
Abstract
Noninflammatory alopecia is common in dogs and is a frequent cause to consult a veterinarian. It is also a common reason to take biopsies. Noninflammatory alopecia can be attributed to a decreased formation or cytodifferentiation of the hair follicle or the hair shaft in utero, resulting in congenital alopecia. Congenital alopecia often has a hereditary cause, and examples of such disorders are ectodermal dysplasias associated with gene variants of the ectodysplasin A gene. Noninflammatory alopecia may also be caused by impaired postnatal regeneration of hair follicles or shafts. Such disorders may have a clear breed predilection, and alopecia starts early in life. A hereditary background is suspected in those cases but has not been proven. They are referred to as follicular dysplasia although some of these disorders present histologically like a hair cycle disturbance. Late-onset alopecia is usually acquired and may be associated with endocrinopathies. Other possible causes are impaired vascular perfusion or stress. As the hair follicle has limited possible responses to altered regulation, and histopathology may change during the course of a disease, a detailed clinical history, thorough clinical examination including blood work, appropriate biopsy site selection, and detailed histological findings need to be combined to achieve a final diagnosis. This review aims to provide an overview about the known noninflammatory alopecic disorders in dogs. As the pathogenesis of most disorders is unknown, some statements are based on comparative aspects or reflect the authors’ opinion.
Hypotrichosis or alopecia refers to abnormal thinning of hair or complete hair loss in body regions or on the entire body. Alopecia that is not caused by an internal disease with potentially serious consequences to general health is often regarded as a cosmetic problem. However, if the importance of the hair coat for thermal regulation, physical and immunological protection against external insults, dispersion of sebum and pheromones, sensory perception, social interactions, and camouflage are taken into account, alopecia is in any case affecting the physiological and psychological well-being of the affected individual. 124 Dogs with alopecia are more susceptible to sunburn, temperature extremes, comedone formation, as well as bacterial and fungal infections of the skin.
An intact hair coat requires faultless hair follicle (HF) morphogenesis during embryonic life and lifelong recurrent reconstitution of the HF. Both HF morphogenesis and the hair cycle (HC) are tightly regulated and depend on a fully functional stem cell compartment.25,117 As outlined in the review on HF structure, morphogenesis, and regeneration in the same issue of this journal, the hair coat, after initial follicular morphogenesis, is preserved by repetitive cycles of periodic stages of HF growth (anagen), regression (catagen), and quiescence (telogen). Hair shedding occurs during the exogen phase and follicles that lack a hair shaft (HS) are in the kenogen stage. The duration of the different HC phases varies between species, breeds, and body locations, and is influenced by age, sex, hormones, and day light. Thus, hair may be shorter or longer on specific body parts, it may have different textures on some body parts, or it may be, depending on the time of the year, thicker or thinner. Most dog breeds have an asynchronous HC with a mixture of anagen (average 40%), telogen (average 17%), and kenogen (average 14%) HFs, but some breeds (eg, poodles) have an anagen-dominated HC with more than 90% of the HFs being in anagen. 98 Due to the breed depend variation in hair fiber thickness and length, HF density, and seasonal changes, the clinical and histological assessment of mild hypotrichosis may be difficult and breed-specific aspects have to be taken into account when the hair coat is evaluated clinically or histologically.
Alopecia is a common cause for consulting a veterinarian, and a systemic work-up is mandatory to identify the correct underlying cause. Alopecia can be primary or secondary, and in primary cases, it is either noninflammatory or inflammatory. Noninflammatory alopecia is attributed to a decreased formation or cytodifferentiation of HFs or the HSs in utero or an impaired postnatal regeneration of HFs. The decreased formation of HFs or HSs during embryonic life results in congenital onset alopecia and is most often hereditary. In contrast, late-onset alopecia is usually acquired. Certain dog breeds are born with an intact hair coat but may develop noninflammatory alopecia early in life. In these cases, a hereditary cause is likely. However, a monogenetic trait has not been identified in any of these disorders. Inflammatory primary alopecia is caused by increased destruction of HFs and/or HSs. The cause can be either infectious or immune-mediated.49,50 In most cases, inflammation targets primarily the HF; however, the HF can also be affected by inflammatory cells that target other components of the skin (eg, sebaceous glands). Examples of inflammatory alopecia are folliculitis caused by infectious agents (bacteria, dermatophytes, or demodex mites), perifollicular inflammation caused by leishmania, or immune-mediated follicular or perifollicular inflammation (eg, alopecia areata, idiopathic mural folliculitis, perifolliculitis, and sebaceous adenitis).49,50 Alopecia as a secondary lesion develops most often subsequent to dermal inflammation and pruritus due to hypersensitivity reactions or ectoparasites.
This review aims to provide an overview about primary noninflammatory alopecia in dogs.
General Considerations and Definitions
The term “dysplasia” is defined as abnormal or disorganized growth or development of cells, tissues, or organs resulting in alterations in size, shape, or organization of body structures. 132 The term “follicular dysplasia” is used for genetically predisposed HF defects, which may be present at birth or develop at young age. With regard to alopecia, it is important to distinguish between (ectodermal) follicular dysplasia and ectodermal dysplasia. While in follicular dysplasia only HFs are affected, in ectodermal dysplasia 2 or more tissues of ectodermal origin are impaired. In human medical literature, ectodermal follicular dysplasia is referred to as nonsyndromic ectodermal dysplasia, whereas ectodermal dysplasia in general is reported as syndromic. 55
Follicular dysplasia can manifest at birth as congenital alopecia. Follicular dysplasias that manifest at birth have been reported in cattle and are known in hairless cat breeds.44,58 In dogs, to my knowledge, no congenital follicular dysplasia has been reported, and all congenital alopecic disorders reported in the literature are associated with dental abnormalities and thus are categorized as ectodermal dysplasia. Sole follicular dysplasia in dogs is seen with postnatal onset, and clinical signs appear within the first few years of life.85,149 Canine follicular dysplasia that develops after birth is associated with abnormal reconstitution of the inferior portion of the HF or the HS and may present histologically as a cycling defect.12,24,31,39,48,78,84,99,128,140 In the later cases, the histology is often comparable to acquired HC disturbances, and only the knowledge about the breed affected, the age of onset, and the exclusion of endocrinopathies allows for a definite diagnosis. The fact that hormonal imbalances have been identified in HF dysplasias that develop postnatally suggests a genetic predisposition and explains their histological similarity to acquired HC disorders caused by endocrinopathies.13,24,41,122 Beside hormones, dysregulation of signaling of molecules derived from ectodermal or mesodermal tissue or a defective stem cell compartment may also affect the cycling capacity of the HF. If the dysplasia results in structural defects of the HS, histological evaluation may fail to see these defects and analysis of the HSs by trichograms or electron microscopy may be required. 14 Follicular dysplasia may also be associated with an impaired function of tissue derived from the neuroectoderm such as in color dilution alopecia. 34
