
Abstract
Introduction: Although migraine is classified as a headache disorder, a key part of migraine pathophysiology is a heightened excitability of visual cortices in between headache events. The goal of our study was to examine the behavioral impact of this visuocortical hyperexcitability, in terms of its effect on reflexive visual attentional orienting.
Methods and results: In Experiment 1, using a non-predictive spatial cuing task that relied on sensory-evoked responses in the visual cortex for triggering attentional orienting, we found that migraineurs had greater attentional enhancement of manual target responses, relative to non-migraine controls. In two control experiments we confirmed that this heightened attention effect in migraineurs is not due to exaggerated reflexive orienting responses in general, but rather, it appears to be specifically associated with sensory-evoked attentional triggers.
Discussion: Taken together, this confirms that the functional consequences of hyperexcitable visual cortex in migraineurs are not just purely sensory in nature, but directly impact at least some forms of reflexive attention. This provides evidence of at least one cognitive implication of hyperexcitable visual cortical responses in migraineurs, namely heightened reflexive visual-spatial orienting specific to sudden-onset peripheral events.
Introduction
Although migraine is classified as a headache disorder, a key part of migraine pathophysiology is a heightened excitability of sensory cortices in between headache events. In vision, for example, migraineurs are more susceptible to visual illusions (1), have more discomfort in the presence of intense or repetitive patterns of visual stimulation (2), and show reduced sensory habituation to repetitive visual stimuli (3–7). Given these visual sensory issues in migraine, the goal of our study here was to examine their possible impact on reflexive visual attentional orienting.
At issue in our study is whether visual cortical hyperexcitability in migraineurs may have functional consequences beyond the initial sensory responses themselves. In particular, visual-spatial attention is automatically drawn to the location of sudden-onset, non-foveal visual stimuli in an automatic, bottom-up manner (8–10). On the possibility that the magnitude of reflexive orienting may increase with the intensity of the sensory-evoked response to a visual stimulus, this suggests that migraineurs may not just be affected by an exaggerated sensory response itself, but also from the subsequent effect this has on reflexive attentional orienting. In other words, does visuocortical hyperexcitability in migraine lead to heightened reflexive orienting to sudden-onset events in the visual periphery?
That it might is consistent with several lines of converging evidence. For one, at a subjective level, migraineurs frequently report having difficulty ignoring distracting visual stimuli (11). Likewise, in the laboratory, migraineurs have been shown to have a reduced ability to suppress extraneous visual noise (12). Indeed, even when migraineurs consciously orient their attention to a discrete location in visual space, they nevertheless manifest heightened sensory responses to visual events outside their zone of attentional focus, relative to non-migraine controls (13). Collectively, this suggests that migraineurs have systematic impairments in attenuating attentional responses to non-foveal visual stimuli, and precisely aligns with what would be predicted if they showed heightened reflexive visual-spatial orienting to sudden-onset non-foveal events.
Towards testing this prediction, we examined migraineurs reflexive visual-spatial orienting in Experiment 1 using a canonical peripheral attentional cuing paradigm. On each trial participants made speeded detection responses to small, brief targets presented in either the upper left or upper right visual field quadrant. The target locations themselves were demarcated by boxes that remained onscreen throughout each trial block. Prior to each target presentation, one of the two box outlines brightened briefly, serving as the sudden-onset, sensory-driven attentional cue. On half of the trials the target was presented at the ‘cued’ location, and on the other half of the trials the target was presented at the opposite, or ‘uncued’, location. In these paradigms, people are faster and more accurate in their responses when the target is presented at the cued location within 200–300 ms of cue onset, relative to targets presented at the uncued location, the behavioral signature of attention being reflexively oriented to the cued location (8,14). If migraineurs do in fact have heightened reflexive visual-spatial orienting to sudden-onset non-foveal events, then it is predicted that they should have a larger effect of attention in reaction time (RT) performance, relative to non-migraine controls.
Experiment 1
Materials and methods
Participants
A total of 40 paid volunteers participated, 20 in the non-migraine control group (15 women and 5 men) and 20 in the migraine group (17 women and 3 men). The control group mean age was 22.2 (SD 5.1) years and the migraineurs were 23.9 (SD 4.1) years, with no significant difference between group age [F(1,38) = 1.344; p = 0.254]. The migraineurs had 15.4 (SD 22.6) headaches a year, while the controls had 3.0 (SD 3.1) headaches a year (non-classified as migraine; see below).
Headache classification
All migraine participants were required to meet the migraine criteria specified by the International Headache Society (15) and determined by an interview. Specifically, in order to be included in the migraine group, participants had to meet a basic minimum criterion including five or more attacks with headache lasting 4–72 hours. In turn, the headaches needed to include two of the following: unilateral pain; pulsating pain quality; moderate to severe pain; and pain aggravated by routine physical activity such as walking or climbing stairs. The headache also had to be accompanied by nausea and vomiting or photophobia and phonophobia. All our migraine participants had not suffered from a migraine for at least 48 hours before testing, nor did they have a migraine for at least 48 hours after testing. In addition to our headache classification criterion, migraineurs were excluded if they were taking any form of migraine prophylactics.
