
Abstract
Background
Impaired sensory processing in migraine can reflect diminished habituation, increased activation, or even increased gain or amplification of activity from the primary synapse in the brainstem to higher cortical/subcortical brain regions.
Methods
We scanned 16 episodic migraine (interictal) and 16 healthy controls (cross-sectional study), and evaluated brain response to innocuous air-puff stimulation over the right forehead in the ophthalmic nerve (V1) trigeminal territory. We further evaluated habituation, and cortical/subcortical amplification relative to spinal trigeminal nucleus (Sp5) activation.
Results
Migraine subjects showed greater amplification from Sp5 to the posterior insula and hypothalamus. In addition, while controls showed habituation to repetitive sensory stimulation in all activated cortical regions (e.g. the bilateral posterior insula and secondary somatosensory cortices), for migraine subjects, habituation was not found in the posterior insula. Moreover, in migraine, the habituation slope was correlated with the amplification ratio in the posterior insula and secondary somatosensory cortex, i.e. greater amplification was associated with reduced habituation in these regions.
Conclusions
These findings suggest that in episodic migraine, amplified information processing from spinal trigeminal relay nuclei is linked to an impaired habituation response. This phenomenon was localized in the posterior insula, highlighting the important role of this structure in mechanisms supporting altered sensory processing in episodic migraine.
Introduction
Migraine is a neurovascular disorder characterized by altered neural processing in the central nervous system (1,2). Importantly, hyperalgesia, allodynia, and impaired habituation have been commonly reported in migraine patients, even during the interictal phase (between attacks) (3,4). While most migraine neuroimaging studies assessing altered neural processing (e.g., impaired habituation) have focused on cortical responses and used evoked potentials and electrophysiological methods, functional MRI may be more appropriate to evaluate longer duration stimuli and deeper brain structures, including the brainstem. In fact, increased gain or amplification of activity from the primary synapse in the brainstem to cortical/subcortical brain regions has not been assessed in migraine.
Altered sensory processing can occur on multiple levels of the central nervous system. For instance, amplification of peripherally generated signaling can occur at the level of the first synapse in the spinal cord, higher up in cortical or subcortical regions, or both. No studies, however, have explicitly evaluated amplification of activation focusing on both the brainstem and cortical/subcortical brain regions in migraine, particularly with functional MRI (fMRI).
Habituation is a neurophysiological phenomenon characterized by a reduced response to repeated sensory stimulation (5). Neural habituation is considered protective to the organism, as it shields the brain from excessive information processing and limits energy consumption (6). For instance, habituation is critical for learning, in order to discriminate relevant stimuli and focus on selective properties of external stimuli (5). In migraine, impaired habituation to not just pain but also non-noxious sensory stimuli of various modalities (visual, auditory, olfactory, somatosensory) has been commonly reported, mainly using electrophysiological approaches (7,8). For instance, the late component of nociceptive laser-evoked potentials, which may be generated from the deep insular and anterior cingulate cortices, demonstrates reduced habituation in migraine patients (9–11). FMRI neuroimaging has also assessed habituation in the brain to repeating blocks of sensory stimulation (12,13). Additionally, a recent study found that migraine patients do not report decreasing intensity ratings for repeated painful stimuli, and when such ratings are used to guide the fMRI analysis, pain-evoked activity in the insular and anterior cingulate cortices similarly demonstrates a lack of habituation (14).
Afferent sensory inputs (tactile, thermosensory, and nociceptive signals) from the face are transmitted to the trigeminal sensory complex in the brainstem through the afferent trigeminal nerve, then to the thalamus (lateral and medial nuclei) and primary somatosensory cortex (SI, face area). This pathway is known as the primary somatosensory trigeminal pathway (15-18). The trigeminal sensory complex in the brainstem consists of the main sensory nucleus and spinal trigeminal nucleus (Sp5). The main sensory nucleus (rostral portion of the trigeminal sensory complex) is activated by low-threshold tactile (e.g., touch and pressure, Aβ mediated afference) stimulation. Sp5 is also activated by Aβ-mediated tactile (touch and pressure) afference, mainly in the oralis subdivision, while the caudalis subdivision is known to mainly process Aδ-/C-mediated nociceptive and thermosensory afference (15-18). In addition, while multiple fMRI studies have demonstrated Sp5 activation in response to nociceptive stimulation in patients with trigeminal tractotomy (16), trigeminal neuropathy (19), migraine (20), as well as in healthy adults (21), few studies have evaluated the trigeminal sensory complex response to non-painful stimulation.
