
Abstract
Background
The neurobiological basis of migraine remains incompletely understood. Magnetic resonance spectroscopy (MRS) allows non-invasive quantification of neurochemical and metabolic patterns in the brain, offering unique insights into biochemical processes during distinct migraine phases. This systematic review provides a critical appraisal of existing evidence describing MRS-derived neurochemical and metabolic alterations during spontaneous and experimentally provoked migraine attacks.
Methods
A systematic review was conducted in accordance with the PRISMA statement and prospectively registered in PROSPERO. Comprehensive searches of PubMed, Embase, and Scopus were performed from database inception through August 1, 2025. Eligible studies included observational or interventional investigations acquiring 1H-MRS or 31P-MRS data during the ictal phase in adults with migraine, incorporating either non-ictal comparisons or healthy controls. Considerable variability in study design, brain regions, and metabolite outcomes precluded quantitative synthesis, necessitating a structured qualitative analysis organized by MRS technique and anatomical region.
Results
Eight studies published between 1988 and 2022 met inclusion criteria, comprising five 1H-MRS investigations and three 31P-MRS studies, some of which derived from overlapping participant cohorts. Brain regions examined included occipital cortex, pons, frontal cortex, basal ganglia, and parieto-occipital areas. In individual 1H-MRS studies, occipital cortex analyses demonstrated ictal elevations in total choline and total N-acetyl aspartate, while lower glutathione concentrations were observed. A single 1H-MRS study targeting the pons identified ictal increases in total creatine and total N-acetyl aspartate. Findings from 31P-MRS studies indicated altered cerebral energy metabolism during migraine attacks.
Conclusions
Available MRS evidence suggests that migraine attacks are associated with altered cerebral energy metabolism, particularly within visual cortical and brainstem regions. However, existing studies differ substantially in design, acquisition parameters, regions of interest, and analytical approaches, such that few directly address comparable questions. Thus, the reproducibility of reported findings remains uncertain. Establishing reliable attack-related metabolic signatures will require well-designed longitudinal MRS investigations capable of directly probing ictal dynamics.
This is a visual representation of the abstract.
Introduction
Migraine is a prevalent and disabling neurological disorder, 1 yet its underlying disease mechanisms remain incompletely understood despite extensive clinical and pathophysiological investigation. 2 Migraine attacks are often conceptualized as progressing through interictal, prodromal, aura, headache, and postdromal phases; however, the presence, sequence, and duration of these phases vary substantially across individuals and attacks. 2 Among these phases, the headache, or ictal, phase constitutes the most debilitating stage and can persist from several hours to multiple days. 2
The ictal phase is thought to reflect the convergence of altered cortical excitability, activation of pain-processing networks, and disruptions in neurotransmitter signaling and cerebral energy metabolism.3–6 Characterizing the neurochemical environment during this phase is therefore critical for advancing mechanistic models of migraine pathophysiology and for identifying potential disease biomarkers. However, direct in vivo assessment of these processes during active migraine attacks remains challenging.
Magnetic resonance spectroscopy (MRS) provides a non-invasive means of quantifying brain neurochemicals and metabolites in vivo, offering information complementary to structural and functional neuroimaging approaches.7–9 Using proton (1H-MRS) and phosphorus (31P-MRS) techniques,7–10 MRS enables the assessment of metabolites related to excitatory and inhibitory neurotransmission, neuronal and glial integrity, and cellular energy homeostasis.7–10 Interictal MRS studies have yielded important insights into migraine pathophysiology, including evidence for altered neurotransmitter balance and changes in brain metabolism.3–5
In contrast, relatively few studies have applied MRS during the ictal phase.11–18 Existing ictal-phase studies report methodologically diverse findings, likely reflecting substantial variability in study design, brain regions examined, magnetic field strength, and the use of spontaneous versus experimentally provoked attacks. To date, no systematic synthesis has focused specifically on MRS findings acquired during the ictal phase of migraine.
To address this gap, we synthesized and critically appraised the available evidence describing neurochemical and metabolic changes measured with MRS during migraine attacks. Specifically, we summarize reported ictal metabolite alterations by brain region and MRS modality, evaluate methodological variability across studies, and identify key priorities for future research.
Methods
This study was conducted as a systematic review without meta-analysis, given anticipated methodological variability, and was reported in accordance with the PRISMA 2020 statement and the SWiM reporting guideline.19,20 The review protocol was prospectively registered in PROSPERO (Identifier: CRD420251125199). Informed consent was not required for this study, as it is a systematic review of previously published data and did not involve direct participation of human subjects.
Search strategy and selection criteria
A comprehensive literature search was performed in PubMed, Embase via Ovid, and Scopus from database inception through August 1, 2025. Searches used free-text terms related to “migraine” and “magnetic resonance spectroscopy”, with full database-specific search strategies provided in Supplemental Table 1. Reference lists of all included articles were manually screened to identify additional relevant publications not captured by the initial database searches.
Eligible articles reported original investigations using proton (1H)-MRS or phosphorus (31P)-MRS and included adult participants (≥18 years) diagnosed with migraine with aura, migraine without aura, or chronic migraine. To be eligible, studies were required to acquire MRS data during the ictal phase and to include a comparator. The comparator was defined as either an interictal scan obtained from the same participants with migraine or a healthy control group. Both observational and interventional designs were eligible, with no restrictions on language or publication date. Reviews, case reports, conference abstracts, animal studies, technical validation studies without metabolite outcomes, and articles lacking ictal-phase metabolite comparisons were excluded. A complete list of eligibility criteria is outlined in Table 1.
Eligibility criteria.
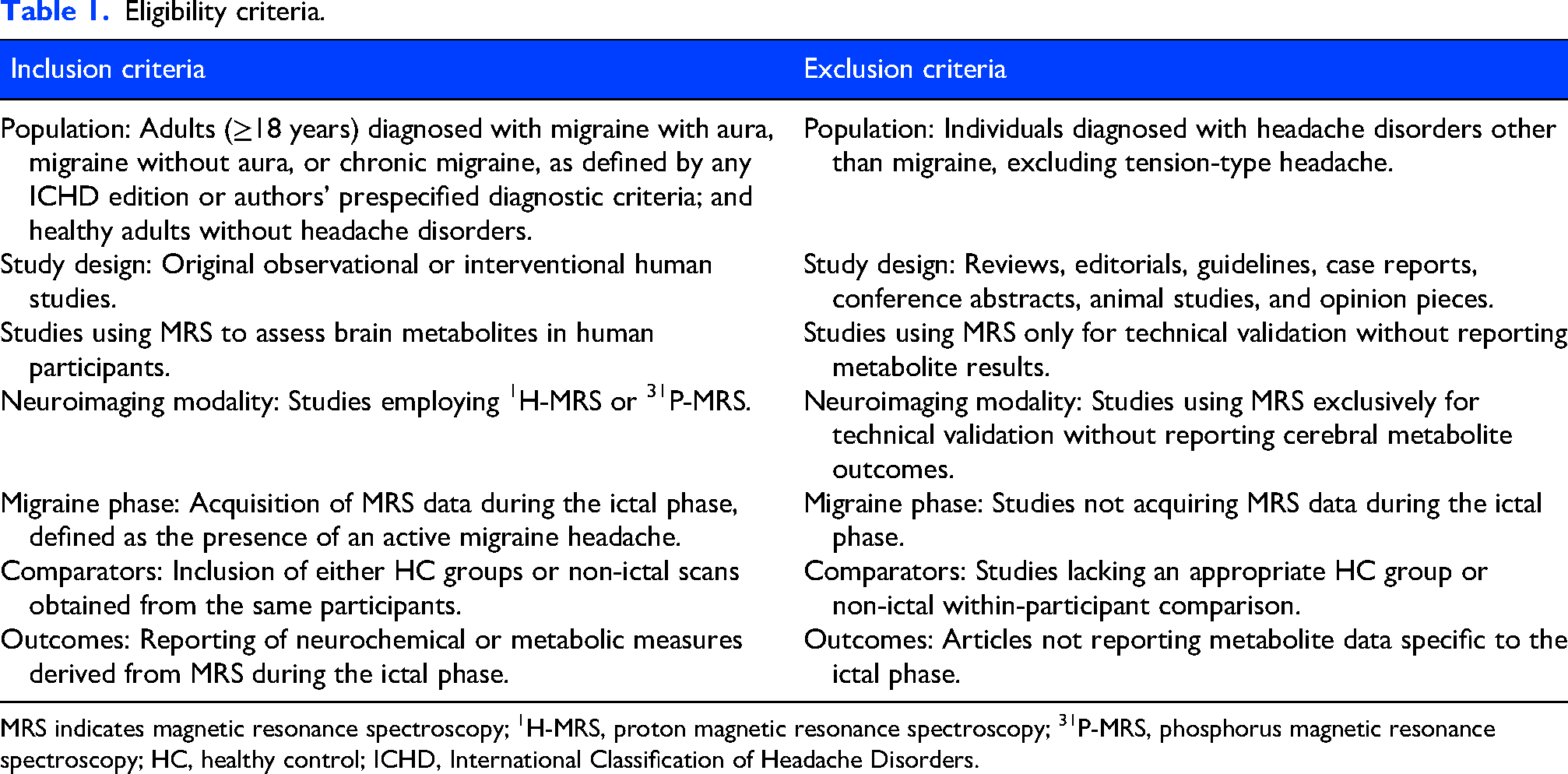
MRS indicates magnetic resonance spectroscopy; 1H-MRS, proton magnetic resonance spectroscopy; 31P-MRS, phosphorus magnetic resonance spectroscopy; HC, healthy control; ICHD, International Classification of Headache Disorders.
