
Abstract
Background
Migraine is a common neurological condition that often involves differences in visual processing. These sensory processing differences provide important information about the underlying causes of the condition, and for the development of treatments.
Review of psychophysical literature
Psychophysical experiments have shown consistent impairments in contrast sensitivity, orientation acuity, and the perception of global form and motion. They have also established that the addition of task-irrelevant visual noise has a greater effect, and that surround suppression, masking and adaptation are all stronger in migraine.
Theoretical signal processing model
We propose utilising an established model of visual processing, based on signal processing theory, to account for the behavioural differences seen in migraine. This has the advantage of precision and clarity, and generating clear, falsifiable predictions.
Conclusion
Increased effects of noise and differences in excitation and inhibition can account for the differences in migraine visual perception. Consolidating existing research and creating a unified, defined theoretical account is needed to better understand the disorder.
Introduction
Migraine is a debilitating and common condition that, by definition, is associated with differences in visual perception. The International Headache Society criteria for classifying migraine list photophobia, an increased sensitivity to light, as one of the key signs of a migraine attack, alongside a severe headache lasting four to 72 hours and nausea/vomiting (1). Some people with migraine report visual triggers of attacks (2–4), and are particularly sensitive to light stimulation during an attack (5). In some people, a migraine attack is preceded by a set of sensory, motor or speech disturbances known as the migraine aura. Although these disturbances can be in any modality, for example auditory or olfactory, the aura is often visual, which is experienced in 57% of cases (1,6). The visual aura consists of a fortification pattern surrounding a central scotoma. Illustrations of these patterns can be seen in (Sacks (7), Schott (8), Lashley (9)). Other phenomena include patterns such as tunnels and spirals, flashes of light (phosphenes), changes in apparent scale (micropsia and macropsia), and a disjointedness of the visual image, such that objects appear as they do in Cubist paintings (7).
Photosensitivity is aversion to light, which is a feature of migraine. Photosensitivity is increased in migraine (e.g. Hay et al. (10)). Eighty per cent of people with migraine indicated that pain could be induced by light stimulation, and pain thresholds were lower than those for a control group (11). Long wavelength, reddish light (398–503 nm) caused the most discomfort for people with migraine (12). As the trigeminal ganglion responds to light, it could account for some of the photosensitivity in migraine (13). Applying an ice-block to the centre of the forehead, so as to induce trigeminal pain, reduced visual discomfort thresholds for people with migraine (14–16). A thalamic pathway for the exacerbation of migraine headache by light has also been proposed (17). There is evidence of a possible mechanism for this, as the same positron emission tomography (PET) activation from light and pain in controls has been found as from light alone in migraine (18).
Spatially and temporally periodic stimuli are particularly uncomfortable and can induce migraine attacks (2). Stripes caused discomfort for 82% of people with migraine, compared with only 18% of a control sample (19). The tendency of a striped pattern to cause discomfort depends on its spatial frequency, or the width of the stripes (Figure 1). Striped patterns of around three cycles/degree cause greater discomfort than either higher or lower frequencies (20). People with migraine reported more illusions, and experienced greater pattern glare, when viewing 1.2 cycles/degrees gratings, which was accompanied by greater functional magnetic resonance imaging (fMRI) blood oxygen level-dependent (BOLD) response (21). The temporal properties of a stimulus, as well as its spatial properties, affect discomfort judgements. Haigh and colleagues (22) found that the contrast threshold at which two cycles/degree gratings started to become uncomfortable to view was lower in people with migraine than in controls. Drifting and vibrating stimuli were found to be more aversive than static stimuli. People with migraine also report greater discomfort scores for a large, uniform field flickering at 10 Hz (23,24).
(a)–(c) Gratings with progressively increasing spatial frequency. (d) A Gabor patch stimulus, which is a sine grating multiplied by a Gaussian envelope.
There is a substantial body of research into sensory processing in migraine, using clinical, neurophysiological and electrophysiological approaches. This research has established clear differences in visual perception in migraine, accompanied by differences in neural responses to visual stimuli. This has led to the proposal of several theoretical accounts of migraine that are linked to abnormalities in sensory processing. In turn, these accounts have suggested that the visual cortex in migraine is either hyper-responsive or hyperexcited, does not habituate to stimuli, and shows a reduction in the degree of inhibitory interactions between neurons. The distinctions between these accounts can be very fine, and can turn on the precise definition of key terminology, which varies between groups of researchers. The current article provides a review of the literature on visual processing in migraine, and a novel theoretical account that brings together a number of strands of research, and builds on previous explanations. We conclude that, when viewed as a whole, visual processing differences in migraine can be characterised as showing greater effects of internal noise. This term ‘noise’ is used here to refer broadly to any activity in the visual system that can potentially hinder performance on a given perceptual task, and covers a wide range of sources and types of such activity. As such, we introduce a mathematically defined model, based on signal detection theory, in order to help describe and explain the nature of differences in visual perception in migraine. In this review, we concentrate on studies that have researched differences on psychophysical tasks. These differences are accompanied by differences in neural activity in migraine. We do not discuss these in detail, as they have recently been reviewed (25).