In this study, we applied fMRI to investigate two aspects of tactile sensory processing in episodic migraine: impaired habituation and cortical/subcortical amplification of the brainstem response to non-painful ophthalmic nerve somatosensory input. We hypothesized that interictal migraine patients would demonstrate reduced habituation in multiple stimulus-activated brain regions, which would be associated with increased amplification relative to activation at Sp5, the site of the primary brainstem synapse.
Methods
All studies were performed at the Martinos Center for Biomedical Imaging in Boston, Massachusetts. The experimental procedure was approved by the Partners Human Research Committee (approval number: 2005P-000466), and research was performed in accordance with the principles of the Declaration of Helsinki. All participants in the study were fully informed of the procedure and provided written informed consent.
Demographics and clinical characterization
Migraine diagnosis was based on the classification of the International Headache Society (Headache Classification Committee of the International Headache Society, 2004). Inclusion criteria allowed for the enrollment of episodic migraine patients with 2–15 episodes per month. Exclusion criteria included other neurological or major psychiatric disorders. Sixteen migraine patients (MIG, 15 females, 35.8 ± 13.4 years old, mean ± SD) and sixteen sex/age-matched healthy controls (HC, 15 females, 36.0 ± 13.7 years old, p = 0.96) participated in this cross-sectional study (22). In migraine patients, clinical data such as episodes per month (times), presence of aura, migraine duration (years), lateralization of migraine attack (right, left, or variable right/left), and spatial extent (frontal, temporal, occipital, vertex, or a combination) were collected. All MIG patients were scanned interictally (i.e. between episodes). Subjects were asked on the day of the scan when their preceding ictal event took place (PRE_I: the number of days between the previous migraine episode and the scan visit), and were followed up by phone regarding the subsequent migraine episode, after the scan visit (NEXT_I: the number of days between the scan visit and the subsequent migraine episode). We then calculated the “interictal phase”, defined for the day the subject was scanned (see equation 1), to take into account individual patients’ attack-to-attack cycle, which resulted in a metric scaled from 0 (immediately after the last attack) to 100 (immediately preceding the next attack).

Air-puff stimulation
To investigate the stimulus-related brain response in migraine patients and healthy controls, the outlet of MR-compatible air tubing (inner diameter = 12 mm) was positioned over the right supraorbital region of the forehead, approximately 2 cm above the medial aspect of the eyebrow. This location targeted the ophthalmic (V1) spinal trigeminal nerve branch (Figure 1(a)). The tubing was passed through the MR-scanner penetration panel and connected to an air compressor controller (AIRSTIM, San Diego Instruments, Inc., CA, USA). Non-painful air-puff stimulation (80 Psi, 5 Hz) was delivered through the tubing to the subjects using a block fMRI design (14 seconds ON and 20 seconds OFF, 11 repetitions, a total of 370 seconds (Figure 1(b)), see below for more detail). We chose a stimulus frequency, 5 Hz, on the same order but higher than that reported in previous studies demonstrating reduced habituation (e.g. 3 Hz somatosensory stimulus (23)). After the fMRI experiment, the intensity of air-puff sensations during the scan (i.e. psychometric outcome) was rated by subjects on a numerical rating scale (NRS) of 0 to 10 (0: no sensation, 10: pain detection threshold, i.e. on the verge of painful sensation). A two-tailed Student’s t-test (unpaired, SPSS v. 10.0.7, Chicago, IL, USA) was used to evaluate the difference between MIG and HC, significant at p < 0.05.
Innocuous somatosensory (air-puff) stimulation, which was used for ROI selection. (a) Right forehead (V1, ophthalmic nerve territory) was stimulated. (b) In a block design (air pressure = 80 Psi with 5 Hz, 11 repetitions, 14 second stimulation and 20 second rest).
MRI data collection
This fMRI study was performed using a 3.0T scanner (Trio, Siemens Medical, Germany) with a 12-channel head coil. Subjects were asked to lay supine in the scanner with their eyes closed while staying alert and awake. They were also asked to focus on the air-puff stimulation and remain still during structural and functional scan runs, which were assisted by padding the head inside the head coil. Earplugs were provided to attenuate noise during data collection.