The term “ectodermal dysplasia” in general refers to clinically and genetically heterogeneous inherited conditions in which the development of 2 or more tissues of ectodermal origin is impaired.29,111 Thus, affected animals may present with congenital hereditary alopecia/hypotrichosis and defects in other body locations, including brachygnathism, dental, thymic, or genital abnormalities.111,112,153
Ectodermal dysplasia, which is associated with congenital alopecia, is mostly caused by variants of the ectodysplasin A (EDA) gene. EDA is a member of the tumor necrosis factor (TNF) family and is involved in ectodermal/mesodermal interaction. This interaction is essential for the formation of several structures that arise from the ectoderm, including the skin, sweat glands, nails, and teeth. 30 During HF morphogenesis, EDA signaling is important for early placode formation. 88 Binding of EDA-A1 to its receptor (EDAR) results in the recruitment of the intracellular EDAR-associated death domain (EDARADD) adapter protein and the activation of the NF-κB signaling pathway, while EDA-A2 binds to the EDA2R, also known as X-linked ectodermal dysplasia receptor (XEDAR). 73 EDARADD can contribute to hypohidrotic ectodermal dysplasia (HED) in mice, human beings, and bovine.32,37,45,61,101,102,125,133 The clinical phenotype depends on the gene variant. Some variants lead to a complete loss of the protein function and a severe clinical picture, whereas other variants are associated with residual EDA activity and milder phenotypes.15,46
Hair Cycle Disturbance/Arrest
As outlined in detail in the review on HF structure, morphogenesis, and regeneration in this journal issue, the maintenance of the HC and thus the reconstitution of the inferior portion of the HF and the HS are highly conserved processes and are tightly regulated. It involves follicular stem cells as well as numerous molecules derived from epithelial, mesenchymal, and neuroectodermal cells as well as the extracellular matrix of the follicular and dermal environment and is influenced by systemic factors such as hormones, age, genetics, and environmental factors, such as the time of the year. 25
If 1 or several extrinsic or intrinsic factors of this complex regulatory network are deregulated, the delicately balanced crosstalk of the various signals that influence the HC is disturbed and results in deregulation of the HC. It is important, although not known in the dog, to consider that the different primary and secondary HF types may be regulated differently, and thus, a HC disturbance may primarily affect primary or secondary HFs. The distinct regulation of primary and secondary HF morphogenesis and cycling has been shown in mice and Cashmere goat.35,155 Depending on the factors that are deregulated and in which stage the different HFs are in at the onset of the deregulation, different HC stages or HF types may be affected. As a consequence, either anagen induction, anagen promotion or cytodifferentiation, catagen induction, catagen promotion, telogen induction or telogen promotion or several of these stages are impaired and the HC disturbance may affect all or only some HF types. 86 As each HC phase has an impact on the consecutive phase, an ongoing HC disturbance will result histologically in a similar clinical phenotype. For example, failure of anagen induction results in a higher number of telogen follicles, which will eventually lose their HSs (kenogen follicles). If lack of anagen induction remains kenogen follicles will become atrophic over time. In a study histologically investigating some of the HC arrest disorders (endocrinopathies, alopecia X, alopecia of unknown origin), we could show that in all of these diseases 3-fold to 4-fold increases of kenogen follicles were seen histologically, whereas the number of anagen follicles was decreased dramatically. The kenogen follicles underwent subsequent atrophy. 98 The abovementioned increased amount of kenogen follicles in alopecia is also observed on the human scalp.114,115 It is important to understand that only an increased number of kenogen and atrophic follicles result in the clinical picture of alopecia (Fig. 1a, b). Telogen follicles still have a club hair, and thus, a higher number of telogen follicles do not result in alopecia. Of course during the disease course, it may happen that the number of telogen follicles is transiently increased. It also has to be noted that up to 20% of kenogen HFs may be normal in many dog breeds. They serve as reserve follicles to grow a thicker hair coat, if needed. In contrast, in anagen-dominated dog breeds, and on the human scalp, kenogen HFs are only a small percentage of the HFs present,98,115 and in anagen-dominated breeds, some anagen follicles may be present in biopsies from dogs with a HC disorders. Besides the decreased amount of anagen follicles and the increased number of kenogen and atrophic follicles additional histological findings in HC arrest disorders may be (1) increased infundibular keratin and dilation of the infundibula (Fig. 1b), (2) excessive trichilemmal cornification (Fig. 1a), (3) distorted (dysplastic) follicles (see Fig. 7a), (4) transiently increased numbers of telogen follicles (Fig. 1a), (5) transiently increased numbers of catagen follicles, (6) epidermal atrophy (Fig. 1b), (7) epidermal hyperplasia (if secondary infections are present), (8) epidermal hyperkeratosis (Fig. 1a), (9) dermal atrophy, and (10) epidermal hyperpigmentation (Fig. 1b).
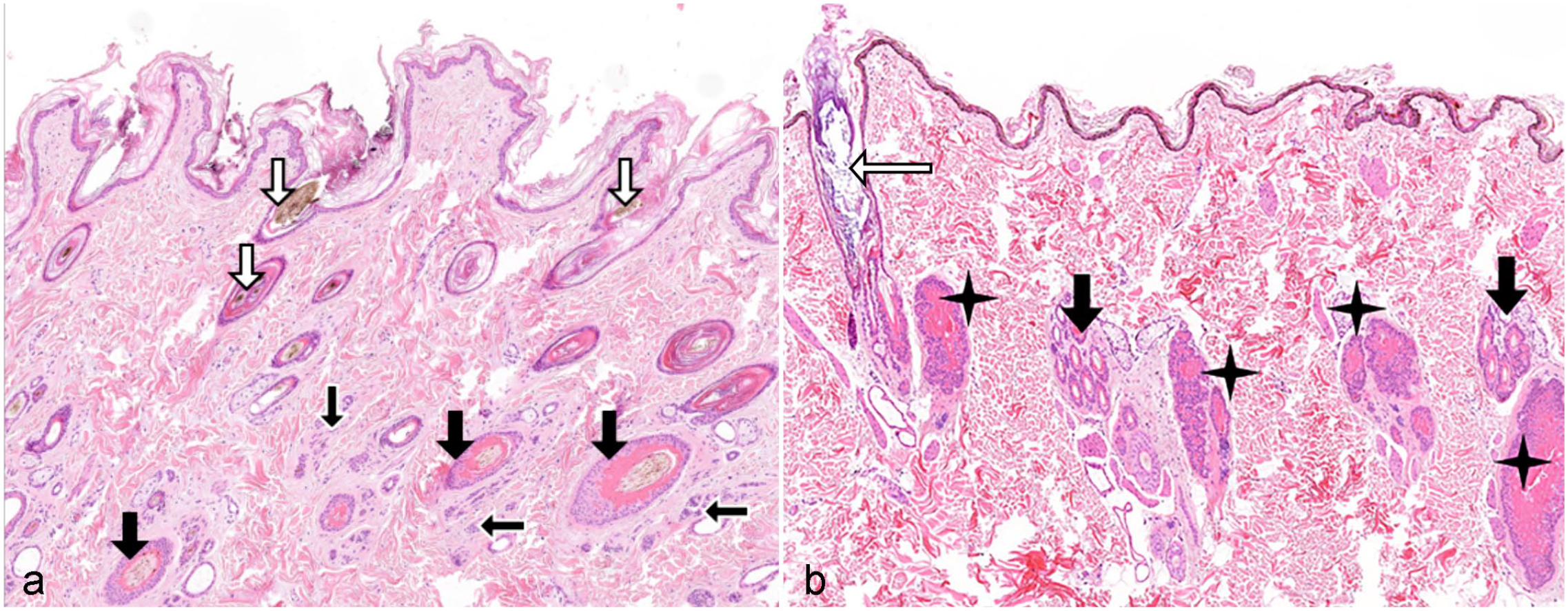
Canine skin showing typical features of a hair cycle disturbance. Hematoxylin and eosin.
The increased percentage of kenogen follicles may be due to different reasons:
Signals that induce a new anagen phase at the end of the telogen phase are missing/reduced. The club hair will be shed eventually during exogen, and the empty follicles remain (hairless telogen/kenogen) and become atrophic over time.
Exogen has occurred prematurely, and the empty telogen follicle is not yet competent to enter a new anagen phase.
Signals which are maintaining the anagen phase (anagen promotion) are missing; the HFs enter catagen prematurely and thus enter the subsequent telogen phase earlier. At the same time, the signals which are necessary for anagen initiation are not more effective. This results in a higher number of telogen follicles, which eventually lose the HS and remain in kenogen until adequate signals initiate a new anagen phase.
