
Abstract
Background and methods: Data on the association between the SLC6A4 5-HTTLPR polymorphism and migraine are conflicting. We performed a systematic review and meta-analysis among studies published up to September 2009. For each study with genotype information, we calculated odds ratios (OR) and 95% confidence intervals (CI) assuming additive, dominant, and recessive genetic models. We then calculated pooled ORs and 95% CIs.
Results: Among the ten studies identified there was no overall association between the polymorphism and any migraine for Europeans or Asians. However, European women carrying the S allele had an increased risk for any migraine (dominant model: pooled OR = 2.02; 95% CI 1.24–3.28). Results among Europeans further suggested an increased risk for migraine with aura among carriers of the S/S genotype (recessive model: pooled OR = 1.41; 95% CI 0.83–2.40).
Conclusions: While our results indicate no overall association between the SLC6A4 5-HTTLPR polymorphism and migraine among Europeans and Asians, gender and migraine aura status may have modifying roles among Europeans.
Introduction
Migraine is a common, chronic disorder affecting 10–20% of the population, women 3–4 times more often than men (1). Up to one-third of migraine patients experience an aura prior to, or during, the migraine headache.
Despite many advances in migraine research over the last decades, migraine pathophysiology is still incompletely understood. Current concepts view migraine as an inherited brain disorder, characterised by neurotransmitter imbalances that lead to neuronal dysfunctions in the form of recurrent headache attacks and combinations of gastrointestinal and autonomic nervous system symptoms (1,2). The serotonergic system appears to play a pivotal role and alterations in serotonin metabolism and in the processing of central serotonin-mediated responses have been described as typical for migraineurs (3).
Following axonal release, serotonin (5-hydroxytryptamine; 5-HT) action is rapidly terminated by presynaptic re-uptake (4). This is performed by the serotonin transporter SLC6A4 (5-HTT) (5), which is mostly expressed in the brainstem and midbrain (6), areas implicated in migraine pathophysiology. A 44-bp insertion/deletion polymorphism in the promoter region of the SLC6A4 gene, termed 5-HTTLPR (5-HTT linked polymorphic region), has been described (7,8). This 5-HTTLPR polymorphism translates into a long (L) and short (S) allele, which show functional differences. The S allele is associated with half the number of serotonin transporters compared with carriers of the L/L genotype (9). This results in a slower synaptic clearing of serotonin in carriers of the S allele. Subsequently, it was speculated that the SLC6A4 5-HTTLPR polymorphism accounts for altered serotonin levels in migraineurs and may thus be associated with migraine.
However, results from studies looking at the association between the SLC6A4 5-HTTLPR polymorphism and migraine are conflicting. Some have reported an association with any migraine (10,11), some only with migraine with aura (12,13), and others did not find an association (14–20). This may, in part, be due to differences in genotype and allele frequencies between ethnic groups (21,22), sample size, and study characteristics.
We sought to summarise the current evidence on the association between the SLC6A4 5-HTTLPR polymorphism and migraine including migraine with aura (MA) and migraine without aura (MO) by systematically reviewing the literature and performing a meta-analysis.
Materials and methods
Selection of studies
We followed the guidelines for systematic reviews of genetic association studies (23). Two investigators (MS and PMR) independently searched MEDLINE, EMBASE, and Science Citation Index from inception to September 2009 combining text words and MESH terms, where appropriate, for serotonin and serotonin transporter (‘Serotonin Plasma Membrane Transport Proteins’ or ‘serotonin transporter’ or ‘serotonin’ or ‘SLC6A4’) with terms for genetic variations (‘gene’ or ‘polymorphism’ or ‘genetic variation’) and terms for headache and migraine (‘headache’ or ‘headache disorders’ or ‘migraine’ or ‘migraine disorders’). The search terms were combined with the ‘explode’ feature where applicable. We considered full articles without language restrictions. In addition, we manually searched the reference list of all primary articles and review articles.
A priori, we defined the following criteria for inclusion:
Studies must have a cross-sectional, case-control or cohort design and must be published as full papers. Authors must investigate patients with migraine, diagnosed according to the criteria of the International Headache Society (IHS) (24,25) and healthy control subjects. Authors must provide information on genotype frequencies of the SLC6A4 5-HTTLPR polymorphism or sufficient data to calculate these. In studies with overlapping cases and/or controls, the largest study with extractable data was included.
