
Abstract
Background
Migraine attacks are often triggered by normally innocuous stimuli, suggesting that sensitization within the nervous system is present. One mechanism that may contribute to neuronal sensitization in this context is translation regulation of new protein synthesis. The goal of this study was to determine whether protein synthesis contributes to behavioral responses and priming in preclinical models of migraine.
Methods
Mice received a dural injection of interleukin-6 in the absence or presence of the protein synthesis inhibitor anisomycin or the translation initiation inhibitor 4EGI-1 and were tested for facial hypersensitivity. Upon returning to baseline, mice were given a second, non-noxious dural injection of pH 7.0 to test for priming. Additionally, eIF4ES209A mice lacking phosphorylation of mRNA cap-binding protein eIF4E received dural interleukin-6 or were subjected to repeated restraint stress and then tested for facial hypersensitivity. After returning to baseline, mice were given either dural pH 7.0 or a systemic sub-threshold dose of the nitric oxide donor sodium nitroprusside and tested for priming.
Results
Dural injection of interleukin-6 in the presence of anisomycin or 4EGI-1 or in eIF4ES209A mice resulted in the partial attenuation of acute facial hypersensitivity and complete block of hyperalgesic priming. Additionally, hyperalgesic priming following repeated restraint stress was blocked in eIF4ES209A mice.
Conclusions
These studies show that de novo protein synthesis regulated by activity-dependent translation is critical to the development of priming in two preclinical models of migraine. This suggests that targeting the regulation of protein synthesis may be a novel approach for new migraine treatment strategies.
Introduction
A distinct characteristic of migraine patients is their enhanced sensitivity to innocuous stimuli, which can trigger and exacerbate a migraine headache (1,2). In preclinical models of migraine, repetitive stimulation of dural afferents not only contributes to peripheral plasticity, but also to central plasticity at synapses in the trigeminal nucleus caudalis (TNC), which is thought to account for the cutaneous facial allodynia present in humans during attacks (3,4). Activation of meningeal afferents results in the release of numerous pro-inflammatory cytokines, growth factors, excitatory neurotransmitters, and neuropeptides from primary sensory neurons, degranulated mast cells, and dural fibroblasts (5–8). Many of these endogenous factors, including IL-6 and calcitonin gene-related peptide (CGRP), directly contribute to the sensitization of primary sensory neurons as well as their downstream targets in the TNC (6,8–13). Based on this, mechanisms underlying peripheral and central sensitization have been proposed to contribute to these symptoms.
Repeated or intense noxious stimulation can cause maladaptive changes in synaptic plasticity within nociceptive circuits, leading to peripheral and central sensitization (14). Nascent protein synthesis in response to noxious stimuli can induce long-term changes in nociceptor activity and gene expression that can lead to the development of chronic pain (15–19). Additionally, regulation of nascent protein synthesis via activity-dependent translation has been shown to be a highly critical molecular event for neuroplasticity and plays a key role in changing nociceptor functionality (17,20–23). Activity-dependent translation can be induced by various endogenous compounds and membrane receptors and is regulated via the mammalian/mechanistic target of rapamycin complex 1 (mTORC1) and the extracellular-signal-regulated kinase (ERK) pathways, which converge on eukaryotic initiation factor 4E (eIF4E) of the eIF4F translation pre-initiation complex. Interestingly, in the dorsal root ganglion and dorsal horn of the spinal cord, these pathways are robustly activated following peripheral nerve inflammation and nerve injury and have been demonstrated as being essential for the persistence of chronic pain. For example, inhibition of mTORC1, activity-dependent translation, or general translation by local administration of rapamycin, 4EGI-1, or anisomycin, respectively, reduces mechanical hypersensitivity and injury-induced changes in nociceptor excitability, further implicating mechanisms of translation in the maintenance of pain (22,24).
Despite the overwhelming evidence for a role of translational regulation in chronic pain, no studies to date have examined this mechanism in the development and persistence of migraine headache. Here, using a preclinical model of dural stimulation and a model of repeated stress-induced hypersensitivity, we demonstrate a critical role for de novo protein synthesis in the behavioral responses and priming in these models.