The lack of signals to initiate the next HC phase may also be the reason for a transiently increased percentage of catagen or telogen follicles.
The impact of hormones, associated with canine alopecia on the different HC phases, is shown in Figure 2. This figure shows that the same hormone has an effect on various HC phases and underlines why histology may look alike in all HC arrest disorders and the histologic features are dependent on the disease duration and the HC phase at the time of disease onset.
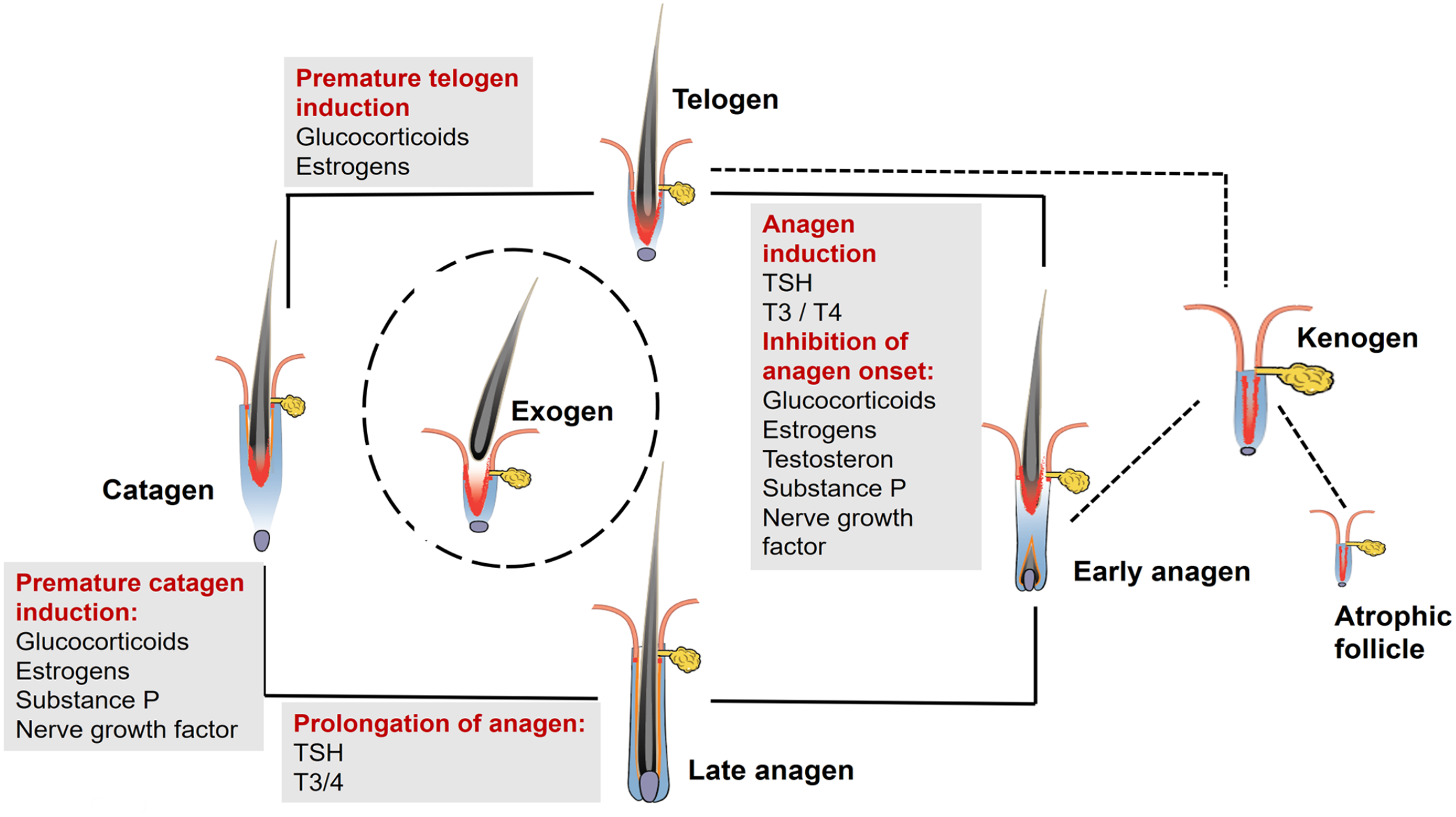
The impact of glucocorticoids, estrogens, thyroid hormones, and stress-related molecules on the hair cycle. The same hormone or molecule may have an effect on various hair cycle phases, which explains that endrocinopathies may histologically look alike and also that other factors such as stress might result in a hair cycle disturbance. Note that thyroid-stimulating hormone (TSH), triiodothyronine (T3), and thyroxine (T3/4) have an stimulating effect on the hair cycle, whereas estrogens and glucocorticoids have an inhibitory effect.
Alopecia With Congenital Onset
Congenital alopecia refers to the widespread to complete absence of hair at birth. It is seen regularly by breeders and in veterinary clinics but only a limited number of cases have been reported in the literature, and even a smaller number have been worked up genetically. Novel techniques, such as whole genome sequencing, facilitate the discovery of the causative gene variants, and the number of cases with a known underlying gene variant is increasing. Congenital alopecia is not always inherited, but may also be caused by a metabolic imbalance or an in utero infection. However, noninherited cases, such as maternal dietary iodine deficiency or intrauterine virus infections, have been reported in farm animals only.62,121
Congenital alopecias in dogs follow monogenetic inheritance patterns and are inherited through autosomal recessive, autosomal dominant, or X-linked modes.33,54,57,92,106 De novo mutations have been seen as well. 85 Depending on the gene affected, HF morphogenesis can be disturbed during induction, organogenesis, or cytodifferentiation.
Accordingly, histology will reveal complete absence of HFs (aplasia caused by impaired HF induction), incomplete HF formation (dysplasia caused by impaired organogenesis), or inner root sheath and/or HS defects (dysplasia caused by impaired cytodifferentiation). If cytodifferentiation of the HS is impaired, HFs may look histologically normal or only the inner root sheath of the inferior portion may present with histological changes such as vacuoles or altered trichohyalin granules. The latter has not been reported in dogs but has in certain cat breeds and cattle.44,58 If only HS defects are present, the histological evaluation might fail to make the diagnosis, but trichograms or electron microscopy of HSs may be helpful to identify structural HS defects, which result in hair breakage.