There are clear changes in sensory processing across the migraine cycle, seen in both behavioural performance and electrophysiological recordings (e.g. Khalil (26)). Many studies make efforts to avoid testing near the headache event (e.g. McKendrick and Sampson (27)), some aim to investigate these cycle effects directly (e.g. Coppola et al. (28)). Results are not always unidirectional – some electrophysiological studies find more pronounced differences in migraine compared to control in the days before an attack (29–31), whereas other studies showed visual-evoked potential (VEP) habituation normalises just before, and during (32), and only during, a migraine attack (33). Other researchers only found differences in the electroencephalogram (EEG) response 10 days after the migraine attack (34). To further complicate matters, some researchers find cycle effects only for behavioural performance measures, not EEG responses (35). All the psychophysical studies reviewed here have excluded individuals close to a migraine attack. However, it should be borne in mind that, for one-off experiments, migraine cycle effects are likely to add variability to the results.
Visual processing differences in migraine
The visual discomfort and visual aura that can occur in migraine, together with the fact that visual stimulation can trigger a migraine attack, and the association with increased neural activity, all suggest differences in visual processing in migraine. Such differences have been found across many domains, for example, the perception of colour (36–39), binocular vision (40) and the higher-level visual processing involved in the perception of facial expressions (41). Here, we concentrate on studies of low-level visual processing in migraine.
Contrast sensitivity
A summary of the literature on contrast sensitivity in migraine.

F: female; MA: migraine with aura, MO: migraine without aura; C: control; MOC: method of constant stimuli; 2IFC: two-interval-forced-choice, photopic: full light levels; scotopic: reduced light levels; cpd: cycles per degree.
To summarise, some studies have found reduced contrast sensitivity in migraine, but this is not unanimous. The variability in results across studies is likely to reflect the heterogeneity of the condition, the migraine cycle effects discussed, the very different stimuli and methods used in different studies, and the small sample sizes often employed. Differences in stimuli and psychophysical methods, in particular, make it difficult to compare studies fairly. Overall, there does seem to be some evidence for poorer contrast sensitivity in migraine.
Inhibitory interactions
The activity of visual neurons is determined by both direct stimulation by external sources, and by inputs from other neurons, which can be both inhibitory and excitatory. Some of these interactions occur at the scale of the receptive fields of individual neurons. For example, the orientation selectivity of individual neurons depends on both feed-forward input from the lateral geniculate nucleus (LGN), and inhibitory input from other cortical neurons (53–55). Suppression effects from surrounding units also operate on larger scales, for example, those that are responsible for contrast normalisation (56) and surround suppression (57). Evidence for this has been found in V1 single-cell recordings in macaques (58). It has also been suggested that V1 cells respond to entire feature combinations (e.g. a red, vertical line). This will allow for suppression not only across single characteristics of the target, but across combined features. This iso-feature suppression has been proposed as an important mechanism in visual search tasks (59). In addition, inhibitory and excitatory interactions form the basis of the mechanisms that group individual features into extended contours, and coherent global form (60). If lateral inhibition processes are reduced in migraine, there will be differences on performance in these tasks compared to control populations.
Orientation acuity
Orientation discrimination is thought to rely on lateral inhibition between neighbouring neural units (53–55). Therefore, differences in inhibitory processes could lead to different performance in migraine. Orientation acuity was measured with a derivative of Gaussian stimulus, and a sinusoidal grating, both foveally (nine cycles/degree) and peripherally (3.6 cycles/degree). No significant differences in orientation thresholds were found (61). Other researchers found poorer sensitivity to orientation differences for foveally presented Gabor patches with a spatial frequency of 0.5 cycles/degree, but not for peripherally presented stimuli, or those with a spatial frequency of four cycles/degree (50). Researchers measured orientation sensitivity using a four cycles/degree Gabor patch, and a virtual line, defined by the presentation of two circles presented either side of fixation (43). Poorer orientation discrimination was found for both stimuli when they were at oblique orientations, but not when they were horizontal or vertical (43). Poorer performance with the virtual line stimuli is interesting because is indicates that the deficit in orientation acuity occurs for stimuli that are processed in extrastriate cortex, and is not therefore restricted to V1; in other words, the deficits are found in higher visual areas also.
Surround suppression
Battista and colleagues (62) measured the size of the Chubb illusion (63), in which the apparent contrast of a central region is reduced in the presence of a high-contrast surround (Figure 2). They found a greater effect in migraine for moving stimuli (two cycles/degree grating, drifting at two cycles/second), but not for static stimuli (four cycles/degree grating). These results show a greater surround suppression effect in migraine, but only for moving stimuli.
An illustration of the Chubb illusion, as used by Battista et al. in 2011 (62). The small circular region has the same contrast in both figures, but appears to have a higher contrast when (a) it is surrounded by a uniform background than (b) when it is surrounded by a high-contrast background striped background.
Visual search
In simple visual search tasks, observers are asked to detect a target that differs in some way from a background of distracters, and it has been proposed that this relies on interactions at the feature level, called iso-feature-suppression (59). The difference between the target and the distracting elements determines the ease with which the task can be completed. For example, the elements might be lines, with the target differing from the distractors in orientation. People with migraine are able to complete search tasks more quickly than others, while maintaining the same level of accuracy (64). However, other studies that have assessed visual search performance in migraine have failed to find any such improvements (38,65–67). There is thus no evidence that performance on visual search tasks is different in migraine.
