
Abstract
Our previous high-throughput sequencing indicated that rno-miR-1298 was down-regulated in ischemia-reperfusion model of rat. However, little is known about the function and molecular mechanism of rno-miR-1298 in rat tumor cell. In this study, rno-miR-1298 was detected to be significantly down-regulated in rat tumor C6 cells. Moreover, overexpression of rno-miR-1298 obviously inhibited the proliferation and induced apoptosis in C6 cells. SET domain containing 7 (SETD 7) was identified to be a target of rno-miR-1298 using bioinformatics and luciferase reporter assays. Overexpression of rno-miR-1298 markedly reduced the expression of SETD 7 at protein level. Knockdown of SETD 7 also suppressed proliferation and promoted apoptosis in C6 cells. It was indicated that rno-miR-1298 affected cell proliferation and apoptosis of rat tumor cells by targeting SETD 7. Thus, the newly identified miR-1298/SETD 7 expands the elaboration of the mechanisms of the development and progression of tumors and may provide therapeutic target for tumors of nervous system.
Introduction
MicroRNAs (miRNAs) are a class of non-coding small RNAs that are 19–25 nucleotides long. Generally, miRNAs negatively regulate gene expression by imperfect pairing with the 3′-untranslated region (3′-UTR) of target messenger RNAs (mRNAs). 1 Increasing evidences have reported that miRNAs play critical roles in a wide range of physiological and pathological processes.2–5 The most typical example is lin4, which regulates lin-14 translation via an antisense RNA–RNA interaction with the 3′-UTR of lin-14 mRNA, causing a temporal decrease in LIN-14 protein starting in the first larval stage in Caenorhabditis elegans. 6
Additionally, miRNAs are dysregulated and could function as oncogenes or tumor suppressors in most tumor types. The first demonstration of a link between miRNA genes and cancer was miR-15/16, which was found to be frequently deleted or down-regulated in chronic lymphocytic leukemia. 7 To date, more and more miRNAs have been identified to be dysregulated and could affect various function of tumor cell in nervous system. The expression of miR-183 was found significantly higher in glioma tissues, and high expression of miR-183 was significantly correlated to larger tumor size, higher WHO grade, and worse Karnofsky performance score. 8 The expression of miR-383 was down-regulated in human glioma cells U251 and U87 and was negatively related to the pathological grading of glioma. 9 These findings indicate that miRNAs play important roles in tumorigenesis.
Recently, our high-throughput sequencing analysis demonstrated that rno-miR-1298 was obviously down-regulated in ischemia-reperfusion model of rat. 10 However, the biological roles of rno-miR-1298 have not yet been elucidated. In this study, we are aimed at investigating the expression level of rno-miR-1298 in rat C6 cells and the biological functions of rno-miR-1298 on tumor cells and try to further elaborate the molecular mechanisms of rno-miR-1298.
Materials and methods
Cell lines
Rat glioma C6 cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM; Gibco, Grand Island, USA) supplemented with 10% fetal bovine serum (FBS; Gibco). Penicillin (100 U/mL) and streptomycin (100 µg/mL) were added in culture medium. The cells were maintained in an incubator with a humidified atmosphere with 5% CO2 at 37°C.
Quantitative real time polymerase chain reaction for rno-miR-1298
Total RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad, USA) according to the manufacturer’s instructions. Reverse transcription was carried out using M-MLV reverse transcriptase (Promega, Madison, USA). rno-miR-1298 primer was purchased from RiboBio (Guangzhou, China). Quantitative real time polymerase chain reaction (qRT-PCR) was performed in a Roche Real time PCR system (LightCycler480) using SYBR premix ex taq reagents (RiboBio, Guangzhou, China). The two-step polymerase chain reaction (PCR) procedure was as follows: 95°C for 30 s, followed by 40 cycles of denaturation at 95°C for 5 s and annealing/extension at 60°C for 30 s. U6 was used as internal control for rno-miR-1298, and the relative expression level was calculated by ΔCt (ΔCt = Experimental Ct − Control Ct) and 2−ΔΔCt method. The experiment was performed for three times.
