
Abstract
Background
Non-invasive stimulation of the vagus nerve has been proposed as a new neuromodulation therapy to treat primary headache disorders, as the vagus nerve is hypothesized to modulate the headache pain pathways in the brain. Vagus nerve stimulation can be performed by placing an electrode on the ear to stimulate the tragus nerve, which contains about 1% of the vagus fibers. Non-invasive vagus nerve stimulation (nVNS) conventionally refers to stimulation of the cervical branch of the vagus nerve, which is made up entirely of vagal nerve fibers. While used interchangeably, most of the research to date has been performed with nVNS or an implanted vagus nerve stimulation device. However, the exact mechanism of action of nVNS remains hypothetical and no clear overview of the effectiveness of nVNS in primary headache disorders is available.
Methods
In the present study, the clinical trials that investigated the effectiveness, tolerability and safety of nVNS in primary headache disorders were systematically reviewed. The second part of this study reviewed the central connections of the vagus nerve. Papers on the clinical use of nVNS and the anatomical investigations were included based on predefined criteria, evaluated, and results were reported in a narrative way.
Results
The first part of this review shows that nVNS in primary headache disorders is moderately effective, safe and well-tolerated. Regarding the anatomical review, it was reported that fibers from the vagus nerve intertwine with fibers from the trigeminal, facial, glossopharyngeal and hypoglossal nerves, mostly in the trigeminal spinal tract. Second, the four nuclei of the vagus nerve (nuclei of the solitary tract, nucleus ambiguus, spinal nucleus of the trigeminal nerve and dorsal motor nucleus (DMX)) show extensive interconnections. Third, the efferents from the vagal nuclei that receive sensory and visceral input (i.e. nuclei of the solitary tract and spinal nucleus of the trigeminal nerve) mainly course towards the main parts of the neural pain matrix directly or indirectly via other vagal nuclei.
Conclusion
The moderate effectiveness of nVNS in treating primary headache disorders can possibly be linked to the connections between the trigeminal and vagal systems as described in animals.
Keywords
Introduction
Headache is a common symptom and, collectively, headache disorders have a prevalence of 49% (1). Approximately 95% of the general population have experienced headache during some stage in their life (2). In addition to having a significant impact on patients’ daily life due to pain and disability, the socioeconomic burden of headaches are considerable, with a cumulative burden of $14 billion per year (3). Although the pathophysiological mechanisms involved remain partially elusive, chronic headache is believed to have a neurogenic basis (4–11). For example, a recent fMRI study showed that areas within the hypothalamus and brainstem, including the trigeminal system, were involved in mediating migraine (12). Although various treatments are available for alleviating pain in headache, the results are often unsatisfactory (13–15). Therefore, experimental interventions that treat headache disorders more effectively are being investigated and show promising results. One of the options includes (non-invasive) vagus nerve stimulation (nVNS), which was found to inhibit nociceptive behavior in animals (16–21). In humans, multiple clinical trials have been conducted, showing the clinical relevance of nVNS in treating chronic headache disorders (22–25). However, the underlying mechanism supporting these effects is difficult to elucidate (26–29).
Distribution of the types of afferents over the four vagus nuclei (adapted from Yuan and Silberstein 2016 (28)).
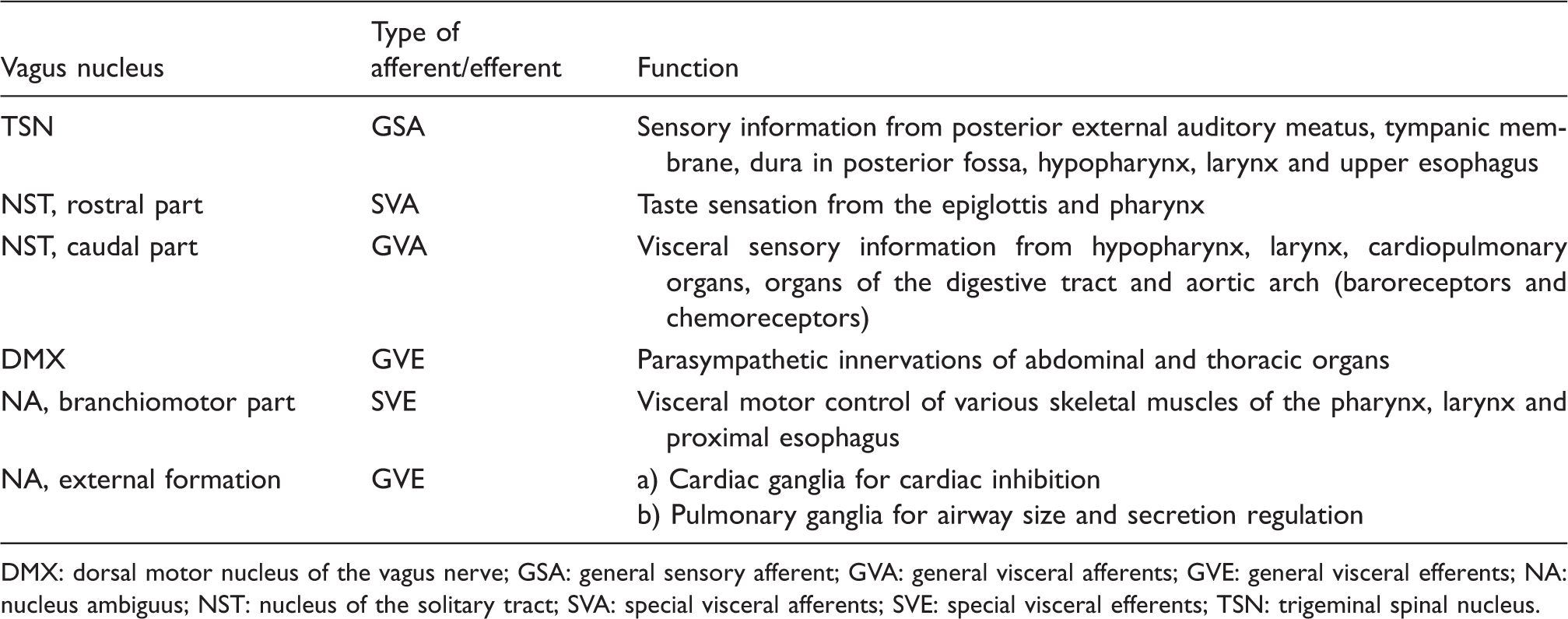
DMX: dorsal motor nucleus of the vagus nerve; GSA: general sensory afferent; GVA: general visceral afferents; GVE: general visceral efferents; NA: nucleus ambiguus; NST: nucleus of the solitary tract; SVA: special visceral afferents; SVE: special visceral efferents; TSN: trigeminal spinal nucleus.
