
Abstract
The marine fossiliferous Baripada Beds (Upper Miocene), on the eastern coast of India, are well known for yielding diverse species of sharks, batoids and teleost fishes, reptiles, mammals, invertebrates and microfossils (Foraminifera and Ostracoda). A recent study conducted at Mukurmatia and Itamundia localities yielded more than 200 additional specimens of shark and batoid teeth from these rocks. A total of nine families of sharks and batoids belonging to Carcharhinidae, Hemigaleidae, Alopiidae, Myliobatidae, Dasyatidae, Pristidae, Rhynchobatidae, Rajidae, and Sphyrnidae have been recorded from Mukurmatia and Itamundia localities. The present study also records additional elasmobranch species (Carcharhinus amblyrhynchoides, C. brevipinna, C. perezi, C. plumbeus, Physogaleus hemmooriensis, Physogaleus sp., Aetobatus cappettai and Taeniurops sp.) for the first time from these beds. These additional findings from Baripada Beds also represent the first report of elasmobranch species, namely: Carcharhinus amblyrhynchoides, C. brevipinna, Physogaleus hemmooriensis and Aetobatus cappettai from the Miocene deposits of the Indian subcontinent. The collected fauna in association with foraminifera and molluscs reported from here suggests the deposition of Baripada Beds in a coastal, lagoonal, near-shore environment under neritic conditions connected to the open ocean. Beta diversity (Sørensen–Dice coefficient) correlation study of the Miocene elasmobranchs of India and their global counterparts shows that the Upper Miocene fauna had a close affinity to that of the South Pacific and the Indo-Pacific oceans, suggesting a shift in migration path after the permanent closure of the east Mediterranean seaway.
INTRODUCTION
The Baripada Beds, deposited through the Miocene marine transgression event along east coast of India, have yielded a diverse fossil assemblages of sharks, batoids, teleosts, reptiles and mammals, invertebrates (bivalves, gastropods, bryozoans, echinoids, decapods) and microfossils (foraminifera and ostracods) (see Bhalla & Dev, 1988; Bhaumik et al., 2017; Chatterjee, 1973; Chaudhuri, 1958; Eames, 1936; Ghosh, 1956, 1959; Mehrotra, 1979; Mehrotra et al., 1973; Modak, 1952; Mohanty, 1966, 1980; Mondal et al., 2009; Sahni & Mehrotra, 1981; Sharma, 2013; Sharma & Patnaik, 2010, 2013, 2014; Tewari & Awasthi, 1960 and references therein). The first investigation of these beds can be traced back to the discovery of Ostrea-bearing limestone beds exposed at Mahulia (Bose, 1904). Among the fossil vertebrates, sharks and batoids are the most dominant groups. A considerable amount of work has been carried out on fossil elasmobranchs from the area (Ghosh, 1956, 1959; Jena, 1943; Modak, 1952; Mondal et al., 2009; Mohanty, 1966; Sahni & Mehrotra, 1981; Sahni et al., 1971; Sharma & Patnaik, 2013, 2014; Tewari & Awasthi, 1960 and references therein), yet a detailed revision of the systematics of the elasmobranch fauna from the Miocene deposits of India is the need of the hour (Adnet et al., 2007; Sharma et al., 2021). Moreover, these beds also serve as a prospective site for Upper Miocene elasmobranch diversity and migration patterns of Upper Miocene global paleobiogeography. Taking this into account, fossil and rock samples were collected from 2017 to 2022 from Mukurmatia (N 21° 50 39.34″: E 86° 43′ 5.6″) and Itamundia (N 21° 53′ 35.9″: E 86°43′ 35″) sections that yielded additional teeth of sharks and batoids. Here, we record the recovered elasmobranch fauna from these sections, some of which are being reported for the first time from the study area. The study also shows a scope for taxonomic revision of some of the chondrichthyan fauna and enriches the record of elasmobranchs from the east coast of India. The sharks and batoids being reported here, together with previously described fossils, have been used to reconstruct the palaeoenvironmental condition of the Baripada Beds. The current data have been integrated with the previous studies on Miocene elasmobranchs of India to discuss the Miocene paleobiogeographic conditions of the region.
GEOLOGICAL SETTING
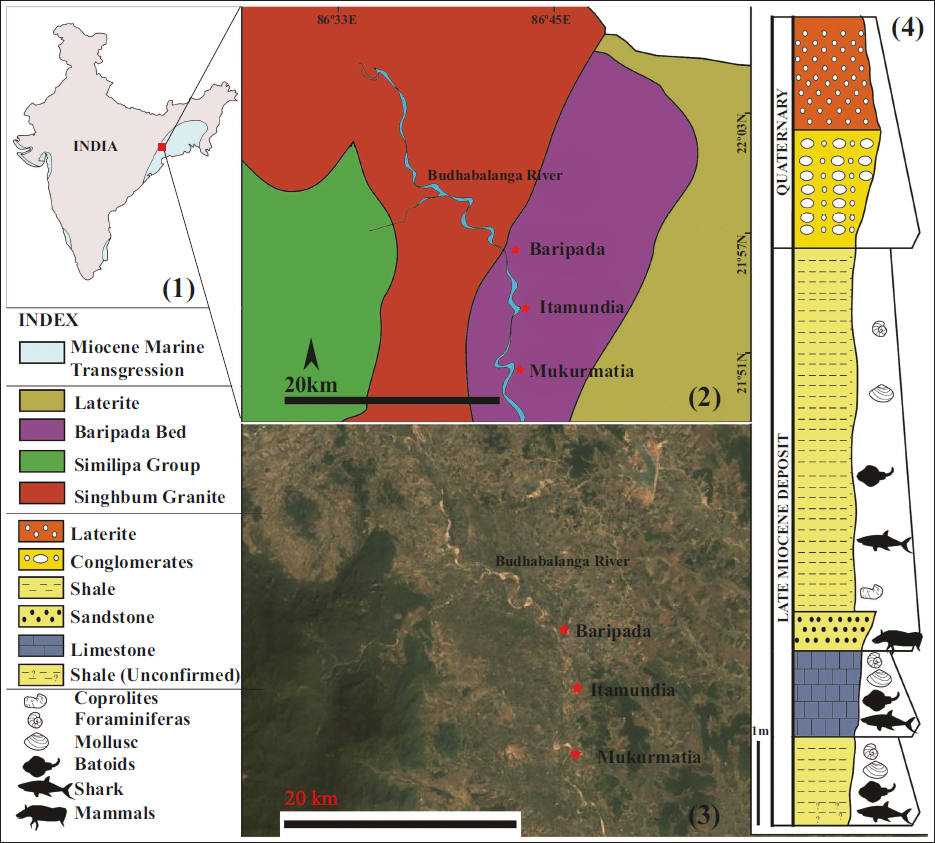
The Baripada Beds, exposed along the banks of the Budhabalanga River (Krishnan, 1982; Sharma & Patnaik, 2013, 2014), are deposited over the Precambrian basement along the east coast of Odisha (also see Bose, 1904; Faruque, 2006). The Cenozoic deposits, including Baripada Beds, overlie the Archean Singhbhum Granite, comprising Archean granitoids, gneisses, and migmatites interspersed with supracrustal assemblages, which in turn are intruded by mafic-ultramafic intrusive and layered magmatic complexes (Sharma, 2013 and references therein). The exposure of Similipal Group consists of an alternate spilitic lava and quartzite sequence lying unconformably above the Singhbhum Granite and occasionally intruded by pyroxene granite, granophyres, gabbro and anorthosite (Iyengar & Banerjee, 1964; Sharma, 2013). The exposure of a 14 m thick section of Baripada Beds along the bank of Budhabalanga River has yielded a variety of fossils (Figure 1). Lithologically, the beds comprise greenish to yellowish colour shales, soft sandstones and limestones. The limestone bed (~1.5 m) overlies the lowermost greenish to bluish coloured shales. The limestone is arenaceous towards the top, from where a few remains of sharks, batoids, bivalves, and foraminifera were reported (Bhaumik et al., 2017; Sahni & Mehrotra, 1981; Sharma & Patnaik, 2013 and references therein). The lower shale is comparatively less fossiliferous than the overlying limestone, sandstone and shales. The hard, compacted, yellowish to brownish colour limestone bed is dominated by fossil oysters, gastropods, foraminifera, teeth and bone remains of sharks and batoids (Bhalla & Dev, 1988; Eames, 1936; Sahni & Mehrotra, 1981 and references therein). A coarsening upward sequence is observed from the limestone bed to the sandstone bed, overlain by the shale bed. The shale and sandstone beds also yielded foraminifera, molluscs, teeth and bone remains of sharks, batoids, teleosts, crocodiles, turtles, etc. (Bhalla & Dev, 1972, 1988; Sahni & Mehrotra, 1981; Sharma & Patnaik, 2010, 2013, 2014). The shale bed is overlain by unfossiliferous thick, hard, compact Quaternary deposits of conglomerate/laterite beds (Figure 1). There is a sharp unconformable boundary between the Miocene Baripada Beds and the successive younger Quaternary deposits (Sharma, 2013).