Follicular Aplasia
Aplasia of HFs is seen when early placode formation during organogenesis does not occur. The mechanisms that regulate placode formation are complex and insufficiently understood. They have only been studied in detail in rodents.35,79 As outlined above, the EDA/EDAR signaling pathway plays an important role in placode formation. 88
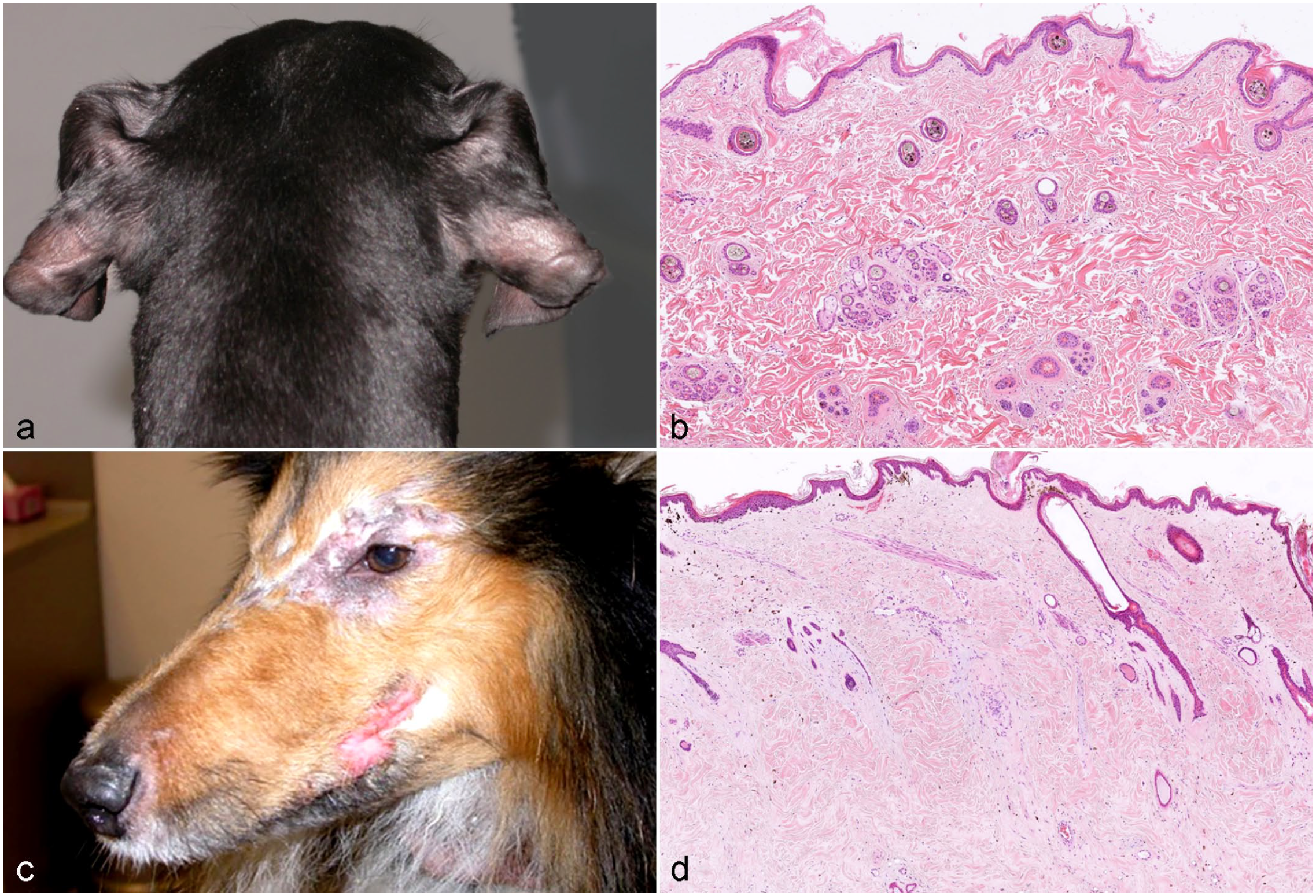
Canine ectodermal dysplasia resulting in follicular aplasia has been reported as congenital ectodermal defect, congenital ectodermal dysplasia, and X-linked ectodermal dysplasia.16,76,92,129 Dogs with ectodermal dysplasia may lack, like other species with the same disease, other structures of ectodermal origin such as eccrine sweat glands, sebaceous glands, lacrimal glands, or nasolabial glands (Fig. 3a).
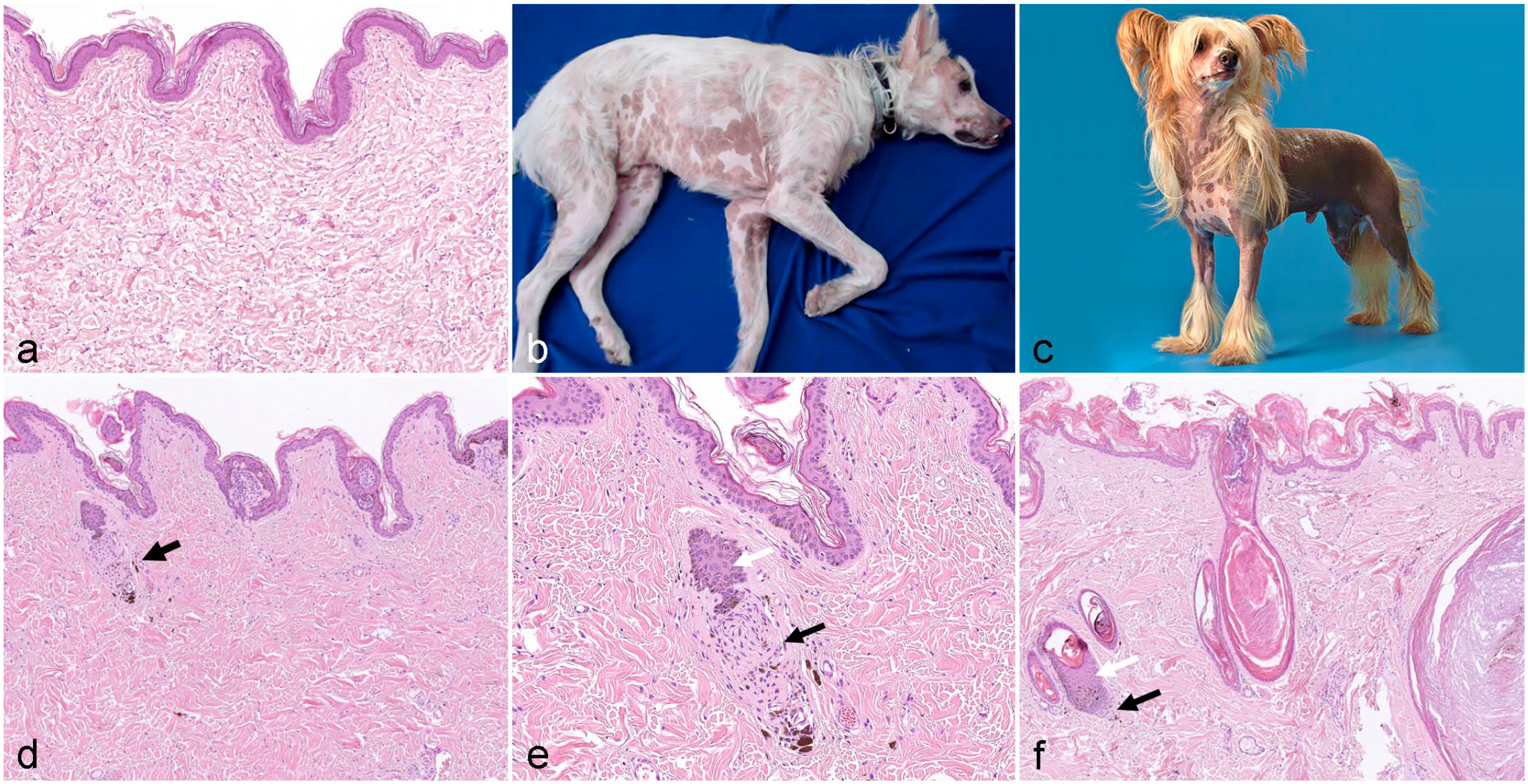
Clinical and histological images from dogs with congenital follicular dysplasia.
The clinical presentations of canine cases of various breeds have been nicely summarized in reviews and several case reports.17,54,92,94,95,129,141,146 The affected animals are completely alopecic or have a reduced number of hairs in various anatomic locations (especially the head, pinnae, neck, back, and tail) (Fig. 3b). Eyelashes and vibrissae may be lacking. The remaining hair has an abnormal texture, and is either coarse or fine, breaks easily, and can be epilated without effort. In some cases, the alopecia and hypotrichosis are most severe in newborn animals, and some fine hairs grow over time.