Material and methods
Methodology for retrieval and assessment of nVNS trials
Quality assessment of the individual trials.

1. Was the method of randomization adequate?
2. Was the treatment allocation concealed?
3. Was the patient blinded to the intervention?
4. Was the care provider blinded to the intervention?
5. Was the outcome assessor blinded to the intervention?
6. Was the dropout rate described and acceptable?
7. Were all randomized participants analyzed in the group to which they were allocated?
8. Are reports of the study free of suggestion of selective outcome reporting?
9. Were the groups similar at baseline regarding the most important prognostic indicators?
10. Were co-interventions avoided or similar?
11. Was the compliance acceptable in all groups?
12. Was the timing of the outcome assessment similar in all groups?
+, criterion achieved; −, criterion not achieved; ∗, assessors initially disagreed.
High: > 75% of the criteria have been fulfilled (≥ 10/12). Where they have not been fulfilled, the conclusions of the study or review are thought very unlikely to have been altered.
Moderate: 50–75% of the criteria have been fulfilled (6–9/12). Those criteria that have not been fulfilled or not adequately described are thought unlikely to have altered the conclusions.
Low: Less than 50% of the checklist criteria were fulfilled (< 6/12). The conclusions of the study are thought likely or very likely to alter had those criteria been fulfilled.
Methodology for retrieval and assessment of anatomical papers
Regarding the anatomical review, various anatomical atlases and textbooks (31,32,38–42) were used to create a preliminary overview of vagus nerve anatomy and central connectivity. Thereafter, cross-referencing was carried out in order to enrich the anatomical results. PubMed, Medline, EMBASE and Google Scholar were searched using: [Vagus Nerve]; [Dorsal Motor Nucleus]; [Ambiguus Nucleus]; [Trigeminal Spinal Nucleus]; [Nucleus of the Solitary Tract]; [Anatomy]; [Neural Pathways]; [Afferent Pathways]; and [Efferent Pathways]. Medical subject headings were used to enrich the results. All searches were conducted until July 2018. Inclusion criteria were: a) Anatomy of the central portion of the vagus nerve was discussed; b) studies implemented techniques like tracing studies, dissections or neuroimaging; and c) investigation was carried out in either humans or animals. Exclusion criteria were: a) Articles were other than original research papers (i.e. reviews); b) articles were written in other languages than English, Dutch, German or French; c) articles investigated the peripheral portion of the vagus nerve; and d) articles investigated the anatomy by use of computational modeling. Each paper was reviewed using the inclusion and exclusion criteria by two investigators (BD, MvD and/or NV) independently. When in doubt, a third investigator (DH) was consulted. A total of 507 articles were retrieved, 358 of which remained after the removal of duplicates. Finally, 94 anatomical papers could be included based on the inclusion and exclusion criteria. Each paper was assessed by two researchers independently (BD, MvD, NV and/or DH). Findings were grouped per vagus nucleus, afferent pathway or efferent pathway and presented narratively and summarized in schematic figures.
Results
nVNS in primary headache disorders
Twelve clinical trials were included, representing a total of 866 patients who underwent the treatments as defined per protocol. Sex of the patients was reported in 836 cases (373 males; 44.6%). Primary headache disorders that were treated with nVNS constituted migraine (n = 469), cluster headache (n = 376), and other trigeminal autonomic cephalgias, including hemicrania continua (n = 13), paroxysmal hemicrania (n = 6) and short-lasting unilateral neuralgiform headache attacks with cranial autonomic symptoms (SUNA) (n = 2). In episodic migraine, it was found that nVNS elicited a significant pain reduction (22,43–47). One paper described a significant reduction of the number of headache days per month (44), although the number of headache days or tolerability of the attacks were not influenced by nVNS according to another paper (48). nVNS also has been described as capable of reducing the intake of rescue medication in cases of migraine attack (23). In cluster headaches, nVNS improved the mean overall condition of the patients (22,49), significantly reduced weekly attack frequency (50) and induced significantly higher pain-free rates with active nVNS compared with sham nVNS (24,50,51). In hemicrania continua, the majority of patients reported reduced severity of continuous pain (22,52). A more modest, but similar trend was observed in patients suffering from paroxysmal hemicrania (52). Neither of the patients suffering from SUNA benefited from nVNS therapy (22). The quality assessment of the individual trials is provided in Table 2. An extensive overview of the included papers is given in Supplemental Tables 1 and 2.