(1) Outline map of India showing the Middle and Upper Miocene marine deposits along the Indian coast (Redrawn after Sahni & Mitra, 1980); (2) Geological map of the study area (Faruque, 2006). (3) Location map of the study area (source: Google Earth); (4) Generalised stratigraphic log of Baripada Beds exposed around Makurmatia and Itamundia (Odisha, India).
Age of Baripada Beds
The Baripada Beds are considered to be deposited during the late Miocene within a time frame of ~12.6 to ~8.3 Ma, based on the faunal assemblage of planktic foraminifera (Globorotalia menardii, Neogloboquadrina continuousa, etc) (see Bhaumik et al., 2017). An earlier report of Tetraconodon intermedius from the bluish grey sandy shale bed overlaying the limestone deposits of Itamundia assigned an age range of ~10-8 Ma (Sharma & Patnaik, 2013) to the marine beds of 10–14 metres thick, exposed at the banks of the Burabalang River. The maximum thickness of the Baripada Beds exposed at the Makurmatia section along the right bank of the Burbhalang River is ~14 m (Figure 1).
METHODOLOGY
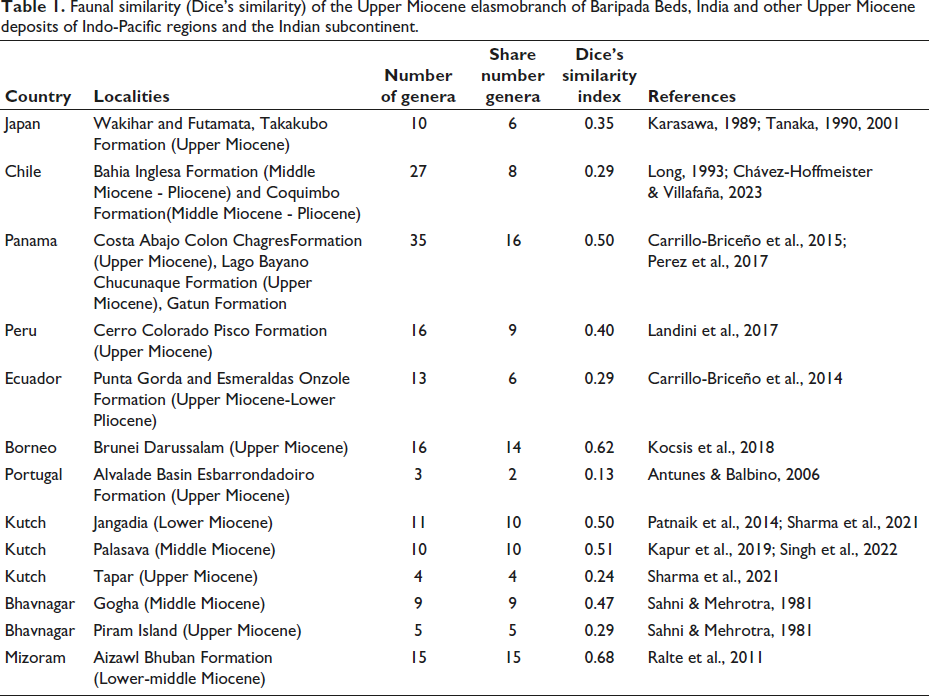
In situ samples of sharks’ and batoids’ teeth were collected from the field sites of Mukurmatia and Itamundia localities of Odisha, India. The microscopic remains were recovered from approximately 450 kg of shale samples after maceration in the laboratory following the technique suggested by Sharma and Patnaik (2014). The disintegrated samples were then sieved using ASTM sieves ranging in size from 74 to 595 µm. The residues were air-dried, and microfossils present in the macerates were picked up using a binocular microscope. Photography of the collected fossil samples was carried out using a Leica M205C stereo zoom trinocular microscope housed at the Department of Geology, Central University of Punjab, Bathinda. Scanning Electron Microscopy of a few of the samples was done using JSM JEOL, 6460, housed at the Centre of Advanced Study in Geology, Panjab University, Chandigarh. Teeth terminology after Cappetta (2012) is adopted in this article. Dice similarity coefficient is calculated for the generic level taxa from Baripada Beds and other Upper Miocene deposits of Indo-Pacific regions and Indian subcontinent (Table 1) for studying their faunal similarity using the formula Dice Coefficient = 2 * |A ∩ B|/(|A| + |B|) where |A ∩ B| represents the number of genera common to both locality A and B and |A| and |B| are the sizes of genera in both locality A and B. The specimens are deposited at the Department of Geology, Central University of South Bihar, under the catalogue DGCUSB/BB.
Faunal similarity (Dice’s similarity) of the Upper Miocene elasmobranch of Baripada Beds, India and other Upper Miocene deposits of Indo-Pacific regions and the Indian subcontinent.
SYSTEMATIC PALAEONTOLOGY
Order Carcharhiniformes Compagno, 1977
Family Carcharhinidae Jordan & Evermann, 1896
Genus Carcharhinus de Blainville 1816
Carcharhinus amblyrhynchoides Whitley, 1934
(Plate 1, Figures 1–4)
Figures 1–4: Carcharhinus amblyrhynchoides (DGCUSB/BB-115,112), upper antero-lateral, Figures 5–6: Carcharhinus brevipinna (DGCUSB/BB-136), upper tooth, Figures 7–10: Carcharhinus perezi (DGCUSB/BB-140,143), upper lateral, Figures 11–18: Carcharhinus aff. C. plumbeus (DGCUSB/BB-19, 20), upper lateral, Figures 19–24: Carcharhinus sp. (DGCUSB/BB-111, 99), upper teeth, Figures 23–24: Physogaleus hemmooriensis (DGCUSB/BB-23), lower antero-lateral tooth, Figures 25–26: Physogaleus sp. (DGCUSB/BB-45), lower tooth. Figures 27–36 Upper teeth of Glyphis sp. (DGCUSB/BB-113,119, 148, 150, 160). 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35 are in lingual view and 2, 4, 6, 8,10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36 are in labial view.

Material and Horizon: Two isolated upper teeth, DGCUSB/BB-112, 115, from Upper Miocene yellowish shale of Mukurmatia, Odisha, India.
Description: The specimens DGCUSB/BB-112, 115 are medium in size, cusps are erected, relatively narrow and triangular in shape. The central cusp is slightly arched lingually (Plate 1, Figures 1 and 3) and flattened labially (Plate 1, Figures 2 and 4). The crown possesses mesial and distal cutting edges, which are straight and finely serrated (Plate 1, Figures 1 and 4). The heels of the crown are coarser and separated from the finer serrated edges by a distinct notch on both sides. The root is robust, extended mesiodistally and bilobed. A deep transverse nutritive groove is present on the lingual face of the root, dividing the root into two lobes, and the groove penetrates nearly through the basal margin of the root. TH (teeth height): 6.57–7.53 mm; CH (crown height): 3.97–4.78 mm; RW (root width): 9.45–9.91 mm.
Remarks: Carcharhinus amblyrhynchoides (graceful shark) is being reported for the first time from Upper Miocene deposits of Baripada Beds, Odisha. The teeth of C. amblyrhynchoides are known for their narrow, erect crowns, fine serrations on cutting edges, which become coarser at the heel, resembling small cusplets, separating it from other Carcharhinus taxa (Plate 1, Figures 1–4) (Kocsis et al., 2018). The upper teeth of C. amblyrhynchoides are closely comparable to C. limbatus in having a narrower, more erect crown with fine serrations on cutting edges and coarser at the heel, but C. limbatus has symmetrical cusps except for the posterior teeth that are slightly oblique with concave distal edges and finer serrations on the heel (Morey et al., 2008). Earlier, C. amblyrhynchoides was considered identical to C. wheeleri (Compagno et al., 2005). However, the presence of a straight mesial cutting edge and a broader crown base in C. wheeleri makes it different (Kocsis et al., 2018).
C. brevipinna Valenciennes, 1839
(Plate 1, Figures 5 and 6)
Material and Horizon: Isolated upper teeth, DGCUSB/BB-136 from the Upper Miocene yellowish shale of Mukurmatia, Odisha, India.
Description: DGCUSB/BB-136 is a medium-sized tooth having a thin and erect cusp, slightly inclined distally. The edges bear worn-out serrations on both sides. The fairly erected triangular crown is convex on the lingual side and nearly flattened on the labial side. The root is broader than the height of the crown. The heels of the crown bear fine serrations; the root is bilobed, compressed, and a deep nutritive groove divides the two lobes. The basal margin of the root is nearly straight.
Remarks: Sahni and Mehrotra (1981) reported dermal denticles of C. brevipinna from Baripada Beds as C. maculipinnis. The presence of a lingually curled crown, shorter cusp with wider root in C. brevipinna makes it distinguishable from other species of the genus Carcharhinus (Perez et al., 2017). Teeth of C. limbatus closely resemble the upper teeth of C. brevipinna. However, the presence of an asymmetric and short crown, fine serrations on the heel distinguished it from those of C. limbatus (Perez et al., 2017). The extant shark of C. brevipinna is mainly known for its adaptation to warm temperate to tropical waters in the Indo-West Pacific Ocean, Mediterranean Sea and the Atlantic Ocean (Burgess, 2009; Compagno, 1984).