Several variants in the EDA gene encoding for EDA have been identified causing X-linked HED, the most common form of ectodermal dysplasia in dogs.18,54,63,141,146 In a poodle, however, which presented with clinical signs of X-linked HED, no variants in the EDA gene were identified, suggesting that other genes may also cause this disease in dogs. 93
Aplasia of HFs without dental dysplasia have, to the best of our knowledge, not been reported in the literature in dogs, although they most likely exist. 84 There are reports from swine in which the number of HFs is significantly reduced but the apocrine sweat glands and the teeth are not affected. 84
Congenital Alopecia Caused by Impaired Hair Follicle Organogenesis
Congenital alopecia/hypotrichosis also occurs when the causative gene variants affect HF organogenesis. In these cases, HF placodes were formed, but the morphogenesis of HFs is altered during the different stages of organogenesis. Histologic lesions in these cases vary depending on the underlying gene variant. In any case, HFs are not fully developed, are too short, and are malformed. The infundibulum and the dermal papilla (DP) may or may not be present. If HSs are produced, they are very thin and break easily. There are several dog breeds where a spontaneous mutation has been propagated on purpose and the phenotype is now recognized as breed standard (eg, Chinese crested dog, Mexican hairless dog, and Peruvian Inca Orchid) (Fig. 3c–f). Hairlessness in these 3 breeds is inherited as a monogenic autosomal semidominant trait. Hairless dogs are always heterozygous. 33 Whereas the Mexican hairless dog is truly hairless, some HFs develop in Chinese crested dogs. There are 3 phenotypes in the Chinese crested dogs: the true hairless, the semicoated, and the powderpuff (haired phenotype). The HF density in the hairless phenotype is much lower than in the 2 haired phenotypes and if HFs are present they are simple. 151 As these dog breeds also have dental abnormalities, the phenotype is considered as ectodermal dysplasia. A frameshift mutation the forkhead box I3 (FOXI3) gene has been identified as the cause of the hairless phenotype in the Chinese crested dog, the Mexican hairless dog, and the Peruvian Inca Orchid dog. 33 The hairless phenotype in these dog breeds can be explained by the biological function of FOXI3. In mice, it has been shown that the expression of Foxi3 is regulated by ectodysplasin in HF placodes and that mutations in Foxi3 result in the downregulation of several stem cell signature genes.130,131 It has also been shown that FOXI3 in mice is essential for molar crown patterning, which most likely explains the dental phenotype in these dog breeds. 59
In dogs, no reports are available in the literature that describe failure of HF organogenesis in dogs other than the above-described breeds. However, there are reports of failure of HF organogenesis in other domestic animal species.1,7
Dysplasia Associated With Insufficient Hair Shaft Quality Due to Impaired Cytodifferentiation
Despite the fact that coat abnormalities are common in veterinary medicine, only 7 inherited HS defects have been reported so far in domestic animals.44,58,69,84,118 Some of these defects result in alopecia present already at birth, whereas others most likely develop during cytodifferentiation of the cycling anagen HF. In dogs, trichorrhexis nodosa, trichoptilosis, medullary trichomalacia, spiculosis, and bald thigh syndrome in sight hounds have been reported. In trichorrhexis nodosa, nodes are formed along the HS and the hair is abnormally fragile following trivial injury. This disorder can be acquired or congenital and has been described in golden retrievers.84,128 Congenital trichoptilosis has been reported in related male golden retrievers.84,128 Medullary trichomalacia has been reported in German shepherd dogs, but it is unknown if this disorder is acquired or congenital. In medullary trichomalacia, the medulla of the HS is vacuolated resulting in hyperfragility of the HS. 128 Bald thigh syndrome is seen in greyhounds and related sighthound breeds (Fig. 4a). Because of the clear breed predisposition, a genetic component is most likely involved. In the literature, this syndrome has also been reported as pattern baldness and as follicular dysplasia;47,126 however, in a recent study, an impaired cytodifferentiation resulting in HS fractures has been identified as an underlying cause. On the molecular level, the phenotype is associated with downregulation of genes and the corresponding proteins in the HS (eg, keratin 1 and desmocollin), which are important for the proper assembly of the HS (Fig. 4b, c). 14 Recently, a canine case with bulbous swellings of the HSs, which resembled lanceolate hair phenotype seen in mice and 2 cats, has been observed. Genetic analysis determined a desmoglein 4 variant similar to mice and cats with the same phenotype( Sarah Kiener and Tosso Leeb, personal communication).
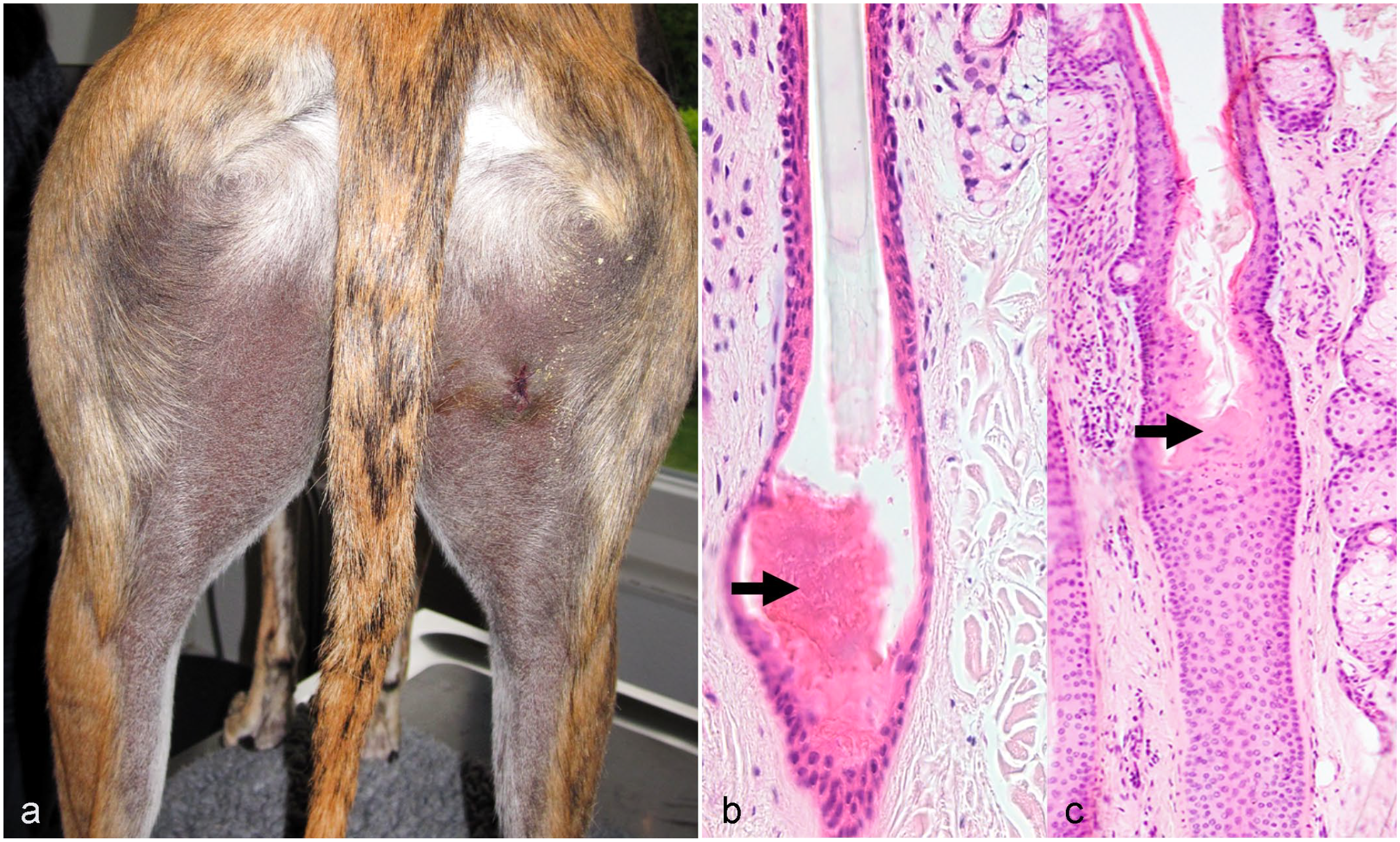
Greyhound with bald thigh syndrome.