Anatomical characteristics of the vagal central connections
Central connections of the NST
The NST has been reported to receive afferents from the vagus nerve, the facial nerve, the trigeminal nerve and the glossopharyngeal nerve (53–55). Labeled fibers of the vagus nerve could be traced bilaterally to the NST and ipsilaterally to the external cuneate nucleus. However, the majority of the fibers that headed to the external cuneate nucleus continued and entered the solitary tract (56,57). Projections from the NST terminated in the vagal preganglionic motor neurons of the DMX and the NA (58,59). A prominent projection traversed towards the dorsomedial reticular formation and several medullary nuclei (i.e. the dorsal medullary raphe nuclei (60), the gigantocellular reticular nucleus and the paragigantocellular nucleus (60,61)). From the NST, fibers were observed to terminate in the dorsal or ventral part of the dorsal raphe nuclei, the locus coeruleus (A6 noradrenergic cell groups) (61), the A1 and A5 noradrenergic cell groups (61–63), the dorsal medial medulla (C3 adrenergic cell groups) (57), the lateral paragigantocellular reticular nucleus (C1 adrenergic cell groups) (61) and the medial and lateral parabrachial nuclei (58,59,64,65). Together with the parabrachial nuclei, the A1 and A5 catecholamine cell groups were reported to receive input from the NST and together they are believed to play a role as relay nuclei for sensory and gustatory information, which is processed in different areas of the forebrain. From the caudal part of the NST, projections arise that travel to the vagal preganglionic neurons in the DMX (66–68) and the NA (58,59,63,69), the main sites of vagal motor neurons. Each area that receives a direct projection from the NST, with the exception of the motor nuclei of the cranial nerves V, VII, X and XII, projects back to the DMX. In addition, projections have been described to the motor nuclei of the trigeminal, the facial and the hypoglossal nerves. These motor nuclei have an important role in the motor functions of the ingestive behavior (63,65). Furthermore, the trigeminal sensory nucleus complex (including the principal sensory nucleus and the TSN) has connections with the NST (57). Ipsilateral vestibular afferents were labeled and could be traced via two different routes: a) Fibers that traveled within the medial vestibular nucleus before entering the NST (this route is known as the lateral pathway), and b) fibers that traveled within the nucleus prepositus hypoglossi to its caudal part, whereupon they entered the NST (this route is known as the medial pathway). Besides, labeled fibers from the caudal part of the medial vestibular nucleus and the inferior vestibular nucleus could