Materials and methods
Experimental animals
Male and female eIF4ES209A mice on a C57BL/6 background were generated in the Sonenberg laboratory at McGill University as previously described (25) and bred at The University of Texas at Dallas to generate experimental animals. These animals were genotyped using DNA from tail clips taken at the time of weaning and were backcrossed to C57BL/6 background for at least 10 generations before experiments. In experiments involving transgenic mice, the control mice used were wild-type (WT) mice generated in the UT Dallas breeding colony from crossings of heterozygous eIF4ES209A mice with WT C57BL/6 mice (Envigo). Male and female ICR (CD-1) mice were outbred and purchased from Envigo. All behavior experiments were performed using mice aged 6–8 weeks (∼25–30 g) at the start of the experiment. All mice were housed on 12 h light/dark cycles with lights on at 7:00 AM. All mice were housed in groups of four animals per cage and had food and water available ad libitum. All behavioral experiments were performed between the hours of 9:00 AM and 5:00 PM. Mice were randomized to groups from multiple cages and investigators were blinded to treatment groups in all experiments. All animal procedures were approved by the Institutional Animal Care and Use Committees at The University of Texas at Dallas and were performed in accordance with the ARRIVE guidelines as well as the policies of the International Association for the Study of Pain and the National Institutes of Health guidelines for animal research.
Drugs and antibodies
Human recombinant IL-6 protein (R&D Systems) stock solution (100 µg/mL) was prepared in sterile 0.1% BSA and diluted to 1 ng/µL in synthetic interstitial fluid (SIF) consisting of 135 mM NaCl, 5 mM KCl, 10 mM HEPES, 2 mM CaCl2, 10 mM glucose, 1 mM MgCl2 (pH 7.4, 310 mOsm). Anisomycin (Tocris) stock solution (135 µg/µL) and 4EGI-1 (Enzo) stock solution (20 µg/µL) were both prepared in sterile 0.1% BSA and diluted to 5 µg/µL in SIF. Sodium nitroprusside (SNP) (Sigma-Aldrich) was prepared in sterile phosphate buffered saline (PBS) at the time of use and was kept away from light. For dural injections, mice received 5 µL injections of either IL-6, anisomycin, 4EGI-1, SIF, or a combination of IL-6 and anisomycin or IL-6 and 4EGI-1 for acute testing. For testing the ability of mice to prime to the initial stimulus in these dural injection experiments, 5 µL of SIF pH 7.0 were administered onto the dura. In repeated restraint stress experiments, which used SNP to test priming, a subthreshold dose of 0.1 mg/kg of SNP was administered intraperitoneally as a 150 µL injection. For western blotting experiments, p-eIF4E (Cell Signaling #9741S) and total-eIF4E (Cell Signaling #9742S) antibodies were used for primary incubation.
Mouse dural injections
Mouse dural injections were performed as previously described (13). Mice were anesthetized under isoflurane for <2 min with <2.5–3% isoflurane via a chamber. While anesthetized, treatments were injected in a volume of 5 µL via a modified internal cannula (Invivo1, part #8IC313ISPCXC, Internal Cannula, standard, 28 gauge, fit to 0.5 mm). The inner projection of the cannula was used to inject through the soft tissue at the intersection of the lambdoidal and sagittal sutures. The length of the projection was adjusted, using calipers, to be from 0.5 to 0.7 mm based on the animal weight (25–30 g) so as to not puncture the dura. Control mice received a 5 µL dural injection of SIF (pH 7.4, 310 mOsm).
Repeated restraint stress
Mice were stressed as previously described (26). Mice were placed right-side up into tail vein injection tubes (Stolting #51338) with the nose through the provided breathing hole and the tail through the slotted tail piece. The slotted tail piece was tightened so as to prevent the mouse from rotating in the tube, but loose enough to allow the animal to breathe. Mice were stressed between the hours of 10:00 AM to 12.00 PM for 2 h per day for three consecutive days. Control mice were placed into a separate room and deprived of food and water for the same 2-h interval for three consecutive days.
Von Frey testing
Mice were conditioned for five continuous minutes by handling, 24 h before habituation. Mice were habituated to paper cups (Choice 4 oz paper cups: 6.5 cm top diameter, 4.5 cm bottom diameter, 72.5 cm length) while in testing chambers for 2 h per day and for at least 2 days before measuring a baseline (13). Each mouse typically used their same assigned paper cup for the remainder of the experiment. Animals were given food while in testing chambers to allow for testing as previously described. Filament thresholds were determined using the Dixon “up-and-down” method. Testing in mice began with 0.07 g on the face and increased in weight to a maximum of 0.6 g on the face. The testing timelines for dural injection experiments and stress experiments were conducted as previously described in (13) and (26), respectively. In both experimental paradigms, once the mice returned to baseline, a sub-threshold dose of compound was administered either onto the dura (pH 7.0) or intraperitoneally (sodium nitroprusside). Mice were then tested for the ability of the initial stimulus to cause priming to the sub-threshold stimulus. All investigators were blinded to experimental conditions.