In a first step, two investigators (MS and TK) by consensus identified all studies not meeting any of the prespecified criteria by screening the title and abstracts. These studies were excluded. In a second step, the same investigators evaluated the remaining studies in their entirety. Studies were excluded if they did not meet all criteria.
Data extraction
Two investigators (MS and PMR) independently extracted data from the published studies and entered them in a customised database. Disagreements were resolved by consensus. The extracted data included authors and title of study, year of publication, country of origin, ethnicity of the population investigated, setting (clinic vs population), study design, genotyping method, migraine status (any migraine, MA, MO), age range and gender of study individuals, study size, allele and genotype frequencies, and information on additional genetic variants as well as gene–gene and gene–environment interactions, if investigated. If not given, genotype frequencies were calculated where possible. If provided allele or genotype frequencies did not match with the number of patients reported, the respective subgroup was excluded. We did not contact the authors to collect further information.
Statistical analysis
We first used logistic regression to calculate odds ratios (ORs) and 95% confidence intervals (CIs) for the association between the SLC6A4 5-HTTLPR polymorphism and migraine assuming additive, dominant, and recessive genetic models. The additive model assumes that the risk for migraine among carriers of the heterozygous genotype (L/S) is half way between carriers of the homozygous genotypes (L/L, S/S). While the dominant model assumes that carriers of the heterozygous (L/S) and homozygous mutant genotypes (S/S) have the same risk of developing migraine compared with carriers of the homozygous wild-type genotype (L/L), a recessive model assumes that carrying the homozygous mutant genotype (S/S) is necessary to alter the risk for migraine compared to carriers of the heterozygous (L/S) and homozygous wild-type genotype (L/L). We also determined Hardy–Weinberg Equilibrium (HWE) for each study. The Hardy–Weinberg principle states that allele and genotype frequencies in a population remain constant (are in equilibrium) from generation to generation. Deviations from HWE might, for example, indicate genotyping error, limited population size, population substructure, or newly occurred mutations. A P-value ≥ 0.05 from a chi-squared test used to test for HWE in the control population makes such errors/conditions unlikely (no deviation from HWE) and is considered a quality measure of a study. We investigated any migraine, MA, and MO.
We a priori decided to perform separate analyses among European and Asian populations since the genotype and allele frequencies differ between ethnic groups (21,22). We then pooled results from studies stratified by country of origin. We grouped the Turkish study (20) with the other studies in European populations based on the similarity of allele and genotype frequencies in the controls (Table 2).
We weighted the log of the ORs by the inverse of their variance to obtain pooled estimates. We ran random-effects models, which include assumptions about the variability between studies and provide more conservative estimates. We performed the DerSimonian and Laird Q test for heterogeneity and also calculated the I 2 statistic for each analysis (26). This statistic describes the percentage of total variation across studies that is due to heterogeneity rather than chance (25% low, 50% medium, 75% high heterogeneity). We used Galbraith plots to examine visually the impact of individual studies on the overall homogeneity test statistic. We evaluated potential publication bias visually by examining for possible skewness in funnel plots (27) and statistically with the methods described by Begg and Mazumdar (27) and Egger (28). The former method uses adjusted rank correlations, the latter a weighted regression approach to investigate the association between outcome effects (log odds ratio) and its standard error in each study. P-values < 0.05 may be regarded as an indication for publication bias.
We considered a two-tailed P-value <0.05 as statistically significant. All analyses were performed using SAS v.9.1 (SAS Institute Inc., Cary, NC, USA) and STATA v.10.1 (Stata, College Station, TX, USA).
Since we only utilised previously published data, we did not obtain approval of an ethics committee or written informed consent.
Results
Figure 1 summarises the process of identifying eligible studies. After title and abstract evaluation, we were left with 18 studies (10–20,29–35). We excluded eight more studies (19,29–35) after evaluating the full paper versions of the remaining articles and were left with 10 studies for this analysis (10,12–18,20,30).
Process of identifying studies.
Study characteristics
Characteristics of included studies investigating the association between the SLC6A4 5-HTTLPR polymorphism and migraine

MA, migraine with aura; MO, migraine without aura; NS, not specified.
Allele and genotype frequencies for the SLC6A4 5-HTTLPR polymorphism

MA, migraine with aura; MO, migraine without aura.