be traced bilaterally to the NST (70–72). Another tract originating from the NST occupies a region known as the intermediate reticular zone as it courses towards the intermediolateral nucleus (64). Finally, there is a group of projections that ascend from the NST to thalamic, hypothalamus and other limbic regions (e.g. the midline thalamic nuclei (73,74), the bed nucleus of the stria terminalis (59), the central nucleus of the amygdala (59), lateral hypothalamic area (75), paraventricular hypothalamic nucleus (59,63,76), the arcuate nucleus (73) and the medial preoptic area (75,77)). The aforementioned connections are depicted in Figure 1.
Schematic overview of the central projections of the NST. Latin numbers indicate cranial nerves or their corresponding nuclei; color of the tract is consistent with the color of the nucleus/area it sprouts from.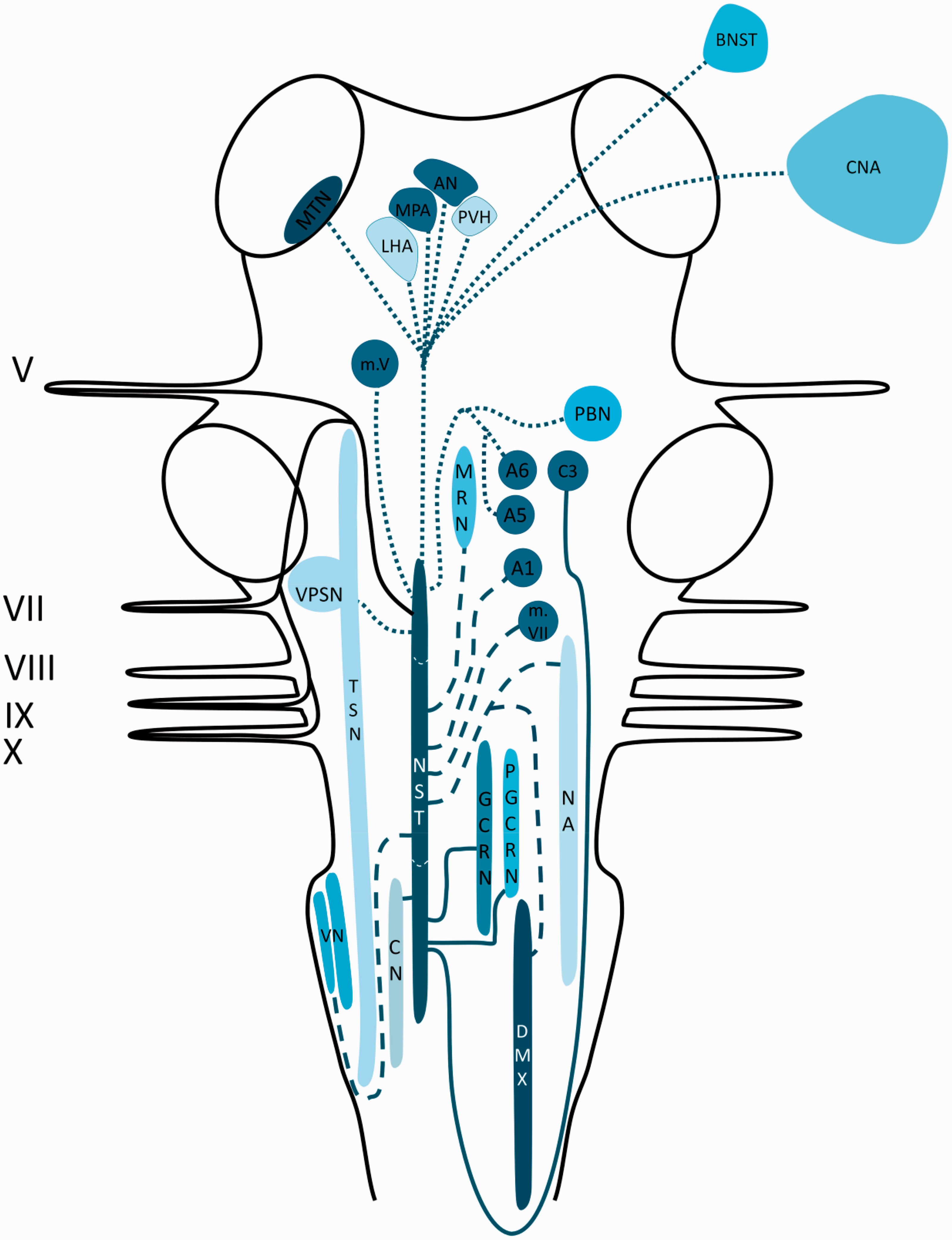
Central connections of the NA
Afferents were reported to terminate in the NA originate from the periaqueductal grey (PAG), the cuneiform nuclei, the red nucleus and the frontal cortex (78,79). Efferent projections to the area postrema, the contralateral NA and the lateral reticular formation were found (78–81). Furthermore, connections to the medial parts of the medulla oblongata and the parabrachial nuclei were described as well (78,79). The aforementioned connections are depicted in Figure 2.
Schematic overview of the central projections of the NA. Latin numbers indicate cranial nerves or their corresponding nuclei; color of the tract is consistent with the color of the nucleus/area it sprouts from.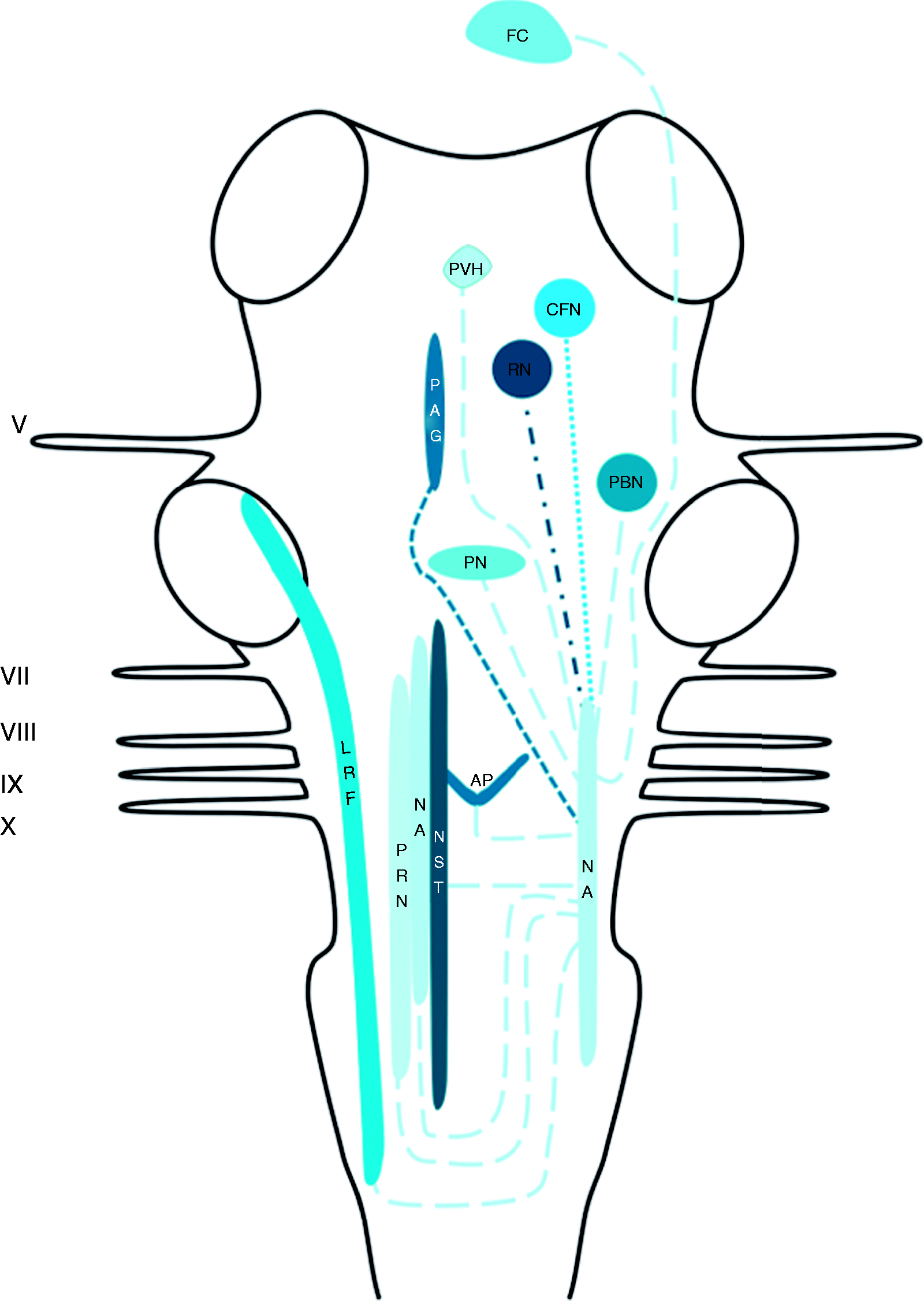
Central connections of the TSN
The trigeminal connections have been described in depth in a recent review by Henssen et al. (2016) (82). Afferents of the trigeminal, vagus, facial and glossopharyngeal nerves contribute to the trigeminal spinal tract and the TSN (53–55). The TSN can be subdivided into three subnuclei: The oral part, the interpolar part and the caudal part. The efferents from the oral part of the TSN project mainly to the contralateral thalamic subnuclei, the ventral posteromedial nucleus (VPM) in particular, via the ventral trigeminothalamic tract. However, the oral part of the TSN was also observed to have projections to lamina III and IV of the medullary dorsal horn (83). These connections were found to contribute to a minor ascending tract that terminates partially in the PAG. This tract is named the intranuclear pathway and it receives most of its input from the interpolar and caudal parts of the TSN (83–85). The interpolar part of the TSN also projects via the ventral trigeminothalamic tract to the contralateral VPM
85
. Lastly, the interpolar part of the TSN
56
