ridge. The neck and aperture are rounded to oval in cross-section. The observed length and width of the boring vary from 8 to 15 mm and 3 to 10 mm, respectively.
Remarks: Gastrochaenolite cluniformis is known from the Pliocene of Gujarat, India (Kulkarni et al., 2008); the middle Miocene of the northern Western Desert of Egypt (El-Hedeny & El-Sabbagh, 2018) and the middle Miocene rocky coast of the Para Tethys, Poland (Uchman et al., 2018).
Horizon and Locality: Carbonate mudstone of Fm facies and carbonate shale of Sll facies, upper unit of the Boka Bil Formation exposed at Kaiphundai section (Outcrop no. 1), Thangjing Hill Range, Manipur, NE India.
Gastrochaenolites lapidicus Kelly and Bromley (1984)
(Plate 2(1))
Material: Five specimens (ESD/MU/IF/B: 0717 - 0721).
Description: Smooth, elongated, clavate borings with a narrow apertural neck, showing a circular outline with a parabolic to rounded truncated base. The apertural region is narrower than the elongated main chamber. The main chamber has the widest dimension near its centre. The aperture neck and main chamber exhibit a circular cross-section. The length of the borings varies from 11 to 14 mm and the width from 3 to 8 mm. Inner surfaces of the borings are mostly affected by the second generation of the borings.
Remarks: Gastrochaenolites lapidicus is known from the Pliocene of the northwestern Mediterranean (Gibert et al., 1998); the Pliocene of Gujarat, India (Kulkarni et al., 2008); the Miocene rocky shores in southern Iberia (Santos et al., 2008); the Miocene Reefs of the northwest Red Sea, Egypt (Abdel-Fattah & Assal, 2016); the Middle Miocene of the northern Western Desert of Egypt (El-Hedeny & El-Sabbagh, 2018); the Early Oligocene of the Tertiary Piedmont Basin, Italy (Uchman et al., 2017); the Middle Miocene of the Carpathians, Poland (Uchman et al., 2018) and the Late Pleistocene of the Red Sea Coast, Saudi Arabia (Demircan et al., 2021).
Horizon and Locality: Carbonate mudstone of the Fm facies and carbonate shale of the Sll facies upper unit of the Boka Bil Formation exposed at Kaiphundai section (Outcrop no. 1), Thangjing Hill Range, Manipur, NE India.
Gastrochaenolites orbicularis Kelly and Bromley (1984)
(Plate 2 (5))
(1) Gastrochaenolites Lapidicus Showing Elongated Nature; (2) G. torpedo Showing Club-Shaped Configuration; (3) G. anauchen Showing Elongated Nature (4) G. cluniformis Showing Elongated Nature; (5) G. orbicularis Showing Bilobate Nature at the Basal Section; (6) G. torpedo Showing Orbicular Main Chamber; (7) G. torpedo (Gt) and G. lapidicus (Gl) Showing Club-Shaped Configuration from Kaiphundai First Outcrop; and (8) Photographs of Entobia ovula (Eo) and the ‘Worm’ Borings Showing Substrate-Parallel Trypanites Solitarius (Ts)), Vertically Inclined T. weisei (Tw) from Kaiphundai First Outcrop. Scale Bar: 5 mm.

Material: Three specimens (ESD/MU/IF/B: 0733 – 0735).
Description: Borings with smooth, clavate, and circular cross-section throughout. The main chamber is orbicular, and neck region is elongated or may be short. Borings with the orbicular main chamber, slightly shallow depressions with subcircular bases, 3 to 4 mm deep. A short or long neck separates the aperture.
Remarks: Gastrochaenolitetes orbicularis is known from the Pliocene of Gujarat, India (Kulkarni et al., 2008); the Early Oligocene of Tertiary Piedmont Basin, Italy (Uchman et al., 2017); the Middle Miocene of the northern Western Desert of Egypt (El-Hedeny & El-Sabbagh, 2018) and the Middle Miocene rocky coast of the Paratethys, Poland (Uchman et al., 2018).
Horizon and Locality: Carbonate mudstone of Fm facies and carbonate shale of Sll facies, upper unit of the Boka Bil Formation exposed at Kaiphundai section (Outcrop no. 1), Thangjing Hill Range, Manipur, NE India.