Stimuli and timing
Stimulus timing and sequence are shown in Figure 1. Each trial started with the presentation of a non-foveal (peripheral) cue to the left or right of fixation. Following the cue, a small target dot was presented in one of the two peripheral locations. On half the trials the target was presented at the cued location, and on the other half of the trials the target was presented at the uncued location. As such, the cue was unpredictive as to the location of the pending target. As well, because reflexive visual attentional orienting to sudden-onset peripheral events at short cue-target delays gives way to inhibition of return (or IOR; faster RTs for uncued vs. cued targets) at longer cue-target delays (8,14), we randomly varied the onset of cue to the onset of target between trials (170 ms vs. 960 ms) to confirm this normative pattern of reflexive attentional orienting. The target locations, one in each upper visual-field quadrant, were demarcated by the outlines of 1.0° white square boxes. These boxes were located 3.2° (to center) from fixation and 1.3° (to center) above the horizontal meridian. The non-foveal cue was a brief brightening of one of the two boxes. The fixation was a 0.2° fixation cross; the target dot was 0.1°.
Basic trial conditions for Experiment 1. Cued and uncued trials are shown.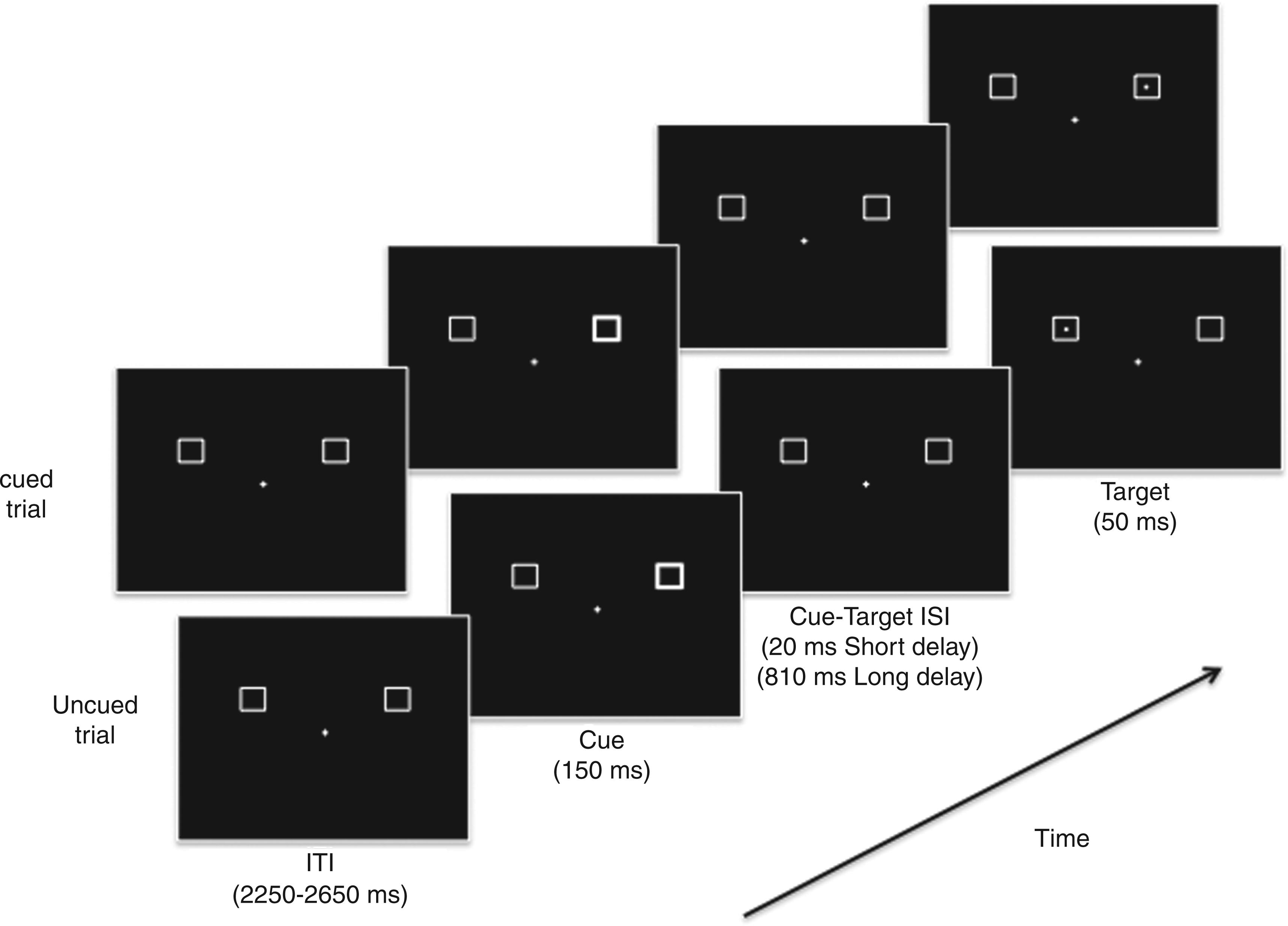
Procedure
Participants were required to press a button as quickly as possible after the appearance of the target dot on the screen. A cued trial was defined as a target at the cued peripheral location and an uncued trial was defined as a target at the uncued peripheral location. On 20% of the trials (catch trials), only a cue was presented, in order to eliminate anticipatory responses. Each participant completed 12 blocks of 50 trials (20 cued, 20 uncued and 10 catch trials per block), with the target trials equally split (but randomly varying) between short and long cue-target delays, so as to decrease expectation effects. No feedback was given on hit or miss rates.