, together with the caudal part of the TSN and the principal sensory nucleus, is known to have connections with the NST (54,57). The caudal part is the most investigated subnucleus of the TSN. The caudal part of the TSN was found to project to the contralateral VPM mainly, although a bilateral projection to the mediodorsal nucleus of the thalamus was also reported. Furthermore, connections between the principal sensory nucleus of the trigeminal nerve and the caudal part of the TSN were observed (86). The caudal part of the TSN was furthermore connected to the VPM and dorsomedial region of the thalamus in a bilateral fashion and to the contralateral posterior nucleus of the thalamus (87,88). As stated, the TSN contributes to the intranuclear pathway and therefore has extensive connections with the PAG and the NST (54,57,85). The aforementioned connections are depicted in Figure 3.
Schematic overview of the central projections of the TSN. Latin numbers indicate cranial nerves or their corresponding nuclei; color of the tract is consistent with the color of the nucleus/area it sprouts from.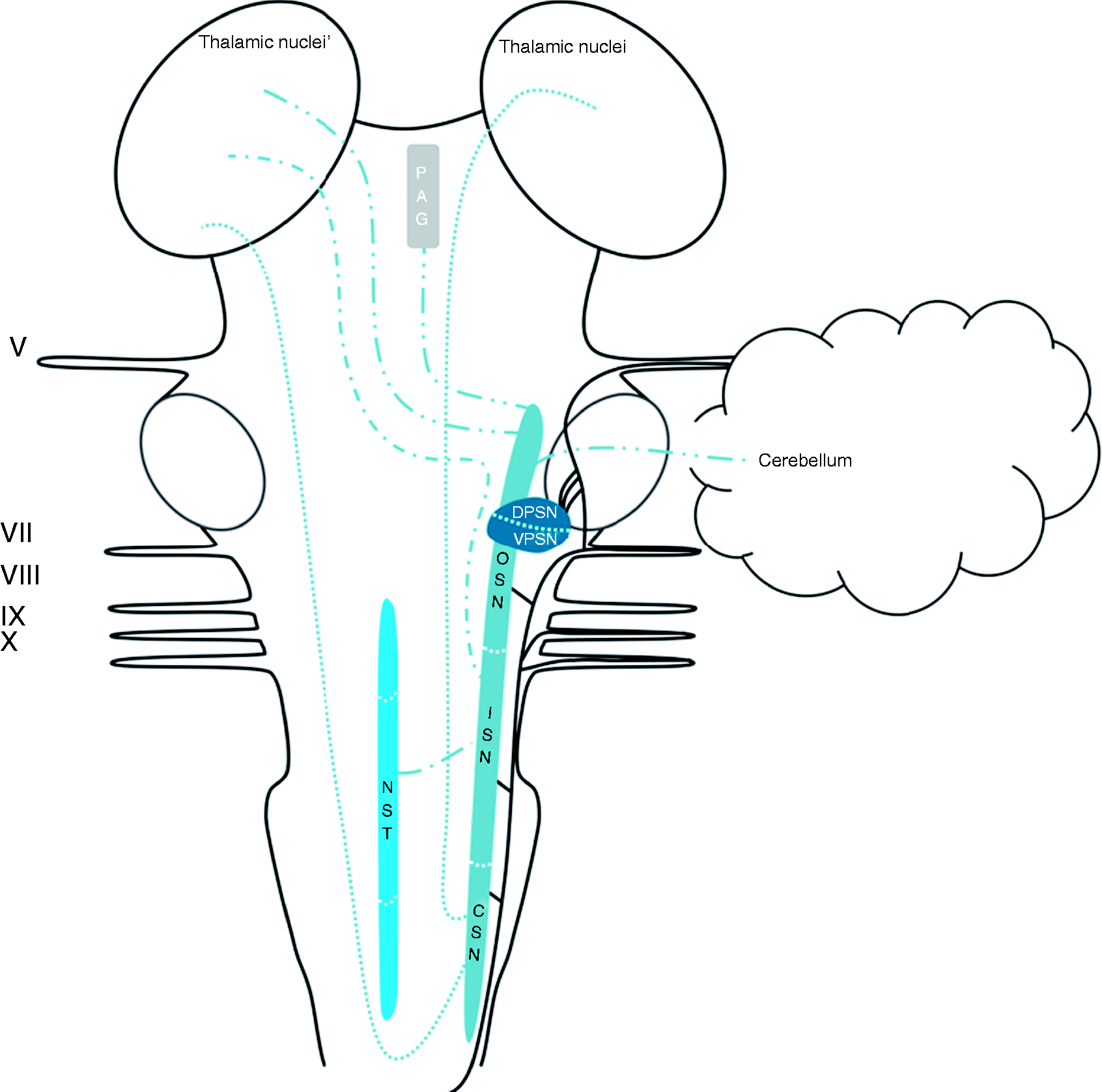
Central connections of the DMX
Rogers et al. (1980) reviewed afferent projections to the DMX in the late 20th century and described three major sources of afferent input to the DMX: The NST, the magnocellular paraventricular nucleus and several medullary nuclei (89). Projections of the NST to the DMX have been described by others as well (90,91). The paraventricular nucleus, the magnocellular neurons in particular, has been reported as a sprouting zone of a substantial amount of fibers that terminate in the DMX in rats (92). Concerning the medullary nuclei, the nucleus reticularis and the nucleus gigantocellularis have been shown to project to the DMX (89). Additionally, thyrotropin-releasing hormone-immunoreactive neurons residing in the nucleus raphe pallidus, nucleus raphe obscurus and the parapyramidal region of the ventral medulla have been suggested to project to the DMX and NST in rats (93). Furthermore, a fourth, minor, afferent pathway has been described by Berk and Finkelstein. They found that, in the pigeon, a minority of afferents of the nucleus periventricularis magnocellularis of the hypothalamus project to the ventral parvocellular subnucleus of the DMX (94). Zardetto-Smith et al. also found evidence in 1988 for a dynorphinergic innervation of the DMX by the perifornical nucleus, another hypothalamic area believed to be involved in the general arousal associated with emotive behaviours, as dynorphin has been implicated in mediating a wide variety of behavioral and autonomic-related processes including pain modulation (95). A fifth pathway was described by Ter Horst et