Western blotting
Female mice were used for all western blotting experiments and were killed by decapitation following anesthesia with tissues being flash frozen on dry ice. Frozen tissues were homogenized using a pestle in lysis buffer (50 mM Tris, pH 7.4, 150 mM NaCl, 1 mM EDTA, pH 8.0, and 1% Triton X-100) containing protease and phosphatase inhibitors (Sigma-Aldrich) and then sonicated for 10 sec. TG and dura tissues were harvested following 3 days of repeated restraint stress at the time-points provided in the Results section. To clear debris, samples were centrifuged at 14,000 rpm for 15 min at 4°C. 15 µg of protein was loaded into each well and separated by a 10% SDS-PAGE gel. Proteins were transferred to a 0.45 PVDF membrane (Millipore) at 30 V overnight at 4°C. Membranes were then blocked with 5% non-fat dry milk in 1× Tris buffer solution containing Tween 20 (TTBS) for 2 h. Membranes were washed in 1× TTBS three times for 5 min each then incubated with primary antibody overnight at 4°C. The following day, membranes were washed three times in 1× TTBS for 5 min each then incubated with the goat anti-rabbit secondary antibody (Jackson Immunoresearch) at room temperature for 1 h. Membranes were then washed with 1× TTBS six times for 5 min each. Signals were detected using Immobilon Western Chemiluminescent HRP Substrate (Millipore). Bands were visualized with a Bio-Rad ChemiDoc Touch and over-saturated pixels were excluded from the final analysis. Blots were first probed for phosphorylated eIF4E (peIF4E) (Cell Signaling; 1:3000), then stripped and re-probed for total eIF4E (teIF4E) (Cell Signaling; 1:3000). Equal loading was verified using GAPDH (Cell Signaling; 1:5000) as a control. For quantitative analysis, peIF4E was normalized to teIF4E. Analysis was performed using Image Lab version 6.0.1.
Experimental design and statistical analysis
We used only female mice in the dural IL-6 experiments involving ICR mice, given the higher frequency of migraine among women. The rationale for this was based on pilot studies in which co-injection of dural IL-6 with either anisomycin or 4EGI-1 revealed no sex differences in these animals. Additionally, previously reported findings from our laboratory revealed similar effects in males when IL-6 is co-injected with anisomycin or 4EGI-1 into the hindpaw (27). Since there have been no comparable studies in eIF4ES209A mice, we used both females and males to explore the possibility of a sex difference in this genotype with these stimuli. All behavioral data are represented as individual data points with means (lines). Western blot data are represented as means ± SEM. Behavioral data were analyzed for multiple comparisons at each time point via two-way ANOVA and Bonferroni’s post-hoc test. F-values for each analysis are presented (Table 1). Student’s unpaired two-tailed t-test was used for individual mean comparisons when appropriate. Data analysis was performed using Prism version 8.3 for Mac OS X. Significance was set at p < 0.05 for all analyses. Power analysis was performed using G power for comparison of the means between groups using expected effect sizes based on pilot studies and previously published data in other models (24). All investigators were blinded to genotype and treatment during testing and scoring. Each experiment was independently replicated twice.
F-values obtained from two-way ANOVA analysis comparing mean effects within rows are presented for each figure.