C. perezi Poey, 1876
(Plate 1, Figures 7–10)
Material and Horizon: Two isolated upper teeth DGCUSB/BB-140,143 from the Upper Miocene yellowish shale of Mukurmatia, Odisha, India.
Description: Small-sized teeth, triangular in shape and distally inclined cusp. The cutting edge of the distal side is nearly vertical to concave to some extent. The mesial cutting edge is convex. Both the mesial and distal cutting edges are ornamented with coarse serration. The coarsely serrated heel is separated from the central cusp by a shallow and slightly rounded notch on the distal edge. The crown possesses finer serrations toward the apex, and the serrations are coarser on the heel. The labial face of the teeth is nearly flattened, whereas the lingual surface is curved. The root of the tooth is robust with a transverse groove on the lingual side and a slightly curved root base. TH: 11.12–12.11 mm; CH: 6.19–6.95 mm; RW: 13.56–15.49 mm.
Remarks: This is the first report of this species from the Miocene of the eastern coast of India. The presence of a distally inclined cusp, a coarsely serrated cutting edge on both sides, and a straight distal cutting edge in the teeth of C. perezi makes it distinct from other species of the genus Carcharhinus. C. perezi presents upper teeth, which closely resemble C. obscurus and C. brachyurus. However, C. obscurus has a broader crown, and the presence of a continuous transition from cusp to heel on the mesial side without a distinctive notch and coarser serration at the crown base (Pimiento et al., 2013). In the presence of a wavy mesial cutting edge, clear apical convexity, and a truncated cusp, C. brachyurus differs from C. perezi. The teeth of C. perezi are known from the Pliocene of Tuscany, Italy (Marsili, 2006), the Caribbean Sea, Upper Miocene Gutun Formation of Panama (Carrillo-Briceño et al., 2019; Pimiento et al., 2013; Sánchez-Villagra et al., 2000); Lower Miocene Pirabas Formation of Brazil and Cantaure Formation, Venezuela (Aguilera et al., 2017; Carrillo-Briceño et al., 2016; Reis, 2005), Miocene Lee Creek Mines of North Carolina (Purdy et al., 2001), Alvalade Basin of Portugal (Antunes & Balbino, 2004; Antunes et al., 1999).
Carcharhinus aff. C. plumbeus Nardo, 1827
(Plate 1, Figures 11–18)
Material and Horizon: Four isolated upper lateral teeth DGCUSB/BB-19, 20, 137, 138 from the Upper Miocene yellowish shale of Mukurmatia, Odisha, India.
Descriptions: The specimens (DGCUSB/BB-19, 20) (Plate 1, Figures 11–14) are triangular, small-sized teeth with slightly curved distal cutting edges. However, the mesial edges are nearly convex (Plate 1, Figures 17 and 18) to slightly concave (Plate 1, Figures 11 and 12) near the apex with a truncation. The crown on the lingual side is curved and is flattened labially. Both the cutting edges bear fine serrations, which are coarser towards the heel. The coarser serrations at the distal heel show the presence of secondary serrations. DGCUSB/BB-20 has a deep distal notch, whereas DGCUSB/BB-19 has a shallow distal notch. The roots are partially broken in both specimens, and a nutritive groove is observed at the base of the root. The other two specimens (DGCUSB/BB-137,138) (Plate 1, Figures 15–18) are also triangular in shape with an erected crown and a pointed apex. The lingual side of the crown is convex, and the labial face is flattened. The cutting edge of the distal side is almost straight and nearly perpendicular to the basal margin of the teeth, and a shallow notch separates the central cusp from the heel. The mesial cutting edge is inclined distally and continues to the heel. The cutting edges on both the mesial and distal sides bear moderate serrations. Roots are bilobed, and a strong, prominent nutritive groove is present; the root-crown boundary is curved in the lingual face and straight in labial view.
Remarks: The upper lateral teeth of C. plumbeus previously reported from the Pleistocene of the Ogushi Formation, Japan (Ando et al., 2014), closely resemble the present specimens (DGCUSB/BB-19, 20) in having a posteriorly compressed crown and the presence of secondary serrations on the heel. DGCUSB/BB-137, 138 are closely comparable with the upper teeth of C. plumbeus from the Upper Miocene Gatun, Charges and Chucunaque formations of Panama in having inclined mesial cutting edges, moderately sized uniform serrations, a broadly broad crown and the presence of slightly curved to straight distal cutting edges (also see Carrillo-Briceño et al., 2015; Perez et al., 2017; Pimiento et al., 2013). However, C. plumbeus reported from Panama lacks a notch on either side of the cutting edge (Perez et al., 2017). This is the first report of Carcharhinus aff. plumbeus from the Baripada Beds.
Carcharhinus sp.
(Plate 1, Figures 19–22)
Material and Horizon: Two isolated upper teeth with catalogue numbers DGCUSB/BB-99, 111 from the Upper Miocene greenish to yellowish shale bed of Mukurmatia, Odisha, India.
Description: DGCUSB/BB-99 and 111 are small, triangular-shaped teeth. The crown is thin and pointed with a slight distal inclination. Both the lingual and labial faces of the cusps are slightly curved and convex with finely uniform serration on both the cutting edges, and serrations are finer towards the apex (Plate 1, Figures 19–22). The heels on both the mesial and distal sides bear fine, uniform serrations. The cutting edge is separated from the distal and mesial heels by distinct notches. The roots are incomplete, bilobed, and a prominent median nutritive groove separates each lobe.
Remarks: Certain teeth of Carcharhinus sp. have been reported earlier from the bluish grey shale bed of Mukurmatia, Baripada Beds (Sahni & Mehrotra, 1981; Sharma & Patnaik, 2014 and reference therein). DGCUSB/BB-99, 111 are closely similar to Carcharhinus limbatus reported from the Neogene of Guajira Peninsula of Colombia (Carrillo-Briceño et al., 2019) in possessing strong serrations on both cutting edges of the erect cusp. The upper teeth of C. limbatus have finer serrations on the heel. The specimens are also distinguishable from those of C. longimanus teeth in having a thinner cusp and a shallow notch separating the heel from the crown. The absence of a root in the current specimen makes it difficult to assign it to a particular species.
Genus Physogaleus Cappetta, 1980
Physogaleus hemmooriensis Reinecke & Hoedemakers, 2006
(Plate 1, Figures 23 and 24)
Material and Horizon: Two isolated teeth with catalogue number DGCUSB/BB-23 from the Upper Miocene greenish shale bed of Mukurmatia, Odisha, India
Description: DGCUSB/BB-23 is a small tooth with a narrowly erect crown. The crown is smooth with no visible serration on the cutting edges (Plate 1, Figures 23 and 24). The mesial edge is somewhat concave, wavy, and the cusp is distally inclined. The cusp is again deflected slightly toward the apex on the mesial side. A prominent distal notch separates the slightly curved distal edge from the heel. The distal heel possesses two to three cusplets (Plate 1, Figures 23 and 24). The cusp in the labial face is flattened, and the lingual view of the crown presents a sigmoidal outline and is compressed toward the base of the crown. In the lingual view, the crown-root boundary is curved, and it is straightened in the case of the labial face. The root of the tooth is present with two lobes, a prominent nutritive groove separates each in the lingual view, and the root is convex toward the basal margin. The height of the crown is 2.6 mm, the width of the root is 3.6 mm, and the total height of the tooth is 4.22 mm.
Remarks: This is the first report of the Physogaleus hemmooriensis from the Miocene sequences of the Indian subcontinent. Teeth of Physogaleus hemmooriensis are distinguished from other species of Physogaleus in the presence of non-serrated cutting edges and a distal heel. P. hemmooriensis differs from P. maltzani in having narrower main cusps with a relatively higher crown. Teeth of P. secundus also differ from P. hemmooriensis in having less upright main cusp and more strong roots (see, Maisch et al., 2015; Reinecke & Hodemakers, 2006). P. hemmooriensis can be distinguished from the teeth of P. contortus in having a narrowly extended cusp with smooth cutting edges and the presence of cusplets at the distal heel. Further, the absence of serrations on the mesial heel of P. hemmooriensis separates it from P. latus. P. hemmooriensis has also been described from the Lower Hemmoorian (Behrendorfian, Upper Burdigalian, Lower Miocene) of Werder, Lower Saxony, Germany; Edegem and Antwerpen Sands Members of the Berchem Formation of Belgium, and from the Miste Bed, Aalten Member of the Breda Formation of the Netherlands (also see Reinecke & Hoedemakers, 2006).
Physogaleus sp.
(Plate 1, Figures 25 and 26)
Material and Horizon: Isolated tooth DGCUSB/BB-45 from the Upper Miocene greenish shale bed exposed at Mukurmatia, Odisha, India.