Definition of ictal phase
To ensure consistency across included studies, the ictal phase was defined as the period during which an active migraine headache was present. In spontaneous attacks, this definition corresponded to diagnostic criteria for migraine without aura specified in any edition of the International Classification of Headache Disorders (ICHD). 1 When explicitly reported, authors’ prespecified case definitions of a migraine attack were also accepted. For experimentally provoked attacks, ictal-phase classification followed each article's prespecified criteria, reflecting variability in provocation paradigms and outcome definitions across studies.
Overview of magnetic resonance spectroscopy
MRS is a non-invasive neuroimaging technique that enables in vivo quantification of cerebral neurochemical and metabolic composition from predefined regions of interest. Unlike conventional magnetic resonance imaging (MRI), which primarily provides anatomical information, MRS yields biochemical profiles reflecting molecular constituents.
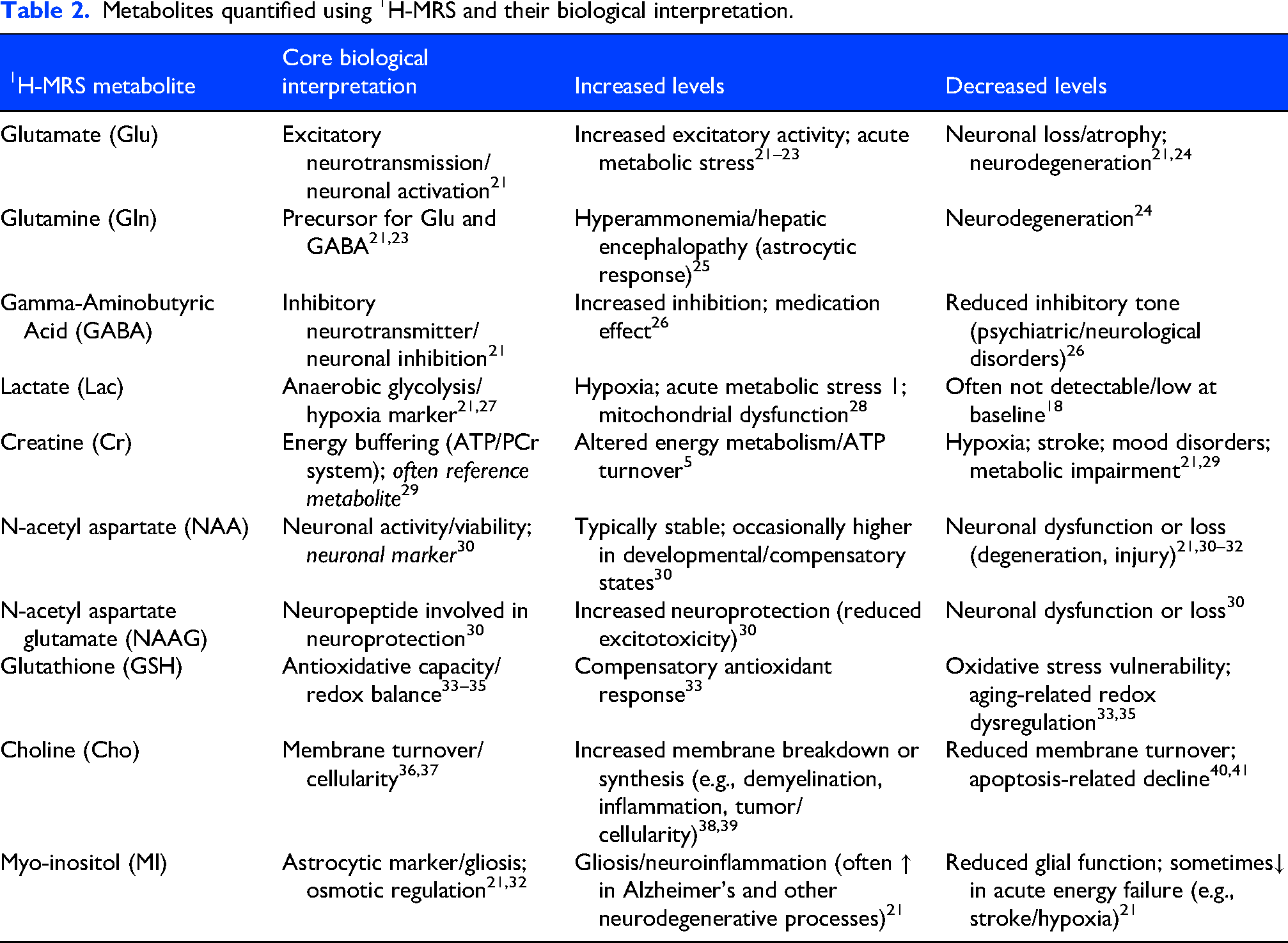
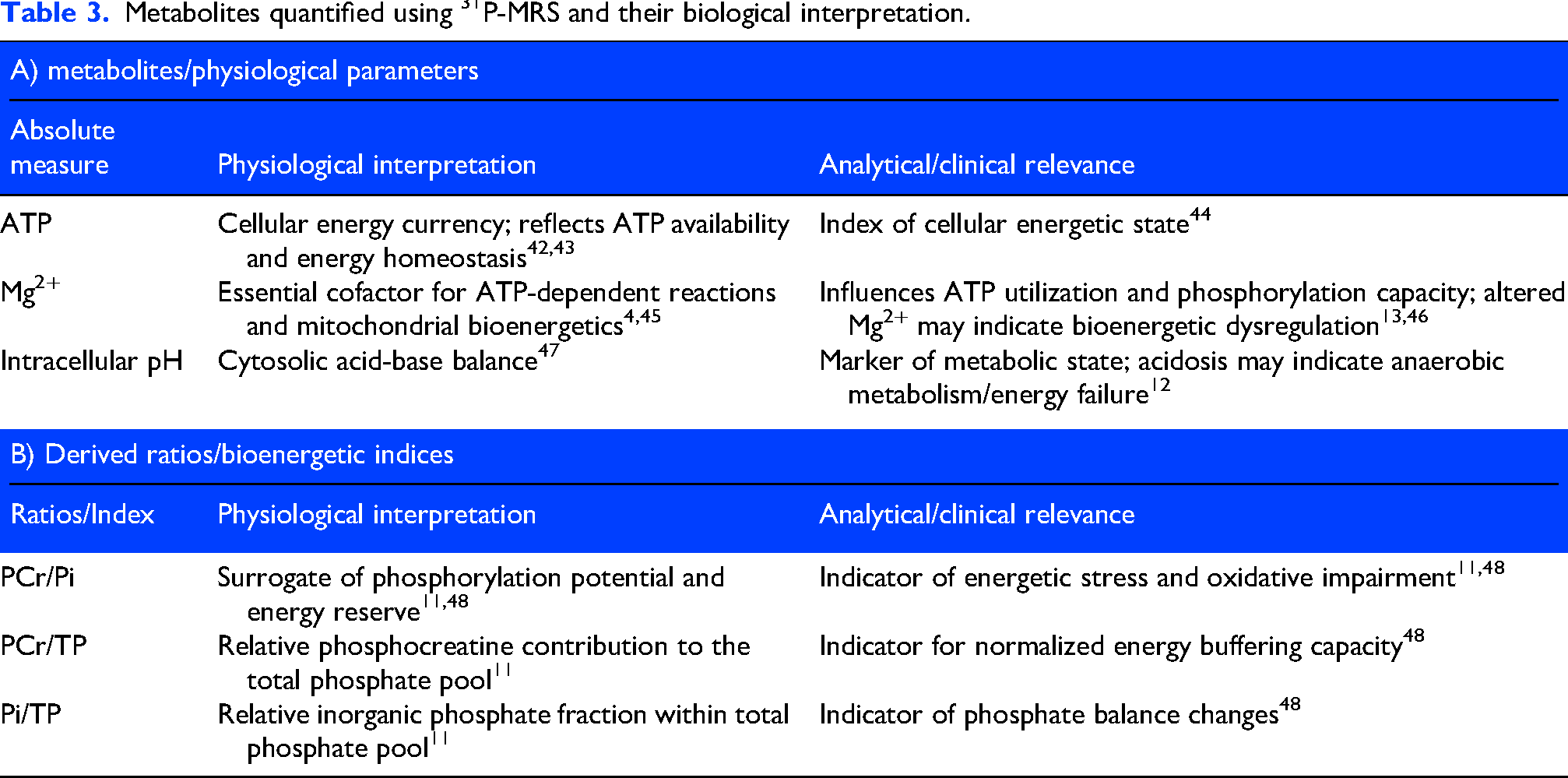
Both 1H-MRS and phosphorus-based 31P-MRS techniques are used in human neuroimaging research. 1H-MRS permits quantification of metabolites related to excitatory and inhibitory neurotransmission, including glutamate (Glu) and γ-aminobutyric acid (GABA), as well as markers of neuronal integrity such as N-acetyl aspartate (NAA) and glial markers including myoinositol (MI). 31P-MRS enables assessment of compounds involved in cellular energy metabolism, including creatine (Cr), phosphocreatine (PCr), inorganic phosphate (Pi), and adenosine triphosphate (ATP). Additional commonly reported composite and redox-related measures include total creatine (tCr), total choline (tCho), total N-acetyl aspartate (tNAA), choline (Cho), glutathione (GSH), intracellular magnesium (pMg2+), total phosphate (TP), and intracellular pH. A detailed overview of the investigated MRS metabolites and their biological interpretation is presented in Tables 2 and 3.
Metabolites quantified using 1H-MRS and their biological interpretation.
Metabolites quantified using 31P-MRS and their biological interpretation.