Results
Priming induced by dural IL-6 is blocked by general protein synthesis inhibition
Previously, our laboratory has demonstrated the ability of dural IL-6 to sensitize mice to cutaneous mechanical stimulation following a sub-threshold stimulus in a model of hyperalgesic priming (13). Given recent data supporting a role for general protein synthesis in the development of hyperalgesia via DRG and spinal pathways (28,29), and given the potential greater dependence on translation regulation signalling for sensitization in TG versus DRG neurons (30), we tested the hypothesis that protein synthesis is necessary for facial sensitization in this dural stimulation priming model. We first administered either 0.1 ng of IL-6, 25 µg of anisomycin (a general protein synthesis inhibitor), or a co-injection of both onto the dura of female mice to induce mechanical facial allodynia that persisted for more than 24 h and resolved by 72 h (Figure 1). Upon returning to baseline, we administered a second stimulus, SIF (pH 7.0), onto the dura to reveal the presence of hyperalgesic priming from the initial IL-6 stimulus. Although there were only minor differences in acute mechanical hypersensitivity, mice that were initially administered only IL-6 exhibited robust facial hypersensitivity when exposed to low pH. This hypersensitivity persisted for more than 5 h. Conversely, mice that received a co-injection of both IL-6 and anisomycin did not respond to the low pH, suggesting that general protein synthesis is required for the generation of a primed state in mice.

Dural co-injection of IL-6 with the general protein synthesis inhibitor anisomycin blocks hyperalgesic priming to dural pH 7.0 in female ICR mice (n = 10 for all groups). Analysis of groups was performed using two-way ANOVA followed by Bonferroni post-hoc test (see Table 1 for F-values).
Inhibiting cap-dependent translation prevents long-lasting facial hypersensitivity
Previous reports have demonstrated that cap-dependent translation induced by IL-6 is dependent on the binding of eIF4E/eIF4G to induce eIF4F complex formation and interruption of this binding via stabilizing 4E-BP1 to eIF4E prevents the priming induced by pronociceptive mediators (27). Translation control by 4E-BP1 has been shown to regulate mechanical hypersensitivity and genetic loss of 4E-BP1 increases excitatory synaptic transmission in the spinal cord, thereby enhancing mechanical nociception (31). To gain a better understanding of the molecular mechanisms underlying the priming induced by IL-6, we sought a similar approach to investigate the role of eIF4E phosphorylation and subsequent cap-dependent translation. Similar to the experiments in Figure 1, we administered 0.1 ng of IL-6, 25 µg of 4EGI-1, a compound that mimics the activity of 4E-BP1, or a co-injection of both onto the dura of female mice (Figure 2). The resulting mechanical allodynia induced by IL-6 persisted for more than 24 h and resolved after 72 h. Upon returning to baseline, mice were exposed to a dural injection of low pH. Mice previously treated with IL-6 alone demonstrated cutaneous mechanical hypersensitivity. As with anisomycin, acute facial hypersensitivity was partially attenuated and the response to low pH was robustly blocked in mice that received a co-injection of IL-6 and 4EGI-1. Thus, assembly of the eIF4F complex locally in the dura appears to be critical to the development of long-lasting mechanical hypersensitivity following repeated stress.

IL-6-induced priming to pH 7.0 is blocked by co-treatment with 4EGI-1 in female ICR mice (n = 8 for all groups). Analysis of groups was performed using two-way ANOVA followed by Bonferroni post-hoc test (see Table 1 for F-values).
eIF4E phosphorylation is necessary for priming following repeated stress
Previous studies have shown that eIF4ES209A mice, which lack phosphorylation of eIF4E at Ser209, exhibit reduced nociceptor sensitization and hyperalgesic priming in response to pro-nociceptive and inflammatory factors, including NGF and IL-6 (24,32). To test the hypothesis that eIF4E phosphorylation is important for priming following dural stimulation or repeated stress, we utilized these same eIF4ES209A in these models. As previously described, we once again administered IL-6 onto the dura of female and male eIF4ES209A mice, followed by dural pH 7.0 (Figure 3). Unlike WT mice, acute mechanical hypersensitivity was partially attenuated and priming to dural pH 7.0 was blocked in both sexes of eIF4ES209A mice, supporting our hypothesis that eIF4E phosphorylation is critical to establishing long-lasting hypersensitivity induced by dural IL-6.

Female (a) and male (b) eIF4ES209A mice have decreased mechanical hypersensitivity to dural IL-6 and do not prime to dural pH 7.0. For (a) n = 8 for WT groups; n = 7 for 4EKI/SIF and n = 8 for 4EKI/IL-6. For (b) n = 8 for WT/SIF and 4EKI/IL-6; n = 7 for WT/IL-6 and 4EKI/SIF. Analysis of groups was performed using two-way ANOVA followed by Bonferroni post-hoc test (see Table 1 for F-values).