Description: DGCUSB/BB-45 is a small-sized tooth with a narrowly erect crown. The crown is triangular, broader at the base, distally inclined; cutting edges of the cusp are smooth and non-serrated. The mesial side of the tooth is slightly concave to straight; the apex of the cusp is slightly deflected towards the mesial side. The cutting edges on the distal side are slightly curved, and a deep distal notch separates the cusp edge from the distal heel. The distal heel possesses two to three cusplets with secondary serrations. The tooth is slightly convex towards the apical portion of the cusp and flattened near the root on the labial face, and in the lingual view, it is sigmoidal in shape and compressed near the base of the crown. The crown-root boundary is curved; the root is worn out, bilobate, and a deep median nutritive groove divides the roots into two halves, comprising each lobe. The root is concave in labial view, while the basal margin of the root is convex. The height of the crown is 1.64 mm, the width of the root is 2.9 mm, and the total height of the root is 2.61 mm.
Remarks: The present specimen is similar to the tooth of P. hemmooriensis in having non-serrated cutting edges and the absence of coarse serrations on the mesial heel. However, it differs from P. hemmooriensis in having secondary serrations in distal cusplets. The present specimen is also distinct from the P. latus in the absence of serration on the mesial heel. It also differs from Sphryna in being smaller in size, in the presence of a straight edge and cusplets on the distal heel. Galeorhinus, on the other hand, has shorter and more inclined cups compared to P. hemmooriensis.
Genus Glyphis Agassiz, 1843
Glyphis sp.
(Plate 1, Figures 27–36)
Material and Horizon: Upper teeth under the specimen (DGCUSB/BB-113, 119, 148, 150, 160) from the Upper Miocene greenish shale beds, exposed at Mukurmatia, Odisha, India.
Descriptions: The teeth are small with thin, symmetric to asymmetric, triangular crowns. The crown is flat on the labial side, and the lingual face is slightly curved; both the cutting edges present fine and uniform serrations. The basal margins of the roots are curved, and a shallow nutritive groove is visible in the lingual view. DGCUSB/BB-113,119 (Plate 1, Figures 27–30) possess a triangularly extended symmetrical crown with prominent heels. The labial side of the tooth crown is thin and flattened, whereas it is slightly curved on the lingual face. Both distal and mesial cutting edges are serrated. Serrations are fine and uniform, slightly coarser on the heel. DGCUSB/BB-148 (Plate 2, Figures 31 and 32) has a thinly extended crown; the cusp is slightly tilted distally. The cutting edges are finely serrated, curved on the distal side, while it is straight from tip to heel base on the mesial side. DGCUSB/BB-150, 160 (Plate 2, Figures 35 and 36) present asymmetrical cusps; the distal cutting edge is curved, but the mesial edge is also slightly curved. The cusps of the crown are distally inclined, and fine serrations on both the cutting edges are increasingly coarser toward the base.
Remarks: The current specimens are similar to Glyphis teeth described from the Neogene of Borneo, Ambug hill (Kocsis et al., 2018) and Miocene of Myanmar (Shimada et al., 2016). Kocsis et al. (2018) suggested that the upper anterior teeth of Glyphis are characterised by a triangular, symmetric, thin crown with a broader base; lateral and posterior teeth are characterised by a straight or slightly concave mesial cutting edge and an asymmetric crown. Posterior teeth are small with a highly curved distal cutting edge extended to the heel. The lower teeth possess a narrow, slightly curved and elongated crown. The tip of the teeth looks like a spearhead in shape and lacks prominent serrations. DGCUSB/BB-113 and 150 are anterior teeth, whereas DGCUSB/BB-119 and 150 are the lateral teeth, and DGCUSB/BB-160 is a posterior tooth.
Order Rajiformes Berg, 1940
Family Myliobatidae Bonaparte, 1838
Genus Aetobatus Blainville, 1816
Aetobatus cappettai Antunes & Balbino, 2006
(Plate 2, Figures 1–10)
Figures 1–10: Aetobatus cappettai (DGCUSB/BB-180, 181, 182, 183), upper teeth, Figures 11–13: Myliobatis quila (DGCUSB/BB-189), lower median tooth. 1, 4, 11 occlusal view, 2, 5, 7, 9, 12 lingual view, 3, 6, 8, 10, and 13 are in basal view.

Material and Horizon: Isolated teeth of upper dentition catalogued as the specimen numbers DGCUSB/BB-180, 181, 182, 183 from the Upper Miocene greenish shale of Mukurmatia and Itamundia, Odisha, India
Description: Specimens DGCUSB/BB-180, 181 are small, narrow and long teeth of the upper jaw. The teeth are thick, possess a slightly convex low crown, and bear a prominent lateral extremity (Plate 4, Figures 1 and 4). The crown and root are divided by a thin ridge. The crown is thinner than the root. In the basal surface of the root, alternate ridges and grooves, which are of nearly equal width, are visible (Plate 2, Figures 3 and 6). DGCUSB/BB-182 and 183 are incomplete lower teeth; the labial side of the crown is arched and convex, flattened labio-lingually (Plate 2, Figures 7 and 9). The crown surface is low, smooth and curved; the width of the crown decreases towards the lingual edge. The lingual view of the teeth shows the presence of a deep and wide depression separating the crown from the root. The thick root is high; the thickness of the root diminishes towards the lateral ends, and in the labial face, the roots are arched and slightly convex (Plate 2, Figures 7 and 9). The crowns of the lower teeth are comparatively broader than those of the upper teeth.
Remarks: Aetobatus cappettai has been earlier reported from the Alvalade basin (late Miocene), Portugal (Antunes & Balbino, 2006, Figures 1–8), and the Lower Tagus basin of the Middle Miocene deposit, Portugal (Fialho et al., 2019). The lower teeth of Aetobatus cappettai are distinguished from the closely identical lower teeth of Aetobatus arcuatus in having oblique and prominently developed roots in labial and lingual faces, which are less arched, and also differ from Aetobatus irrigularis, which have slightly arched teeth of constant width and roots that are bent lingually (Antunes & Balbino, 2006). Aetobatus cappettai has not been recorded earlier from India, and it is being reported for the first record from the Indian subcontinent.
Genus Myliobatis Dumeril (In Cuvier, 1817: 137)
Myliobatis aquila Linne, 1758
(Plate 2, Figures 11–13)
Material and Horizon: An isolated lower median tooth under the specimen number DGCUSB/BB-189 from the Upper Miocene yellowish shale bed of Mukurmatia, Odisha, India.
Description: The specimen DGCUSB/BB-189 is a median tooth, 27 mm long and 5 mm wide, and the crown is elongated and longer than wide. The occlusal view is nearly flat with finely pitted ornamentation seen at the crown surface, and is nearly rhombic in outline. The teeth are slightly curved in lingual view (Plate 2, Figure 12). The roots are highly worn out, the crown and roots having nearly equal thickness. The root possesses 19 lobes, which are parallel and alternating, each lobe with a groove. The crown in the occlusal view is more laterally developed than in the labio-lingual view. In the occlusal view, the crown is bounded by a slightly concave latero-anterior border. The labial face of the tooth is higher than the lingual face.
Remarks: Earlier, teeth of Myliobatis aquila have been described from the Paleogene of Belgium (Leriche, 1910) and also from the lower Pliocene deposits of Le-Puget-sur-Argens, France (Cappetta & Nolf, 1991). Our specimen is identical to Myliobatis cf. aquila described from the Upper Miocene Alvade Basin, Portugal (Antunes & Babino, 2006, Figures 9 and 10). This is the first record of Myliobatis quila from the Miocene deposits of India.
Family Dasyatidae Jordan, 1888
Genus Dasyatis Rafinesque, 1810
Dasyatis rugosa Probst, 1877
(Plate 3, Figures 1–8)
Figures 1–8: Dasyatis rugosa (DGCUSB/BB-221, 222, 223, 224), Figures 9–15: Dasyatis probsti (DGCUSB/BB-225, 226, 227), Figures 16–18: Dasyatis sp. (DGCUSB/BB-228, 229), Figures 19–22: Himantura menoni (DGCUSB/BB-230, 231), Figures 23–24: Figures 5–8: Taeniurops sp. (DGCUSB/BB-235), Figures 25–30. Pastinachus sp. (DGCUSB/BB-232, 233, 234).1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27 and 29 are in lingual view and 2, 4, 6, 8,10, 12, 14, 16, 18, 20, 22, 24, 26, 28 and 30 are in labial view.

Material and Horizon: Isolated teeth (DGCUSB/BB-221, 222, 223, 224) from the Upper Miocene (Tortonian age) yellowish shale bed of Mukurmatia, Odisha, India.