Summary of inhibitory processes
Behavioural effects that rely on lateral inhibition processes (e.g. orientation tuning, suppression effects, and visual search) are on the whole unimpaired in migraine.
Masking effects in migraine
Adding visual noise to a stimulus can cause masking, reducing the observer’s ability to accurately detect or discriminate a target. A noise mask can take many different forms, and can affect performance by either spatially or temporally disrupting processing. Masking is often accounted for by considering the fact that the mechanisms responding to the target stimulus will also respond to the masking noise (68). However, even when the target and mask activate separate mechanisms, masking can still result from interactions between them, as occurs in surround suppression (69). Baker and Meese noted that some studies of masking might in fact be demonstrating the effects of surround suppression (70). This has important consequences in the interpretation of masking effects in migraine.
Direct visual masking
Direct masking occurs when a mask is presented at the same time, and in the same spatial location, as the target stimulus. Greater masking effects have been found in migraine compared to control groups when mask (grating) and target (Gabor) were presented simultaneously (44). Poorer performance has been found in migraine to detect a small circular target against a random noise background in migraine (51). These results were replicated (52), suggesting that task-irrelevant backgrounds can create greater interference with the successful detection of target stimuli in migraine groups.
Metacontrast masking
The accuracy with which a target can be detected can be affected by the presence of other, task-irrelevant stimuli, even if the two do not overlap in time or space. The effects of such metacontrast masking have been assessed in migraine. Increased masking has been shown for MA compared to MO and control groups, both when a mask was presented at the same time as the target, or with a delay of up to 156 ms (71). Since this study did not include a control in which the target was presented without a mask, it is not known whether the improved performance was due to a decrease in masking, or simply to greater accuracy in the target-detection task. This experiment was replicated (72), this time including forward masking conditions, in which the mask preceded the target, and a condition in which forward (mask presented before the stimulus) and backward (mask presented after the stimulus) masking were present. As found previously (71), better performance was associated with migraine, both with and without aura (72). This difference reflected an overall higher level of performance in the migraine groups, rather than a reduction in metacontrast masking; when baseline performance in the absence of a mask was taken into account, no difference in the degree of masking was observed. Metacontrast masking was masking using a vertical bar target, with both forward and backward masking (73). However, in contrast with a previous work (71), no difference in performance associated with migraine was found (73). It is notable that across these studies, although no evidence for increased metacontrast masking was found, baseline performance was better in the migraine group.
Greater backward masking was found in migraine when participants were asked to detect small deviations of a contour from circularity (69). The main difference between this study and the others discussed here is that it assessed the effects on the just-noticeable differences in the target shape, rather than simply its presence or absence. Thus, the effects of masking are very much dependent on the type of mask and the difficulty of the task itself, and theoretically motivated experiments are needed to more systematically investigate the possibility of increased masking in migraine, taking account of baseline performance. On the whole, there seems to be some indication that migraine groups might be more affected by some effects of masking.