We recently showed that mice exposed to repeated restraint stress become primed to sub-threshold doses of the nitric oxide-donor sodium nitroprusside (SNP). Stress is the number one reported trigger of migraine among humans (33) and NO donors are among the most reliable experimental triggers of migraine attacks (34,35). We subjected both WT and eIF4ES209A mice to repeated restraint stress as previously described. Following their return to baseline nociceptive thresholds, we administered a 0.1 mg/kg IP injection of SNP (Figure 4). Interestingly, priming to SNP was completely blocked in stressed eIF4ES209A mice compared to a robust increase in mechanical hypersensitivity in stressed WT mice. This identifies a key role for eIF4E phosphorylation in the transition from acute to long-lasting hypersensitivity and suggests eIF4E phosphorylation as a potential mechanism underlying neuronal plasticity in migraine.

Female (a) and male (b) eIF4ES209A mice exhibit acute mechanical hypersensitivity similar to WT mice following repeated restraint stress but fail to prime to a sub-threshold dose of the nitric oxide donor SNP (n ≥ 6). For (a), n = 6 for all groups except 4EKI/Stress, for which n = 7. For (b), n = 6 for all groups. Analysis of groups was performed using two-way ANOVA followed by Bonferroni post-hoc test (see Table 1 for F-values).
Phosphorylation of eIF4E is differentially regulated after repeated restraint stress
In an effort to determine the time points at which eIF4E phosphorylation is altered in the stress paradigm, we examined protein lysates from the dura and TG of WT C57BL/6 mice at multiple time points after day 3 of the stress protocol (Figure 5). Our data indicate an almost 50% increase in the expression level of p-eIF4E in the TG 1 h after day 3 of stress, an effect that was diminished by 3 h. In the dura, the effects were completely opposite, with an almost 50% decrease in p-eIF4E expression levels 1 h after stress, but an increase of over 75% in expression levels by 3 h. Phosphorylation levels of eIF4E were decreased 24 h after day 3 of stress in both tissues. Although the implications of these data remain unclear, they indicate that eIF4E phosphorylation-mediated events occur earlier in the TG than they do in the dura in response to stress, providing evidence that the temporal components of this key biochemical event are dynamically regulated.
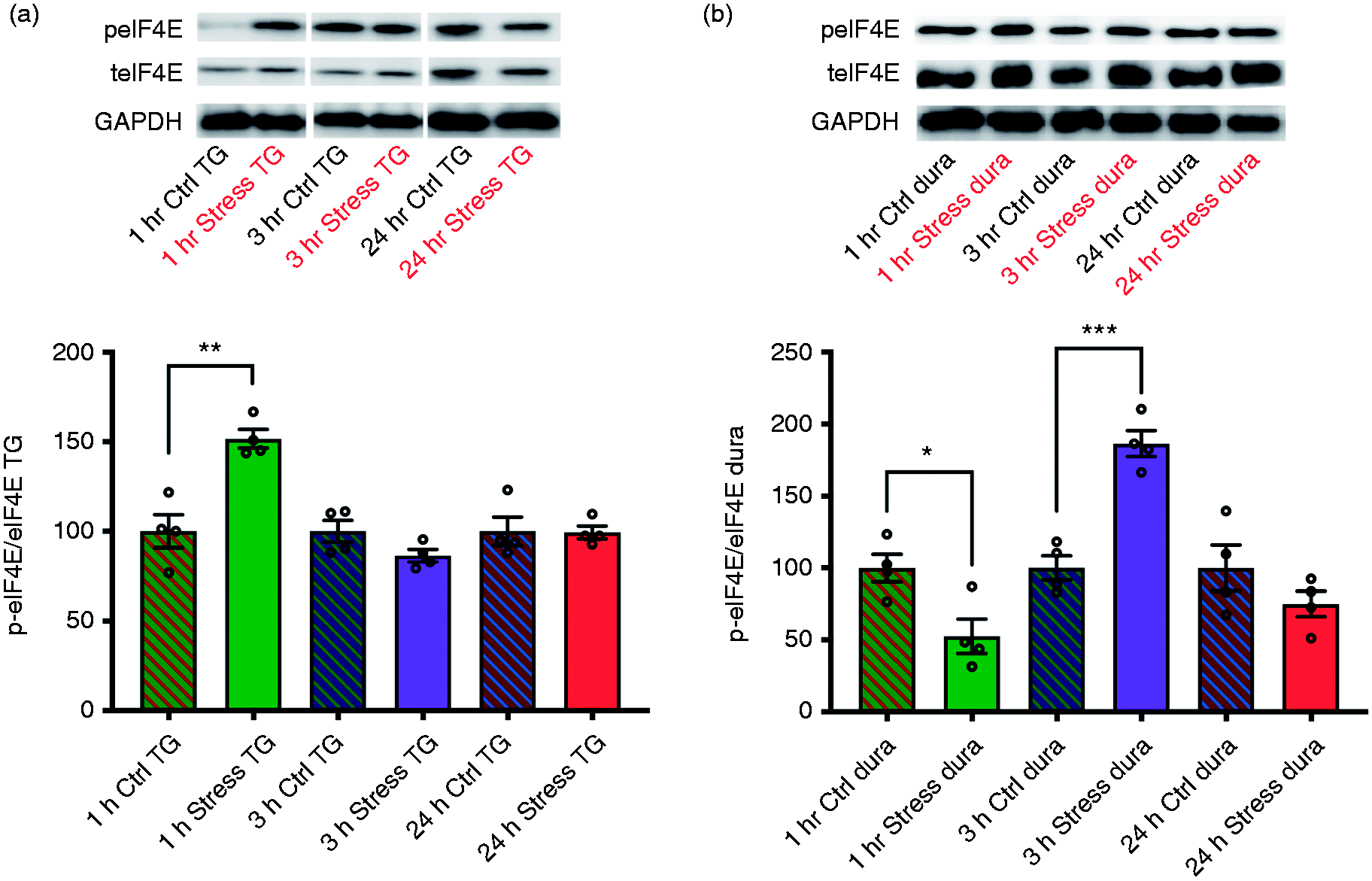
Compared to controls, phosphorylation of eIF4E is robustly increased in the TG of WT C57BL/6 mice at 1 h following repeated restraint stress, an effect that is diminished by 3 h (a). In contrast, decreases in eIF4E phosphorylation initially occur in the dura before robustly increasing to peak levels by 3 h, indicating that eIF4E is differentially regulated in the TG and dura following stress (n = 4 mice pooled for each time point) (b). GAPDH was used as a loading control. Significance between treatments was determined via Student’s unpaired two-tailed t-test. Data are represented as means ± SEM.
Discussion