Gastrochaenolites ornatus Kelly and Bromley (1984)
(Plate 3(1–5))
Material: Five specimens (ESD/MU/IF/B: 0777 - 0781).
(1–5) Photograph of Gastrochaenolites ornatus from the Kaiphundai Second Outcrop: (1–3) Specimens View in Lateral and Basal; (4–5) Specimens with Casts of Internal Surfaces of Bivalves; (6) Field Photograph Showing Meandering Tunnel of Maeandropolydora sulcans (Ms), with Sediment-Filled Cast (White Arrow) from the Kaiphundai Second Outcrop; (7) Hand Specimen Photograph of the ‘Worm’ Borings Showing Substrate-Parallel Trypanites weisei, (Tw) and Entobia ovula (Eo) from Kaiphundai Second Outcrop. Scale Bar: 5 mm.
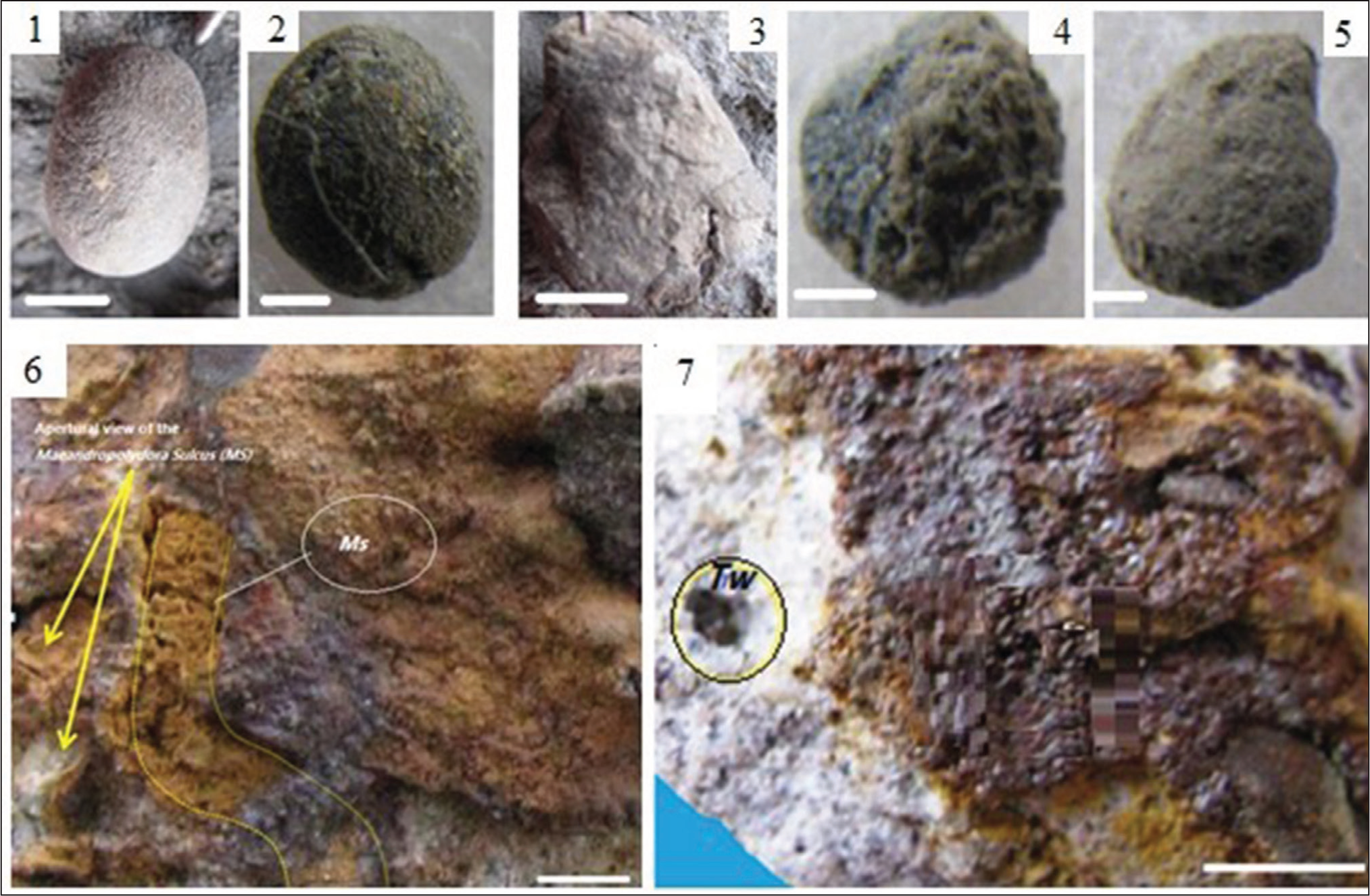
Description: Borings have drop-like chambers, are circular in cross-section, have sub-hemispherical bases, and taper gently upward. Deep parts of the chambers bear spiral scratches. Distance between scratches of a spiral bio glyph ranges from 1 to 2 mm. The maximum diameter of casts is about 33 mm and depth is 45 mm. The diameter/depth ratio varies considerably but, in most cases, increases towards the neck region.