al. in 1991, as they found projections of the locus coeruleus to the DMX in medial and rostral levels (96). In addition, a sixth projection system has been described. The ventral descending pathway, sprouting from the infralimbic cortex in the rat, projects to several autonomic cell groups in the brainstem, including the DMX (97). Furthermore, in 1996 Ruggiero et al. found that vestibular afferents reside in the NST and DMX (72). Earlier, in 1994, Balaban et al. studied central connectivity of the vestibular nuclei and found that some axons ended near somata of DMX neurons (70). Finally, studies conducted by Chiba et al. in 2001 and Kuipers et al. in 2006 found that the anterior cingulate cortex projects to the DMX in a bilateral fashion (98,99). Studies that investigated the morphological characteristics of the efferents from the DMX show that, together with the red nucleus, the DMX projects to the contralateral hemicord via the dorsal part of the lateral funiculus (100). In 1982, Zheng et al. investigated the DMX efferents to the cerebellum in 70 cats, who had been injected with horseradish peroxidase in all cerebellar cortical lobules and cerebellar nuclei. Injections in the anterior cerebellar lobe resulted in labeled neurons situated in the caudal half of the DMX. The cerebellar posterior lobe received input from the rostral half of the DMX, while the medial DMX projects to the vermal regions of the cerebellum in both the anterior and posterior lobe. In a few cases, labeling of the fastigial and anterior interposed nucleus was found (101). Gastric motoneurons of the DMX possess numerous dendrites that penetrate discrete regions of the adjacent NST. The same study showed that a small number of DMX neurons penetrated the ependyma of the fourth ventricle (80). Moreover, the DMX has been shown to interact with the area postrema via both afferents (80,102,103) and efferents (80,104). The central nucleus of the amygdala in the rat sends a considerable projection to, and receives projections from, the parabrachial nucleus and the dorsal vagal complex (DVC; constituents of the DVC are the NST and the DMX) (105). In other animals, this amygdalofugal pathway has also been described (106,107). Also, the posterior hypothalamic area and mostly the lateral hypothalamic area in rats have been shown to project to the DVC (95,108–110). Manaker and Fogarty conducted a study in 1995 that described raphespinal and reticulospinal neurons projecting to the DVC, while focusing primarily on NST afferents (111). Finally, the rostroventrolateral medulla, a brain structure known to be involved in cardiovascular regulation, has restricted terminal fields in the DVC (96). The aforementioned connections are depicted in Figure 4.