The susceptibility of migraine patients to attacks following exposure to normally innocuous triggers strongly implicates sensitization of the trigeminovascular system, leading to lower activation thresholds for nociception to occur. Robust changes in gene expression regulated by activity-dependent translation are key to nociceptor plasticity and phenotypic alterations and are thought to underlie neuronal sensitization (29). IL-6-induced phosphorylation of ERK has been shown to modulate the sodium channel Nav1.7, resulting in prolonged migraine-related pain (36) and activation of ERK/MAPK pathways can ultimately lead to changes in transient receptor potential (TRP) channel function, a family of proteins which have been implicated in migraine pathophysiology (37–39). Co-application of dural IL-6 and anisomycin partially attenuated the resulting acute facial hypersensitivity and completely blocked the development of a primed state in mice, suggesting that protein synthesis may be critical for the development of long-lasting mechanical hypersensitivity. The comparable results between local administration of anisomycin and 4EGI-1 suggest that the newly synthesized proteins required to establish priming are regulated by cap-dependent translation. In accordance with this, our results from injecting IL-6 onto the dura of eIF4ES209A mice suggest that eIF4E phosphorylation is key for long-lasting facial mechanical hypersensitivity. Thus, similar to reports in other preclinical models of pain (24), activity-dependent translation appears to be critical for the sensitization of trigeminal nociceptors by pronociceptive factors.
Headaches can be triggered in migraine patients by a wide range of noxious and innocuous stimuli, making it difficult to parse out which mechanisms may be most relevant for trigeminal activation and sensitization. Since the most common and frequent trigger of migraine is stress (33), use of this trigger is one of the more clinically relevant models for mechanistic investigation. Utilizing our novel repeated stress model with eIF4ES209A mice allowed us to gauge the role of eIF4E phosphorylation in a much more robust model of hypersensitivity that was not isolated to local cephalic regions, but rather the entire body. In eIF4ES209A mice, the loss of NO-donor induced priming normally observed after stress highlights a critical role for eIF4E phosphorylation in the development of long-lasting mechanical hypersensitivity. Additionally, in WT mice, phosphorylation of eIF4E in the dura and TG was differentially expressed across multiple time points following stress, with an increase in p-eIF4E occurring 1 h after stress in the TG and being downregulated by 3 h, while the opposite effect was observed in the dura. While the significance of these data remains unclear, our data indicate that robust changes in eIF4E phosphorylation occur after repeated stress and that a genetic loss of this phosphorylation prevents the development of hyperalgesic priming. Additionally, the experiments in this study are helping to establish the sample sizes needed for significance based on effect sizes in the stress model.