Description: The specimens are small-sized female teeth, globular in shape, having a low crown and a prominent transverse crest, ornamented with distinct wrinkle-like reticulations. The labial face of the crown is convex and decorated with rutted reticulated enamel. The labial visor is convex and nearly semicircular. The lingual zone of the crown presents prominent vertically folded reticulation, the lingual face of the crown is flattened to nearly concave, the lingual visor is straight to slightly convex, the median lingual lobe is absent or weakly developed, and some lateral teeth have shown a weak axial depression on the lingual face. The roots are bilobate and arched lingually. Each lobe of the root is divided by a prominently deep median furrow. The labial part of the root is depressed, separating the root lobes from the nearly flattened lower part of the labial visor. The triangular shape of the basal surface of the root lobes is flattened and lingually projected.
Remarks: Dasyatis rugosa has not been reported earlier from the Upper Miocene Baripada Beds, eastern coast of India. However, earlier workers described the same teeth of the genus Dasyatis from the Baripada Beds (Sahni & Mehrotra, 1981; Sharma, 2013; Sharma & Patnaik, 2014 and references therein) and the Lower Miocene of Mizoram (Tiwari & Ralte, 2012). Sharma et al. (2021) reported Dasyatis rugosa from the Upper Miocene Tapar locality of Kutch, India. The present specimens are identified as female teeth of D. rugosa as they are having typical female teeth characteristics such as the presence sinuous labial crown margin, broad transverse crest, nearly convex labial face and robust root lobes (Cappetta, 1970; Cicimuri & Knight, 2009; Haye et al., 2008; Reinecke et al., 2005; Sharma et al., 2021). Earlier, D. rugosa had been described from the early Upper Miocene of North Carolina USA, Belgium, Austria, the Netherlands, Switzerland, northern and southern Germany and Portugal (Balbino & Antunes, 2006; Barthelt et al., 1991; Bor et al., 2012; Daxner-Höck et al., 2004; Haye et al., 2008; Kent, 2018; Reinecke & Wienrich, 2009; Reinecke et al., 2011), from the Middle Miocene sequences of Central Paratethys (Szabó & Kocsis, 2016), from the Miocene deposits of France (Brisswalter, 2008; Cappetta, 1970; Vialle et al., 2011), the early Miocene of south west Germany (Probst, 1877); from the Upper Oligocene Chandler Bridge Formation of South Carolina, USA (Cicimurri & Knight, 2009).
Genus Dasyatis Rafinesque, 1810
Dasyatis probsti Cappetta, 1970
(Plate 3, Figures 9–14)
Material and Horizon: Several isolated teeth (DGCUSB/BB-225, 226, 227) from the Upper Miocene yellowish shale bed of Mukurmatia, Odisha, India.
Description: DGCUSB/BB-225, 226 and 227 are small-sized female teeth with robust, bulbous and broad crowns; the crown is higher than the root, and a prominent transverse crest showing reticulated ornamentations is present. Crescentic to subcircular shape with a median labial hollow is present at the labial face of the crown; the labial visor is angular and sharp, and the marginal angles are sharp. The crown is flat or slightly concave in the lingual face, a prominent median lingual lobe divides the lingual marginal face, and the lingual notch of the root is clear and sharp. Roots are bilobate, arched lingually, a prominent deep groove separates the lobes, and the median nutritive groove is clearly visible. The labial part of the root is depressed; the basal part of the root lobes is triangular in outline and nearly flat.
Remarks: Earlier, Dasyatis probsti teeth have been described from the Upper Miocene deposit of Kutch, on the western coast of India (Sharma et al., 2021). This is the first-time report of Dasyatis probsti from the Miocene Baripada Beds of the eastern coast of India. Teeth of Dasyatis probsti differ from morphologically similar teeth of Dasyatis rugosa in having a typical, nearly circular median labial hollow in occlusal view and the presence of a prominent median lingual lobe (Cappetta, 1970; Pollerspöck & Beaury, 2014). It also differs from the teeth of the genus Taeniura, which have a sharp transversal edge, a nearly triangular median labial hollow and the presence of a concave occlusal face (Vialle et al., 2011). D. probsti had also been reported from the Middle Miocene of Mazan, France (Vialle et al., 2011), Serravallian of Vaucluse, southern France (Brisswalter, 2008) and the Langhian of Herault (Cappetta, 1970).
Genus Dasyatis Rafinesque, 1810
Dasyatis sp.
(Plate 3, Figures 15–18)
Material and Horizon: Isolated teeth (DGCUSB/BB-228, 229) from the Upper Miocene greenish to yellowish shale bed of Mukurmatia, Odisha, India.
Description: DGCUSB/BB-228, 229 are small teeth with a more or less globular crown, which are slightly longer than wide. The crown presents a bulging transverse crest, and the lingual zone of the crown is ornamented with longitudinal wrinkles. Labial face of the crown is bulging, and the labial visor is subcircular in outline; a distinct median protuberance is visible, and the marginal angles are distinct. Median lingual ridge divides the crown into two halves (Plate 3, Figures 15 and 17). The marginal-lingual surfaces of the crown are slightly concave. The roots are arched lingually, bilobed, and a broad and deep median nutritive groove divides the root into two lobes; the basal surface of the lobe is triangular in shape. The root in the labial faces is well depressed (Plate 3, Figures 16–18).
Remarks: The present specimens resemble Dasyatis aff. D. probsti (Schultz, 1977; Sharma et al., 2021) in having a prominent median lingual lobe dividing the lingual marginal face, the presence of a clear lingual notch of the root and an angular labial visor. However, they differ from Dasyatis aff. D. probsti in having a weakly developed median labial hollow at the labial face. It is also distinguished from Dasyatis rugosa in its large size and the presence of a prominent median lingual lobe on the lingual face. The present specimens have a globular crown with a transversal crest and a lingual zone of the crown that is striated by numerous fine vertical ridges, placed under Dasyatis sp.
Genus Himantura Müller & Henle, 1837
Himantura menoni Sahni & Mehrotra, 1981 (Marramà et al., 2019)
(Plate 3, Figures 19–22)
Material and Horizon: Isolated teeth (DGCUSB/BB-230, 231) from the Upper Miocene greenish to yellowish shale bed of Mukurmatia, Odisha, India.
Remarks: Himantura menoni has been earlier reported from Baripada Beds (Sahni & Mehrotra, 1981; Sharma & Patnaik, 2013) as Dasyatis menoni. Andrianavalona et al. (2015) reassigned Dasyatis menoni under the genus Himantura. Sharma et al. (2021) described the teeth of Himantura menoni from the Tapar locality of Kutch. Marramà et al. (2019) reassigned Dasyatis menoni (Sahni & Mehrotra, 1981) as Himantura menoni. Teeth of Himantura are also reported from the Miocene deposit of Nosy Makamby locality of Mahajanga Basin, Northwestern Madagascar (Andrianavalona et al., 2015). Many of the previously described Dasyatis teeth need taxonomic revision, as there is a possibility that many of the pre-Miocene teeth which were hitherto described under the genus Dasyatis may be representative of other genera, including Himantura (also see Cappetta, 2012).
Family Dasyatidae Jordan, 1888
Genus Taeniurops Garman, 1913
Taeniurops sp. (Probst, 1877)
(Plate 3, Figures 23 and 24)
Material and Horizon: Several isolated female teeth (DGCUSB/BB-235, 236, 237) from the Upper Miocene greenish to yellowish shale bed of Mukurmatia, Odisha, India.
Description: DGCUSB/BB-235, 236 and 237 are small teeth with a transverse ridge sharply dividing the crown into labial and lingual faces. The transverse ridge is smooth and sharp with less curvature. The labial face shows an elongated median hollow; the labial visor is semicircular, less arched than the lingual visor. The crown lacks any ornamentation in the labial face. On the lingual side, the crown is nearly concave, smooth, and a prominent median lingual lobe is present. The labial visor is smooth and slightly curved in the lower part. The root is bilobed, and a broad and deep median groove separates each lobe. The root is depressed in the labial part, basal faces of the root are nearly flat and triangular in outline.
Remarks: The present specimens represent the first report of the genus Taeniurops from the Miocene deposits of the Indian subcontinent. Earlier, teeth of T. cavernosus have been described from early Miocene deposits of Germany (Reinecke et al., 2011; Sach, 2016) as Raja cavernosa (Probst, 1877), Dasyatis cavernosa (Barthelt et al., 1991); from Portugal as Dasyatis cavernosa (Antunes et al., 1981), from Switzerland as Trygon cavernosus (Fischli, 1930) and the USA (Case, 1980). The specimens are diagnosed as female teeth as they have high crowns which are non-cuspidate and nearly quadrangular to irregular in outline. In case of the male teeth, the crowns are raised and cuspidate (also see Laurito, 1999). The genus Taeniurops is very difficult to differentiate from Dasyatis (Cappetta, 2012); however, it differs from Dasyatis in having a distinct depression in the labial visor, which is bordered by a sharp crest, which is lacking in Dasyatis (Villafaña et al., 2020).
Genus Pastinachus Rüppell, 1829
Pastinachus sp.
(Plate 3, Figures 25–30)
Material and Horizon: Several isolated teeth (DGCUSB/BB-232, 232, 234) from the Upper Miocene greenish to yellowish shale bed of Mukurmatia, Odisha, India.