MRS data are acquired using either single-voxel spectroscopy, which samples a single brain region, or multi-voxel chemical shift imaging (2D or 3D CSI), which allows mapping of metabolite concentrations across multiple brain regions. Interpretation of MRS measurements depends on acquisition and processing parameters, including voxel placement, magnetic field strength, sequence design, and whether neurochemical or metabolite levels are reported as absolute concentrations or relative ratios.
Data extraction and analysis
All records were imported into EndNote for de-duplication. Two reviewers (D-IR and ES) independently screened titles and abstracts for eligibility, followed by full-text review of potentially relevant studies. Disagreements were resolved by consensus or consultation with a third reviewer (RHC). Data extraction was performed independently by two reviewers using a standardized form.
Extracted data included publication details, participant characteristics, study design, examined brain regions, MRS modality, scanner field strength, acquisition and sequence parameters, voxel size, and quantification methods. Additional extracted elements comprised post-processing software, statistical analyses, metabolite outcomes, reported limitations, and key findings. Further variables included the use of migraine provocation agents, timing of image acquisition relative to attack onset, and characteristics of comparator groups.
Data synthesis
Given substantial difference in study designs, MRS protocols, brain regions, and outcome measures, quantitative meta-analysis was not performed. Instead, a structured qualitative synthesis was undertaken, with findings grouped by MRS modality and brain region, and quantitatively analyzed to identify recurring patterns of ictal neurochemical and metabolic alteration.
Risk of bias and methodological appraisal
Because no validated risk-of-bias tool exists specifically for MRS studies, methodological quality was assessed using a domain-based appraisal informed by expert recommendations for MRS research. 9 Domains included blinding, placebo control, participant matching, spectral preprocessing, eddy current and motion correction, referencing accuracy, voxel placement consistency, spectral modeling quality control, and correction for multiple comparisons.9,49
Results
The database search identified 1245 records. After duplicate removal, 629 unique titles and abstracts were screened, and 68 full-text articles were assessed for eligibility. Eight studies met inclusion criteria and were included in the qualitative synthesis.11–18 The study selection process is illustrated in Figure 1.
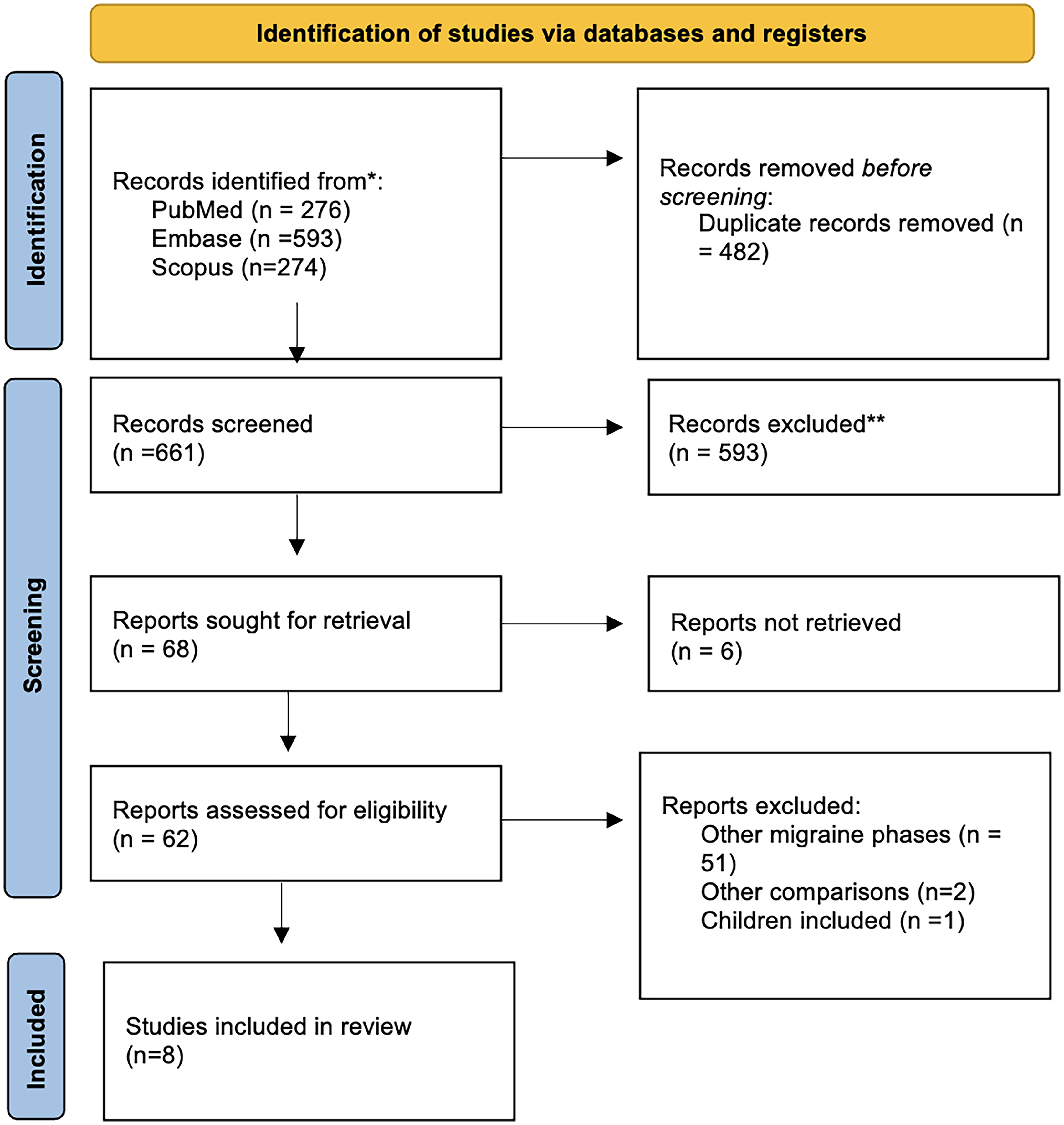
Flow diagram.