MTT for cell proliferation
The C6 cells were seeded into six-well plates at a density of 40% and incubated overnight and infected with lentivirus plasmid GV369 fused enhanced green fluorescent protein (EGFP) as negative control (NC) or lentivirus plasmid of rno-miR-1298 (Genechem, Shanghai, China). After 48 h of infection, the infected cells were seeded into 96-well plates at 2000 cells per well and cultured for 1, 2, 3, 4, and 5 days. In total, 20 μL of MTT reagent (5 mg/mL) was added into the culture medium. Following incubation for 4 h, the MTT/medium was removed and 100 μL of dimethyl sulfoxide (DMSO) was added. The absorbance values were measured at 490 nm using an spectrophotometric plate reader (Bio-Tek, Winooski, USA). Each time point was repeated in five wells and the experiment was independently performed for three times.
Flow cytometry for cell apoptosis
Apoptosis assay of C6 cells was evaluated using an Annexin V-APC apoptosis detection kit (eBioscience, Santiago, USA). After 48 h of infection, the infected cells were collected and washed twice with phosphate-buffered saline (PBS). The cells were re-suspended with 200 μL binding buffer at a density of 5 × 105 and 10 μL of Annexin V-APC was added. The samples were incubated for 5 min at room temperature in the dark. The cell apoptosis was detected using a flow cytometer FACSCalibur (BD, USA). The experiment was performed for three times.
Prediction of rno-miR-1298 targets
Putative targets of rno-miR-1298 were predicted using three algorithms, Target Scan (http://www.targetscan.org/), miRanda (http://microrna.sanger.ac.uk/), and miRDB (http://mirdb.org/miRDB//).
Dual-luciferase reporter assay
Luciferase reporter assay was performed using the Dual-Luciferase Reporter Assay System (Promega, WI, USA) according to the manufacturer’s instructions. Cells were plated in 24-well plates at 1 × 105. When the confluence of cells was 60%, wild-type plasmid, mutant reporter plasmid, or negative control plasmid (termed as WT, MT, or NC) were co-transfected into cells with 100 nM rno-miR-1298 and Renilla plasmid using X-tremegene HP (Roche, Basel, Switzerland). After 24 h of transfection, reporter assays were performed using the dual-luciferase assay system. Firefly luciferase activity was normalized using the corresponding Renilla luciferase activity. All experiments were performed at least three times.
siRNA knockdown
Rat SET domain containing 7 (SETD7) siRNAs (Setd7-rat-803) and NC were purchased from Sangon Biotech (Shanghai, China). The sequence of SETD7 siRNAs was GGACCCAAUACUGUCAUGUTT and ACAUGACAGUAUUGGGUCCTT. The C6 cells were cultured for 24 h, and SETD7 siRNAs were transfected using X-tremegene HP (Roche, Basel, Switzerland) following manufacturer’s protocol. Subsequently, MTT and flow cytometry assays were carried out.
Western blotting
Cells were washed with ice-cold PBS and lysed in radioimmunoprecipitation assay (RIPA) buffer (Beyotime, Shanghai, China) containing protease inhibitor. The supernatants were collected, and the protein concentration was measured using a BCA Assay Kit (Beyotime, Shanghai, China). A total of 50 µg of total proteins was separated on 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred to polyvinylidene fluoride (PVDF) membranes (Millipore, MA, USA). The membranes were blocked with 5% skimmed milk for an hour at room temperature. After washing three times, the membranes were incubated with a primary anti-SETD7 antibody (1:1000; Abcam, Cambridge, UK) at 4°C overnight. Subsequently, the membrane was washed three times and incubated with an anti-rabbit secondary antibody (1:2000; BZSGB Technology, Beijing, China). Thereafter, the membranes were detected by chemiluminescence with ECL Detection Reagent (Beyotime, Shanghai, China). An anti-β-actin antibody (1:1000; BZSGB Technology, Beijing, China) was used as the loading control.