Schematic overview of the central projections of the DMX. Latin numbers indicate cranial nerves or their corresponding nuclei; color of the tract is consistent with the color of the nucleus/area it sprouts from.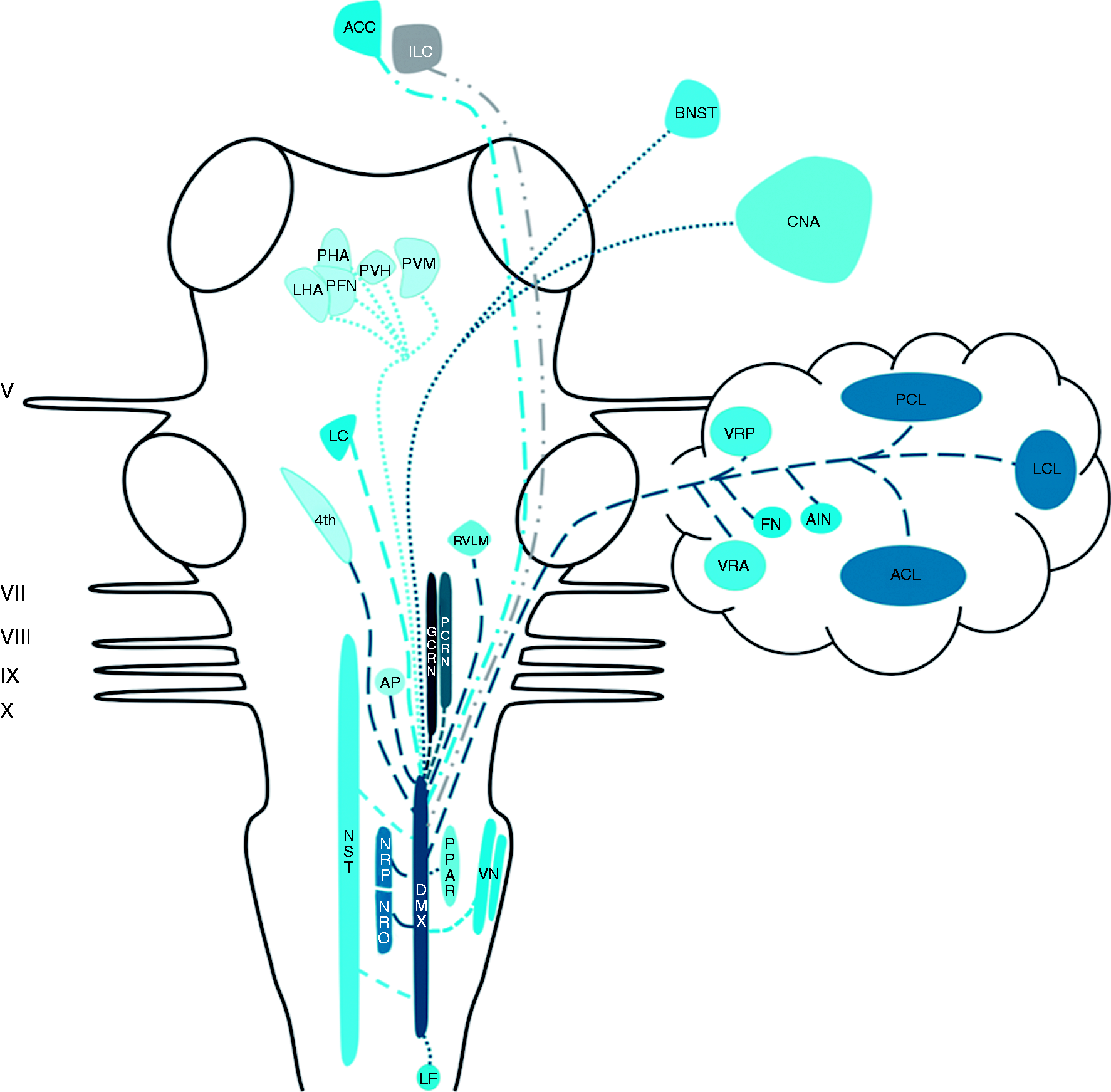
Discussion
The present review synthesized evidence suggesting that nVNS can be beneficial in treating primary headache disorders, migraine and episodic cluster headaches in particular. Two level one studies on episodic and chronic migraine showed effectiveness of nVNS as an acute and prophylactic treatment option (46,48). In episodic cluster headache, on the other hand, only the effectiveness of the acute treatment of nVNS was reported in two level one studies (24,51). In addition, results suggest that nVNS is a well-tolerated and safe therapy. Nevertheless, the exact neural underpinnings of nVNS remain largely elusive, though it has been suggested that modulation of the central connections of the vagus nerve are of importance in alleviating pain in primary headache disorders. Therefore, the second part of this study reviewed the central connections of the vagus nerve. However, no original research papers on this topic were found that included humans. In animals, extensive interconnections between the different vagal nuclei have been described. Also, the TSN and NST, the vagal nuclei that receive sensory and visceral afferents, are the main sprouting areas for fibers that course towards main parts of the neural pain matrix directly or indirectly via other vagal nuclei. Moreover, it has been described that vagal nerve fibers intertwine with fibers from the trigeminal, facial, glossopharyngeal and hypoglossal nerves within the trigeminal spinal tract, forming a trigeminovagal complex.