Study characteristics
A concise summary of the included publications is provided in Tables 4 and 5. Additionally, Figures 2 and 3 offer an illustrative overview of the observed metabolite changes. The eight included studies were published between 1988 and 2022 and comprised five investigations using 1H-MRS14–18 and three using 31P-MRS.11–13 Study designs included five cross-sectional studies11–14,16 and three longitudinal investigations.15,17,18 Sample sizes ranged from a single participant to 34 participants with migraine. Six studies included healthy control groups for comparison,11–16 while two focused on the difference between the interictal and ictal phases.17,18
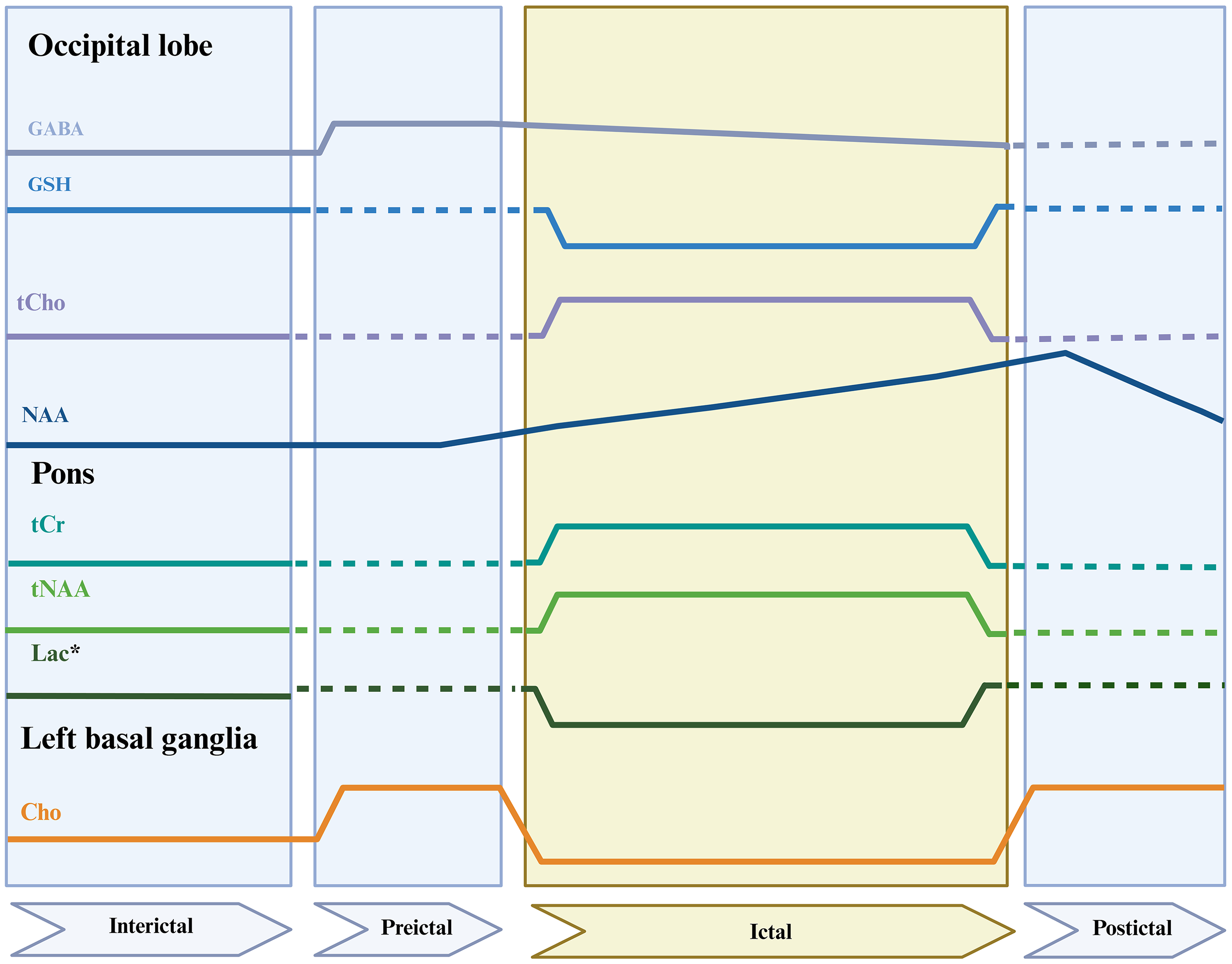
Regional 1H-MRS-derived alterations across migraine phases. Schematic representation of reported changes in 1H-MRS-derived metabolites across interictal, preictal, ictal, and postictal phases, stratified by brain region. Solid lines indicate reported metabolite alterations, dashed lines denote phases or regions for which data were not reported, and the shaded area highlights the ictal phase. Glu, glutamate; GABA, γ-aminobutyric acid; tCho, total choline; GSH, glutathione; NAA, N-acetyl aspartic acid; tNAA, total NAA; tCr, total creatine; Cho, choline; Lac, lactate; - - -, no reported data; *, the results were statistically significant only after the exclusion of two outliner values; For the NAA the parameters are measured only in the left occipital cortex.
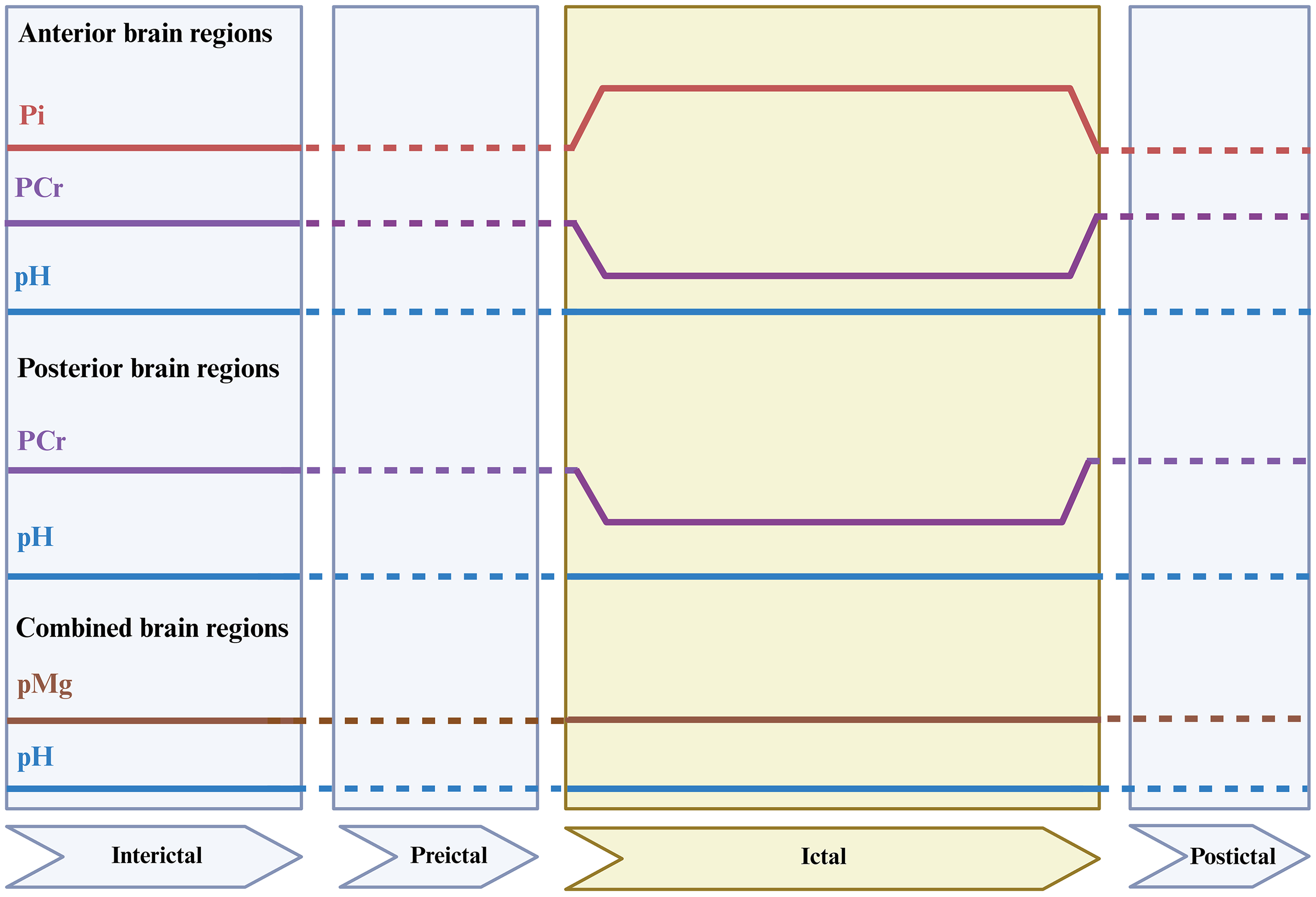
Regional 31P-MRS–derived metabolite alterations across migraine phases. Schematic representation of reported changes in 31P-MRS-derived metabolites across interictal, preictal, ictal, and postictal phases, stratified by brain region. Solid lines indicate reported metabolite alterations, dashed lines denote phases or regions for which data were not reported, and the shaded area highlights the ictal phase. Abbreviations: Pi, inorganic phosphate; PCr, phosphocreatine; pH, intracellular pH; pMg2+, negative logarithm of magnesium ion concentration.