Structural MRI data were collected using a T1-weighted MP-RAGE pulse sequence (TR = 2530 ms, TE = 1.64 ms, flip angle = 7°, FOV = 256 × 256 mm2, 176 axial slices, voxel size = 1 × 1 × 1 mm3), and whole-brain functional MRI data were acquired using a T2*-weighted blood oxygenation level dependent (BOLD) pulse sequence with increased matrix size for improved in-plane spatial resolution to increase sensitivity for brainstem nuclei with small cross-sectional area (TR = 2500 ms, TE = 30 ms, flip angle = 90°, FOV = 220 × 220 mm2, matrix = 84 × 84, 43 axial slices, slice thickness = 2.62 mm, gap = 0.5 mm, voxel size = 2.62 × 2.62 × 3.12 mm3).
We corrected for cardiorespiratory-related physiological artifacts in fMRI data, which is critical for brainstem analysis (24). Peripheral physiological data were acquired using the Powerlab system (ML880, ADInstruments Inc., Colorado Springs, CO) at a 400 Hz sampling rate. Electrocardiogram (ECG) data were filtered via an MR-compatible monitor (InVivo Magnitude CV, Invivo Research Inc., Orlando, Florida) designed to minimize radiofrequency (RF) and gradient switching artifacts generated during the MRI scan. Heartbeat annotation was performed to localize R-peaks using custom-made MATLAB (The MathWorks Inc., MA, USA) scripts. Respiration data were collected using an MR-compatible belt system constructed in-house, based on the system devised by Binks et al. (25) similar to our previous studies (12,26,27).
MRI data preprocessing
Data preprocessing was performed using FSL (FMRIB Software Library, http://www.fsl.fmrib.ox.ac.uk/fsl/), AFNI (Analysis of Functional NeuroImages, http://afni.nimh.nih.gov/afni), and FreeSurfer (http://surfer.nmr.mgh.harvard.edu). FMRI data were corrected for cardiorespiratory physiological artifacts using the retrospective image correction algorithm (RETROICOR) (24). Head motion was corrected using FSL-MCFLIRT (28). FMRI data were smoothed in space (Gaussian kernel with FWHM = 5 mm) and time (high pass cutoff = 0.0147 Hz, consistent with twice the duty cycle of our block design) domains to increase SNR and to remove MR signal drift, respectively. Structural MRI data were registered to fMRI data (FREESURFER-BBREGISTER) (29), allowing for co-registration of both to the standard space (MNI152 template) using non-linear warping (FSL-FNIRT).
MRI data analysis
A first-level general linear model (GLM) was used to estimate brain response to air-puff stimulation using a single regressor for the air-puff stimulation block design, convolved with the canonical hemodynamic response function (Double-Gamma). The results of this analysis (i.e., the parameter estimate and its variance) were transformed into standard space (MNI152), and passed up to a combined group analysis using FMRIB’s Local Analysis of Mixed Effects (FLAME 1 + 2, cluster-corrected for multiple comparisons, Z = 2.3, p < 0.05). A combined activation map was obtained for the purpose of identifying regions of interest (ROIs), which were subsequently used for group comparisons. Both groups were combined in order to maximize SNR for brainstem response and to define ROIs with unbiased localization for the subsequent cortical/sub-cortical amplification and habituation analyses (see below), which were the focus of this study. ROIs identified from the combined group activation map included a ponto-medullary cluster consistent with Sp5 (localized based on brainstem atlas (30)), important for the amplification analysis.
For activated brain regions, we then evaluated the cortical/subcortical amplification of Sp5 activation. Our metric was similar to that used by other studies investigating central amplification at higher levels of the central nervous system, e.g. the cortical (i.e. the primary somatosensory cortex)/peripheral (i.e. the median nerve) amplification ratio using sensory nerve action potentials was calculated for Carpal Tunnel Syndrome patients (31). For our analysis, a 3 mm diameter sphere mask was centered on the peak voxel for each ROI, and the mean percent signal change of activated voxels was calculated. For mathematical stability, the amplification ratio was calculated as an inverted ‘Sp5 ROI activity / cortical/subcortical ROI activity’, due to the fact that Sp5 signal was more likely to approach nil in some subjects. This ratio was contrasted between MIG and HC using a two-tailed Student’s t-test (unpaired, SPSS v. 10.0.7, Chicago, IL, USA), significant at p < 0.05. For easier interpretability, visual plots are provided with the more intuitive ‘cortical/subcortical / Sp5’ ratio.