Remarks: Gastrochaenolites ornatus is known from the Early Miocene discontinuity surface in Patagonia, Argentina (Carmona et al., 2007); the Miocene rocky shores in southern Iberia (Santos et al., 2008); the Miocene disconformity at Foz da Fonte, Portugal (Santos et al., 2010); the Miocene rocky shore in southern Spain (Santos et al., 2011) and the Miocene Reefs of the northwest Red Sea, Egypt (Abdel-Fattah & Assal, 2016).
Horizon and Locality: Carbonate mudstone of Fm facies, upper unit of the Boka Bil Formation exposed at Kaiphundai section (Outcrop no. 1), Thangjing Hill Range, Manipur, NE India.
Gastrochaenolites torpedo Kelly and Bromley (1984)
(Plate 2(2, 6, 7))
Material: Three specimens (ESD/MU/IF/B: 0722 - 0724).
Description: Elongate, smooth borings, with the base acutely parabolic. Their lengths vary from 15 to 18 mm and widths from 4 to 6 mm. The widest section is located midway between the base and the aperture. The neck has a constriction near the aperture. The apertures are circular with diameters of approximately 2 mm. The internal surfaces of borings are either smooth or display a sculpture of concentric lines.
Remarks: Gastrochaenolites torpedo is known from the Miocene rocky shores of many regions in Europe and neighbouring areas (Radwański, 1969); the Pliocene of the northwestern Mediterranean (Gibert et al., 1998); the Pliocene of Gujarat, India (Kulkarni et al., 2008); the Miocene of Mut Basin, Turkey (Uchman et al., 2002); the Miocene rocky shores in southern Iberia (Santos et al., 2008); the Miocene of Lower Tagus Basin, Portugal (Santos et al., 2010); the Late Miocene rocky shores from Menorca (Balearic Islands), Spain (Johnson et al., 2010); the Late Miocene of Cannakale, Turkey (Demircan, 2012); the Miocene Reefs of the northwest Red Sea, Egypt (Abdel-Fattah & Assal, 2016); the Early Oligocene of Tertiary Piedmont Basin, Italy (Uchman et al., 2017); the Middle Miocene of the northern Western Desert of Egypt (El-Hedeny & El-Sabbagh, 2018); the Middle Miocene of the Carpathians, Poland (Uchman et al., 2018) and Late Pleistocene of Red Sea Coast, Saudi Arabia (Demircan et al., 2021).
Horizon and Locality: Upper unit of the Boka Bil Formation exposed at Kaiphundai section (Outcrop no. 1), Thangjing Hill Range, Manipur, NE India in Carbonate mudstone of Fm facies.
Ichnofamily Dendrinidae Bromley et al. (2007)
Ichnogenus Maeandropolydora Voigt (1965)
Maeandropolydora sulcans Voigt (1965)
(Plate 3(6))
Material: Two specimens were observed in the field.
Description: Borings have cylindrical galleries with a constant diameter, irregularly twisted into complicated convolutions and preserved partially only part of the looping limb, without vane or pouch development, associated with another smaller specimen having two apertures. The width of the boring ranges from 0.5 to 2.8 mm, whereas the length ranges in a few tens of mm.