Global coherence
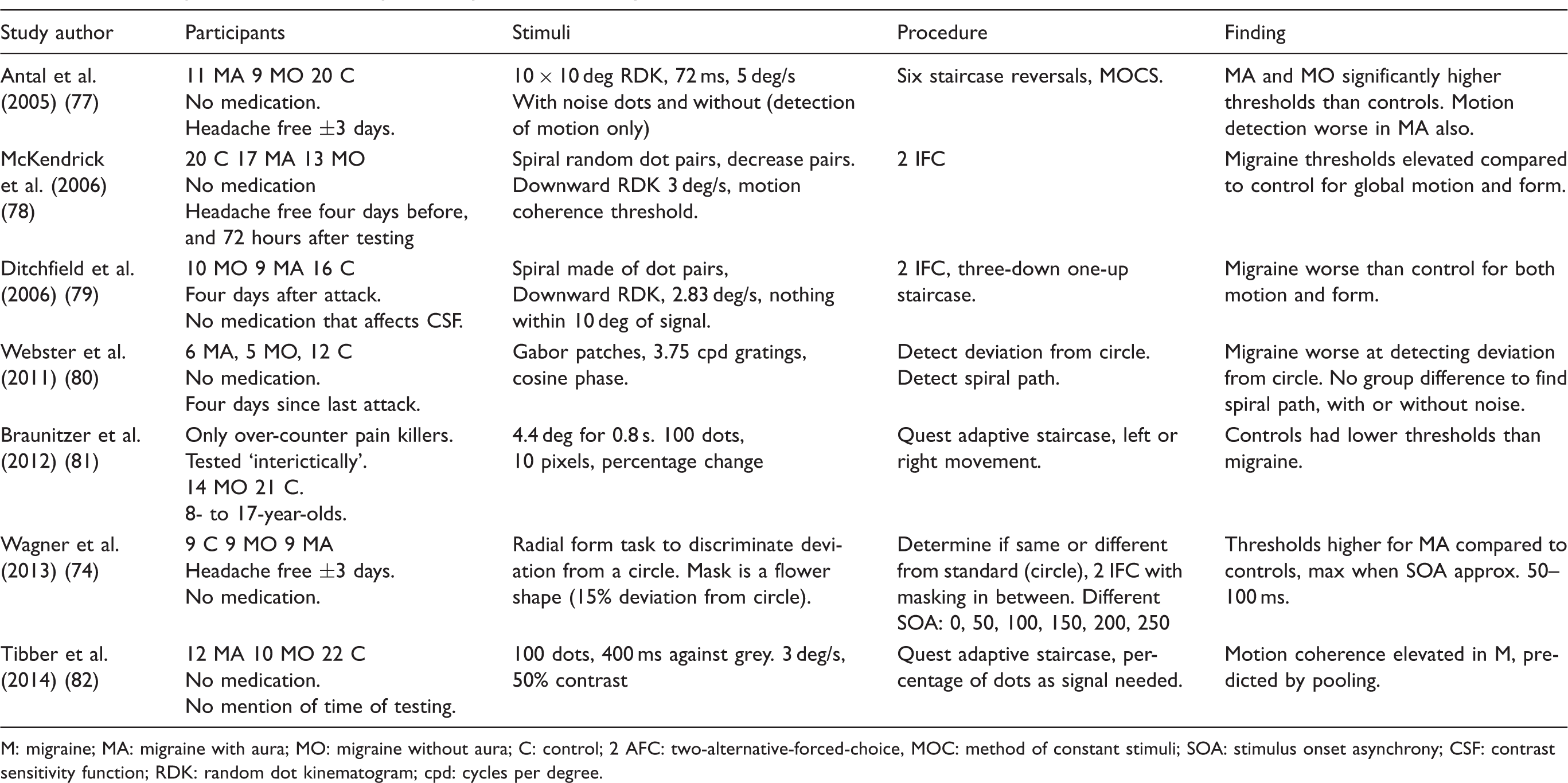
Visual perception requires not only the effective detection of localised information, but also the integration of this information across space and time. The integration of information across space allows for the detection of coherent form, and is achieved through a combination of excitatory and inhibitory interactions between neurons with nearby receptive fields, forming ‘association fields’ (75), as illustrated in Figure 3. Analogously, the integration of local information across time is needed for the successful detection of global motion (76). Successful performance on global motion and form tasks thus relies on successful global pooling processes. A summary of the literature on global form and motion tasks can be seen in Table 2.
An illustration of an association field of the type proposed by Field et al. (1993) (75), adapted from Dakin and Baruch (2009) (83). Each circle represents the receptive field of an orientation-tuned neuron. Excitatory connections (solid lines) exist between neurons whose position and orientation tuning are consistent with smooth, continuous contours. Inhibitory connections (dashed lines) exist between neurons whose orientation tuning is inconsistent with smooth, continuous contours. A summary of the literature on global integration tasks in migraine. M: migraine; MA: migraine with aura; MO: migraine without aura; C: control; 2 AFC: two-alternative-forced-choice, MOC: method of constant stimuli; SOA: stimulus onset asynchrony; CSF: contrast sensitivity function; RDK: random dot kinematogram; cpd: cycles per degree.
Global form perception
The Glass pattern is a stimulus that has been developed to understand how local information is integrated to perceive shape (84). Figure 4(a) shows an example consisting of randomly positioned pairs of dots. The orientation of the dot pair is orthogonal to its direction from the centre of the stimulus, so that the overall pattern appears to consist of a number of concentric circles. Similar Glass-line patterns can be created using lines rather than pairs of dots (Figure 4(b)). The global coherence of Glass patterns can be reduced by randomising the orientation of a proportion of the dot pairs or lines. Some studies have found poorer performance in migraine compared to control groups to detect global shape (78,79). This impairment is also found in children with migraine (10–18 years) but not in younger children (6–9 years) (80).
Stimuli used in the perception of global form and motion. (a) A Glass pattern, in which a random dot pattern, and a rotated version of the pattern, are combined, given the appearance of a circular structure. (b) A Glass line pattern in which the structure is determined by oriented lines, rather than pairs of dots. (c) The global shape task used by Webster et al. (2012) (52), in which observers were asked to detect deviations from a circular contour, as shown here. (d)–(f) illustrate the construction of a typical global motion stimulus. (e) Signal dots that are all moving in the same direction, in this case to the right. (f) Noise dots, moving in randomly chosen direction. (f) A typical stimulus in which signal and noise dots are combined and observers are asked to judge whether the overall direction is rightwards or leftwards.
However, this deficit appears to be task-dependent: (80) used a different task, based on discrimination between two Glass patterns, rather than detection of a single Glass pattern, and found no difference was found for either condition between people with migraine and a control group. Similarly, other researchers (70) found no difference in the magnitude of deviation from the circle that could be detected between people with migraine and a control group. Using a similar task, but with a continuous contour, higher thresholds for people with MA than for a control group and those with MO were reported (74). No difference was found between the latter two groups.