Results
Reaction times
To examine possible age- and gender-related effects in our primary findings reported below, we initially ran repeated-measures analyses of covariance (ANCOVAS) with sigma-restricted coding of categorical predictors for sex and age covariates. As we found no significant effects of age or gender (all F < 2.773; all p > 0.105), we did not include these covariates in the main results.
Target-present trials
Mean RTs to targets are presented in Figure 2 as a function of group, attention, and cue-target delay. As can be seen, it appeared that migraineurs had a greater attention effect at short cue-target delays, relative to controls. To confirm this data pattern we assessed RTs via repeated-measures analyses of variance (ANOVAs) that included headache category (migraine vs. control) as a between-subjects factor and attention (cued vs. uncued targets) and cue-target delay (short vs. long delay) as within-subjects factors. We found a significant interaction between group, cue and cue-target delay, [F(1,38) = 4.178; p < 0.05). Performing separate follow-up t-tests within each group, both groups did indeed show a significant cuing effect at the short cue-target delay [for controls, t(19) = −2.891; p < 0.01; for migraineurs, t(19) = −3.853; p < 0.001] and a significant IOR effect at the long cue-target delay [for controls, t(19) = 7.477; p < 0.001; for migraineurs, t(19) = 8.592; p < 0.001). Importantly, however, separate follow-up between-groups ANOVA within each cue-target delay condition revealed a significant group × cue interaction at the short cue-target delay [F(1,38) = 5.892; p < 0.05], indicating a greater cuing effect for migraineurs vs. controls, but no such interaction was found at the long cue-target delay [F(1,38) = 0.955; p = 0.335].
Mean reaction times (RTs) to targets as a function of group, attention, and cue-target delay.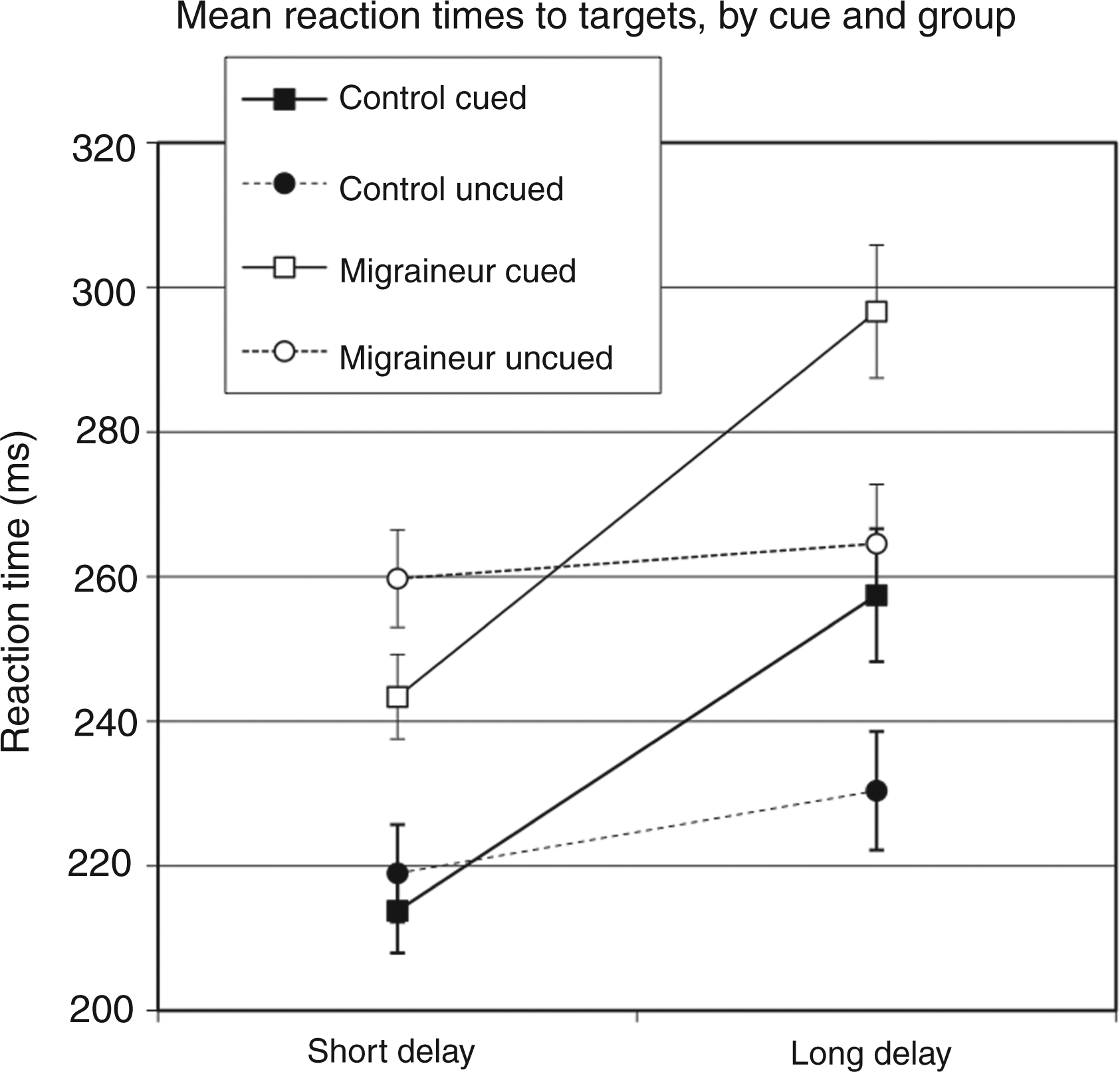
Catch trials
On 20% of the trials (catch trials), only a cue was presented, in order to eliminate anticipatory responses. The proportion of catch trials in which participants had a button press did not differ between groups [F(1,38) = 0.008; p = 0.931], with both groups responding on about 2% of catch trials (migraineurs mean 2.0%, SD 2.7%; controls mean 1.9%, SD 2.5%).