Inherited Alopecia With Early Postnatal Clinical Manifestation
Hairlessness in the American Terrier (Hairless Rat Terrier)
Hairlessness is the desired phenotype of the American hairless terrier and is inherited as an autosomal recessive trait. Puppies are born with a sparse, fuzzy coat over their entire body and then gradually lose their coat from the nose backward until they are entirely bald. This typically occurs by 6 weeks of age. Eyebrows and whiskers are retained. Teeth may be missing but most dogs have normal dentition. The dogs are otherwise healthy. Histological features have not been described. The condition is associated with a deletion in the serum/glucocorticoid regulated kinase family member 3 gene (SGK3). 106 SGK3 has been shown to be important in postnatal HF development in mice.3,104
Juvenile Alopecia in the Scottish Deerhound
In contrast to the American hairless terrier, hairlessness in the Scottish deerhounds is undesired. It is caused by a frameshift insertion in the SGK3 gene and is inherited as a recessive trait. 57 Affected puppies are born with sparse hair and lose it within the first 2 months of life. The hair does not regrow. Histology has not been described, and dogs are otherwise healthy.
Noninflammatory Alopecia With a Clear Breed Predisposition, Postnatal Onset, and a Most Likely Hereditary Cause
There are several noninflammatory alopecic conditions with a clear breed predisposition and development of the alopecia during the first years of life, which indicates an impaired postnatal reconstitution of the HF. Due to the clear breed disposition, an underlying hereditary cause is likely, although no causative genes have been identified in the disorders that have been genetically evaluated.14,60,81,82,99,134,148 Thus, it may be speculated that a polygenic trait and other nongenetic factors, such as diet and an altered hormone metabolism, are involved in the pathogenesis.20,31,60,99 Histological features in most of those conditions are insufficiently described. If dogs were biopsied, only a small number of affected dogs have been investigated and different pathologists were involved.
Known diseases associated with postnatal follicular dysplasia are described.
Hypotrichosis and Alopecia in the Irish Water Spaniel
Hypotrichosis and alopecia in the Irish water spaniel is characterized by nonpruritic, noninflammatory, regionalized hair loss affecting the ventral and latero-dorsal neck, the flanks, the dorsum, the rump, the caudal part of the thighs, and the distal tail. The coat color changes from reddish brown to grayish brown. Pedigree analysis suggests a dominant mode of inheritance. A similar clinical presentation is also seen in the American water spaniel, although it has not been described in the literature. Hormone testing suggests an abnormality of steroidogenesis, and the hair coat regrew in some dogs after dietary changes. 20 Histologically, the HFs’ infundibula are dilated and filled with abundant keratin in the alopecic areas. The proximal HFs are atrophic. Melanin aggregates are present in the HSs, the follicular outer root sheath, and around the base of the follicles. These changes are also seen in the haired areas.20,22 As no control dogs were examined in this histologic study, it cannot be determined whether the pigment aggregates are normal for this breed.
Follicular Dysplasia of the Portuguese and Spanish Water Dog
This type of follicular dysplasia has only been described in the Portuguese water dog, but is seen also in the Spanish water dog, which is a related breed. The clinical and histological findings are similar in both breeds and resemble those of Irish and American water spaniels (Fig. 5a–d). The coat color may change from black to reddish brown. Hair loss occurs at a median age of 2 years. Both males and females are affected. Histologically, the HF infundibula are severely dilated and plugged with keratin. HFs may have an irregular outer contour and are either in telogen or in anagen. Anagen follicles may be small. Hairs may be absent from HFs or may appear fragmented and thin. Hair shaft dysplasia starts in the lower part of the inferior portion of the HF (Fig. 5b–d). There may be scattered clumps of melanin within follicular keratin and within follicular epithelium. Vacuolar changes in the inner and outer root sheath and the matrix cells have been reported.48,89
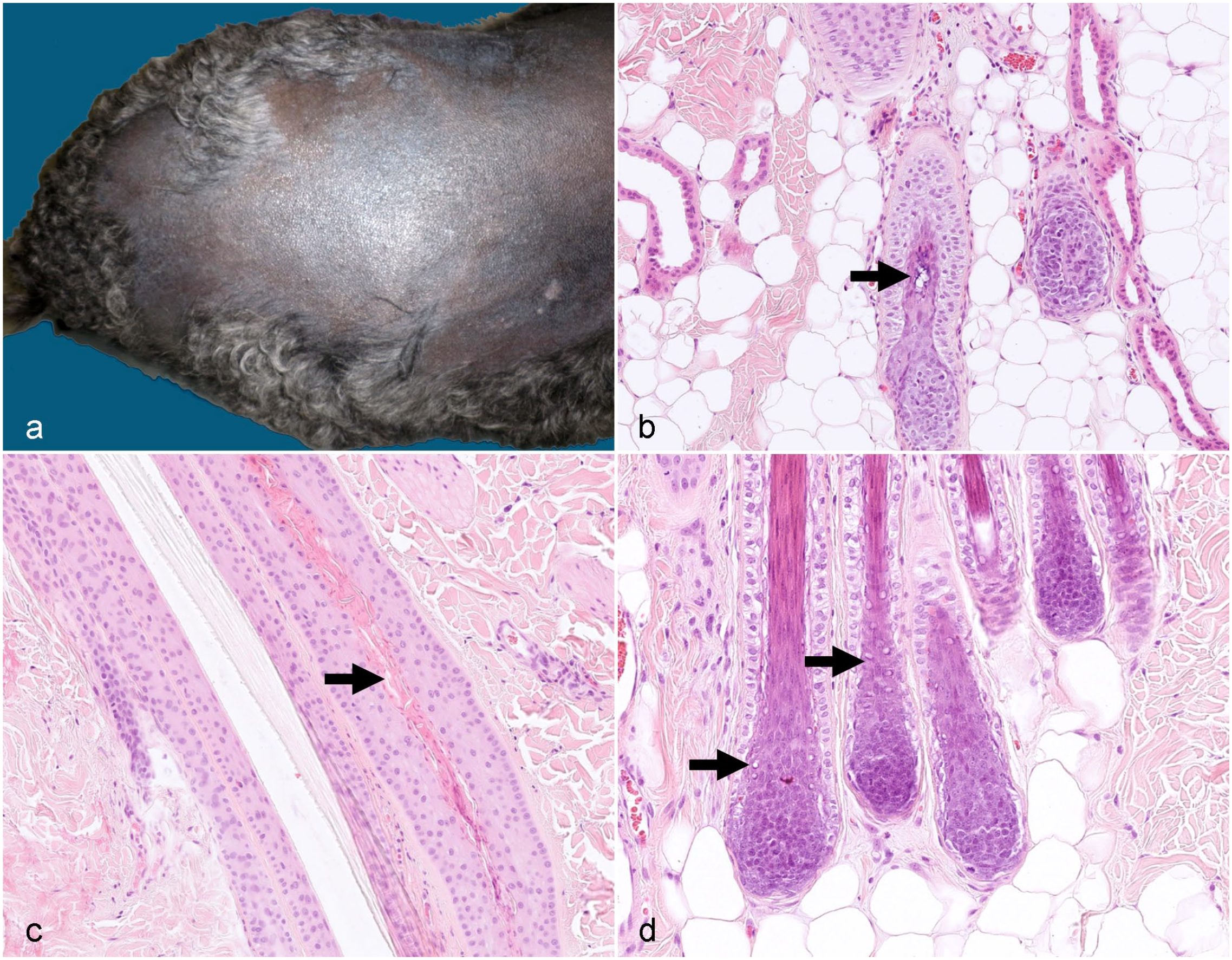
Follicular dysplasia in Spanish and Portuguese water dogs.