Remarks: Maeandropolydora is categorised as ‘winding boring’ with regard to the architectural designs of trace fossils (Buatois et al., 2017). Ethologically Maeandropolydora has been classified as domichnion (Buatois et al., 2016; Gibert et al., 1998) and ranges in age from the Jurassic to the Pleistocene (Bromley & D’Alessandro, 1983; Buatois et al., 2016; Taylor & Wilson, 2003; Wilson, 2007; Zıtt & Mikuláš, 1994). Maeandropolydora sulcans is known from the Pliocene of the northwestern Mediterranean (Gibert et al., 1998); the Late Miocene of Portugal (Santos & Mayoral, 2008); the Miocene rocky shores in southern Iberia (Santos et al., 2008); the Miocene rocky shore in southern Spain (Santos et al., 2011); the Late Miocene of Cannakale, Turkey (Demircan, 2012); the Miocene Reefs of the northwest Red Sea, Egypt (Abdel-Fattah & Assal, 2016) and the Middle Miocene of the northern Western Desert of Egypt (El-Hedeny & El-Sabbagh 2018).
Horizon and Locality: Argillaceous, sandy sandstone of Sr facies, upper unit of the Boka Bil Formation, exposed in Kaiphundai section (Outcrop no. 2), Thangjing Hill Range, Manipur, NE India.
Ichnofamily Trypanitidae Mägdefrau (1932)
Ichnogenus Trypanites Mägdefrau (1932)
Trypanites solitarius Hagenow (1840)
(Plate 2(8))
Material: Two specimens (ESD/MU/IF/B: 0746 - 0747).
Description: Borings are curvilinear to slightly sinuous cylindrical to subcylindrical, circular in cross-section with single apertural openings and distal portions taper to a blind end. The borings are parallel or sub-parallel to the surface. The borings have a length of a few tens of mm with an average diameter of 1.5 mm.
Remarks: Trypanites is categorised as ‘cylindrical vertical to oblique boring’ with regard to the architectural designs of trace fossils (Buatois et al., 2017). Ethologically Trypanites is classified as domichnion (Buatois et al., 2016; Gibert et al., 1998) and ranges in age from the Cambrian to the Holocene (Bromley, 1972; Buatois et al., 2016; Cole & Palmer, 1999; James et al., 1977; Kelly & Bromley, 1984; Kobluk et al., 1978; Kobluk & Nemcsok, 1982; Taylor & Wilson, 2003; Wilson, 2007). Trypanites solitarius is known from the Miocene Reefs of the northwest Red Sea, Egypt (Abdel-Fattah & Assal, 2016); the Miocene Reefs of the northwest Red Sea, Egypt (Abdel-Fattah & Assal, 2016) and the Early Oligocene of Tertiary Piedmont Basin, Italy (Uchman et al., 2017).
Horizon and Locality: Carbonate silty shale of Sll and Ssc facies, upper unit of the Boka Bil Formation, exposed in Kaiphundai section (Outcrop no-2), Thangjing Hill Range, Manipur, NE India.
Trypanites weisei Mägdefrau (1932)
Material: Three specimens (ESD/MU/IF/B: 0751 and 0753).
Description: Single entrance, cylindrical to sub-cylindrical, unlined and unbranched narrow borings in hard substrates, mostly oriented perpendicular to the bedding plane. The axes of the borings may be straight, curved or irregular. It is 1.2 to 2.3 mm in diameter, and its length varies from specimen to specimen within a few tens of mm. The base of the borings is blind and has a tapered to rounded termination.
Remarks: Tripanite weisei is known from the Pliocene of the northwestern Mediterranean (Gibert et al., 1998); the Miocene of Lower Tagus Basin, Portugal (Santos et al., 2010); the Miocene Reefs of the northwest Red Sea, Egypt (Abdel-Fattah & Assal, 2016) and the Middle Miocene of the northern Western Desert of Egypt (El-Hedeny & El-Sabbagh, 2018).
Horizon and Locality: Carbonate silty shale of Sll and Ssc facies, upper unit of the Boka Bil Formation, exposed in Kaiphundai section (Outcrop no-2), Thangjing Hill Range, Manipur, NE India.