Control analyses
Proportional analysis
In addition, it appears that migraineurs respond more slowly overall to targets than controls, and this was confirmed statistically [F(1,38) = 6.429; p < 0.05). Although our RT findings suggest that migraineurs show greater cuing effects than non-migraineurs, an alternative explanation is that the apparent effect may be an artifact of these overall slower RTs in migraineurs. For example, if migraineurs simply respond at some constant rate slower than controls, we would expect to see an equal proportional increase in RTs between cued vs. uncued conditions in both groups. This would suggest that both groups are equally affected by the cue. To address this possibility that the increased magnitude in effects found may be a consequence of this overall speeded difference, we ran an additional analysis comparing the proportional difference between the cued and uncued targets. The proportional analyses revealed that indeed, the difference between groups held for the short cue-target delay [F(1,38) = 5.918; p < 0.05], but not for the long cue-target interval [F(1,38) = 0.027; p = 0.870), which demonstrates that the difference is not simply due to a slower overall response time by migraineurs.
Migraine subgroups
Some studies report differences between migraine with aura (MA) and migraine without aura (MO) groups. Accordingly, we divided the migraine group into more specific headache categories, with 8 MA and 12 MO. There were no interactions between subgroup and attention [all F(1,18) < 1.01; all p > 0.328] and no overall differences in reaction speeds between subgroups [F(1,18) = 0.05; p = 0.836).
Discussion
The results of Experiment 1 suggest that migraineurs do in fact have heightened reflexive visual-spatial orienting to sudden-onset non-foveal events, an effect on attention consistent with known visual cortical hyperexcitability in migraine populations (16,17). However, an alternative possibility is that migraineurs may simply have greater reflexive attentional orienting responses in general, the by-product of a learned response to visual hypersensitivities rather than a direct result of heightened sensory responses per se. To test this possibility, we performed a second experiment that triggered reflexive visual-spatial orienting via so-called ‘feature singletons’.
In particular, in attentional capture paradigms, participants are asked to respond to targets that are not presented in isolation, but rather in the context of multiple surrounding distractors that are presented simultaneously. On some trials the distractors are identical in terms of features like shape and color, and only the target is unique among the array of items. On other trials, however, one of the distractors is uniquely different from the others (e.g. a different color or shape), a manipulation designed to reflexively draw attention to this ‘feature singleton’. The extent to which attention is actually ‘captured’ by the singleton is determined by comparing target RTs on singleton present vs. absent trials – longer RTs on singleton present trials indicate ‘capture’, or reflexive attentional orienting (18). If the results of Experiment 1 were really tied to overall greater attentional sensitivities in migraineurs rather than visual cortical hyperexcitability, then they should show similar evidence of increased reflexive orienting in attentional capture paradigms. Conversely, if the results of Experiment 1 were due to sensory cortical hyperexcitability, migraineurs should show no difference from controls.
Experiment 2
Materials and methods
Participants
A total of 56 paid volunteers participated in our study, 28 in the non-migraine control group (20 women and 8 men) and 28 in the migraine group (21 women and 7 men). The control group mean age was 22.8 (SD 3.3) years and the migraineurs were 25.5 (SD 7.1) years, with no significant difference between group age [F(1,55) = 3.491; p = 0.067). The migraineurs had a mean of 22.7 (SD 32.9) migraine headaches a year, while the controls had a mean of 11.7 (SD 21.9) non-migraine headaches a year. Headache classification was performed in the same manner as in Experiment 1.
Stimuli and procedures
Stimulus sequence and timing are shown in Figure 3. A black central circular fixation point (0.5 cm diameter) was located in the center of the computer screen and remained on screen for the duration of each trial block. On each trial, four stimulus elements – consisting of three squares and one diamond – were presented, spaced randomly at equal intervals above, below, left, or right of fixation around the boundary of an imaginary circle surrounding fixation (0.57° radius). The open black squares had a line weight of 0.5 pt (0.38° sides) and served as the distractors on each trial. The target shape singleton was an open black diamond shape with similar dimensions and luminance to the squares except for being oriented at a 90° angle to create the diamond shape. Presented within each stimulus element was a small black bar oriented either vertically or horizontally (0.19° in length). The task required participants to signal on each trial the orientation of the bar within the diamond (vertical vs. horizontal). In the singleton distractor condition, which occurred in half the presentations, one of the three distractor squares was red in color, rather than black. All variables in the paradigm – diamond location, target bar orientation, orientation of bars within the distractors, and location of the singleton distractor (when present) – were randomly varied within each block of trials. In terms of responding, horizontal vs. vertical line was indicated using button presses on a hand-held response device with the left vs. right thumb, with the orientation/thumb mapping counterbalanced between participants. Participants were told to ignore the singleton distractor, to maintain fixation at all times and to respond as quickly and as accurately as possible during the task. No feedback was given on hit or miss rates. Stimuli were viewed at a distance of 150 cm. Stimuli were presented in 24 blocks of 24 trials. Each session was completed in approximately 1 hour.