Follicular Dysplasia of the Curly Coated Retriever
Follicular dysplasia of the curly coated retriever has been described as symmetrical, nonpruritic alopecia and/or frizzy coat changes, usually affecting the caudal thighs, axillae, dorsum, and the neck (Fig. 6a). The age of onset varies from 4 months to 6 years (median 13 months). Alopecia may be waxing and waning. Histology is characterized by a reduced number of anagen follicles and increased numbers of kenogen and atrophic follicles. Infundibula are dilated and filled with abundant keratin. Melanin aggregates are present in the HSs and the outer root sheath; however, this may be normal in brown or black dogs (Fig. 6b, c). 12
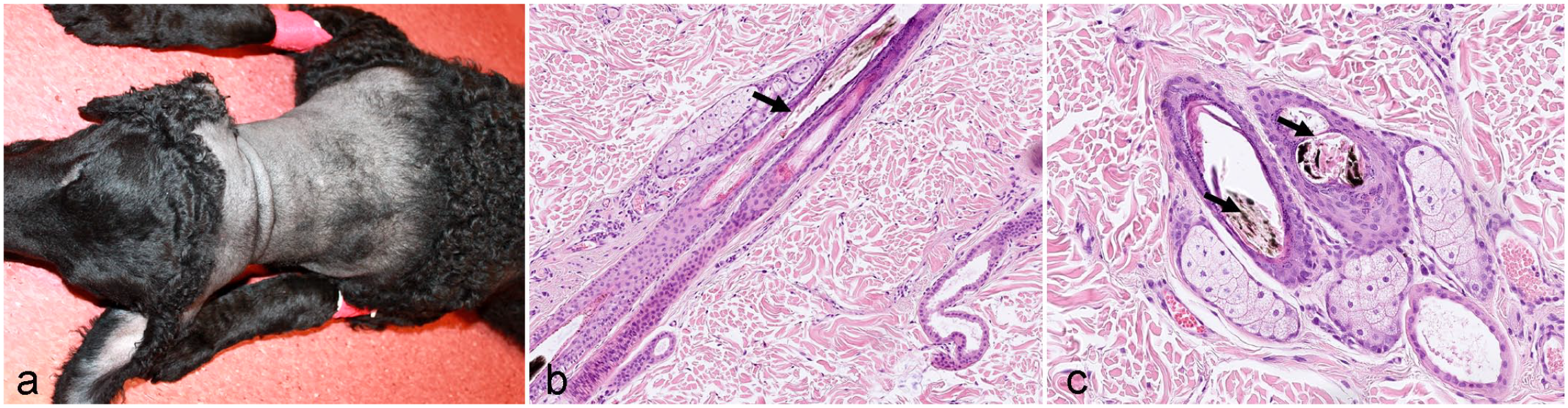
Follicular dysplasia with postnatal onset.
Alopecia of the Chesapeake Bay Retriever
Alopecia of the Chesapeake Bay retriever is characterized by hair loss affecting the axillae, the latero-ventral thorax, the flanks, the ventrum, the dorsum, the rump, and/or the caudal part of the thighs. Hormonal investigations showed increased adrenal and sex steroid concentration in some cases. Histopathology is characterized by severe infundibular hyperkeratosis, follicular atrophy, and occasional melanin clumping with malformed, often thin HSs. 24
Alopecia in the Pont Audemer Spaniel
The Pont Audemer spaniel is derived from the Irish water spaniel. Alopecia in the Pont Audemer spaniel is limited to the brown-haired areas of the trunk and ears, and develops within the first 2 years of life. Histological findings include severely dilated HF infundibula with hyperkeratosis. No information is available about the HC stages in affected dogs. However, in 1 study, vacuolar degeneration and apoptosis of keratinocytes of the inner and outer root sheath, suggesting that anagen follicles are present, have been reported. Melanin aggregates in the HSs and outer root sheath have also been reported. 53 However, this may be associated with the coat color of the affected areas.
Recurrent Flank Alopecia
Recurrent flank alopecia is characterized by recurrent episodes of well-demarcated alopecia that affects several canine breeds, but boxers, Rhodesian ridgebacks, Airedale terriers, French bulldogs, English bulldogs, and schnauzers are predisposed.89,140 In these dog breeds, recurrent flank alopecia is relatively common. It has been described also as seasonal flank alopecia, seasonal growth hormone deficiency, canine idiopathic cyclic flank alopecia, cyclic follicular dysplasia, and follicular dysplasia. 31
The existence of a strong breed predilections implies a hereditary component; however, the photoperiod and seasonal climactic variations influence the onset of alopecia and hair regrowth. The response to melatonin in many cases underlines this association.77,120 Nevertheless, there is evidence that systemic intrinsic factors play a role in the pathogenesis since lesional skin grafts of dogs with recurrent flank alopecia on the back of athymic mice resulted in hair regrowth faster than in the donor dogs. 140
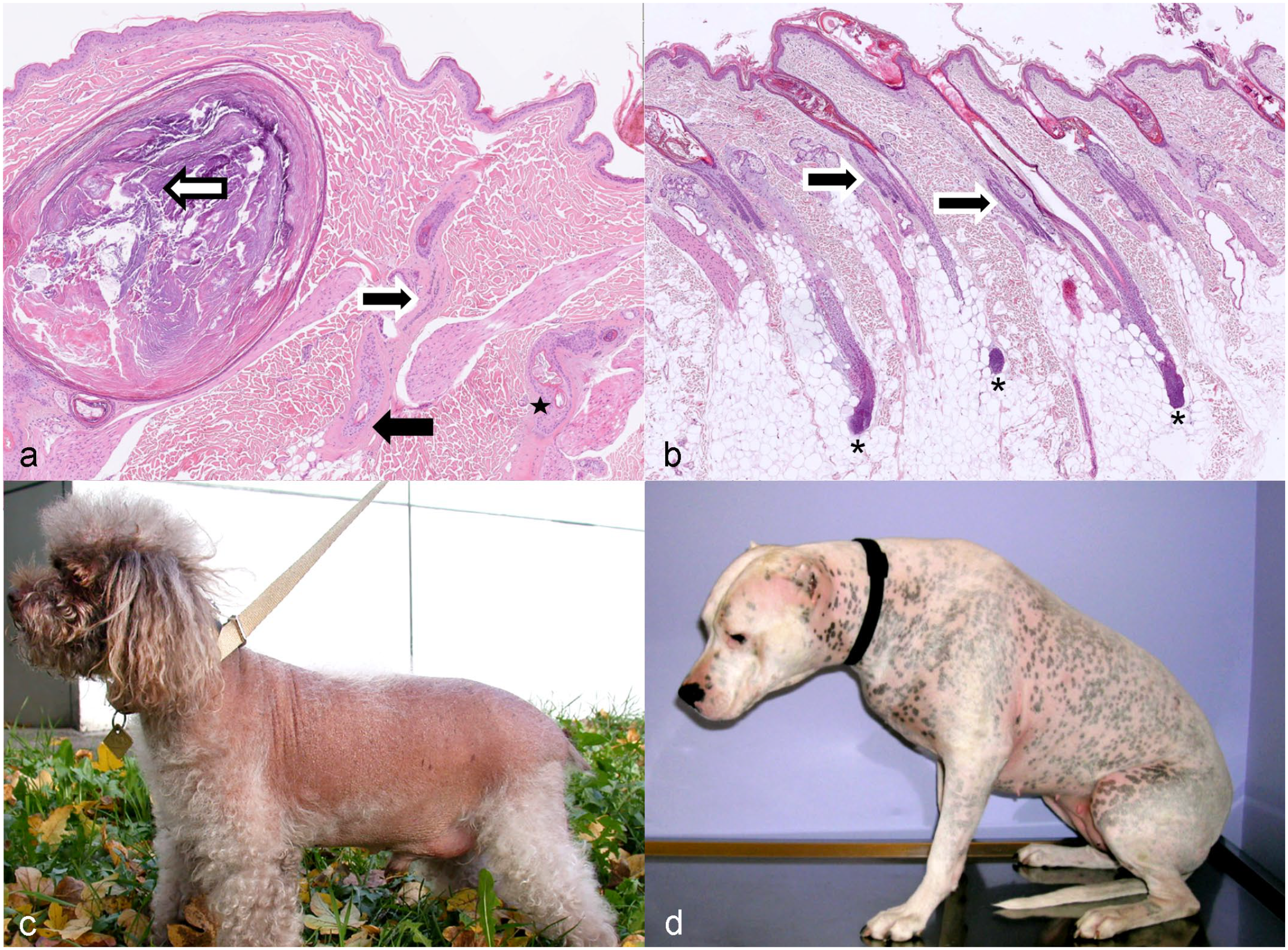