The allele and genotype frequencies for the SLC6A4 5-HTTLPR polymorphism for migraineurs and controls in each of the studies are summarised in Table 2.
Hardy–Weinberg Equilibrium and odds ratios (95% confidence intervals) for the association between the SLC6A4 5-HTTLPR polymorphism and migraine

MA, migraine with aura; MO, migraine without aura.
P-value for Hardy–Weinberg Equilibrium.
Association between the SLC6A4 5-HTTLPR polymorphisms and migraine, heterogeneity, and publication bias
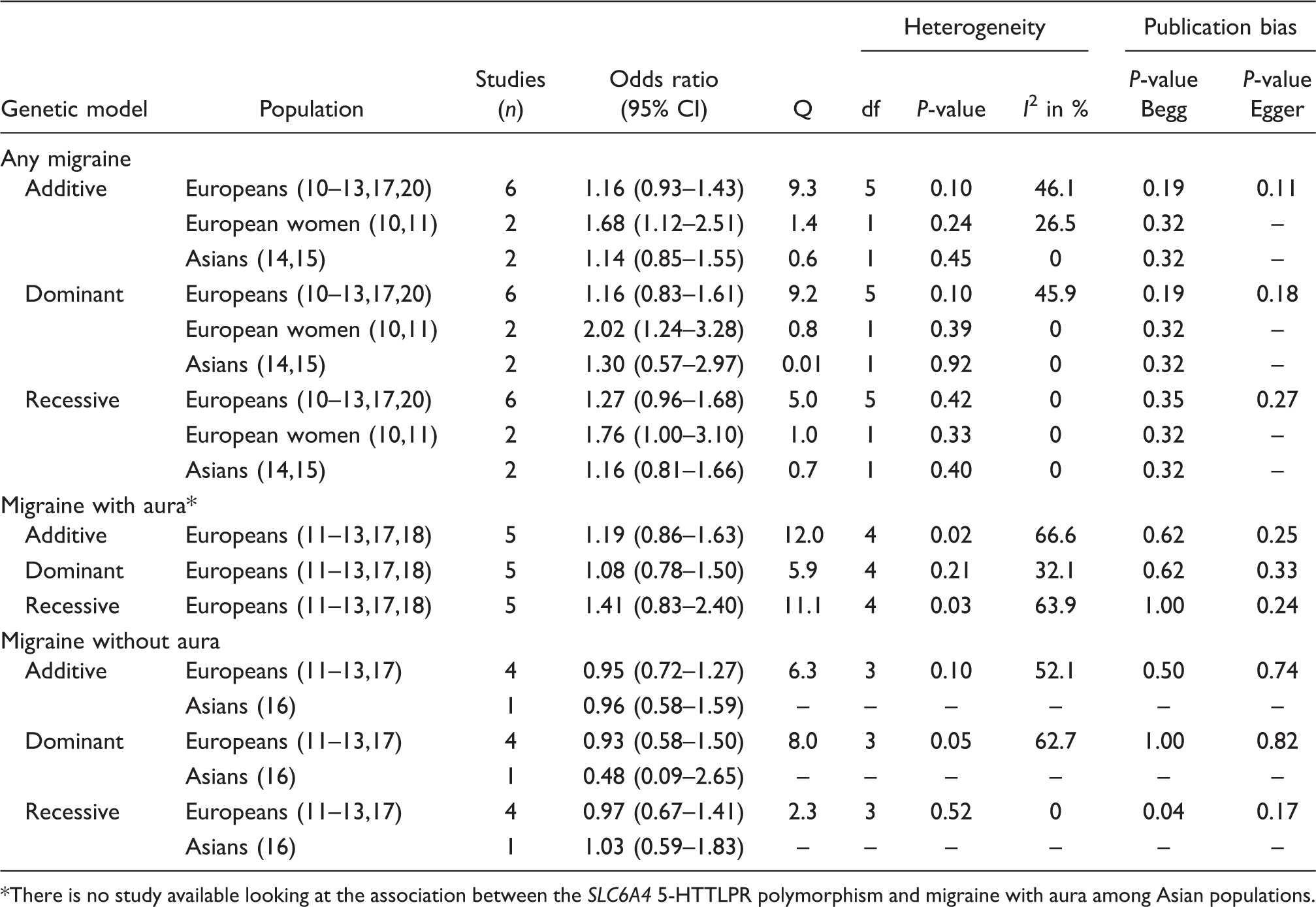
There is no study available looking at the association between the SLC6A4 5-HTTLPR polymorphism and migraine with aura among Asian populations.