Basic trial conditions for Experiment 2. No distractor and distractor singleton trials are shown.
Results
To examine possible age- and gender-related effects in our primary findings reported below, we initially ran repeated-measures ANCOVAS with sigma-restricted coding of categorical predictors of sex and age covariates. As we found no significant effects of either age or gender (all F < 2.201, all p > 0.144), we did not include these covariates in the main results.
Reaction times
Mean RTs are presented in Figure 4 as a function of group and attention. Both groups appeared to show attentional capture effects in RT, such that the RTs were slower when a singleton distractor was present vs. absent, but this effect did not appear to differ in magnitude between groups. This was confirmed via an omnibus ANOVA that included group (migraine vs. control) as a between-group factor and capture condition (singleton present vs. absent) as a within-groups factor. This analysis revealed a main effect of capture [F(1,54) = 27.144; p < 0.001), but no interaction between capture and group [F(1,54) = 0.078; p = 0.781). As well, there was no significant main effect of group [F(1,54) = 0.009; p = 0.926), indicating overall RTs were comparable between groups.
Mean reaction times (RTs) to targets as a function of group and capture.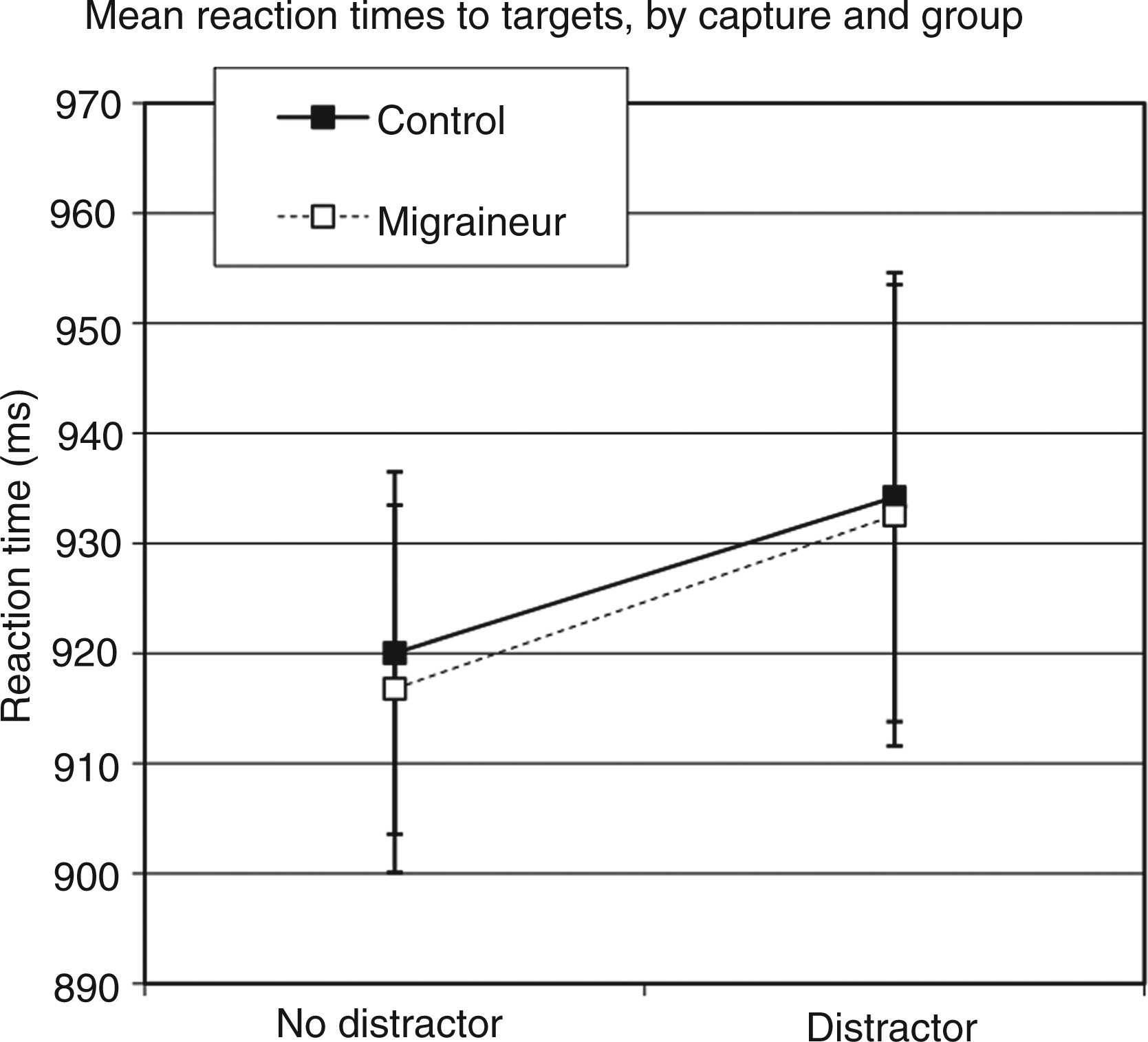
Accuracy
Accuracies for Experiment 2 as a function of group and singleton condition

Control analyses
Subgroups
To examine whether the type of headache affected task performance, we divided the participants into more specific headache categories, with 6 MA, 22 MO, 21 controls with tension-type headache, and 7 controls with no headache history. There were no interactions between subgroup and capture condition [for RTs F(3,52) = 0.276; p = 0.843; for accuracy F(3,52) = 1.440, p = 0.242) and no overall differences in reaction speeds between subgroups [for RTs F(3,52) = 1.457; p = 0.237; for accuracy F(3,52) = 0.557; p = 0.646].