Summary of eligible 1H-MRS studies.
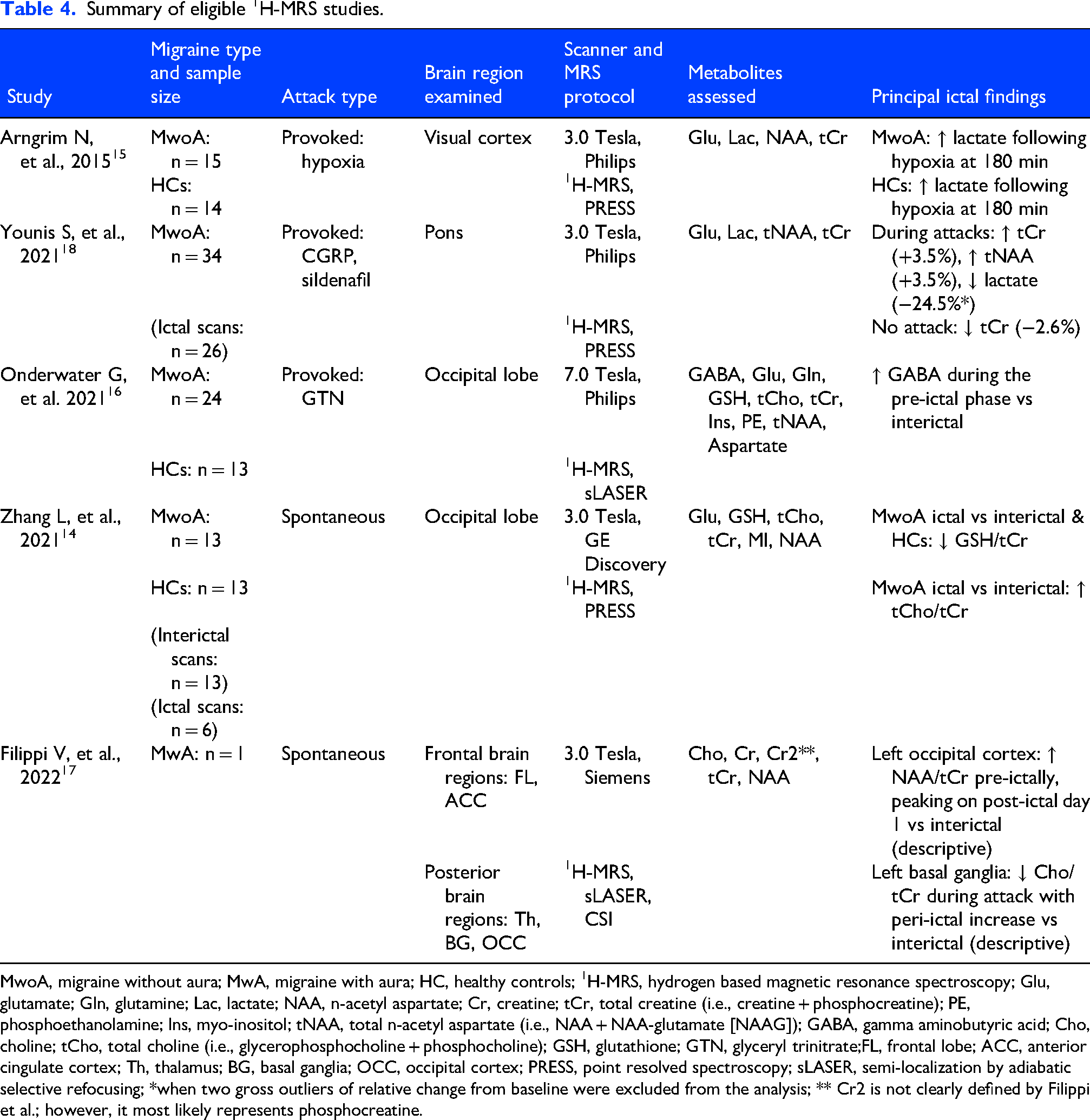
MwoA, migraine without aura; MwA, migraine with aura; HC, healthy controls; 1H-MRS, hydrogen based magnetic resonance spectroscopy; Glu, glutamate; Gln, glutamine; Lac, lactate; NAA, n-acetyl aspartate; Cr, creatine; tCr, total creatine (i.e., creatine + phosphocreatine); PE, phosphoethanolamine; Ins, myo-inositol; tNAA, total n-acetyl aspartate (i.e., NAA + NAA-glutamate [NAAG]); GABA, gamma aminobutyric acid; Cho, choline; tCho, total choline (i.e., glycerophosphocholine + phosphocholine); GSH, glutathione; GTN, glyceryl trinitrate;FL, frontal lobe; ACC, anterior cingulate cortex; Th, thalamus; BG, basal ganglia; OCC, occipital cortex; PRESS, point resolved spectroscopy; sLASER, semi-localization by adiabatic selective refocusing; *when two gross outliers of relative change from baseline were excluded from the analysis; ** Cr2 is not clearly defined by Filippi et al.; however, it most likely represents phosphocreatine.
Summary of eligible 31P-MRS studies.