Clinical relevance of findings
Several studies investigated the possible mechanisms of action of nVNS in both animals and humans (53–55,57,85,112–116). The results of the study of Nonis et al. showed that nVNS in humans was capable of activating vagal afferents as measured by electrophysiological recordings, indicating that efficacy of nVNS can be monitored in future trials (112). An extensive body of literature provides evidence for the anti-inflammatory effect of VNS (113), which may also play a role in the mechanisms by which (n)VNS ameliorates pain (114). Another mechanism of action includes modulation of the trigeminal system by nVNS. In a rat model of migraine-like headaches, nVNS affected the levels of extracellular glutamate in the caudal part of the TSN (115). Another study investigated the firing rate of trigeminocervical neurons in rats treated with nVNS. They suggested that nVNS may inhibit the firing rates of trigeminocervical neurons directly or indirectly, although the exact physiological mechanisms involved remain elusive (116). Based on animal research, direct and indirect connections of the trigeminal and vagal nerves at the level of the brainstem are suggested to be part of the pathways involved in headache pain (53–55,57,85). It is therefore hypothesized that inhibition of the firing pattern of the trigeminal neurons occurs due to the existence of these reciprocal connections between the TSN and the NST, which may serve as the main targets for afferent fiber tracts from the vagus nerve (117,118). This was also in agreement with the study of Frangos et al., which showed that, next to other brain regions, the NTS and STN were respectively activated and deactivated by nVNS (119).
Our review further contributes to elucidating the working mechanism of nVNS in primary headache disorders by synthesizing the evidence of the presence of a trigeminovagal complex in the brainstem of a wide range of animals. However, the existence of an anatomically distinguishable trigeminovagal complex in humans remains uncertain. Future research should focus on imaging human neuroanatomy, especially of the trigeminovagal complex. New, high-resolution imaging techniques could contribute to revealing such neuroanatomical pathways in humans. For example, diffusion magnetic resonance imaging (dMRI) and tractography have become well-known methods to study white matter anatomy (120). To study white matter architecture in even greater detail, an innovative technique called polarized light imaging microscopy (121) might be a valuable asset to study the hypothesized trigeminovagal complex in humans.
Strengths and limitations
This systematic review of the clinical studies on nVNS reveals four level one studies in primary headache disorders which are supplemented by numerous prospective and retrospective reviews. There is heterogeneity in applied methodologies, disease states, duration of headache and outcome measures. These aspects make it more difficult to perform a single meta-analysis on the results. One particular problem concerned the higher than expected sham results with the sham device in both the cluster headache and migraine studies. This suggests the possibility of a placebo effect (46,122,123). Alternatively, we now know that in several of these studies the sham was later determined to be active stimulation, and thus there was no true inactive device or sham stimulation (124). The methods that we have used in this review strengthen our conclusion. Another strength of this study concerns the combination of a systematic review on nVNS in primary headache disorders with an anatomical review. By combining both reviews, we aimed to contribute to the elucidation of the underpinnings involved in this clinical phenomenon. Therefore, based on the reviewed evidence, we put forth two hypotheses. First, we hypothesize the existence of a trigeminovagal complex in humans. Second, we hypothesize the involvement of this complex in nVNS to treat primary headache disorders. Furthermore, the combination of anatomical evidence, and the suggestions that come forth from the clinical trials, can be regarded as a strength of this paper. However, the complete absence of original research on the central connections of the vagus nerve in humans uncovers a remarkable imperfection in knowledge of human neuroanatomy.
Conclusion
Based on the results from several clinical trials reviewed in this paper, moderate effectiveness of nVNS in primary headache disorders is suggested, although the heterogeneity of the included studies precludes the drawing of a sound recommendation. Furthermore, the hypothesized trigeminovagal complex in humans, similar to an anatomical feature in the brainstems of various animal species, could further elucidate the neural underpinnings of nVNS in alleviating pain in primary headache disorders, migraine and cluster headaches in particular.
Article highlights
nVNS is a moderately effective, safe and well-tolerated therapy for migraine and cluster headache. In animals, connections between the trigeminal and vagus systems were found at the level of the brainstem. These connections could contribute to the neural underpinnings of nVNS in primary headache disorders. The existence of a trigeminovagal complex remains elusive in humans.
Declaration of conflicting interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Dr Staats is an employee of both National Spine and Pain Centers and ElectroCore.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Supplemental Material
Supplemental Material1 - Supplemental material for Vagus nerve stimulation for primary headache disorders: An anatomical review to explain a clinical phenomenon
Supplemental material, Supplemental Material1 for Vagus nerve stimulation for primary headache disorders: An anatomical review to explain a clinical phenomenon by Dylan Jozef Hendrik Augustinus Henssen, Berend Derks, Mats van Doorn, Niels Verhoogt, Anne-Marie Van Cappellen van Walsum, Peter Staats and Kris Vissers in Cephalalgia
Supplemental Material
Supplemental Material2 - Supplemental material for Vagus nerve stimulation for primary headache disorders: An anatomical review to explain a clinical phenomenon
Supplemental material, Supplemental Material2 for Vagus nerve stimulation for primary headache disorders: An anatomical review to explain a clinical phenomenon by Dylan Jozef Hendrik Augustinus Henssen, Berend Derks, Mats van Doorn, Niels Verhoogt, Anne-Marie Van Cappellen van Walsum, Peter Staats and Kris Vissers in Cephalalgia
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.