Discussion
In Experiment 2 we found no difference between migraineurs and controls in terms of reflexive attentional orienting as captured by color feature singletons. This suggests that migraineurs don’t necessarily have heightened reflexive attentional orienting in general, but rather, they have heightened reflexive orienting linked to hyperexcitable visual cortical responses to sudden-onset peripheral events. However, with respect to this conclusion, it could also be the case that the need to attend peripherally – which was present in Experiment 1 but not Experiment 2 – may drive the cuing effects seen in Experiment 1, and not visual cortical hyperexcitability per se. In other words, migraineurs might show heightened reflexive orienting to the visual periphery regardless of the attentional trigger, something the paradigm in Experiment 2 could not assess due to the relatively close spatial proximity all the stimuli had to fixation.
To test this possibility, we thus ran a third experiment, using non-foveal targets as in Experiment 1 but that triggered reflexive visual orienting via eye gaze cues presented at fixation. The notion that people orient their attention to where someone else is looking has been confirmed in studies revealing that eye gaze cues trigger reflexive shifts in attention (19,20). If migraineurs show heightened reflexive orienting to the visual periphery in general, then it predicted that they should have a greater attentional cuing effect in this paradigm, relative to controls. Conversely, if heightened reflexive orienting in migraineurs is really driven by hyperexcitable responses to peripheral visual stimuli, then it predicted that migraineurs should have comparable attentional responses in this paradigm, relative to controls.
Experiment 3
Materials and methods
Participants
A total of 34 paid volunteers participated in our study, 17 in the non-migraine control group (14 women and 3 men) and 17 in the migraine group (14 women and 3 men). The control group mean age was 20.2 (SD 1.6) years and the migraineurs were 21.7 (SD 3.1) years, with no significant difference between group age [F(1,32) = 3.314; p = 0.078). The migraineurs had a mean of 26.8 (SD 26.6) headaches per year while the controls had a mean of 16.2 (SD 13.8) non-migraine headaches a year. Headache classification was performed in the same manner as in Experiment 1.
Stimuli and timing
Stimulus timing and sequence are shown in Figure 5. Each trial started with the foveal presentation of an image of eyes cuing either the left or the right visual field. Following the cue, a target X was presented in one of the two peripheral locations. On half the trials the target was presented at the cued location, and on the other half of the trials the target was presented at the uncued location. As such, the cue was unpredictive as to the location of the pending target. The target locations, one in each upper visual-field quadrant, were located 3.1° (to center) from fixation and 0.4° (to center) above the horizontal meridian. The foveal cue was an image of eyes 2.1° × 1.1°, with pupils directing attention to either the left or right visual field. The fixation was a 0.3° fixation cross; the target X was 0.3° × 0.5°.
Basic trial conditions for Experiment 3. Cued and uncued trials are shown.
Procedure
Participants were required to press a button as quickly as possible after the appearance of the target on the screen. A cued trial was defined as a target at the cued peripheral location and an uncued trial was defined as a target at the uncued peripheral location. Each participant completed an average of 30 blocks of 20 trials (50% cued and 50% uncued) per block. The time cue is on screen before the target appears was varied to decrease anticipatory responses. No feedback was given on hit or miss rates.
Results
To examine possible age- and gender-related effects in our primary findings reported below, we initially ran repeated-measures ANCOVAS with sigma-restricted coding of categorical predictors of sex and age covariates. As we found no significant effects of either age or gender (all F < 1.460, all p > 0.236), we did not include these covariates in the main results.
Reaction times
Mean RTs are presented in Figure 6 as a function of group and attention. Both groups appeared to show attention effect in RT, such that the RTs were faster for cued than uncued targets, but this effect did not appear to differ in magnitude between groups. This was confirmed via an omnibus ANOVA that included group (migraine vs. control) as a between-groups factor and attention (cued vs. uncued) as a within-groups factor. This analysis revealed a main effect of attention [F(1,32) = 24.172; p < 0.001], but no interaction between attention and group [F(1,32) = 0.110; p = 0.742). As well, there was no significant main effect of group [F(1,32) = 0.724; p = 0.401], indicating overall RTs were comparable between groups.