It appears that deficits are found in migraine when the stimulus consists of discrete signal and noise elements (78,79,85) but not when all elements contribute to the signal, but are corrupted by noise (52,80). This is theoretically interesting because it might indicate that there is not an issue with integration of elements per se, but that the increasing external noise from less coherent stimuli has a greater impact on migraine groups compared to controls for detection tasks. This pattern of results is reflected in the perception of global motion (see below) and suggests an inability to exclude visual noise in migraine (82).
Motion coherence
In global motion coherence tasks, observers are required to detect the direction of a signal motion (e.g. movement to the right) in a set of randomly positioned dots, in the presence of incoherent motion noise (Figure 4(d)–(f)). This task requires the integration of motion information across space and time, and is linked to extra-striate processing in the cortical area V5/MT+ (86,87). Deficits in global motion processing in migraine have been shown in a number of studies (50,77–80,88–90), and these may be related to anatomical differences, such as cortical thickening of V3A and MT+, in the motion processing pathway (91). Other researchers also found worse performance on a motion coherence task in a group of children with migraine (81). Differences in performance were particularly pronounced in the youngest age group (8 years old), and improved with age for the group with migraine, but not for the control group. The authors concluded that the development of visual motion processing is delayed in migraine, but normalises with time. However, since deficits in global motion tasks have been found in adults in other studies, these differences cannot be purely attributed to delayed development.
Tibber and colleagues measured sensitivity to global motion in two ways: by varying the proportion of signal to noise dots in a stimulus, as in other studies, and by disrupting the motion information conveyed by all dots (82). In the latter case, dots are not divided into distinct signal and noise groups. The direction of each dot is instead drawn from a Gaussian distribution. The observer’s task is then to determine the mean direction of motion, and the difficulty of this task is varied by changing the standard deviation of the distribution. They found that performance was worse in migraine in the former case, but not the latter. They attributed this to a failure to exclude the motion information conveyed by the noise dots, rather than an increase in internal neural noise, which would have affected performance for both types of stimulus. It was found that motion coherence thresholds were higher in people with migraine, and that these covaried with differences in contrast sensitivity (90). This is important as it could be the case that low-level visual deficits could account for the higher-level findings. Alternatively, the two deficits might have a common mechanism. In any case, the relationship between low- and higher-level deficits should be considered in research. An interesting finding of this study was that performance was slightly better in people with migraine at low levels of added dynamic noise. Dynamic noise involved a task-irrelevant stimulus in the background, in this case stationary, flickering noise dots in the random dot array. This difference was not found in the control group. This is an interesting finding, as it shows that adding task-irrelevant noise improved performance in migraine, when it is generally considered that adding a particular type of task-irrelevant stimuli would impair performance, for example in the case of masking.
Adaptation and aftereffects
Viewing a stimulus for a period of time can lead to adaptation and habituation, resulting in aftereffects. The processes underlying aftereffects can include neural fatigue, and possibly gain control, and have been reviewed (92).
Tilt aftereffects
After viewing a tilted stimulus for a period of time, other similar stimuli appear to be tilted in the opposite direction. These tilt aftereffects are generally increased in migraine, both for stimuli presented one after the other (93) and also for stimuli presented simultaneously, alongside each other (94). This is at first sight curious because, as previously discussed, sensitivity to differences in orientation is lower in migraine. Therefore it is plausible that the greater aftereffect is due to a less selective neural response in the first place, as a result of less inhibition between neurons. This might then lead to a greater initial response and therefore a larger aftereffect.
Motion aftereffects (MAEs)
After viewing a moving image for a prolonged period of time, there is a tendency for subsequently viewed stimuli to appear as though they are moving in the opposite direction. Shepherd (89,93) showed that the duration of this MAE is longer in migraine.
Battista and colleagues (95) assessed the MAEs and also used it to measure the effects of surround suppression, and showed reduced MAEs only for large, high-contrast stimuli in migraine. This is consistent with increased surround suppression: The strength of the aftereffect reduces for large stimuli, due to surround suppression of the response of motion detectors as the stimulus extends beyond their receptive field (96).
Palinopsia
Another form of aftereffect that is more common in migraine than controls is palinopsia. This refers to the persistence or recurrence of images once the stimulus has been removed (97). It can take many forms, which typically involve the persistent perception of objects, actions, parts of objects or textures, and the incorporation of these into the currently viewed scene. Palinopsia is more common in migraine than those without. It has been reported that palinopsia occurred in around 10% of people with migraine, and no incidence of this condition was found in their sample of 226 healthy controls (98). Palinopsia was more common in patients with MA, than in those without aura. Patients with palinopsia tended to have less frequent migraine attacks. Like migraine, palinopsia has been associated with cortical hyperactivity and cortical spreading depression (97).
Effects of adaptation on flicker detection and discrimination
Elevated thresholds in migraine have been found for the detection and discrimination of changes in contrast to blurred spots or edges, after adaptation to 70 Hz and 10 Hz flicker (25). The authors suggested that this increased adaptation was due to either increased levels of neural fatigue, or a difference in gain control. A similar pattern of results was found (99) at a cortical level of processing, at which neurons respond to inputs from both eyes.