Discussion
Global Occurrences of Trypanites Ichnofacies During the Miocene
The Miocene Epoch was significant in the geological past as the modern continents and oceans assumed their present positions (Torsvik et al., 2017), the climates remained moderately warm, and the contemporary flora and fauna evolved. Important Miocene deposits occur in North and South America, southern Europe, India, Mongolia, East Africa and Pakistan. Bioerosional structures preserved as macroborings belonging to the Trypanites Ichnofacies are known from the Miocene of Poland (Radwański, 1969, 1970); Italy (Bromley & D’Alessandro, 1984); Spain (Belaústegui et al., 2012; Domènech et al., 2001); Argentina (Carmona et al., 2007); Portugal (Santos & Mayoral, 2008; Santos et al., 2010); Iberia (Santos et al., 2008); Turkey (Demircan, 2012; Uchman et al., 2002); Egypt (Abdel-Fattah & Assal, 2016; El-Hedeny & El-Sabbagh, 2018); Poland, (Uchman et al., 2018); Spain (Rodríguez-Tovar et al., 2017) and Saudi Arabia (Demircan et al., 2021). Thus, the aforementioned publications documenting macroborings belonging to the Trypanites Ichnofacies from the Miocene of different countries suggest that, in the geological past, the Trypanites Ichnofacies witnessed its peak during the Miocene of the Cenozoic Era. Only two ichnospecies, Entobia cateniformis and E. laquea, belonging to the Trypanites Ichnofacies, are known from the Early Miocene Chhasra Formation of Kachchh, India (Kulkarni et al., 2007). Hence, the present record of 11 ichnospecies of the Trypanites Ichnofacies from the Miocene Boka Bil Formation of Manipur, NE India, provides important information on the presence of bioerosional communities.
Bioerosional Structures and Bioeroding Communities During Miocene, Manipur
The relative abundance of the 11 ichnospecies recorded in the Miocene of Boka Bil Formation, Manipur (Table 3) reveals two distinguishing assemblages of macroboring, such as Gastrochaenolites-dominated and Entobia-dominated assemblage. The Gastrochaenolites-dominated assemblage mainly consists of macroborings of Gastrochaenolites of relatively large sizes and six species, namely, G. anauchen, G. cluniformis, G. lapidicus, G. orbicularis, G. ornatus and G. torpedo; out of these, G. anauchen, G. lapidicus, G. orbicularis and G. ornatus are the most common borings, followed by those of the G. torpedo and meagre occurrences of G. cluniformis. The