Statistical analysis
Data are presented as mean ± standard error of mean (SEM). All statistical analysis was performed using GraphPad Prism 5. Two-tailed Student’s t-tests were used for comparisons between experimental groups, and P < 0.05 was considered statistically significant.
Results
The expression of rno-miR-1298 was decreased in C6 cells
First, the expression of rno-miR-1298 was evaluated in C6 cells using a qRT-PCR assay. As shown in Figure 1 and Table 1, the Ct value of rno-miR-1298 expression exceeds 35. The mean value of ΔCt was 23.56 in C6 cells. It is generally considered that when the ΔCt value is more than 16, the abundance of gene expression is low. Therefore, the abundance of rno-miR-1298 expression was low in C6 cells.

Rno-miR-1298 was down-regulated in C6 cells. The relative expression of rno-miR-1298 was examined in C6 cells by qRT-PCR. The Ct value of rno-miR-1298 expression exceeds 35, indicating the abundance of rno-miR-1298 expression was low in C6 cells.
The Ct of amplification.

The lentiviral vector of rno-miR-1298 was successfully constructed
In order to study the function of rno-miR-1298 in C6 cells, we successfully constructed the lentiviral vector of rno-miR-1298 fusing EGFP. As shown in Figure 2(a), the efficiency of cell infection was better, so subsequent experiment can be carried out using the lentiviral vector of rno-miR-1298.
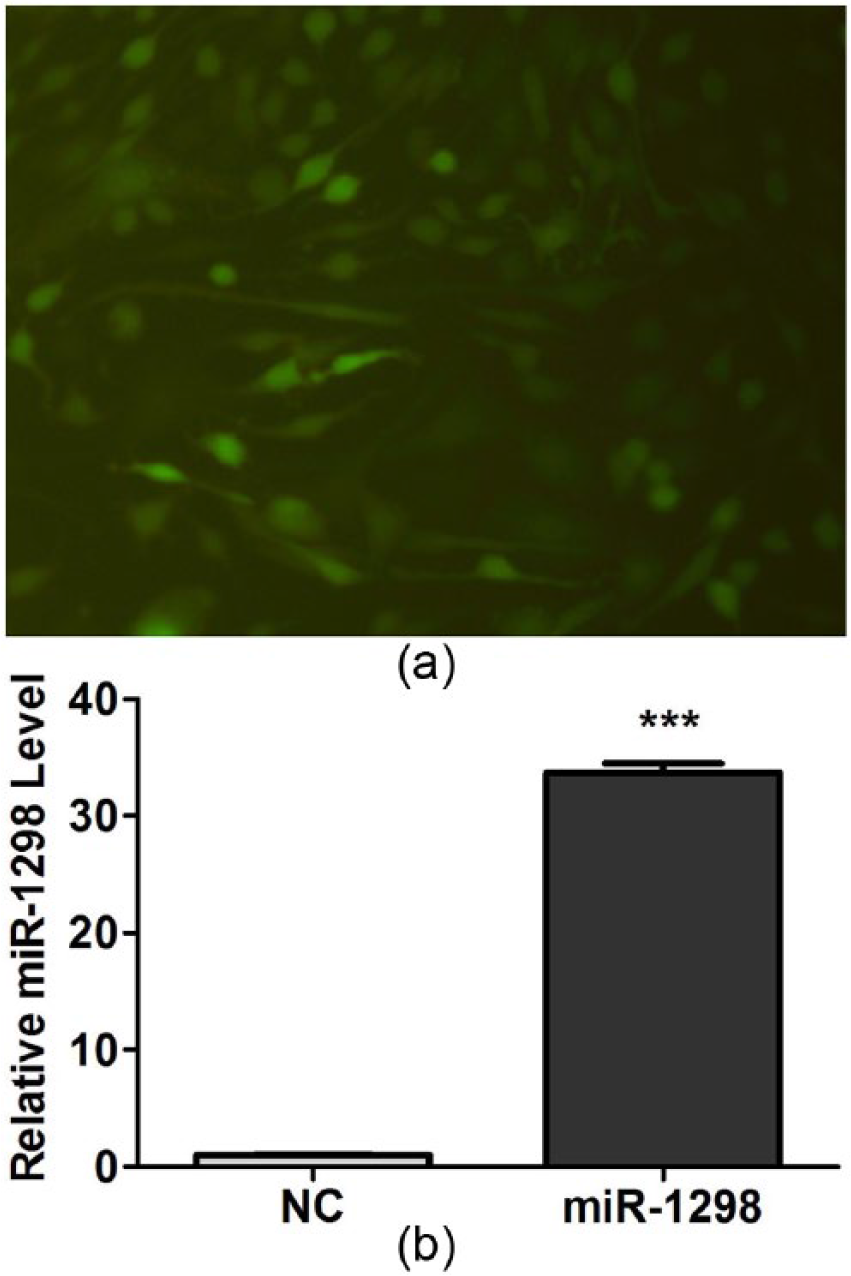