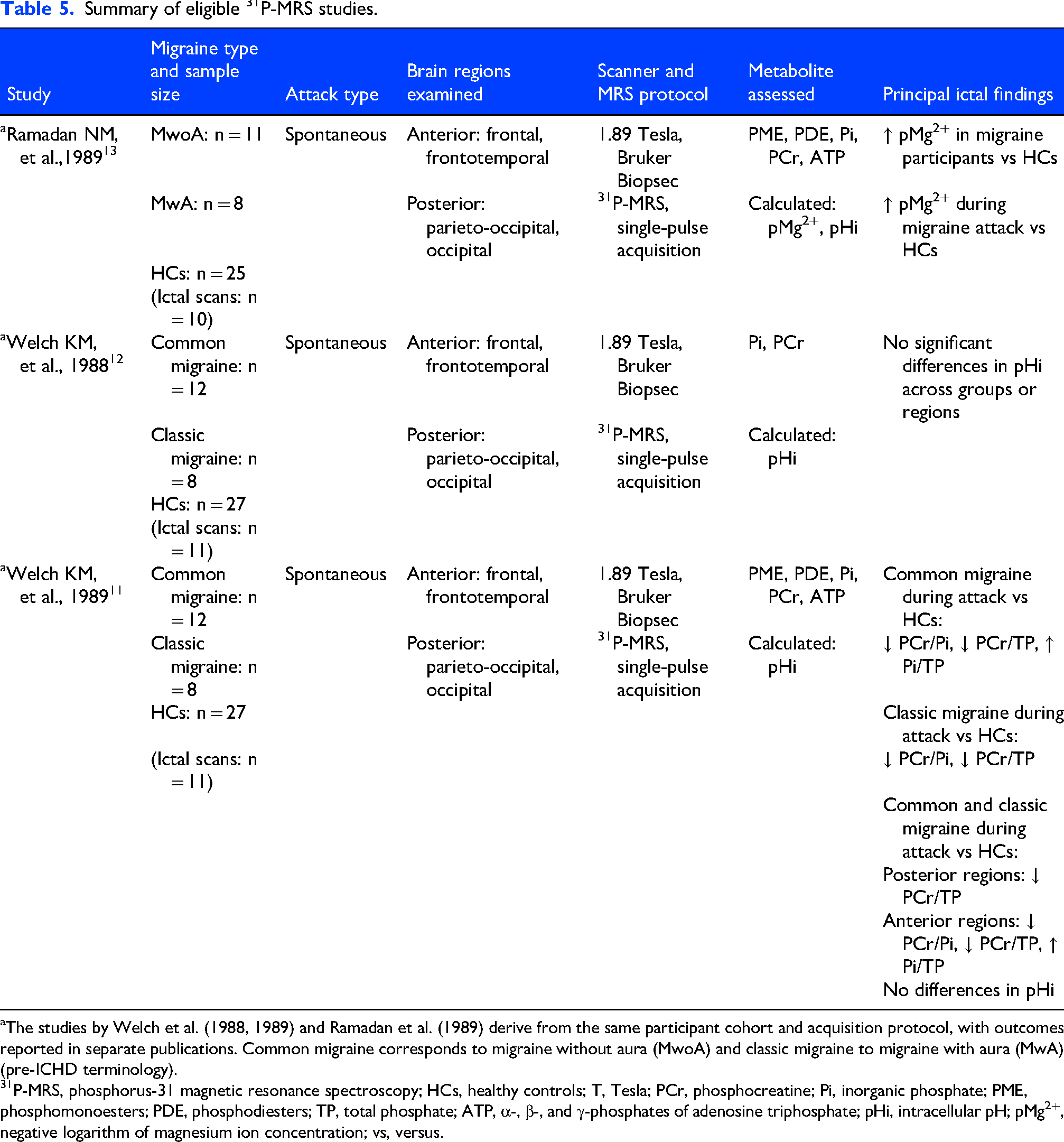
The studies by Welch et al. (1988, 1989) and Ramadan et al. (1989) derive from the same participant cohort and acquisition protocol, with outcomes reported in separate publications. Common migraine corresponds to migraine without aura (MwoA) and classic migraine to migraine with aura (MwA) (pre-ICHD terminology).
P-MRS, phosphorus-31 magnetic resonance spectroscopy; HCs, healthy controls; T, Tesla; PCr, phosphocreatine; Pi, inorganic phosphate; PME, phosphomonoesters; PDE, phosphodiesters; TP, total phosphate; ATP, α-, β-, and γ-phosphates of adenosine triphosphate; pHi, intracellular pH; pMg2+, negative logarithm of magnesium ion concentration; vs, versus.
Most studies (n = 6) diagnosed migraine according to ICHD criteria.13–18 Two earlier studies used historical classifications, including “common” migraine and “classic” migraine.11,12 Three publications derived from the same underlying dataset but reported distinct metabolic outcomes.11–13
Five studies examined spontaneous migraine attacks,11–14,17 whereas three used experimental provocation paradigms, including hypoxia, 15 glyceryl trinitrate (GTN), 16 calcitonin gene-related peptide (CGRP), 18 and sildenafil. 18 All provocation studies employed 1H-MRS,15,16,18 whereas all 31P-MRS studies examined spontaneous attacks only.11–13
Magnetic field strengths varied across studies. Four 1H-MRS investigations used 3-Tesla scanners,14,15,17,18 one used a 7-Tesla scanner, 16 and all 31P-MRS studies used 1.89-Tesla scanners.11–13 Moreover, targeted brain regions also differed. Among 1H-MRS studies, three examined the occipital cortex,14–16 one focused on the pons, 18 and one assessed multiple cortical and subcortical regions. 17 The 31P-MRS studies all evaluated frontal, frontotemporal, parieto-occipital, and occipital regions.11–13
Findings from 1H-MRS studies of spontaneous migraine attacks
Zhang et al. conducted a cross-sectional 1H-MRS study examining occipital cortex metabolites in adult participants with migraine without aura. 14 The study included six participants scanned during the ictal phase, 13 scanned during the interictal phase, and 13 HCs. Ictal scans demonstrated significantly lower GSH/tCr compared with both interictal scans and HCs across the occipital cortex. In contrast, interictal scans showed significantly lower tCho relative to both ictal scans and HCs. No significant group differences were detected for other metabolites, including NAA, Glu, or MI across participant groups.
Filippi et al. reported a longitudinal case study involving a single participant with migraine with aura who underwent daily 1H-MRS acquisitions over a continuous 21-day period. 17 During this interval, two spontaneous migraine attacks without aura occurred, enabling MRS assessment across interictal, pre-ictal, ictal, and post-ictal phases. The peri-ictal phase was defined as the combined pre-ictal interval, preceding headache onset by four hours, and the post-ictal interval extending 24 h after headache resolution. 1H-MRS data were obtained from multiple regions, including bilateral frontal lobes, anterior cingulate cortex, thalamus, basal ganglia, and occipital cortex. Within the left occipital cortex, NAA/tCr increased during the pre-ictal interval and peaked on the first post-ictal day relative to interictal measurements. In the left basal ganglia, Cho/tCr decreased during the ictal phase and increased during the peri-ictal phase. All reported observations were descriptive in nature, and no formal statistical analyses were conducted.
Findings from 1H-MRS studies of provoked migraine attacks
Younis et al. performed a randomized, double-blind, double-dummy, crossover MRI study examining pontine metabolites and perfusion during experimentally provoked migraine attacks. 18 Twenty-six participants with migraine without aura underwent imaging across 41 induced attacks, including 20 following intravenous CGRP infusion and 21 following oral sildenafil administration. 1H-MRS was used to quantify pontine metabolite concentrations, while arterial spin labeling assessed regional cerebral blood flow. Imaging targeted the pons ipsilateral to the predominant pain side, or the right pons in bilateral headaches, with scans acquired at baseline and six hours after dosing. During migraine attacks, regional cerebral blood flow increased significantly within the dorsolateral pons ipsilateral to maximal pain intensity relative to baseline measurements. Pontine Glu levels remained unchanged, whereas tCr and tNAA increased during attacks compared with baseline values. Lactate concentrations showed no overall change; however, following exclusion of two outliers, lactate levels decreased significantly relative to baseline. No significant differences in metabolite levels were observed between CGRP-induced and sildenafil-induced migraine attacks.
Arngrim et al. reported from data a randomized, double-blind, sham-controlled, crossover study examining vascular and metabolic responses during hypoxia-induced migraine attacks. 15 Fifteen participants with migraine with aura underwent 180 min of normobaric hypoxia and sham air inhalation on separate study days, while 14 matched HCs were exposed to hypoxia only. Using 1H-MRS, glutamate, lactate, NAA, and tCr were quantified within the visual cortex at baseline, during hypoxic inhalation, and following recovery. Hypoxia provoked migraine attacks in eight participants, including three attacks fulfilling criteria for migraine with aura, whereas only one migraine without aura attack occurred during sham exposure. Visual cortical lactate concentrations increased significantly during hypoxia, whereas glutamate, NAA, and tCr levels remained unchanged. Comparable lactate increases were also observed among HCs, and no significant between-group differences in metabolite responses were detected.
Onderwater et al. conducted a cross-sectional 7-Tesla 1H-MRS study including 24 female participants with migraine without aura and 13 age-matched HCs. 16 All participants received an intravenous infusion of glyceryl trinitrate, with MRS acquisitions performed at baseline, 90 min, and 270 min following infusion onset. Metabolite responses were evaluated across interictal, pre-ictal, and ictal phases, with the pre-ictal phase defined as the interval between infusion administration and headache onset. Glutamate concentrations remained stable across all phases, whereas GABA levels increased during the transition from interictal to pre-ictal phases in participants with migraine. Given the absence of correction for multiple comparisons, the reported results should be interpreted cautiously.