Gastrochaenolites-dominated assemblage is commonly associated with Entobia ovula and Trypanites weisei, as well as rare occurrences of Maeandropolydora sulcans. The Entobia-dominated assemblage is dominated by dense networks and boxworks of the two ichnospecies of Entobia, E. ovula and E. cateniformis. Gastrochaenolites torpedo is the most common in this assemblage, followed by G. lapidicus. Trypanites solitarius and Maeandropolydora sulcans.
The Probable Tracemakers, Bored Substrates and Relative Abundance of the Ichnospecies (After Bromley, 1970, 1994 and Perry, 1996, 1998).
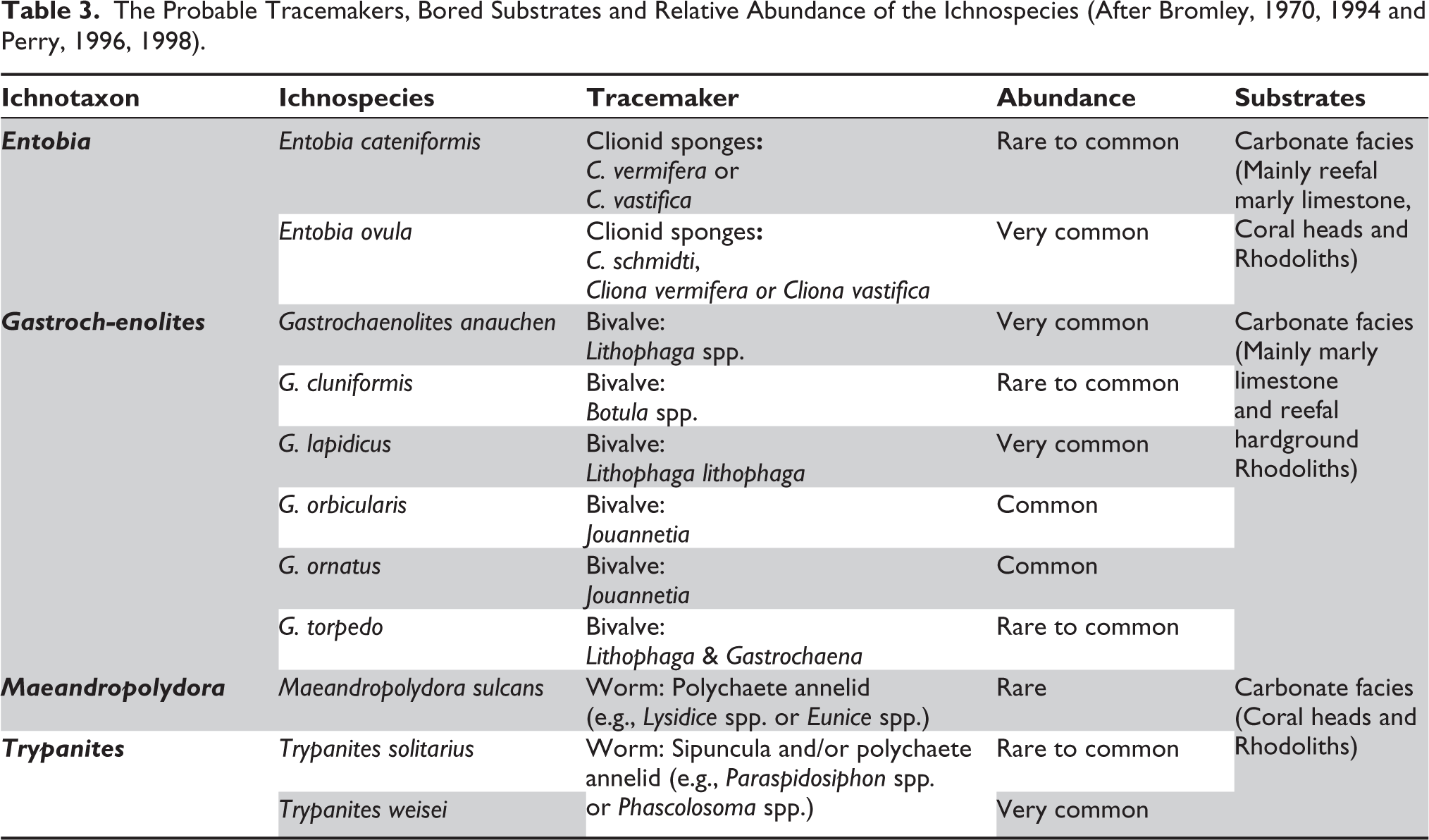
Entobia is produced by sponge Cliona (Bromley & Asgaard, 1993b), and in the Mediterranean Sea, Entobia ovula is produced by Cliona schmidtii (Ridley, 1881); C. vermifera (Hancock, 1867) and/or C. vastifica (Bromley & D’Alessandro, 1989; Bromley & Asgaard, 1993a; Hancock, 1849). The bivalve Lithophaga is considered to be the main producer of Gastrochaenolites (Bromley & D’Alessandro, 1987; Kelly & Bromley, 1984; Uchman et al., 2017, 2018). Gastrochaenolite cluniformis is produced by the bivalves Botula and Gastrochaena (Kelly & Bromley, 1984; Radwański, 1969). G. orbicularis and G. ornatus are produced by the bivalve Jouannetia (Kelly & Bromley, 1984; Uchman et al., 2017). G. torpedo is produced by bivalves, Lithophaga and Gastrochaena (Kelly & Bromley, 1984), and Maeandropolydora sulcans is produced by suspension-feeding spionid polychaetes (Bromley & D’Alessandro, 1983, 1987), like Recent Polydora hoplura Claparède. Trypanites is produced by polychaetes, sipunculoids, and acrothoracican barnacles (Bromley, 1992; Ekdale et al., 1984; Pemberton et al., 1980). Thus, the borings documented here represent bioeroding communities of bivalves, polychaetes, sponges, barnacles and worms.
Paleoenvironmental Implications of Bioerosional Structures of Miocene, Manipur
During the Miocene, the Kaiphundai region, the western part of the Thangjing Hill Range, NE India, was affected by marine transgression, which is reflected by the presence of macroborings characterised by Gastrochaenolites borings, especially G. torpedo, associated with carbonate rocks that can be interpreted as horizontal wave-cut platforms with low or no sedimentation (Demircan, 2012; Santos et al., 2010). Moreover, the occurrences of Gastrochaenolites supports shallow water environments (Bromley, 1992; Bromley & Asgaard, 1993).