Habituation and adaptation are different in migraine
The fact that, on the whole, aftereffects are increased in migraine indicates differences in habituation and adaptation processes. A lack of habituation to stimuli might plausibly result in a longer-lasting response, and possibly even a build-up of the response, without time to return to baseline in between. However, these differences are difficult to interpret as there are multiple explanations of aftereffects (e.g. Webster (92)).
Theoretical accounts of visual processing differences in migraine
There are a number of differences in visual processing in migraine. Despite variation between individuals and studies, some broad generalisations can be made. Vision in migraine can be characterised as showing (i) reduced contrast sensitivity, (ii) increased effects of adding noise to visual stimuli, (iii) poorer global motion and form perception and (iv) stronger, longer-lived aftereffects.
A number of theoretical accounts of these differences have been proposed. Once such theory relates to evidence suggesting that the brains of people with migraine show greater levels of activity than others’. This has been termed ‘hyperexcitability’, but there is some variation in how this term is used. Coppola and colleagues use hyperexcitability specifically to mean a response to an external stimulus that is larger than that typically observed (100). Other researchers (e.g. Shepherd (89)) use a broader definition to include both increased baseline activation and an increased response to visual stimulation. This variation in the use of terminology highlights the need for precisely specified theoretical models to understand visual processing. Here, we consider the way that these models have been developed in vision science, and how they may be applied to understand migraine.
A general signal processing model
At a very general level, we can consider the activity of visual neurons to consist of a component that is informative about the input visual image, and a component that is not. This has been summarised mathematically in many papers, but a simple, general expression following Zhaoping (101) is given by:

Here, The signal processing model outlined in Equations 1 and 2. The input signal is transformed by the encoding function. The resulting output is the sum of the transformed signal and additive and (in Equation 2) multiplicative noise. The amplitude of the latter depends on the amplitude of the input signal.
The idea that the gain of the response
A further complication to the model outlined in Equation (1) is that it may also be necessary to consider multiplicative noise; that is, random variation in the response, the magnitude of which depends on the input stimulus (Figure 5). Adding a multiplicative noise (
The important distinction here is that multiplicative noise refers to an increase in activity in response to an input stimulus, but one that does not provide any information about the nature of the stimulus. It is important to note however that, despite the theoretical difference, there is no way of discriminating between multiplicative noise and a combination of gain control and late additive noise from psychophysical experiments, as their predictions are mathematically equivalent (68).
Henceforth, hyperexcitation will be used to describe either of these possibilities; the more specific terms will be used wherever possible. As we shall see, this distinction is critical to understanding the results of physiological and psychophysical data.
Accounting for visual processing differences in migraine
With this very general theoretical framework in place, we can now consider some of the explanations of sensory processing differences, and how they relate to the components outlined in the previous section. It has been proposed that the cortex is hyper-responsive (100) or hyperexcitable (103) in migraine, and also that it is associated with a reduction in neural inhibition (e.g. Palmer et al. (71)) or habituation (104). Each of these ideas, in turn, may be related to the model of visual encoding outlined above, as well as to subsequent processing of the encoded information.
Hyper-responsiveness
This is the idea that the brain responds more strongly to sensory stimuli in migraine. This would be represented by an increase in the gain of the function
Hyperexcitability
Increased neural activity in migraine need not necessarily result from an increase in responsiveness to sensory stimuli. Thus, while activity may be higher overall, this might arise not from an increase in sensitivity to visual stimulation
Reduced inhibition
Many aspects of visual processing depend on both excitatory and inhibitory interactions between neurons. It has been proposed that inhibitory interactions between neurons are reduced in migraine (105). This would manifest itself as increased
Reduced habituation
If a visual stimulus is repeatedly presented to an observer, the visually evoked potential tends to decrease. This process of habituation is less pronounced in migraine (106). The temporal dynamics of visual processing are not modelled by Equation (2), but this could be extended to allow its parameters to change over time. In such a model, a difference in habituation would then reflect differences in the way that the gain function
Application of signal processing models of vision to migraine
The ideas outlined above provide a very general way of considering the neural processing of visual signals. A variety of conceptually very similar models are used in vision science (107), and some researchers have applied them to migraine (51,74). Wagner and colleagues concluded that increased masking effects in migraine were due to increased multiplicative noise (51), whilst in later years they claimed that poorer performance to discriminate a masked shape was due to differences in gain control in the migraine group (74). Mathematically, the two formulations make identical predictions for the types of psychophysical data modelled. This highlights the difficulty, if not impossibility, of determining the single correct model to account for performance, even when physiological evidence is taken into account (108–112). In addition, the template of the Lu and Dosher model is task-specific, and therefore would take a different form depending on what the observer was doing. For these reasons, Equation (2) is used merely to show the effects that putative differences in visual processing would have in migraine. This is not presented as the single correct model that should be used, but as a simple, generalised formulation. For example, the model here is restricted to interictal visual function. This could be achieved for example if the parameters of the model varied across the migraine cycle. Additionally, this is a model of visual processing; however, a similar principle might plausibly apply to other sensory modalities.
Theoretical explanations of visual processing differences in mirgaine
Reduced contrast sensitivity
The ability to detect a stimulus relies on the sensitivity of the system, but the signal must also be strong enough to overcome the level of noise intrinsic to the system (113). Reduced sensitivity, or responsiveness, of the system, indicated by the encoding function
Increased effects of externally added noise
As previously mentioned, adding noise to a stimulus is in general expected to make performance worse. There is however more than one way of adding external noise to a stimulus, and of how this then affects visual processing (70).