The lentiviral vector of rno-miR-1298 was successfully constructed and expressed: (a) the lentiviral vector of rno-miR-1298 was expressed in cell lines C6 and (b) the expression of rno-miR-1298 was evaluated by qRT-PCR after the lentiviral vector of rno-miR-1298 infected C6 cells. The expression of rno-miR-1298 was obviously increased in C6 cells. ***P < 0.001.
The expression of rno-miR-1298 was further evaluated using a qRT-PCR after infection of C6 cells. As shown in Figure 2(b), the expression of rno-miR-1298 was increased by about 33.7 times, confirming the availability of the lentiviral vector of rno-miR-1298.
rno-miR-1298 suppressed cell proliferation and induced cell apoptosis
To preliminarily evaluate whether ectopic expression of rno-miR-1298 affects the proliferation and apoptosis on tumor cells, we performed the MTT and flow cytometry assays after infecting C6 cells. MTT assay revealed that overexpression of rno-miR-1298 significantly inhibited the cellular proliferation. Especially, the inhibited ratio was significant at fourth and fifth days in C6 cells (Figure 3).

rno-miR-1298 suppressed cell proliferation. The effect of rno-miR-1298 on proliferation of C6 cells was detected by MTT. The results indicated that overexpression of rno-miR-1298 significantly inhibited proliferation of C6 cells and the inhibited ratio was significant at fourth and fifth days. *P < 0.05, **P < 0.01.
Flow cytometry assays revealed rno-miR-1298 significantly induced the apoptosis of cells (Figure 4(a) and (b)). As shown in Figure 4(c), compared to NC, rno-miR-1298 resulted in a significant increase in apoptosis rate with 6.38% versus 4.9% in C6 cells. These results indicated that rno-miR-1298 could obviously inhibit the proliferation and induce apoptosis of C6 cells.