Mean reaction times (RTs) to targets as a function of group and attention.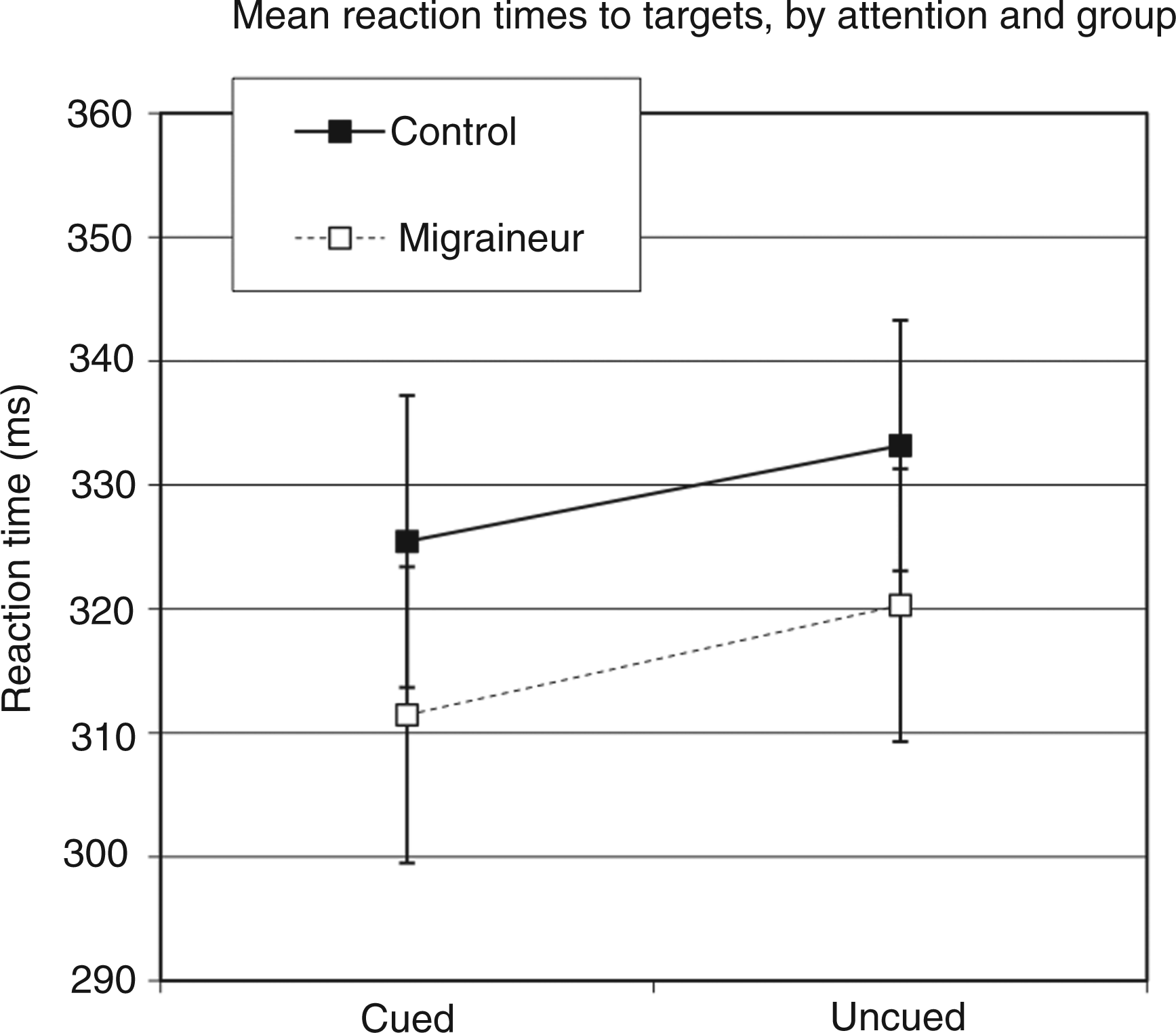
Control analyses
Subgroups
To examine whether the type of headache affected task performance, we divided the participants into more specific headache categories, with 6 MA, 11 MO, and 17 controls with no migraine history. There were no interactions between subgroup and attention [for RTs F(2,31) = 0.066; p = 0.936] and no overall differences in reaction speeds between subgroups [for RTs F(2,31) = 0.780; p = 0.467].
Discussion
In Experiment 3 we found no difference between migraineurs and controls in terms of reflexive attentional orienting as triggered by a central eye gaze cue. This confirms that migraineurs do not have heightened reflexive attentional orienting to the visual periphery in general. Rather, we can conclude that migraineurs have heightened reflexive orienting linked to hyperexcitable visual-cortical responses specific to sudden-onset peripheral events.
Migraineurs across experiments
Notably, in all three of our experiments we found no significant differences between headache classifications (aura vs. non-aura) within migraineurs. One possibility is that we may not have had sufficient power within each experiment to find significant differences between headache groups. Accordingly, to investigate this possibility, we conducted an additional analysis within the migraineurs only, pooled across all three experiments, using a normalized ‘attentional effect score’. To create a normalized score, within each experiment we calculated a difference score for attended vs. unattended stimuli (or the equivalent) and then based on the migraineur group mean within each experiment, calculated a z-score for each migraine participant. Then, for the cross-experiment comparison, we compared the z-scores in an ANOVA with factors of headache classification (MA vs. MO) and Experiment (Experiment 1 short SOA, Experiment 1 long SOA, Experiment 2, and Experiment 3). There were no significant main effects or interactions [all F(1,77) < 1.235; all p > 0.303].