Gastrochaenolites is abundant in carbonate hard substrates (Bromley, 1992; Bromley & Asgaard, 1993a; Kleemann, 1973). In the Mediterranean Sea, Lithophaga is restricted to a depth of up to 10 m (Kleemann, 1973). Trypanites and Maeandropolydora are mostly found in shallow-water settings. In general, the abundance of polychaetes and sipunculans is used as a good indicator of shallow-marine regimes in modern reef environments, such as lagoonal patch reefs to back-reef environments (Hutchings et al., 1992; Macintyre, 1984). Bivalve borings belonging to Gastrocheonlites isp. mixed with Entobia, Trypanities and Maeandropolydora inferred shallow-water, low-energy back-reef to lagoonal facies or nearshore environments are known from the Late Miocene reefs of Mallorca (Perry, 1996); Oligocene–Miocene Bluff Formation, Grand Cayman (Pleydell & Jones, 1988); the Pleistocene coral reef, Bahamas (Wilson et al., 1998) and Pleistocene Falmouth Formation, Jamaica (Perry, 2000). The Miocene bivalves, sponges, barnacles and worms’ borings from the Miocene of Portugal indicate a shallow marine rocky substrate with a negligible sedimentation rate (Santos et al., 2010).
The Gastrochaenolites-dominated assemblage indicates a tidal-flat setting mostly under a water depth of a few metre in a back-reef lagoon paleoenvironment. The ichnospecies of Entobia are the dominant boring communities in lagoonal patch reefs to back reefs, as well as fore-reef and offshore sites (Bromley, 1970; Hernández-Ballesteros et al., 2013; Perry, 1996, 1998, 2000; Risk et al., 1995; Rützler 1975, 2002; Sammarco and Risk, 1990). Entobia-dominated borings assemblage are known from the foreshore of the Holocene Caribbean reefs (Perry, 1998) and the Pleistocene Falmouth Formation, north Jamaica (Perry, 2000), as well as from the Oligocene reefs of Grand Cayman (Pleydell & Jones, 1988) and the Miocene reefs of Mallorca (Perry, 1996). The abundance of ‘worm’ borings associated with Entobia suggests shallow-water environments, that is, lagoon or shallow fore reef (Abdel-Fattah & Assal, 2016; Perry, 2000). Thus, the Entobia-dominated suggests relatively deeper marine conditions in the deeper part of the back reef or fore reef. The Trypanites Ichnofacies is directly associated with different unconformities, either omission or erosive surfaces (Bromley, 1975). Thus, the present ichnoassemblage of the Trypanites Ichnofacies exhibits post-omission suites recognised by Bromley (1975) as borings drilled after lithification.
Conclusion
The following conclusions are drawn from the present study:
Eleven ichnospecies, namely Entobia cateniformis, E. ovula, Gastrochaenolites anauchen, G. cluniformis, G. lapidicus, G. orbicularis, G. ornatus, G. torpedo, Maeandropolydora sulcans, Trypanites solitaries and T. weisei are reported from the upper unit of the Miocene Boka Bil Formation of the Thangjing Hill Range of Manipur, NE India. The observed spatial and sequential distribution of the macroborings reveals Gastrochaenolites and Entobia assemblages pointing to relatively deeper conditions in a deeper part of a back reef or fore reef and shallower marine conditions, underwater depths of a few metres, in the back reef to the patch-reef zone of a back-reef lagoon respectively. The documented ichnospecies belonging to the Trypanites Ichnofacies indicate the prevalence of hard substrate, reef, rocky shore, omission surface and ancient wave-cut platform.
Footnotes
Acknowledgements
The authors thank Professor Dr R. A. S. Kushwaha and Professor Dr S. Ibotombi of Manipur University, India, for their thoughtful comments and technical assistance, respectively, during the fieldwork and writing of this research article. The two anonymous reviewers who offered insightful comments and helpful ideas are gratefully acknowledged. The authors are thankful to Dr Shyam Kishore for improving the manuscript of our article at the editorial level.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors received no financial support for the research, authorship and/or publication of this article.