We also explored habituation across the multiple air-puff stimulation blocks. This analysis used an additional single subject level GLM, in which each of the 11 stimulation blocks was modeled as a separate regressor, thereby assessing activation for each individual stimulus block (similar to our previous fMRI habituation study (12)). Parameter estimates and variances for each of the 11 stimulation blocks were then transformed into standard space for the ROI analysis. The habituation slope (first-order linear regression) of percent-change across the 11 stimulation blocks was calculated for the same ROIs identified above. Habituation (a linear decrease of activation over stimuli) was defined by passing two criteria, significant at p < 0.05: (1) a significant linear correlation (i.e. Pearson’s r-coefficient), and (2) a linearly-decreasing slope significantly different from nil. Additionally, for each ROI, slopes were contrasted between MIG and HC using an unpaired, two-tailed Student’s t-test, significant at p < 0.05.
We also investigated whether the amplification ratio was correlated with the habituation slope within specific brain regions, thereby more closely linking these neurophysiological outcomes. Additionally, the clinical relevance of these brain responses was evaluated by calculating the correlation between activation, the amplification ratio (for regions showing amplification) and the habituation slope (for regions showing habituation in HC but not MIG), and the interictal phase (as defined above). Following testing for normal distribution using a Kolmogorov-Smirnov test, a Pearson’s correlation coefficient, r, was calculated, significant at p < 0.05.
Results
Demographics, psychophysics, and clinical measures.

Data are shown as mean ± SD. Interictal phase = ratio between preceding (=0) and subsequent (=100) attacks from experiment visit. n/a = not applicable.
All subjects tolerated the air-puff procedures and stimulation intensity, which was rated as non-painful (below 10 on the 0–10 NRS) by all subjects. There were no group differences in ratings of air-puff stimulation intensity (Table 1).
Brain response to innocuous somatosensory ophthalmic nerve stimulation