General discussion
We asked if migraine hyperexcitability in visual cortex leads to heightened visual attentional orienting responses. Experiment 1, which triggered attention via sudden-onset peripheral events, confirmed that indeed migraineurs do have heightened reflexive orienting. The two follow-up experiments revealed that this heightened orienting is not due to overall increased reflexive attentional responses (Experiment 2 and 3) or simply to attending to peripheral stimuli (Experiment 3), but specifically to attentional orienting in response to sudden-onset peripheral events. Given our conclusions, several critical questions follow.
What are the implications for migraineurs?
Overall, our study reveals that migraineurs have heightened reflexive visual attentional orienting as a result of a hyperexcitable response to sudden-onset peripheral events. What is the impact of this finding for migraineurs? First, it is important to consider the proposed function of reflexive orienting. Reflexive orienting allows one’s attention to be drawn to new stimuli or salient changes in the environment to aid in our survival in a complex world. With enhanced orienting response, one would expect to be more aware of sudden changes in environmental stimuli, and in fact, migraineurs often report their attention is drawn to unwanted peripheral distractions (11). It is important to note that this is the first study looking at IOR in migraineurs, and it is found to be normal in migraineurs. The function of IOR is thought to encourage proper search of the environment (21). That this is intact in migraineurs means that while their attention is more drawn to locations of sudden-onset stimuli in the environment, their attention is just as able as controls to then block out that already-checked area to the advantage of searching new areas in the environment.
Interestingly, our findings are also consistent with the recent result of Boulloche et al. (22). In their study they found that during visual stimulation, migraineurs showed increased activity in regions of parietal cortex associated with volitional attentional orienting, whereas controls did not. These neuroimaging results paralleled prior behavioral studies suggesting migraineurs have slowed response performance relative to controls due to their attention getting grabbed by visual stimuli (23,24). Not only do our data replicate this latter result – we found overall slowed RTs in migraineurs – but we also provide direct evidence of the hypothesized attentional grabbing in migraineurs, in the form of heightened reflexive orienting. At the same time, this heightened reflexive orienting could also explain the increased activity in migraineurs posterior parietal cortex reported by Boulloche et al. (2010), in that if attention was reflexively grabbed by visual stimuli, controlled attentional processes would need to be invoked to countermand these automatic effects.
Acknowledging that enhanced orienting fits with the migraine profile, are there any advantages to having enhanced orienting and to knowing that this is an issue in migraine? Regarding perceptual advantages, one can imagine many tasks, such as driving, where heightened reflexive orienting could mean vital seconds. On the other hand, to the extent that such heightened orienting may be contributing to triggering migraine events, one could potentially use this information therapeutically to reduce headache frequency. While one can think of very simple adjustments a migraineur could do to limit distracting stimuli, such as sitting so a flashing television is behind them in a restaurant, we would also use attention literature to envision clinical therapeutic possibilities. For example, recent evidence suggests that action video-game playing leads to enhanced ability to suppress distracting irrelevant information (25). Given migraineurs’ heightened orienting to sudden-onset peripheral events, and the potential for video-gaming to suppress this heightened orienting, future research could assess the therapeutic option of using video-gaming to reduce sensory-triggered migraine events. This is just one example of how a growing understanding of the cognitive consequences of hyperexcitable sensory cortices may be used to improve daily life in migraineurs.
What are the implications for attention?
In Experiment 1, we found that migraineurs had increased attentional orienting at short cue-target intervals, but normal IOR for long cue-target intervals. Why might we find heightened facilitation but intact IOR? Facilitation effects from reflexive orienting are known to reflect the earliest sensory-level enhancement of visual processing, modulating the early posterior P1 ERP component occurring around 100 ms post-target (26). Indeed, that migraineurs have a heightened response specific to early-sensory level processing fits well with our previous findings that migraineurs have altered voluntary attentional modulation of sensory-level visual processing (13), as both of these effects are driven by early modulation of extrastriate cortex (26,27). On the other hand, IOR and attentional capture are both thought to reflect later processing, modulating slightly later ERPs, such as the posterior N2pc ERP component, around 175–300 ms post-stimulus (26,28). This suggests that these hyperexcitable cortical implications of migraine are specific to the earliest sensory processing.
This finding is also an example of how migraineurs can provide neuropsychological evidence for dissociating reflexive attentional paradigms. Essentially, a sensory model of reflexive orienting would suggest that the sensory response to a peripheral flash (i.e. cue) draws attention to a location, producing the heightened sensory cortical response (9) and faster RTs (29) to the subsequent target. Because migraineurs have heightened excitability of sensory cortices, that they show altered reflexive orienting only to sudden-onset peripheral targets supports a dissociation between this type of early sensory response reflexive orienting and other types of reflexive orienting in which attention is reflexively cued to the periphery (e.g. eye gaze) or when attention is captured by a salient object (attentional capture). Thus, migraineurs provide novel neuropsychological evidence to support the theory that there are multiple reflexive mechanisms acting in the brain to bias different stages of sensory information processing and that these mechanisms are triggered by different stimulus attributes (26).
Footnotes
Funding
This research was supported by the Migraine Trust, UK, NSERC, and MSFHR.