Findings from 31P-MRS studies of spontaneous migraine attacks
Ramadan et al. conducted a cross-sectional 31P-MRS study examining intracellular pMg2+ and intracellular pH during spontaneous migraine attacks. 13 The study included 19 participants with migraine, with and without aura, and 25 HCs, of whom 10 participants with migraine were scanned ictally and nine interictally. 31P-MRS data were acquired from frontal, frontotemporal, parieto-occipital, and occipital cortical regions, sampled unilaterally or bilaterally depending on feasibility. Intracellular pMg2+ values were significantly elevated during migraine attacks relative to HCs, indicating reduced intracellular magnesium concentrations during the ictal phase. Intracellular pH values did not differ across any participant group or migraine phase.
In two related studies derived from the same but slightly expanded dataset, Welch et al. applied 31P-MRS to investigate cortical metabolism in 12 participants with common migraine, 8 participants with classic migraine, and 27 HCs.11–13 The initial report assessed intracellular pH across frontal, frontotemporal, parieto-occipital, and occipital regions, including 11 participants scanned during the ictal phase, and identified no group differences. A subsequent analysis of phosphate metabolism demonstrated significantly reduced PCr/Pi and PCr/TP ratios and increased Pi/TP values in participants with migraine,during attack, compared with HCs. These alterations were most pronounced in anterior cortical regions, whereas posterior regions showed isolated reductions in PCr/Pi. During the interictal phase, a modest but significant elevation in Pi/TP was also observed.
Quality assessment
Across the eight included studies, methodological quality varied substantially in key domains relevant to MRS reliability and interpretability.11–18 Blinded data analysis procedures were reported in four studies,11,15,16,18 while two of the three interventional investigations incorporated a placebo-controlled design.15,16,18 Age and sex matching was performed in three studies,14–16 whereas this criterion was not applicable in two investigations using within-subject or single-participant designs.17,18
Reporting of MRS-specific methodological details varied across included studies, with inconsistent description of acquisition parameters, preprocessing pipelines, and quality control procedures. Spectral preprocessing was described in five studies,13,14,16–18 eddy current correction was reported in four publications,12,13,16,18 and motion correction procedures were explicitly reported in two investigations.16,18 Consistent voxel placement across scanning sessions was reported in five studies.14–18
With respect to voxel tissue composition, explicit segmentation was reported in only two studies.16,18 Several others attempted to minimize cerebrospinal fluid contamination by placing voxels within brain parenchyma.14,15,50 However, formal correction for gray matter, white matter, and cerebrospinal fluid fractions was not applied. Basis set specification was provided in a single publication, 16 whereas spectral quality metrics were reported in five studies.11,12,14,16,17 Although all investigations described spectral modeling procedures, correction for multiple comparisons was implemented in only three studies11,14,18 and one investigation relied solely on descriptive analyses. 17
Discussion
This systematic review provides the first focused synthesis of MRS studies examining neurochemical and metabolic alterations during the ictal phase of migraine. Despite limited data and considerable methodological heterogeneity, the available findings suggest alterations in cerebral energy metabolism rather than consistent changes in excitatory or inhibitory neurotransmission during attacks. Most investigations focused on visual cortical regions, consistent with their established role in migraine-related sensory processing. However, the small number of studies, variability in methodology, and restricted regional coverage limit definitive mechanistic conclusions. Taken together, the current evidence indicates that migraine attacks are associated with metabolic alterations consistent with increased neuronal energy demand, particularly within posterior cortical and selected brainstem regions.
The occipital lobe and visual Cortex
The occipital lobe has been the most frequently examined region in ictal MRS studies,14–16 reflecting its established involvement in aura, photophobia, and visual sensory hyperexcitability associated with migraine. 22 Most investigations targeted neurotransmitters implicated in cortical excitability, particularly glutamate and GABA, given their central roles in excitatory–inhibitory balance.25,51,52 Across studies, however, ictal glutamate concentrations in the occipital cortex remained largely unchanged compared to interictal states or HCs.14–16 This absence of consistent excitatory alterations contrasts with interictal MRS literature reporting elevated glutamate or reduced GABA in migraine,53–56 possibly suggesting phase-specific neurochemical dynamics.
Several factors might contribute to these negative findings, including small sample sizes and modest signal-to-noise ratios for glutamate quantification. One study reported stable ictal GABA levels but increased pre-ictal concentrations, although this observation arose from a glyceryl trinitrate paradigm with rapid headache onset, potentially blurring phase boundaries. 16 Taken together, current data do not support robust ictal shifts in classical excitatory or inhibitory neurotransmitters within the visual cortex.
In contrast, some occipital cortex findings implicate altered energy metabolism during attacks.11–13 Reduced PCr and altered phosphate ratios in posterior regions indicate increased energetic demand during migraine, consistent with heightened neuronal activation and cortical hyperexcitability. 11 Experimental evidence demonstrating PCr depletion during visual stimulation supports this interpretation. 57 Additional observations of reduced GSH suggest increased oxidative stress accompanying elevated metabolic activity, although this finding has been reported only once and requires replication. 14 In hypoxia-induced attacks, increased lactate concentrations were observed but mirrored changes in HCs, 15 indicating an effect driven by hypoxic exposure rather than migraine-specific mechanisms.
Most ictal MRS studies have focused on the occipital cortex, reflecting its established role in migraine aura and its relative accessibility for voxel placement.58,59 However, the large voxel volumes required for reliable metabolite quantification limit spatial specificity and preclude detailed differentiation of functionally distinct subregions within the occipital cortex. Moreover, metabolic abnormalities in the occipital cortex are not specific to migraine and have been reported in other neurological disorders, including cerebral autosomal dominant arteriopathy with subcortical infarcts and leukoencephalopathy,60,61 and posterior reversible encephalopathy syndrome.62–64 Interestingly, both disorders can present with symptoms resembling migraine.65,66
In contrast, ictal MRS data from central pain processing regions such as the anterior cingulate cortex, insula, somatosensory cortex, and thalamus remain limited. Future investigations should prioritize theory driven and strategically selected regions of interest to better characterize migraine-related neurochemical alterations.
The pons and other regions
Despite extensive evidence implicating subcortical and brainstem structures in migraine pathophysiology,67–73 MRS investigations outside the occipital cortex remain sparse. Among these regions, the dorsal pons has received particular attention due to its supposed activation during migraine attacks and its anatomical connections with pain-modulatory networks,67,68 including the periaqueductal gray. 72 Only one study directly examined pontine neurochemistry during ictal states using provoked attacks. 18
In that investigation, 18 increased regional blood flow within the dorsolateral pons was accompanied by stable glutamate levels but elevated tCr and tNAA during attacks, suggesting increased metabolic turnover and altered neuronal energetics rather than excitatory neurotransmission. However, tNAA elevations were observed irrespective of headache development, complicating interpretation of its ictal specificity. Following exclusion of outliers, pontine lactate levels were reported to decrease. Since lactate is closely linked to brain energy metabolism, this finding may suggest altered metabolic processes during heightened neuronal activity, 74 although the result requires cautious interpretation.
Other non-occipital regions have been examined only sporadically.11–13,17 Frontal and frontotemporal cortices demonstrated phosphate shifts resembling those observed in posterior cerebral regions, 11 consistent with increased energetic demand during attacks. A single descriptive report noted reduced basal ganglia choline during the ictal phase, 17 although the absence of statistical testing and the single-participant design preclude inference. Together, the limited availability of ictal MRS data from pain processing regions, including the anterior cingulate cortex, insula, somatosensory cortex and thalamus, constrains conclusions regarding the regional specificity of attack-related metabolic alterations.
Methodological considerations and future directions
Given the limited evidence base, marked by small sample sizes, methodological limitations, and a lack of replication, it is not scientifically justifiable to describe a consistent metabolic “signature” of the migraine attack. Most investigations applied cross-sectional designs,11–14,16 comparing participants with migraine during headache attacks to HCs. Such designs risk conflating ictal phase specific alterations with broader migraine-related traits that persist outside the attack phase. For instance, previous work has demonstrated increased interictal glutamate in the visual cortex of participants with migraine, consistent with cortical hyperexcitability. 55 Alterations in GABA and Glx levels, as well as reduced NAA in cortical and subcortical regions, have also been reported independent of attack timing, suggesting disrupted excitatory and inhibitory balance as a migraine trait. 75 Although these specific alterations were not consistently observed in the included ictal studies, the possibility remains that pre-existing metabolic differences might obscure true attack-related changes in cross-sectional comparisons.