Description: DGCUSB/BB-232, 232, 234 are small, teeth are massive, crowns are thick and well developed, rhombic to hexagonal in shape, as a characteristic of teeth of the upper jaw. The crown is elevated wider than in length, the lingual zone of the crown is smooth and lacks any visible ornamentations; the marginal angle of the crown is sharp. The lingual face of the crown is depressed, and the labial face is nearly flat with a distinct transverse crest. The roots are robust, nearly vertical, bilobed, and a wide nutritive groove divides each lobe into two halves. The base of the root is smooth, flat and triangular (Plate 3, Figures 26) to pentagonal in outline (Plate 3, Figures 30).
Remarks: Adnet et al. (2019) reassigned Dasyatis sylvestris, which was earlier described from the limestone of Baripada Beds (Sahni & Mehrotra, 1981; Sharma & Patnaik, 2013), to the genus Pastinachus. Teeth of Pastinachus were also described from the Upper Miocene deposits of Tapar section, Kutch in the western coast of India (Sharma et al., 2021), Neogene of Taiwan (Uyeno, 1978), Lower Miocene Marada Formation of Jabal Zlatan, Libya (Argyriou et al., 2015), and the late Middle Eocene of Central Tunisia (Adnet et al., 2019).
DISCUSSION
Faunal Composition of the Baripada Beds
As mentioned above, the Baripada Beds have yielded a variety of fossils, including vertebrates, invertebrates and microfossils. The recovered vertebrate fauna of Baripada Beds comprises sharks, batoids, teleosts, and mammals (also see Sahni & Mehrotra, 1981; Sharma & Patnaik, 2013, 2014). The vertebrate remains are found in association with certain fragmentary remains of bryozoans, balanids, echinoids, molluscs and microfossils (foraminifers and ostracoda). Among the elasmobranchs, Carcharhinidae is the most dominant family and comprises seven genera, including Carcharhinus, Galeocerdo, Glyphis, Negaprion, Physogaleus, Rhizoprionodon and Scoliodon. Family Lamnidae comprises three genera (Carcharodon, Isurus and Lamna), and the other families Hemigalidae (Hemispristis), Trakidae (Galeorhinus), Sphyrnidae (Sphyrna), Alopiidae (Alopias), Odontaspididae (Odontaspis) and Squalidae (Squalus) are represented by one genus each. The batoid fishes are represented by eight families that include Dasyatidae, Myliobatidae, Rhinopteridae, Rajidae, Gymnuridae, Narcinidae and Pristidae. At the generic level, four genera of Dasystidae (Dasyatis, Himantura, Pastinachus and Taeniurops), two genera of Myliobatidae (Myliobatis and Aetobatus) and two genera of Rhinobatidae (Rhinobatos and Rhynchobatus), and one genus of each remaining families Rhinopteridae (Rhinoptera), Rajidae (Raja), Gymnuridae (Gymnura), Narcinidae (Narcine) and Pristidae (Pristis) have been recorded (Mehrotra, 1979; Sahni & Mehrotra, 1981; Sharma & Patnaik, 2014 and reference therein) (see Supplementary Sheet 1). Besides, the teleost, bony fishes are represented by six families and six genera, namely Sparidae (Diplodus, Sparus and Pagrus), Denticidae (Dentex), Scrombridae (Cybium), Sphyraenidae (Sphyraena) (Mehrotra, 1979; Sahni & Mehrotra, 1981; Sharma & Patnaik, 2014). The three mammalian families with three genera, namely Suidae (Tetraconodon intermedius, Potamochoerus), Bovidae (Gazella) and Rhinoceratidae (gen. indet.) were also reported (Chauduri, 1958; Sharma, 2013; Sharma & Patnaik, 2013).
The foraminifera (Anomalina, Ammonia, Anomalina, Bolivina, Buliminella, Cibicides, Discorbis, Elphidium, Globorotalia, Globigerina, Lagena, Nodosaria, Nonion, Rotalia, Textularia, Uvigerina and Valvulineria) have been described from the same beds (Bhalla & Dev, 1988; Bhaumik et al., 2017; Chatterjee, 1973; Sarma, 1956; Tewari & Awasthi, 1960). Among the molluscs, the reported taxa are namely Ostrea gajensis, Ostrea cf. O. papyracea, Trigonia, Cardida, Nucula, Cardium protosubrogosum and Nucula virgo, Barbatia, Astarte, Cardita, Cardium, Venus, Cardium, Paphia, Leda, Pectene cf. P. kokenianus, Natica cf. N. gracillor Crassatella, Turitella cf. T. lydekkeri, Solen, Thracia, Calyptreae aff. C. rugoss, Ficus (ficus) conditus, Torinia, Cypraea, Vermitus, Ramella, Olivia, Conus: Ginglymostoma sp. (Eames, 1936; Jena, 1942; Modak, 1952; Mohanty, 1966, 1980; Sharma, 1956).
Palaeoenvironment
The dominant family Carcharhinidae among the elasmobranchs in the Upper Miocene of Baripada Beds shows a wide range of environmental adaptations. Majority of the species of this family including Carcharhinus amblyrhynchoides, C. brevipinna, C. falciformis, C. plumbeus, C. longimanus, C. macloti, C. leucas, C. perezi, C. limbatus, C. sorrah as well as Galeocerdo cuvier, Glyphis, Rhizoprionodon, Scoliodon laticaudus, Sphyrna zygaena, Sphyrna lewini, Alopias vulpinus, Isurus oxyrinchus, Carcharodon carcharias and Squalus are found today as extant species in the Indian Ocean (Akhilesh et al., 2014; also see Table 2). The majority of species of the genera Carcharhinus and Sphyrna are found adapted to a variety of habitats ranging from inshore to offshore water under tropical, subtropical, and temperate conditions, and they are usually cosmopolitan in distribution (Cappetta, 2012; Compagno, 1984; Sharma et al., 2021). Carcharhinus sharks such as C. brevipinna, C. falciformes and C. leucas are mostly circum-tropical in distribution, and their habitats extend from coastal shallow marine to offshore environments of tropical to temperate conditions (Cappetta, 2012; Compagno, 1984; Pollerspöck & Straube, 2019; Sharma et al., 2021; Singh et al., 2022; Villafaña et al., 2020). The extant shark of C. brevipinna is mainly known for its adaptation to warm temperate to tropical waters of the Indo-West Pacific Ocean, Mediterranean Sea and the Atlantic Ocean (Burgess, 2009; Compagno, 1984). The extant species of C. amblyrhynchoides are known from the present-day tropical region around Borneo and the Indo-West Pacific Ocean (Compagno et al., 2005). They are commonly adapted to inshore and known as coastal pelagic species (Compagno & Niem, 1998). C. leucas is also well adapted to a freshwater environment (Compagno, 1984; Cook et al., 2010) (Figure 2) with the depths of 30 m (Carrillo-Briceño et al., 2016; Pimiento et al., 2013). Extant species Carcharhinus brevipinna, C. perezi and Negaprion are also found to be adapted to continental, coastal and insular shelves with variable bathymetry ranging from 0 to 200 m (Weigmann, 2016). The extant species of C. perezi is known to inhabit the shallower part of the continental shelf up to a depth of 30 m in tropical water (Compagno, 1984). Carcharhinus brevipinna occurs in warm tropical to temperate waters in the Mediterranean, Atlantic and Indo-West Pacific oceans (Burgess, 2009; Compagno, 1984; Perez et al., 2017; Reiner, 1996)—the extant species Carcharhinus aff. C. plumbeus is commonly found in shallow-water shelves to offshore seas at depths less than 280 m, but standard within the depth of 20–55 m (Compagno, 1984; Compagno et al., 2005). Carcharhinus sorrah mainly inhabits tropical waters at a depth of 140 m (Compagno, 1984). Carcharhinus falciformes are well adapted to open ocean and occasional narrow shelves and coastal regions, with the depths ranging from 18 to 500 m (Compagno, 1984; Pimiento et al., 2013; Purdy et al., 2001) and C. perezi up to the depth of 380 m (Weigmann, 2016). Galeocerdo cuvier occurs in the lagoons, river estuaries and river mouths (Sharma et al., 2021). Negaprion and Pristris are most common dwellers of littoral, mostly shallow, brackish water estuaries, lagoons and intertidal through offshore waters (Sharma, 2013). Negaprion brevirostris is known to be well adapted to estuarine, brackish, and marine waters and frequents mangrove and coral reefs, shallow marine environments of tropical and temperate conditions (Compagno, 1984). This reef-associated, oceanodromous species is also occasionally found in the open ocean (near the water surface) for migration purposes (Compagno, 1984; Compagno et al., 2005; Kent, 1994), generally occurring at depths ranging from 0 to 92 m (Riede, 2004), in subtropical conditions (Florida Museum of Natural History, 2005). Sharks of the genera Alopias, Sphyrna, Carcharadon, Galeocerdo and Carcharhinus are also known to have adapted well to open ocean and normal salinity conditions (Cappetta, 2012; Compagno, 1984; Katuna et al., 1997; Sanders et al., 1982; Sharma, 2013; Weems & Sanders, 1986). Carcharhinus perezi was earlier considered to be mainly restricted to the Caribbean Sea (also see Compagno, 1984; Pollerspöck & Straube, 2019; Villafaña et al., 2020). The sharks of the genus Himantura occur in the Indo-Pacific region, specifically in the coastal Miocene deposits of India and Madagascar (Andrianavalona et al., 2015; Borsa et al., 2013; Sahni & Mehrotra, 1981; Singh et al., 2022; Weigman, 2016).