Association between the SLC6A4 5-HTTLPR polymorphism and migraine
Four of the 10 studies suggested an increased risk for migraine among carriers of the S allele (Table 3) (10–13). In two studies among European women, this occurred for any migraine and appeared most pronounced under a dominant model (10,11), while in the other two among mixed populations this appeared only for MA and seemed to follow a recessive model (12,13).
Our overall pooled analyses do not suggest an association between the SLC6A4 5-HTTLPR polymorphism and any migraine (Table 4). The pooled OR (95% CI) from an additive model for any migraine was 1.16 (0.93–1.43) among Europeans (10–13,17,20) and 1.14 (0.85–1.55) among Asians (14,15). This did not change when assuming dominant or recessive models of inheritance. When we analysed the subgroup of European women, however, the risk for any migraine appeared to be significantly increased, which was most pronounced assuming a dominant model (dominant model: pooled OR = 2.02; 95% CI 1.24–3.28).
The studies looking at MA were all performed among European populations (11–13,17,18). The pooled effect estimates suggested an increased risk for carriers of the S/S genotype; however, this did not become statistically significant (recessive model: pooled OR = 1.41; 95% CI 0.83–2.40). There was medium heterogeneity among these studies (recessive mode: I 2 = 63.9%).
For MO, neither the studies among Europeans (11–13,17) nor the study in an Asian population (16) suggested an association with the SLC6A4 5-HTTLPR polymorphism.
Formal investigation using Begg’s test suggested some publication bias when assuming a recessive model among Europeans for MO (P = 0.04), however, Egger’s test did not. Neither test suggested publication bias for any of the other outcomes or for any of the other models.
Sensitivity analyses
When we repeated the analysis on the association between the SLC6A4 5-HTTLPR polymorphism and any migraine, excluding the study in the Turkish population (20), the results among the European populations were very similar to the overall results (additive model: pooled OR = 1.19; 95% CI 0.92–1.53).
For some of our analyses, Galbraith plots identified individual studies as important sources of heterogeneity. Hence, we performed sensitivity analyses by excluding studies that fell outside the margin set by the z score ± 2 SD.
For all analyses the effect estimates were lower and none indicated a statistically significant association. For example, for the association with any migraine the pooled OR (95% CI) from the additive model was 1.08 (0.91–1.27) after excluding one study (10). Likewise, for the association with MA among Europeans the pooled OR (95% CI) from the recessive model was 1.20 (0.70–2.07) after excluding one study (13).
Discussion
The overall results from this meta-analysis do not indicate an association between the SLC6A4 5-HTTLPR polymorphism and migraine among European and Asian populations. Studies among European women, however, indicate a 2-fold increased risk for any migraine for carriers of the S allele. The association with MA has only been investigated in European populations. While the pooled results suggested a 40% increased risk for MA among carriers of the S/S genotype compared to the other genotypes, these results were not statistically significant and there was remaining heterogeneity among studies.
Family and twin studies have clearly established a genetic component in migraine with a heritability of up to 50% (36). While mutations in individual genes have clearly been implicated in rare migraine forms such as familial hemiplegic migraine, it has been notoriously difficult to identify genetic markers for common forms of migraine (37).
The importance of serotonin in migraine has been acknowledged for decades (3) and a dysfunction in the serotonergic system agrees with both pathophysiological models of a cortical dysbalance (38) as well as treatment success of triptans, which are serotonin receptor agonists (39). Among the main features in migraineurs are alterations in serotonin metabolism and in the processing of central serotonin-mediated responses compared to non-migraineurs (3). Since the SLC6A4 5-HTTLPR polymorphism determines serotonin levels with carriers of the S allele having impaired transporter activities (9), it was hypothesised that this variant may also be a marker for migraine. Despite biological plausibility, however, individual studies on the association between the SLC6A4 5-HTTLPR polymorphism and migraine came to conflicting results and our meta-analysis does not support an association. Results from our overall analysis are in agreement with two family-based studies. One used linkage analysis and did not find significant linkage of markers in the SLC6A4 gene region with migraine (33). The other employed the transmission disequilibrium test and did not find an association of any of the 5-HTTLPR genotypes with overall migraine, MA, or MO (31). There are two possible explanations for the negative results. Either there may be truly no association and the positive studies may have occurred by chance or the pattern of association may be more complex and may depend on additional factors. Our results support the latter notion.