Two studies have found that contrast detection thresholds for small targets are no different in migraine when presented in isolation, but worse in the presence of an externally added noise mask (51,52). Following Lu and Dosher’s noisy perceptual template model (107), these results were interpreted either as showing increased multiplicative noise (
Poorer global pooling
Poorer performance on global form and motion tasks could be accounted for in terms of decreased gain (
However, any differences in the inhibitory and excitatory connections that comprise the local association fields would also be expected to compromise performance on global form and motion tasks. Additionally, a specific deficit for global tasks might be the result of increased noise at a later stage of processing from the detection level, of a form that is not captured by Equation 2.
Increased strength and duration of aftereffects
Aftereffects need to be considered as a multistage process. Firstly, there is the adaptation phase, where cells habituate to the constant stimulation, and secondly, the ‘test’ phase, when the aftereffect is experienced. If there is reduced habituation in the adaptation phase, this would result in reduced aftereffects in the test phase. If inhibition is reduced in the adaptation phase, then this might predict an impoverished adapting stimulus, as there will be reduced selectivity and therefore more noise. An increase in the magnitude and duration of the MAE is unlikely to result from a heightened response to the adapting visual stimulation, since neither of these effects result from an increase in the physical contrast of the adaptor (115).
During the test phase, an increased aftereffect could result from reduced dishabituation to the stimulus (116). Shepherd has argued that the differences between migraine and control groups can best be explained by a slower recovery in migraine resulting in extended suppression of excitatory connections (89). This is consistent with increased effects of externally added noise.
Summary
An increase in neural noise, either increased background noise (
The increased additive noise (
The focus of this review is on vision; however, there are other sensory modalities that are important in migraine. For example, as well as photophobia, phonophobia (increased sensitivity to sound) and osmophobia (increased sensitivity to odours) are also symptoms of migraine attack (1). Analogously to visual stimulation, exposure to sound can cause discomfort (117). As well as analogous sensory modalities, there is also evidence of multisensory processing differences in migraine (118). It has been argued that lowered pain thresholds in migraine are accompanied by lowered thresholds to light, sound, and olfactory stimulation and there is also greater integration of information from different sensory modalities, and greater functional connectivity in migraine (117). Greater functional connectivity could lead to increased noise if other brain regions are being recruited that are not directly involved with the task. Multisensory processing can be measured using the McGurk effect, or the flash-beep illusion, which rely on the integration of visual and auditory inputs. On a mechanistic level, the integration period of the flash-beep illusion in non-clinical populations is related to the frequency of alpha band (8–14 Hz) neural oscillations over the visual cortex. These are ongoing rhythms of the visual cortex, thought to be the readiness state of the visual areas of the brain, and involved in controlling perception (119), termed a ‘window of excitability’ (120). As there is some evidence of a possibility to train alpha band oscillations (121,122), it might be the case that there is a potential future therapy in terms of driving these oscillations to vary the extent to which multisensory information is integrated. This is rather speculative and would need direct testing in migraine. However, there is recent evidence of increased visual/auditory integration in migraine groups (123).
As well as sensory information, the principle of overintegration might also include the vestibular system. Some types of migraine are associated with vertigo, and postural imbalances (124). Vertigo is a feeling of dizziness, associated with increased nystagmus, both spontaneous (in any position) and positional (when the orientation of the individual’s head is changed). It has been shown that while watching a ‘rotating snake’ illusion, which causes the perception of illusory motion in a static image, individuals with migraine showed less postural stability than controls (125). However, when a break was given after viewing the illusion and then measuring postural stability, the authors found no group differences. Thus they concluded that the effect of the illusory motion on postural sway in migraineurs must be related to the motion aftereffect caused by viewing the stimulus. A link between migraine vertigo and the trigeminal system has been shown: More individuals with migraine showed spontaneous nystagmus than controls (126). This number increased in the migraine group on stimulating the trigeminal system, which was not seen in controls. As nystagmus is related to vertigo, this suggests a link between the integration of visual and vestibular information in migraine.