Upper Miocene Chondrichthyes from Baripada Beds with respect to global status and present-day distribution in the Indian Ocean (also see, Akhilesh et al., 2014; Mondal et al.,2009; Sahni & Mehrotra, 1981; Sharma, 2013; Sharma & Patnaik, 2013, 2014; Weigmann, 2016). The dagger symbol (†) indicates extinct taxa/species.

Paleoenvironment distribution of the Upper Miocene fauna of Baripada Beds, Orissa (Odisha), India. Dagger symbol (†) indicates extinct taxa/species (Source: Cappetta, 2012; Carrillo-Briceño et al., 2015, 2016; Compagno, 1984; Compagno et al., 2005; Perez et al., 2017; Pimiento et al., 2013; Sahni & Mehrotra, 1981; Sharma & Patnaik, 2013, 2014; Sharma et al., 2021; Singh et al., 2022; Vialle et al., 2011 and references therein).
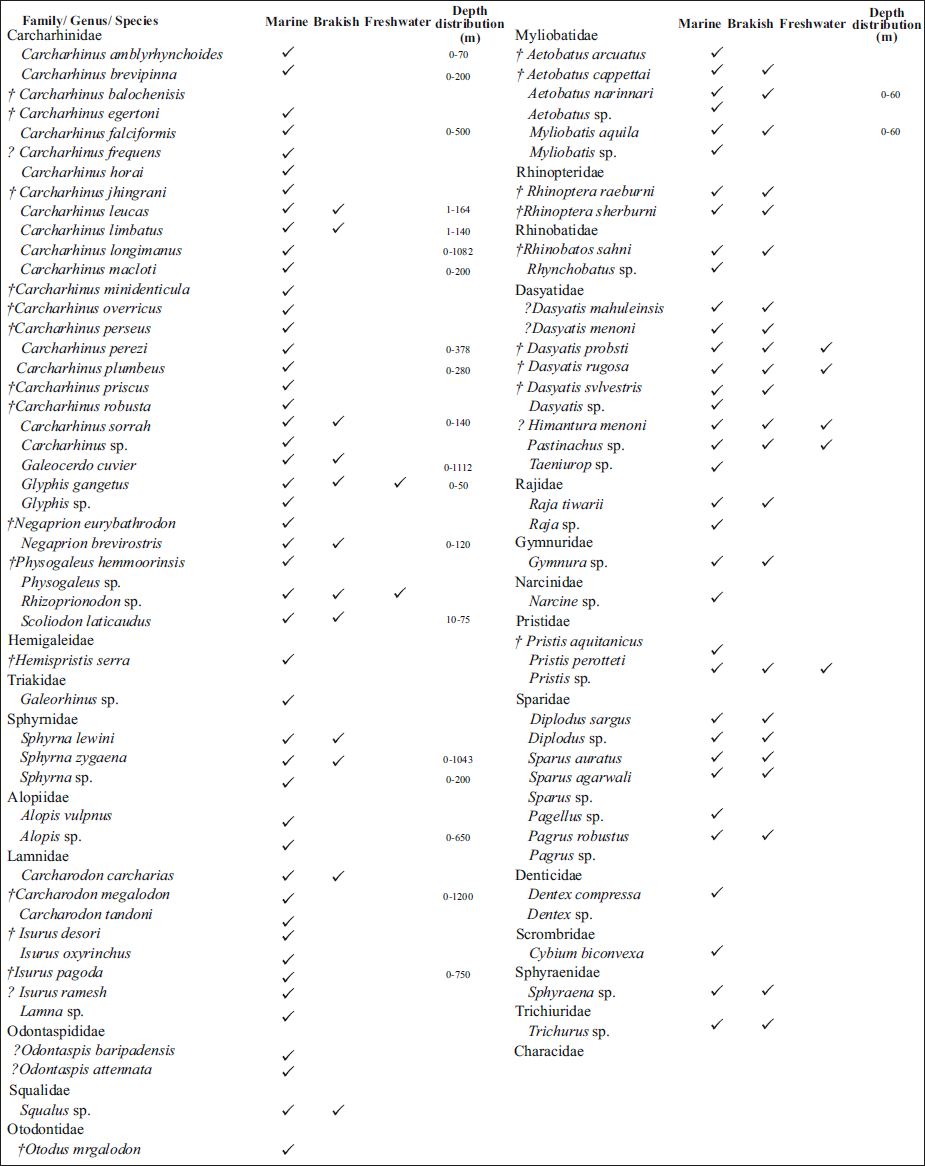
The batoids such as Himantura sp., D. probsti, Dasyatis rugosa, Pastinachus sp., and Pristis sp. were considered to be adapted to a wide range of habitats extending from coastal, shallow marine, inshore, estuaries to freshwater environments (Adnet et al., 2019; Compagno et al., 2005; Sharma et al., 2021; Vialle et al., 2011) (Figure 2). The cosmopolitan species Aetobatus narinari is generally found within the neritic zone in warm tropical to temperate-warm shallow-water environments (Compagno & Last, 1999). Taeniurops cavernosa was considered to be a benthic organism that thrives on the soft bottoms of the shallow marine platform. Himantura is adapted to a variety of environments ranging from tropical, open shallow subtidal environments to coastal, brackish waters, estuaries, and the sandy bottoms of large rivers (Aguilera et al., 2017; Andrianavalona et al., 2015; Kocsis et al., 2018; Marzullo et al., 2011). The species such as Aetobatus narinari, Myliobatis aquila and certain genera of Dasyatis, Himantura, Pastinachus, and Rhynchobatus are also found in the shallow marine, bays, estuaries and river mouths of the present-day Indian Ocean (also see Akhilesh et al., 2014; Compagno, 1984). Certain species of the genus Pastinachus are known for their regular migration between freshwater and the shallow-water coastal seas and frequently migrate towards estuaries, fluvial and freshwater environments (Adnet et al., 2019). The presence of fragmented mammalian remains, teleost fishes and batoids points towards an environment which is very near a fluvial depositional setup (river mouth) with estuaries, very close to the coastal, brackish to very near-shore areas under tropical to subtropical humid conditions.
Based on the occurrence of foraminifera, Bhaumik et al. (2017) suggested that the limestone unit of the Baripada Beds was deposited in the neritic zone (probably within 40 m depth) in warm, tropical, open ocean conditions during the late Miocene marine transgression. Some foraminiferal genera from Baripada Beds, such as Ammonia, prefer to inhabit neritic to upper bathyal zone/estuarine/marsh/tidal flat/subtidal/near-shore areas, deltaic to upper intertidal zone (Bhaumik et al., 2017; Debenay et al., 1998; Hippensteel & Martin, 1999; Murray, 2006; Xue, 2014). Species of the genus Nonionella occur abundantly in the deltaic to neritic zone (Barmawidjaja et al., 1992; Duffield et al., 2015; Murray, 2006; Sabbatini et al., 2010). Even species of the Cibicides group (deep water-well oxygenated species) are also found associated in shallow marine environments (Braga et al., 2012; Majewski & Gazdzicki, 2014; Murray & Elve, 1999). The genus Hanzawaia is considered to be adapted to well-oxygenated coastal shelf areas of India (Kathal & Singh, 2010; Saidova, 2007), and they are found commonly in association with shallow benthic foraminifera in muddy sand of inner shelf to oceanic margins at warm environments (also see Bhaumik et al., 2017; Murray, 2006). The Baripada Beds show scarcity of planktic foraminiferal specimens and records of very few of them (Globigerina falconensis, G. menardii, G. woodi and N. continousa; Bhaumik et al., 2017), indicating the environment of deposition in warm tropical conditions (Kennett & Srinivasan, 1983). The recently recovered fish fossil assemblages in association with other vertebrates and foraminifera suggest a depositional environment of shallow marine, near-shore coastal environment in neritic condition connected to the open ocean for the Baripada Beds (also see Bhaumik et al., 2017; Sharma & Patnaik, 2014).