In this respect, the following limitations of our meta-analysis need to be considered. First, migraine is a complex, heterogeneous disorder with a wide clinical spectrum (37). Hence, patients with different clinical phenotypes are summarised as having migraine and even strict classification according to IHS criteria (24,25) may not sufficiently capture this variability and may be a source of misclassification in all studies. For example, the SLC6A4 5-HTTLPR polymorphism may not simply be associated with any migraine, but with attack frequency, as suggested by one study (15). Further, the association may be different for MA and MO. There is some indication for this from our pooled analyses, since the risk for MA may be increased by 40% for carriers of the S/S genotype. However, this was not statistically significant, possibly due to low sample size, and there was medium heterogeneity (I 2 = 63.9%) leaving residual uncertainties. Moreover, the SLC6A4 5-HTTLPR polymorphism may be associated with early age of migraine onset. In one of the studies, migraineurs were children and adolescents (17); however, this study did not find an association. Second, in addition to the 5-HTTLPR polymorphism, another common polymorphism in the SLC6A4 gene, a variable number tandem repeat in intron 2 (termed STin2 VNTR), has been investigated with regard to its association with migraine, the results being like-wise controversial (32,34). The structural organisation of the STin2 VNTR polymorphism is more complex than that of the 5-HTTLPR polymorphism, since it consists of three alleles and five genotypes. This does not easily allow employing genetic models (additive, dominant, recessive) based on one degree of freedom tests; hence, we did not to include it in our present analysis. Third, SLC6A4 is a large polymorphic gene comprising 37,810 bp organised in 15 exons and 14 introns with 380 single nucleotide polymorphisms (SNPs) reported (40,41), which in addition to the SLC6A4 5-HTTLPR and STin2 VNTR polymorphisms may affect function of the transporter. Fourth, the traditional concept of the SLC6A4 5-HTTLPR polymorphism as being bi-allelic, with the S allele being associated with a lower number of serotonin transporters compared with carriers of the L/L genotype (9) has been challenged. Recent data suggest the existence of a low-frequency LG-allele variant, harbouring an A > G SNP in its sequence, whose functional properties may be similar to the A allele (42,43). However, this additional genotypic information was not available for any of the studies included in our analyses; hence, we could not investigate an association with migraine. Fifth, serotonin’s role in migraine is not just determined by transporter activity, but also by serotonin’s functional effect at the receptor level, which is like-wise under genetic control and unaccounted for in all the studies. Sixth, although serotonin appears to be pivotal, other neurotransmitters like dopamine, orexin, and glutamate are also important in migraine pathophysiology (44). Hence, migraine susceptibility is likely a function of the interaction between various neurotransmitters, which again is determined by the interaction of various genetic variants. Seventh, migraine susceptibility is different between women and men, as indicated by the higher prevalence of migraine among women (45). Our results suggest that this may also be the case with regard to the SLC6A4 5-HTTLPR polymorphism, as European women carrying the S allele have a 2-fold increased risk for any migraine. However, only three studies (10,11,16) were conducted in women and all other studies only reported results for mixed gender populations. Eighth, associations with migraine may differ by ethnicity, especially since the genotype distribution of the SLC6A4 5-HTTLPR polymorphism shows ethnic differences (21,22). We have acknowledged this in our analysis by stratifying studies according to country of origin. However, studies are scarce among Asians and unavailable from other populations of non-European descent. Hence, in particular with regard to MA, a potential link with the SLC6A4 5-HTTLPR polymorphism remains unclear. Finally, we only used extractable data from the papers. However, only one paper did not provide genotypic information (19). The authors reported no association of the SLC6A4 5-HTTLPR polymorphism and migraine; hence, this would likely not have changed our results.
Additional research appears warranted to delineate further the association between the SLC6A4 5-HTTLPR polymorphism and migraine, in particular among women and in MA. In addition, studies in non-European populations are scarce and may yield different results. Future studies need to be adequately powered, should use standardised migraine classification including aura status, and should also present results stratified by gender and migraine aura status. Further, gene–gene interactions should be investigated, even if individual gene variants do not suggest an association with migraine. Finally, other than migraine status, age at onset or markers of migraine severity including attack frequency and aura frequency may be better outcomes.