No subjects were removed from analysis due to excessive head motion (criteria: > 2 mm TR-to-TR translation), and displacement did not differ between groups (RMS displacement: MIG = 0.11 ± 0.09 mm, mean ±SD, HC = 0.11 ± 0.05 mm, p = 0.91). Bilateral activation in the brainstem included pontomedullary junction locations compatible with Sp5 (consistent with the human brainstem atlas (30)) and, more anteriorly, pontine nuclei (Figure 2(a)). Periaqueductal gray (PAG) activation was found in the midbrain (Figure 2(b)). Cortical activation was noted in the bilateral secondary somatosensory cortex (SII), bilateral posterior insula (pINS), and bilateral supramarginal gyrus (SMG). Subcortical activation was noted in the left hypothalamus (Hyp), right putamen, and right caudate nucleus (Figure 2(b)). No group differences for activation response to air-puff stimulation were found within Sp5 or any of the cortical/subcortical brain regions noted above (Table 2).
Brainstem response to somatosensory stimulation. (a) Activation was found in the bilateral spinal trigeminal nuclei (Sp5), consistent with the Duvernoy brainstem atlas (a, left) (30), and overlaid on the MNI-space template (A, middle), rotated for consistency with the Duvernoy brainstem atlas. The right panel of (a) shows the level of the axial slice in green (Obex + 17 mm). (b) Group map of the cortical/subcortical/midbrain response to innocuous air-puff stimulation from all subjects also showed significant activation in cortical areas (the bilateral posterior insula, pINS; the bilateral secondary somatosensory cortex, SII; and the bilateral supramarginal gyrus, SMG), in subcortical regions (hypothalamus, Hyp; right putamen, Put; and right caudate nucleus, Caud), and in a midbrain region (periaqueductal gray, PAG). Both group maps, (a) and (b), were cluster-corrected for multiple comparisons (Z = 2.3, p < 0.05). Brain response to air-puff stimulation. Data are shown as mean ± SD. HC = Healthy controls; MIG = Migraine patients; L = Left side (contralateral to stimulation); R = Right side (ipsilateral to stimulation). Distance from obex was estimated after rotating the brainstem image into the upright position.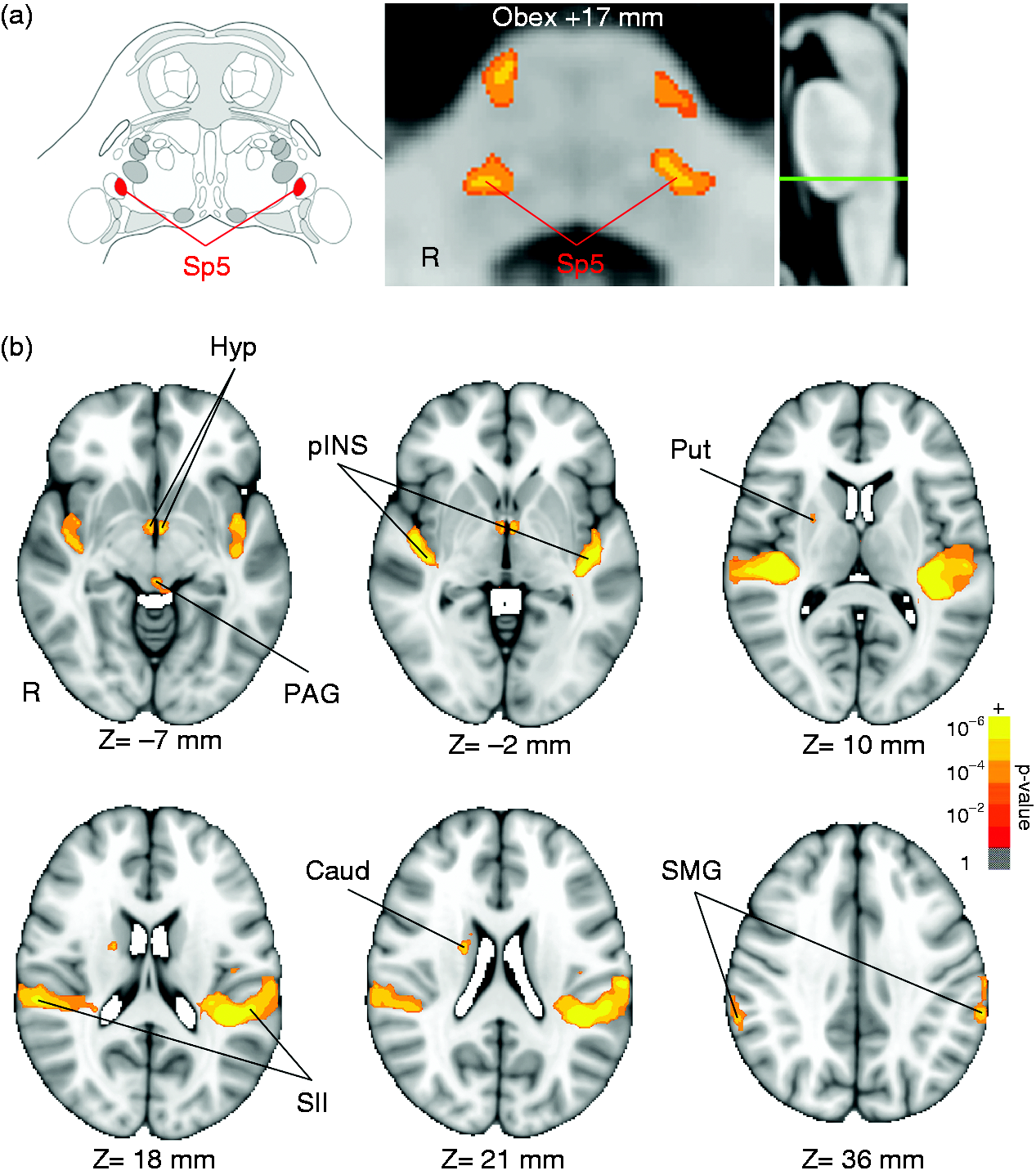

Cortical/subcortical amplification of Sp5 activation
We then calculated the amplification ratios for the cortical/subcortical regions noted above relative to Sp5 activation, and contrasted MIG versus HC groups. A greater amplification ratio was found for MIG compared to HC in two regions: the contralateral posterior insula cortex and hypothalamus (Figure 3, Table 3). Differences between groups in amplification were not seen for other ROIs (Table 3).