Palaeobiogeography
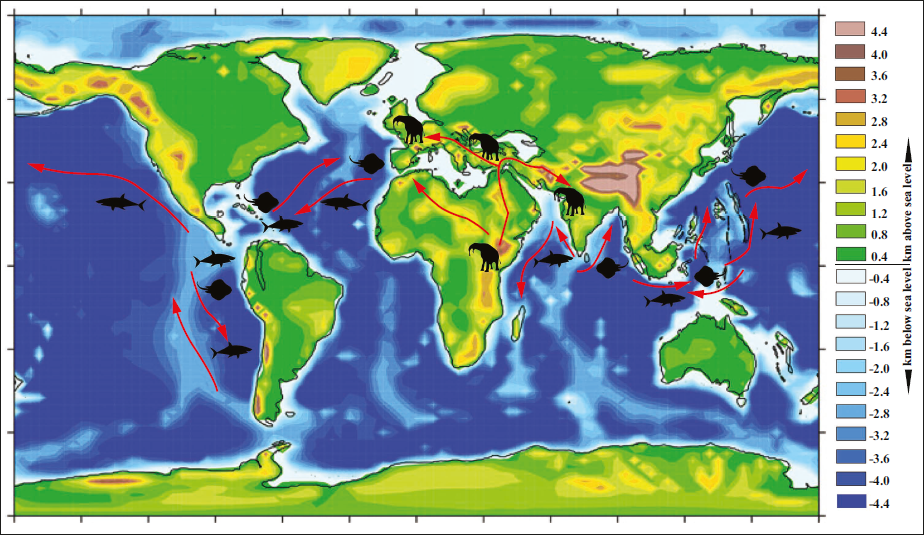
The diversification of Miocene elasmobranchs of the Indian subcontinent and their paleobiogeographic significance were discussed earlier (Modal et al., 2009; Sahni & Mitra, 1980; Sharma et al., 2021; Singh et al., 2022, 2024). The intense diversification of elasmobranch fauna starting from the Eocene-Oligocene epochs was considered to be constrained geographically to the Mediterranean, Atlantic, some parts of the Eastern Tethys and Central Asia (Adnet et al., 2007 and references therein). The Lower Miocene marine fauna of India, comprising the Mediterranean-Tethyan paleogeographic province, shows a similarity index towards the Red Sea and Mediterranean Sea (see Sharma et al., 2021). It indicates the seaway connectivity between the Indian Ocean, the Mediterranean Sea and the Tropical Atlantic-Pacific through the Tethyan Sea and the Isthmus of Panama was open to the migration of marine taxa (Kathal & Bhalla, 1996; Piller et al., 2007; Rögl, 1999; Sharma et al., 2021; Singh et al., 2022). The subsequent tectonic events resulted in the formation of the Gomphotherium Landbridge, a land connection of Africa, Eurasia and the Arabian Plate in the late Burdigalian ~19 Ma (Rögl, 1998). This corridor interrupted the migration of marine fauna, and hence a breakdown in the similarity of marine fauna is observed between the Indian Ocean and the Mediterranean (Harzhauser, 2007; Harzhauser et al., 2002; Sharma et al., 2021; Singh et al., 2022). Further, this paleobiogeographic re-adjudgment is coeval with the late Aquitanian primary biogeographic separation between Mesopotamia, Arabia and northwestern India (Adnet et al., 2007; Harzhauser, 2007; Harzhauser et al., 2002). Singh et al. (2022) also observed a decrease in the similarity index from Lower to Middle Miocene elasmobranch taxa. However, still good affinity may be due to intermittent migration of marine fauna between the Indian Ocean and the Mediterranean region through short-term reopening of Tethyan seaways during ~15–14 Ma (Harzhauser, 2007 and references therein) before the final closure. The Middle Miocene elasmobranchs from the Chhassra Formation of Kutch on the west coast of India exhibit a closure in faunal affinity towards the Pacific Ocean, indicating a gradual shift in migration pattern (Singh et al., 2022). Likewise, the formation of a permanent land bridge between Africa and Eurasia facilitated the land route for the terrestrial fauna to migrate and diversify across two landmasses (also see Harzhauser, 2007; Kapur et al., 2019; Patnaik, 2016; Rögl, 1999; Singh et al., 2022) (Figure 3).
Paleobiogeographic map of the late Miocene Elasmobranchs and Deinotherium/Proboscideans? (Modified after: Steinthorsdottir et al., 2021).
In the present study, the generic level elasmobranch faunal correlation of Upper Miocene deposits of Baripada Beds shows a closer similarity towards elasmobranch fauna from the Upper Miocene Circum-Pacific region (Figure 3 and Table 1). It is observed that the Baripada faunas are closely similar to those from Borneo (62%), Panama (50%), Peru (40%), Japan (35%), Chile (29%), Ecuador (29%) (see Table 1). Similarly, Mondal et al. (2009) observed a higher affinity of the Upper Miocene elasmobranch genera of the Baripada Beds with those of the fauna of the Pacific and Atlantic Oceans. The low faunal affinity at the species level suggests that there must have been faunal endemism in the Indian Ocean (Mondal et al., 2009). Poor faunal similarities in global counterparts at the species level, but a high similarity index in the subcontinent, suggest the faunal endemism in the Indian Ocean (Mondal et al., 2009; Sharma & Patnaik, 2014). The Miocene elasmobranchs from Baripada Beds, India, exhibit a close similarity with elasmobranchs from the Miocene deposits of Kutch basin, Bhavnagar, Mizoram and South Garo Hills District of Meghalaya (Iangrai & Kumar, 2025; Mondal et al., 2009; Sahni & Mehrotra, 1981; Sharma & Patnaik, 2014; Sharma et al., 2021; Singh et al., 2022, 2024) (Table 1). The elasmobranch faunal similarity between the Upper Miocene Baripada Beds on the eastern coast and Tapar sites of Kutch on the western coast of India shows inter-migration of the elasmobranchs from the Bay of Bengal to the Arabian Sea (Sharma et al., 2021). Many of the elasmobranch fauna from the Baripada Beds also exist as present-day living species at the global level, including the Indian Ocean (Table 2). It is observed that the change in faunal migration from the Indian Ocean during the Upper Miocene is evidenced by this close faunal affinity towards the Circum-Pacific region and also very low faunal affinity to those from Portugal (13%) in the Mediterranean-Atlantic region (also see Table 1).
CONCLUSIONS
The present article records additional elasmobranch species, namely Carcharhinus amblyrlynehoides, C. brevipinna, C. perezi, and Carcharhinus aff. C. plumbeus, Physogaleus hemmooriensis, Physogaleus sp., Aetobatus cappettai, and Taeniurops sp. for the first time from the Baripada Beds. Out of these, Carcharhinus amblyrhynchoides, C. brevipinna, Physogaleus hemmooriensis and Aetobatus cappettai are the first reports from the Miocene deposits of the Indian subcontinent. The records of Upper Miocene sharks such as Carcharhinus brevipinna, C. macloti, C. perezi, Carcharhinus aff. plumbeus, Carcharhinus cf. priscus, Carcharhinus cf. sorrah, Carcharhinus sp., Hemipristis serra, Physogaleus hemmooriensis, Physogaleus sp., Glyphis sp., Rhizoprionodon sp., Galeorhinus sp., Sphyrna sp., Alopias sp., and Odontaspis baripadensis and associated foraminifers and molluscs suggest a coastal, lagoonal, near-shore to outer-shelf environment of deposition for the Baripada Beds.
The batoids from Baripada Beds, consisting of Aetobatus cappettai, Aetobatus arcuatus, Aetobatus cf. narinari, Myliobatis quila, Myliobatis sp., Dasyatis probsti, Dasyatis rugosa, Pastinachus sp., Himantura menoni, Taeniurops sp. Raja tiwarii, Rhynchobatus sp., and Pristis sp., which are associated with previously recorded teleost fishes, reptiles, and mammals, indicate an environment closely resembling a fluvial depositional setup (river mouth) with estuaries, situated near the coast, and characterised by brackish to very near-shore conditions under tropical to subtropical humid conditions. Presently reported fossil data from Baripada Beds in association with previously described vertebrates, invertebrates, and foraminifera suggest that these Upper Miocene fossil-bearing beds were deposited in a shallow marine, near-shore coastal environment under neritic conditions connected to the open ocean.
The elasmobranchs from the Baripada Beds have close similarity to those of the Miocene elasmobranchs from the Kutch basin, Bhavnagar, Piram Island, Mizoram. The close faunal affinity of the Upper Miocene Baripada Beds on the eastern coast of India with the Upper Miocene Tapar sites of Kutch shows inter-migration of the elasmobranchs from the Bay of Bengal to the Arabian Sea. The generic level faunal correlation of Upper Miocene deposits of Baripada shows a close affinity to those of different Upper Miocene localities of the Circum-Pacific region. It is observed that the most similar faunas are from Borneo (62%), Panama (50%), Peru (40%), Japan (35%), Chile (29%), Ecuador (29%) (see Table 1). The changes in the faunal migration from the Indian Ocean during the Upper Miocene are evidenced by this close faunal affinity towards the Circum-Pacific region, and also a drastic decline in the faunal affinity with those of the Mediterranean-Atlantic region.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: KMS thanks the Science and Engineering Research Board, India, for the research grants (sanctioned no. CRG/2021/004627). NA Singh is thankful to UGC for providing a JRF for pursuing a PhD.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.