Provoked attack paradigms facilitate ictal data acquisition but could introduce metabolic effects related to the triggering stimulus itself. For example, hypoxia-induced migraine attacks have demonstrated lactate elevations that might reflect hypoxic physiology rather than migraine-specific mechanisms. 15 Although provoked and spontaneous attacks share clinical features, their underlying neurochemical profiles could differ to some extent. Future investigations should therefore prioritize spontaneous ictal acquisitions whenever feasible and directly compare spontaneous and induced attacks to determine the extent to which observed neurochemical alterations reflect migraine pathophysiology vs stimulus-related effects.
Technical factors represent major sources of variability across studies, including differences in magnetic field strength, acquisition sequences, voxel placement strategies, and hemispheric lateralization approaches MRS protocols are often optimized for specific metabolites, which can limit comprehensive metabolic assessment within a single acquisition. Because reliable metabolite quantification requires relatively large voxel sizes, MRS is inherently susceptible to partial volume effects. Individual voxels typically contain mixed gray matter, white matter, and cerebrospinal fluid, each characterized by distinct metabolite concentrations.
These considerations are particularly relevant for early 31P-MRS investigations,11–13 which were conducted at lower field strengths, involved small sample sizes, and lacked tissue composition correction. Large surface coil sampling volumes, although carefully positioned, did not permit adjustment for white matter or cerebrospinal fluid contributions. Most studies did not report partial volume correction,11–15,17 thereby limiting comparability of metabolic measures across investigations. Inconsistent reporting of preprocessing steps and statistical correction procedures further increases the risk of false positive findings and reduces cross-study interpretability.
Previous narrative reviews have summarized MRS findings in migraine across phases.3,4 In contrast, this review is restricted to attack-phase acquisitions and systematically evaluates each eligible ictal dataset. By organizing findings according to study design, comparator type, and attack model, the synthesis clarifies the inferential strength of available evidence and highlights the limited number of datasets that directly inform state-dependent change. A structured methodological appraisal tailored to ictal investigations further identifies specific technical and analytic priorities necessary to improve reproducibility and interpretability. Given that the attack phase represents the defining clinical event in migraine, this focused synthesis delineates the current evidentiary boundaries and provides a framework for future studies aimed at characterizing attack-related neurochemical alterations.
Future investigations should prioritize anatomically and functionally defined regions of interest that are directly implicated in migraine pathogenesis. 22 These regions should include pain processing structures such as the anterior cingulate cortex, insula, somatosensory cortex, and thalamus, together with the visual cortex given its plausible role in cortical hyperexcitability. 22 Analytical strategies should focus on neurochemical markers that reflect excitatory transmission, glial activity, and cellular energy metabolism. Particular emphasis should be placed on glutamate/glutamine and on myo-inositol. When acquisition time permits, spectral editing for GABA should be incorporated to enable more precise characterization of excitatory and inhibitory balance during the attack phase. Such a hypothesis-driven and regionally constrained approach might advance the identification of biologically meaningful signatures associated with migraine attacks. From a methodological standpoint, future studies should implement standardized acquisition protocols and rigorous quality control procedures. These procedures should include correction for multiple comparisons and transparent reporting of quality metrics. Study designs should prioritize longitudinal ictal and interictal assessments within the same participants. Investigators should also carefully account for potential confounding effects of acute and preventive migraine medications at the time of scanning.
Recommended 1H-MRS acquisition should incorporate semi-Localized by Adiabatic Selective Refocusing sequences to minimize chemical shift displacement error, particularly at 3T and higher field strengths.9,76,77 Post-processing pipelines should include tissue segmentation into gray matter, white matter, and cerebrospinal fluid to enable appropriate partial-volume and cerebrospinal fluid correction. 76 For studies targeting inhibitory neurotransmission, spectral editing techniques such as MEGA PRESS should be incorporated as a secondary acquisition when scan time allows, 76 in order to improve detection and quantification reliability of GABA. Longitudinal designs are especially important when examining neurochemical dynamics across ictal and interictal phases. Such approaches strengthen causal inference by distinguishing state-dependent changes from stable interictal traits.
Emerging approaches, including functional MRS, 78 offer the potential to capture dynamic neurochemical responses to migraine-relevant stimuli, providing a complementary perspective to static measurements and advancing mechanistic understanding of migraine attacks.
Conclusions
Current MRS evidence indicates that migraine attacks might be associated with metabolic alterations pointing towards increased cerebral energy demand, rather than reproducible changes in excitatory or inhibitory neurotransmission. However, the available findings are characterized by substantial methodological differences, limited sample sizes, and, in some cases, suboptimal analytical approaches, warranting caution in the data interpretation. Future longitudinal studies using standardized MRS protocols are essential to delineate temporal metabolic dynamics during attacks and to clarify their mechanistic and translational significance.
Key findings
Only limited data is available on neurometabolic alterations during the ictal phase of migraine, and existing investigations have predominantly been confined to the occipital cortex.
Findings have been inconsistent, with some studies suggesting increased cerebral energy metabolism; however, these observations have not been systematically replicated.
Rigorous longitudinal magnetic resonance spectroscopy studies are warranted to clarify neurometabolic dynamics of the migraine attack, particularly within unexplored pain-processing regions.
Supplemental Material
sj-docx-1-cep-10.1177_03331024261441576 - Supplemental material for Magnetic resonance spectroscopy during migraine attacks: A systematic review
Supplemental material, sj-docx-1-cep-10.1177_03331024261441576 for Magnetic resonance spectroscopy during migraine attacks: A systematic review by Delia-Ioana Radutiu, Edina Szabo, Rune Häckert Christensen, Eva-Maria Ratai, Nouchine Hadjikhani, Haidar Muhsen Al-Khazali, Messoud Ashina and Håkan Ashina in Cephalalgia
Footnotes
Acknowledgements
Not applicable.
ORCID iDs
Ethical considerations
Not applicable for reviews
Consent for publishing
All authors have read and approved the final version of the manuscript and agree to its submission to and publication in Cephalalgia.
Consent to participate
Not applicable
Author contributions
Delia-Ioana Radutiu: Conceptualization, Methodology, Formal analysis, Data curation, Writing – original draft, Writing – review & editing. Edina Szabo: Methodology, Data curation, Formal analysis, Writing – review & editing. Rune Häckert Christensen: Conceptualization, Methodology, Supervision, Writing – review & editing. Eva-Maria Ratai: Writing – review & editing. Nouchine Hadjikhani: Writing – review & editing. Haidar Muhsen Al-Khazali: Writing – review & editing. Messoud Ashina: Conceptualization, Supervision, Funding acquisition, Writing – review & editing. Håkan Ashina: Conceptualization, Methodology, Supervision, Funding acquisition, Project administration, Writing – review & editing.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Funding was provided by the Danish National Research Foundation (DNRF188 to MA) and Lundbeck Foundation (R310-2018-3711 to MA; R403-2022-1352 and R481-2024-1392 to HA). The funding sources were not involved in the design or conduct of the study; in the collection, analysis, or interpretation of the data; or in the preparation, review, or approval of the manuscript.
Declaration of conflicting interests
RHC has received personal speaker fees from AbbVie, Pfizer, Lundbeck, and Teva, outside of the submitted work, and serves as Section Editor for the Journal of Pain Research. E.M.-R. declares payments from the NIH (R01 DA047088, R01 NS124065), the Baszucki Group Foundation (2023-005) (funding), and Aletheia (consulting fees), and is an advisor for BrainSpec. HMA has received personal speaker fees from AbbVie, Pfizer, and Lundbeck, outside of the submitted work. MA has also received institutional research grants from the Danish National Research Foundation, Novartis, Novo Nordisk Foundation, and Lundbeck Foundation. MA is also an Associate Editor of Brain and The Journal of Headache and Pain. HA has received personal fees from AbbVie, Lundbeck, Pfizer and Teva, outside of the submitted work. HA is also an Editorial Board Member of The Journal of Headache and Pain. The remaining authors declare no conflicts of interest.
Data availability statement
Upon reasonable request, the corresponding author will provide the necessary data and materials to interested researchers for the purpose of academic scrutiny, reproducibility, and further scientific investigation.
Open practices
Not applicable
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.