Cortical/subcortical amplification of Sp5 activation. Migraine patients showed a greater cortical (i.e. posterior insula, pINS; a) and subcortical (i.e. hypothalamus, Hyp; b) amplification ratio, relative to Sp5 activation. n.b. *p < 0.05, error bars denoted the standard error of the mean. Amplification of Sp5 activation and habituation to ophthalmic nerve somatosensory stimulation. Data are shown as mean ± SEM. Habituation (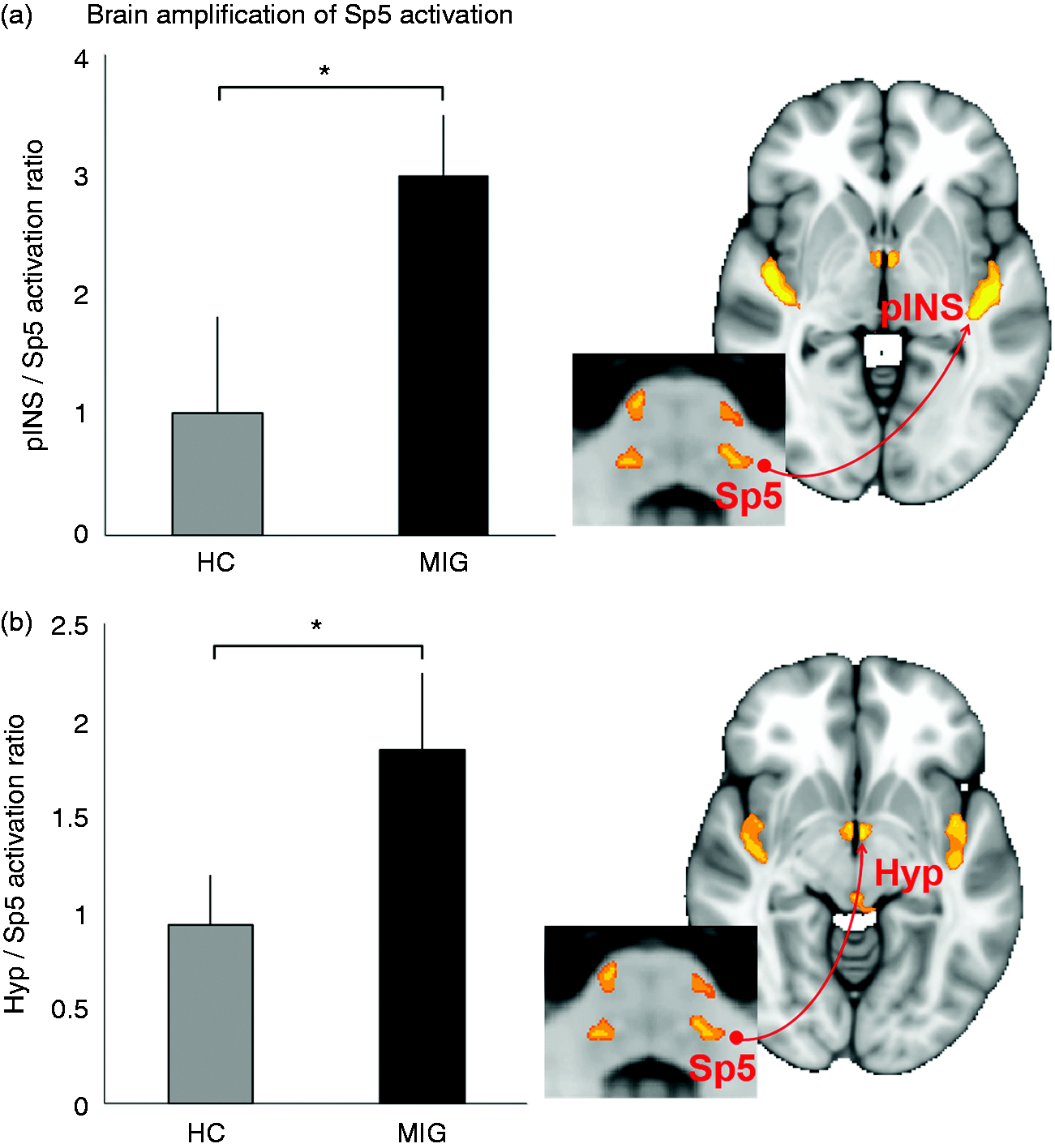

Habituation to repeated blocks of ophthalmic nerve somatosensory stimulation
Neural response habituation was defined by explicit criteria for the fMRI signal (see Methods). For HC, habituation was noted for all activated cortical regions (bilateral SII, bilateral pINS, bilateral SMG), and putamen. For MIG, habituation was noted for bilateral SII and SMG. Directly contrasting MIG and HC, we found a significantly different slope magnitude for the right posterior insula (Figure 4, Table 3). Specifically, for HC, the insula response linearly decreased over time (consistent with habituation criteria), whereas for MIG, the habituation criteria were not met.
Habituation to repeated stimulus blocks for regions activated by innocuous somatosensory stimulation. For healthy controls (HC), habituation was noted for multiple activated cortical/subcortical regions, including the posterior insula (pINS). In contrast, migraine (MIG) subjects demonstrated a significantly (p < 0.05) reduced linear habituation slope in the posterior insula. n.b. **p < 0.01, significant linear fit across blocks for healthy controls.