Targeting local translation in DRG sensory axons has been proposed as a potential treatment for many types of inflammatory and neuropathic pain (23); however, whether the translation of mRNAs that contribute to priming of dural afferents occurs locally in meningeal sensory axons or at distal sites in the TG remains unclear. A previous study from our lab found that while local co-injection of IL-6 with anisomycin into the hindpaw prevented acute mechanical hypersensitivity, co-injection with a transcription inhibitor had no effect, suggesting that local translation contributing to acute pain in the hindpaw is dependent on pre-existing pools of mRNA (27). Given the similarities between the DRG and TG, it is plausible to suggest that the translation events regulating acute and long-lasting hypersensitivity caused by dural IL-6 are dependent on local pools of mRNA as well; however, one key difference between these studies is that attenuation of dural IL-6-induced facial allodynia begins at later time points in the acute phase. One potential explanation for this can be attributed to the location of testing following injection. For example, other studies tested the hindpaw in the same location that the injection was given; here, we inject directly onto the dura, but test the periorbital region of the face. The delay in attenuation of acute hypersensitivity might indicate that nascent protein synthesis in response to dural stimulation occurs some distance away from the initial injection site. Additionally, if these translation events are indeed occurring locally in meningeal sensory axons, then changes in synaptic plasticity in the TG and possibly even the ophthalmic nerve (innervating the periorbital region) may be dynamically and temporally regulated following injection.
Further studies will be necessary to determine both the location and identity of the mRNAs that are translated to mediate long-lasting sensitization of the trigeminal pathway. Recently, studies have demonstrated robust increases in the expression levels of mRNAs that modulate acid-sensing ion channels (ASICs) (40) and TRP channels (41) in the TNC in response to noxious odors and repeated dural stimulation, respectively. Although it is currently unknown which eIF4E-dependent mRNAs are most critical in these models, eIF4E phosphorylation has been shown to regulate the translation of brain-derived neurotropic factor (BDNF) mRNA in mouse DRG (32). BDNF is a key player in the maintenance of long-term potentiation (LTP) (42–44) and has been implicated in maintaining persistent pain states (45,46). Additionally, multiple lines of evidence suggest a role for BDNF in headache (47–50). In support of these claims, our lab has recently demonstrated that afferent input from the meninges is capable of producing BDNF-dependent priming of the trigeminovascular system (51).
Although the exact mechanism is still unclear, our findings are the first to demonstrate that de novo protein synthesis regulated by activity-dependent translation is critical to the development of cutaneous facial hypersensitivity following dural stimulation or repeated stress. Further exploration of how these translation pathways as well as other modes of translation contribute to trigeminal sensitization may provide additional insight into how to develop more efficient therapies for migraine.
Key findings
Local inhibition of de novo protein synthesis prevents the long-lasting facial hypersensitivity that is normally observed following noxious stimulation of the dura mater. Persistent facial hypersensitivity induced by noxious stimulation of the dura mater is dependent on the association of eIF4G with eIF4E and subsequent initiation of cap-dependent translation. The translational regulation of protein synthesis via eIF4E phosphorylation is critical for the development of a primed state following dural stimulation or repeated stress. Phosphorylation of eIF4E is differentially regulated across time points in the dura mater and trigeminal ganglia following repeated restraint stress.
Footnotes
Acknowledgments
We thank Meghna Suresh, Brian Nguyen, and Nikhil Gogineni for their help with the mouse behavior as well as in data acquisition and analysis.
Declaration of conflicting interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: GD and TJP are co-founders of CerSci Therapeutics and Ted’s Brain Sciences. Additionally, TJP is a co-founder of 4E Therapeutics.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: These studies were funded by the National Institutes of Health (NS072204, NS104200, NS104990) and The University of Texas System.