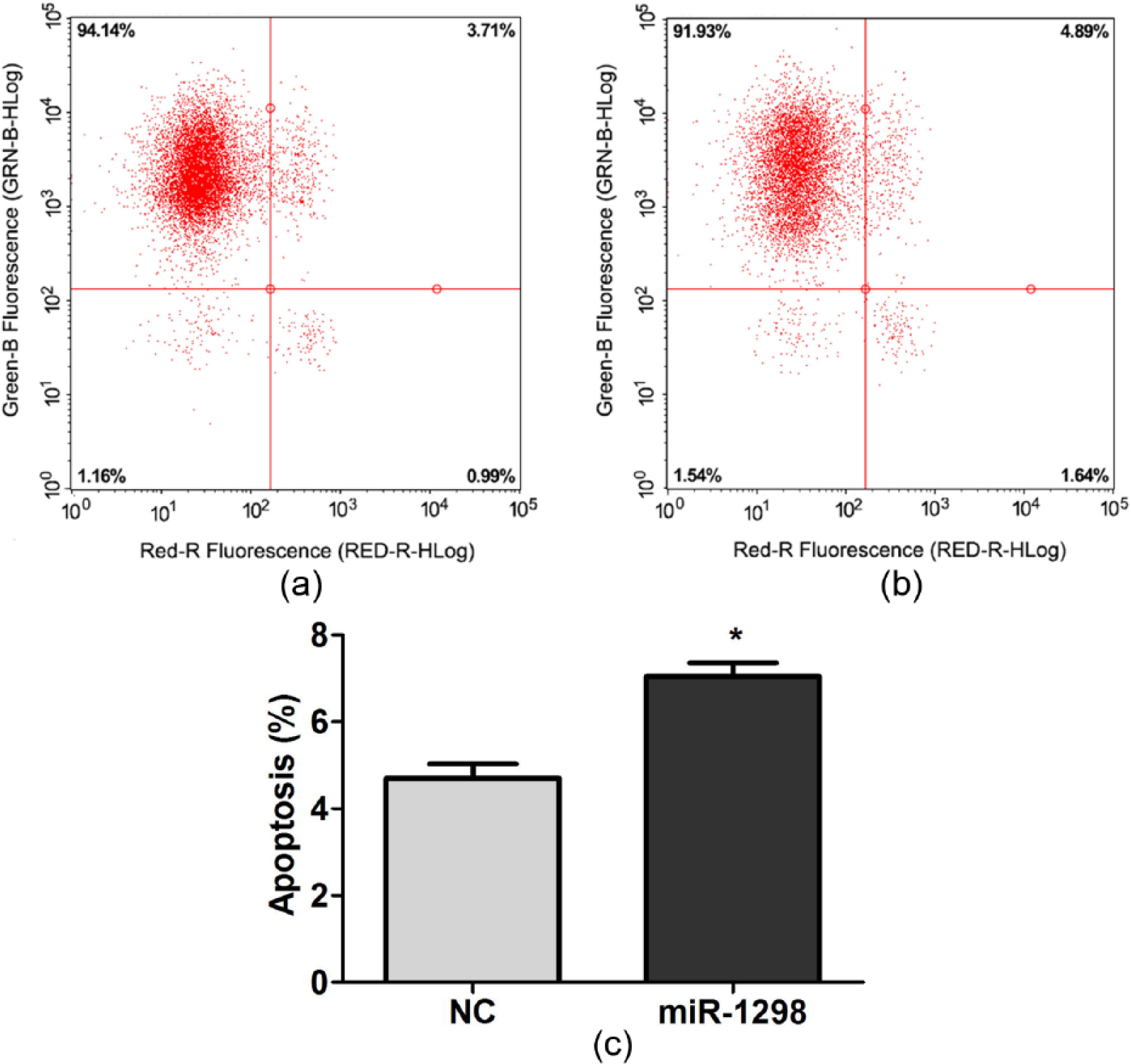
rno-miR-1298 induced cell apoptosis: (a, b) flow cytometry assays the effect of (b) rno-miR-1298 and (a) negative control on apoptosis of C6 cells; (c) statistical analysis of apoptosis indicated that rno-miR-1298 could obviously induced apoptosis of C6 cells. *P < 0.05.
SETD7 is a direct target of rno-miR-1298
To further explain the molecular mechanism by which rno-miR-1298 plays its biological roles, we first predicted SETD7 as a putative target of rno-miR-1298 using public databases (Figure 5(a)). What’s more, overexpression of rno-miR-1298 obviously depressed the expression of SETD7 at protein level in C6 cells. Furthermore, SETD7 expression was lower in C6 cells (73.48%; Figure 5(b)). To confirm whether SETD7 is directly and negatively regulated by rno-miR-1298, we constructed luciferase reporter vectors containing the WT or MT target sequences of rno-miR-1298. As shown in Figure 5(c), rno-miR-1298 significantly inhibited the luciferase activity of the WT compared with negative control (NC). As expected, the luciferase activity of MT was not significantly different compared with NC, suggesting that the miR-1298 binding site was important for miR-1298-mediated suppression. Together, these results demonstrate that SETD7 is indeed a direct downstream target of rno-miR-1298.