Associations between different brain activity metrics and links to clinical measures
For MIG, we found a correlation between the habituation slope and amplification ratio in the right pINS (r = 0.54, p = 0.03, Figure 5(a)) and right SII (r = 0.56, p = 0.02, with outlier removed: r = 0.58, p = 0.02, Figure 5(b)), i.e. since a more negative habituation slope reflects greater habituation, greater amplification was associated with reduced habituation in these regions.
Association between the cortical amplification ratio and the habituation slope. Greater amplification was correlated with impaired habituation (a less negative habituation slope) in (a) the posterior insula and (b) the secondary somatosensory cortex.
We also found a positive correlation between the interictal phase and fMRI signal response in the right, ipsilateral Sp5 for MIG (r = 0.58, p = 0.02, Figure 6). Thus, MIG subjects relatively closer to their next attack demonstrated greater Sp5 activation. Kolmogorov-Smirnov testing confirmed that all variables for significant correlation tests were normally distributed (Sp5: p = 0.32, i.e. data consistent with normal distribution; interictal phase: p = 0.45; amplification ratio, right pINS: p = 0.35, right SII: p = 0.20; habituation slope, right pINS: p = 0.89, right SII: p = 0.77).
Association between the interictal phase and Sp5 activation. Signal increase in Sp5 was positively correlated with the interictal phase (a relative ratio between preceding and subsequent attacks) at the time of the scan – i.e. Sp5 activation was greater as migraine subjects approached their next migraine attack.
Discussion
Our study found that compared to HC, MIG subjects showed enhanced amplification of Sp5 activation and reduced habituation in the posterior insula. Amplification and habituation were correlated, suggesting a shared pathophysiology and highlighting the important role of the posterior insula for mechanisms supporting altered sensory processing in episodic migraine.
Amplification or increased gain for incoming afference can occur at the level of the primary synapse in the spinal cord (or the brainstem for orofacial inputs), higher up in the cortical or subcortical regions, or both. Thus, for migraine, while amplification can exist at the peripheral trigeminal nerve receptor, trigeminal ganglion, or Sp5 in the pontomedullary junction, we specifically investigated amplification from activation occurring at Sp5 up to higher order regions in the brain. Compared to HC, MIG demonstrated amplification in two regions: pINS (significant in left/contralateral and trending in right pINS) and the hypothalamus. The posterior insula cortex, which receives Sp5 afference via the thalamus, is an important region for pain and interoceptive processing (32,33), and has been linked with migraine pathophysiology (34). However, this region also responds to innocuous somatosensory stimulation, as evident in our study and many others, and invasive intracortical stimulation of pINS commonly produces generalized somatosensory (e.g. tingling, vibrating, numbness, warmth) and, when more dorsal, even painful sensations (35). Future studies should evaluate if greater amplification of innocuous somatosensory input to pINS in both ictal and interictal states of episodic migraine are linked to allodynia in these subjects. However, we should note that MIG did not rate the air-puff stimulation as painful, supporting the contention that interictal episodic migraine patients may not show allodynia (36).
The hypothalamus also demonstrated amplification, relative to Sp5 activity, in MIG. The importance of the hypothalamus in migraine pathophysiology has been previously reviewed (37), and an altered response in this area is consistent with commonly reported, non-pain symptoms of migraine, including disturbances in sleep, arousal, and homeostatic/autonomic functioning. Specifically, this region has been hypothesized to play an important role as a trigger of migraine attacks, particularly in patients who can sense oncoming attacks from altered arousal and fatigue (38). Moreover, increased excitation (or decreased inhibition) of the hypothalamus (39,40) has been reported to trigger migraine attacks, and significant hypothalamic activation has been observed during migraine episodes and maintained in the postictal state (41). Thus, amplification of sensory stimuli in the hypothalamus may produce alterations of autonomic and other homeostatic functions associated with oncoming attacks.
Reduced habituation has been noted as one of the most consistently reported interictal neurophysiological phenomena in episodic migraine (8). Habituation is defined as a diminished response to repeated stimulation and is noted in both behavioral and neurophysiological domains (5). It is thought to be protective of excessive information processing and energy consumption (6). Our study demonstrated clear cortical (e.g. SII, pINS, SMG) and even subcortical (putamen) habituation to innocuous somatosensory stimuli in HC. In contrast, for MIG, habituation was noted only in bilateral SII, which is known to process physical aspects of somatosensory stimuli (42,43). Additionally, a direct contrast found a significantly different (less negative) habituation slope in pINS, highlighting the reduced habituation of activation in this brain region. Stankewitz et al. also noted a lack of fMRI activity habituation for episodic migraine in response to nasal trigemino-nociceptive (ammonia) stimuli, which was found in the anterior insula and anterior mid-cingulate cortices (14). However, in the same study, migraine subjects demonstrated marked habituation to an innocuous olfactory stimulus. As our study found reduced habituation for an innocuous somatosensory trigeminal stimulus, this effect may be modality-dependent. Furthermore, in our study the habituation slope was correlated with the amplification ratio in pINS for MIG, i.e. greater amplification was directly associated with reduced habituation. These results highlight the altered stimulus-response physiology for pINS during the interictal phase of migraine.