The origin of the noise is an important outstanding question; however, a thorough account of all possible mechanisms is beyond the scope of this review. To speculate over some of the potential mechanisms, over-recruitment of different areas of the brain is one possible source of noise. Alternatively, there is evidence of genetic malfunctions relating to disturbances in ion channels that also might plausibly be a source of noise. Mice with the CACNA1A gene mutation are found to show the same clinical symptoms as familial hemiplegic migraine (127). Mutated mice showed increased Ca2+ current density, more glutamate release at excitatory cells, and greater susceptibility to cortical spreading depression, but no differences in gamma-aminobutyric acid (GABA)-controlled, inhibitory interneurons (128). The mutated mice also showed increased spontaneous activation, which might be a potential source of noise in the response to stimuli. Research has shown that the mutation on the CACNA1A gene is important in terms of migraine as this gene encodes the P/Q voltage-gated Ca2+ channels (129). It has long been noted that blocking calcium channels using drugs is helpful in both common and classic migraine (130). Pietrobon (131) argued that as P/Q Ca2+ channels act as a gatekeeper for excitatory K+ channels, this relates to the gain function of the cell. It has been shown that mice with a mutated CACNA1A, resulting in P/Q Ca2+ channel differences, showed more irregular Purkinje cell firing rates (132). This irregularity in firing rate was both with and without stimulation. Speculatively, it might be plausible that more irregular firing could be the cause of noisier signal transmission. Although this is indirect, in the fly visual system, voltage dependent gates are related to the gain control of a visual signal (133). P/Q Ca2+ channels are also associated with the increased sensitisation of the trigeminal nucleus (131). Voltage-gated P/Q Ca2+ channels are also responsible for the transmission of pain, and pain is reduced when this channel is blocked (134). Although research using animal models is mostly limited to familial hemiplegic migraine, it is thought that this and MA are part of the same spectrum of disorder (135). Therefore, there might be, in principle, genetic abnormalities leading to Ca2+ ion channel differences and thus irregular neural firing. This might represent a possible source of noise, and a possible area for future research. However, it must be emphasised that all of this is speculative for the time being.
Although GABA-controlled inhibition on a neural level seems unaffected, at least for a subtype of migraine, over-recruitment of different areas of the brain could be due to reduced inhibition between brain areas. Inhibition is a term that has multiple meanings, depending on the scale of the processes involved. For example, there is inhibition between adjacent cells, or groups of cells, responding to different features in the early areas of the visual cortex. There are also more long-range inhibitory processes, involving the suppression of alternative representations, which might be needed in the case of visual search tasks, to enable the viewer to ignore distracters (136). There are inhibitory processes that rely on timing, as is thought to be the case in some aftereffects. It is not helpful to be indiscriminate in the use of the term ‘inhibition’, and theory would benefit from clearly defining the term in context.
The effects of moving stimuli might be particularly relevant, as these are stimuli that are temporally modulated. Studies using prolonged presentations have shown on the whole that the aftereffects last longer in migraine. Aftereffects could be a measure of the ability to adapt to a stimulus, and habituate the response, and it is unclear exactly which processes are involved in an aftereffect (92), therefore more theoretically motivated experiments need to be designed before this can be assessed. It is possible that the time course of neural activity is actually the critical factor in migraine visual performance. For example, differences have been found in habituation in migraine for stimuli presented at 3 Hz only, not at other presentation rates (137). The association with flicker is common in migraine, and needs to be considered in the research. This is not accounted for by the model in its current form, and therefore this will need to be expanded to include the dimension of time.
As well as short-term temporal effects, it is also important to consider the model in terms of the progression of the disease. It has been shown that attack frequency, and duration of the disorder, both associate with more damage to white matter in the brain (138). Although there has been some debate over whether migraine causes damage to the brain, a meta-analysis suggests that overall, migraine is associated with increased white matter lesions (139). Whether the lesions occur due to migraine attacks, or are a cause of the attacks, is as yet unclear. This is of importance to sensory processing as white matter lesions are also found in children with sensory disorder (140), and could therefore be related to poorer visual performance in migraine. White matter lesions in older adults were associated with both increased functional connectivity, and poorer performance on a visual task (141). This could be a possible cause of increased noise in the brain. Although this has been demonstrated in a non-clinical population of older adults, this is not necessarily applicable to migraine, and so the argument remains speculative.
One possible therapy that is relevant to noise is tinted filters. These have been demonstrated to be of benefit in migraine (142). Huang et al. (21) demonstrated a possible mechanism for this in terms of a reduction in fMRI BOLD response to stimuli when viewed through tints. The tints are specific to the individual with migraine in order to have any benefit. Although the mechanism for this is unclear, one might speculate that increasing the specificity of the response would help with increased noise/reduced gain.
Conclusion
Migraine is a common and debilitating neurological disorder. There are many associations between migraine and vision, including visual triggers of attacks, visual auras and differences in performance on visual tasks compared to control groups. Importantly, there are a number of low-level differences that suggest fundamental differences in sensory processing in migraine. These can be summarised as (i) reduced contrast sensitivity, (ii) reduced sensitivity to differences in orientation, (iii) increased effects of adding noise to stimuli, (iv) poorer perception of global motion and form and (v) stronger, longer-lived aftereffects.
A number of theoretical accounts of these differences have been proposed in the literature. One difficulty with comparing these has been some degree of variation in the way that terminology is used, and a lack of precise definition of this terminology (99). Overall, differences in migraine could be accounted for by increased effects of noise, and/or differences in excitatory and inhibitory processes. Increased clarity at the level of theoretical accounts of sensory processing differences is important in driving novel empirical work, with a view to a better understanding of the disorder and the development of novel treatments.
Article highlights
Migraine is a common disorder, associated with visual processing differences. The review focusses on behavioural studies of visual processing in migraine. Migraine groups show reduced sensitivity to contrast, and orientation, and difficulty processing moving stimuli. Suggested theories include hyperexcitation, or an inability to exclude noise. An established mathematically defined signal-processing model is applied, to clarify terminology and highlight future research and treatment implications.
Footnotes
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.