SETD7 was a direct target of rno-miR-1298: (a) sequence alignment of wild-type (WT) and mutated (MT) putative rno-miR-1298 binding sites in the 3′-UTR of SETD7, (b) the expression of SETD7 was examined after infection of rno-miR-1298 in C6 cells by western blot, and (c) relative luciferase activities of plasmids were detected in cells expressed WT or MT of SETD7 3′-UTR. rno-miR-1298 significantly inhibited the luciferase activity of the WT compared with NC. **P < 0.01.
rno-miR-1298 targeted SETD7 to affect cellular proliferation and apoptosis
To identify the function of SETD7 on miR-1298-induced proliferation and apoptosis, C6 cells were transiently transfected with siRNA of SETD7 or NC (Sangon Biotech). As shown in Figure 6(a), knockdown of SETD7 obviously has further suppressive effects on cell proliferation from the fourth day. The similar results from flow cytometry (Figure 6(b)–(d)) showed that knockdown of SETD7 significantly induced effects on cell apoptosis of C6 cells. These results indicated that rno-miR-1298-targeted SETD7 could inhibit the proliferation and induce apoptosis of C6 cells.

rno-miR-1298 targeted SETD7 to affect cellular proliferation and apoptosis: (a) knockdown of SETD7 had obviously suppressive effects on cell proliferation from the fourth day in C6 cells, (b, c) flow cytometry showed that knockdown of SETD7 significantly induced cell apoptosis of C6 cells (b) compared to negative control (c), and (d) statistical analysis of apoptosis in C6 cells. **P < 0.01.
Discussion
Many studies have demonstrated that miRNA dysregulation plays vital roles in various types of tumors and their differential expressions are closely related to proliferation, migration, apoptosis, and so on.11–13 MiR-1298 is identified later and thus the research of miR-1298 is relatively few. It was reported that miR-1298 might inhibit proliferation and migration in the vascular smooth muscle cells (VSMCs). 14 Up to now, there is no report about the expression and function of miR-1298 in nervous system and relative tumors. In this study, we found that rno-miR-1298 was significantly down-regulated in rat tumor C6 cells. Moreover, it was further proved that overexpression of rno-miR-1298 obviously inhibited the proliferation and induced apoptosis of C6 cells. It was supported that miR-1298 functioned as a tumor suppressor and inhibited malignant progression.
SETD7 was a marker for transcriptional activation and might activate promoters of methylation-dependent co-recruitment by mediated methylation of non-histone proteins. 15 It has been indicated that SETD7 plays an important role in inflammation, metabolism-related diseases, and oncogenesis. It was reported that the expression of SETD 7 was significantly higher in hepatocellular carcinoma (HCC) tumor tissues. Overexpression of SETD 7 could promote cell proliferation of HepG2, whereas knockdown of SETD 7 can inhibit cell proliferation of SMMC-7721. 16 Similarly, in this study, we also found that knockdown of SETD 7 can inhibit cell proliferation of C6 cells, indicating that SETD7 might play a critical role in tumors.
It is general that miRNA as regulator exerts its function by modulating with its critical genes and signaling networks involved in tumorigenesis and malignant processes. 17 In this study, we predicted that SETD 7 was a target of miR-1298 using Target Scan, miRanda, and miRDB databases. Then, we further identified that SETD 7 was a direct target of miR-1298 by dual-luciferase activity. Furthermore, restoring the expression of miR-1298 significantly reduced the expression of SETD 7 at protein levels. In addition, knockdown of SETD 7 showed similar suppressive effects of miR-1298 on cell proliferation and induced effects of miR-1298 on cell apoptosis. Taken together, our data suggested that the effects of miR-1298 on proliferation and apoptosis observed in C6 cells may be partly due to its regulation of SETD 7.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was supported by grants from National Nature Science Foundation of China (No. 81501018).