Our study found significant activation in response to trigeminal nerve somatosensory stimulation in bilateral Sp5 (oralis subnucleus). Interestingly, Stankweitz et al. (20) demonstrated a positive correlation between Sp5 activity and time to next ictal event, suggesting that Sp5 response to nociceptive stimulation may be an important predictor of an upcoming migraine attack. In our study, we similarly observed a positive correlation between Sp5 activation and the interictal phase, though not specifically with time to subsequent attack (r = −0.33, p = 0.21). While the two parameters differ in that the interictal phase is a relative measure, and related to individual patients’ attack-to-attack cycles, the relationship with Sp5 activity was similar, such that activation is higher when the next attack is imminent. Moreover, our results suggest that the association between Sp5 activation and subsequent attack onset is not only for nociceptive processing, but also extends to innocuous somatosensory processing in Sp5. Also, bilateral Sp5 activation may have been due to the spatially broad distribution of our air-puff stimuli and the close proximity of the air-puff target region to the face midline, perhaps leading to stimulation of cutaneous receptors on the opposite side of the facial midline.
In addition to Sp5, we also observed significant signal increase in more anterior pontine nuclei, and higher cortical and subcortical regions including basal ganglia areas such as the caudate and putamen. The basal ganglia and cortical regions such pINS, SII, and SMG are known to commonly activate in response to painful and non-painful somatosensory stimulation (43–46), consistent with our innocuous air-puff stimulus. Basal ganglia areas, including the caudate and putamen, have been previously noted to show reduced activation and gray matter volume in migraine patients with more frequent attacks (46), though in our study, activation in these regions was not associated with the interictal phase.
Several limitations in this study need to be mentioned. Firstly, our time-variant analysis assessed linear habituation. While other non-linear response patterns may indeed have occurred, it is important to note that a linear contrast is still sensitive to non-linear time-variant responses. Moreover, a linear contrast has the advantage of easier interpretability. Further, while clinical measures did not differ between aura and non-aura MIG subgroups, our sample size did not allow for direct comparisons of fMRI measures between these subgroups. As another limitation, non-noxious tactile stimuli typically activate Sp5-oralis and the main sensory nucleus of the trigeminal sensory complex. In our study, the lack of activation in the main sensory nucleus may have been due to the small cross-sectional area of this nucleus, and future studies using ultrahigh field (e.g. 7T) fMRI, with better SNR and spatial resolution, should be performed.
In conclusion, our study identified amplification of the cortical and subcortical response, relative to brainstem Sp5 activity, for somatosensory ophthalmic nerve stimulation. Furthermore, we linked this amplification to reduced habituation in several cortical brain regions. Specifically, amplification was found in the hypothalamus, and the posterior insula demonstrated correlated amplification and reduced habituation – i.e. greater amplification was associated with reduced habituation. Our study thus highlights the important role of the posterior insula in mechanisms supporting altered sensory processing in episodic migraine.
Article highlights
Migraine subjects showed greater amplification of trigeminal tactile information transferred from the brainstem to the posterior insula cortex. Migraine subjects also demonstrated significantly reduced habituation, which was correlated to greater amplification, in the posterior insula, a key region for sensory processing in episodic migraine. Neuronal amplification is evident within the central nervous system in migraine.
Footnotes
Acknowledgements
We would like to thank Helen S. Xu and Yazhuo Kong for their helpful comments on data analysis and Hanhee Jung for assistance in recruitment and scanning.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Institutes of Health, National Center for Complementary and Integrative Health [R01-AT007550, P01-AT006663]; National Institute of Mental Health [R21-MH103468]; National Institute of Arthritis and Musculoskeletal and Skin Diseases [R01-AR064367]; National Institute of Neurological Disorders and Stroke [R21-NS087472, R21-NS082926]; the National Center for Research Resources [P41RR14075]; and the Colombian Department of Science, Technology and Innovation [COLCIENCIAS, Grant No. 656664239871].