Clinically, the lesions are distinctive and are characterized by recurrent episodes of sharply demarcated alopecic areas bilaterally in the flank region. The dorsal midline and the thorax may be involved as well. The alopecic skin is hyperpigmented. Hair may regrow partially or complete before it is lost again. Regrown hair may have an altered texture and can be darker. Histologically, the HF infundibula are severely dilated, elongated, and plugged with orthokeratotic follicular keratin that extends into the openings of all follicles assembled in 1 follicular compound. The lower follicular segments of the compounds are short, atrophic, and may have a distorted shape. The term “Witches foot” or “Warlocks foot” is used to describe the dilated infundibulum overlying this group of toe-like, shortened, and irregular primary and secondary follicles (Fig. 7a). 149
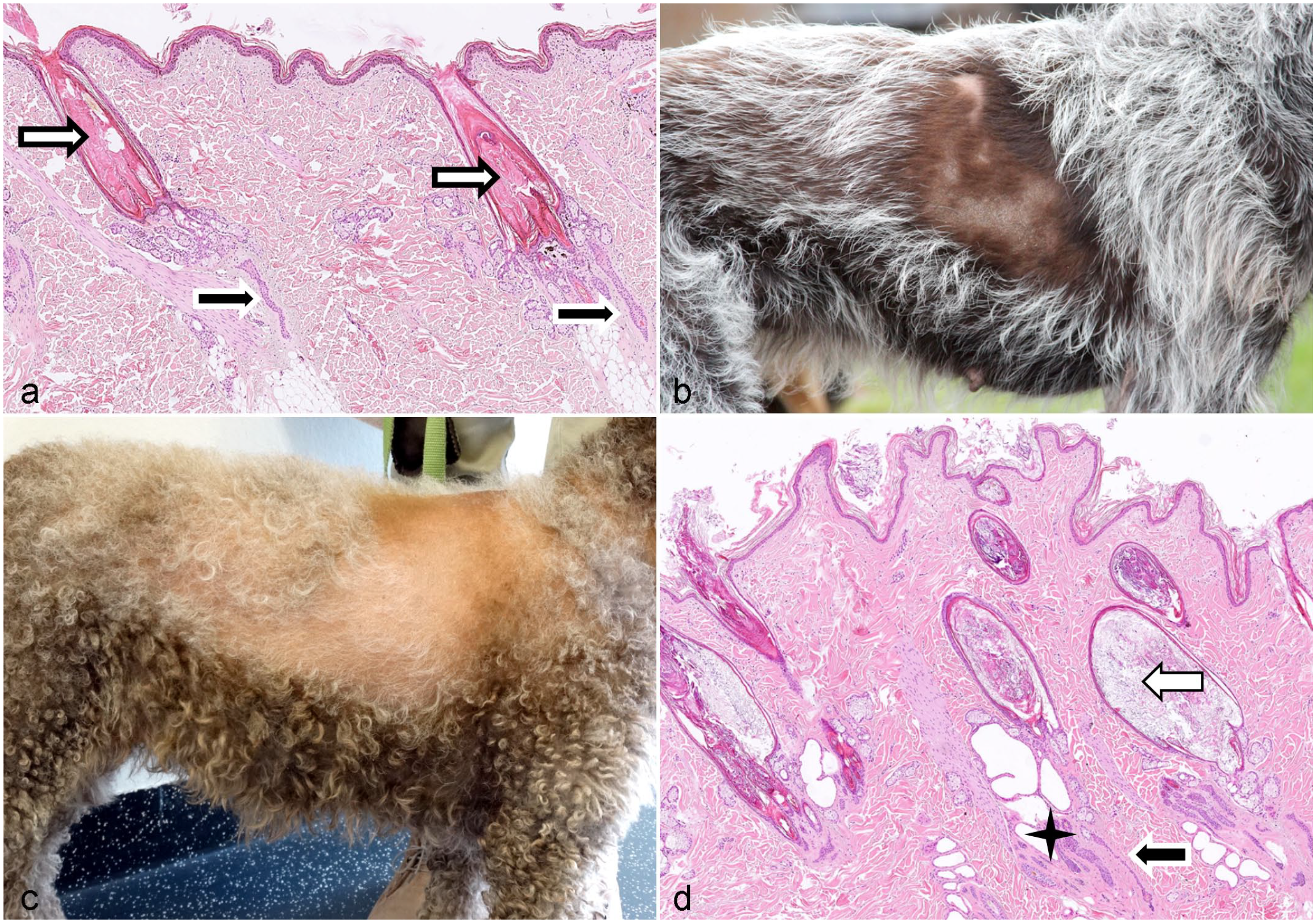
Follicular dysplasia with postnatal onset.
Atypical Recurrent Flank Alopecia
Atypical recurrent alopecia is seen in several hunting dog breeds such as the German short hair pointer and the Cesky Fousek (Fig. 7b). 99 Recurrent alopecia affecting the face only has been described in the Cane Corso and in several Dogues de Bordeaux. 139 Similar to typical recurrent flank alopecia, the existence of a strong breed predilections implies a hereditary component. Atypical recurrent flank alopecia is more severe than typical recurrent flank alopecia and may become permanent with time. In contrast to the typical recurrent flank alopecia, the skin in the alopecic areas is not hyperpigmented. Histologic findings in cases of atypical recurrent flank alopecia are comparable to those observed in typical recurrent flank alopecia. 99
Alopecia in the Lagotto Romagolo
Alopecia in the Lagotto Romagolo is characterized by bilateral hair loss on the trunk and usually starts in autumn/winter (Fig. 7c). This disease has been seen for a few years and started in the Swedish Lagotto population. It has been reported also as follicular dysplasia. 136
Seasonal cycling is observed in about half of the affected dogs. Alopecia onset or worsening of the condition is associated with the estrus, but hormonal profiles are normal. Histology is characterized by severely dilated infundibula filled with abundant keratin and often fragmented HSs. Follicles are in telogen, kenogen, or atrophic (Fig. 7d). 136
Pomeranians With Alopecia X
Alopecia X has been described in the past as adult-onset growth hormone deficiency (hyposomatotropism), growth hormone-responsive alopecia, castration-responsive alopecia, biopsy-responsive alopecia, and congenital adrenal hyperplasia-like syndrome. 39 By breeders, it is known as “black skin disease” or “coat funk.” It has been reported in several dog breeds that have a plush or wooly coat with a dense undercoat. Reported breeds are Pomeranians, Keeshonds, Schipperke dogs, chow chows, Samoyeds, Siberian Huskies, and Alaskan malamutes. In addition, alopecia X has been reported in toy and miniature poodles, although these breeds have a completely different coat type.23,39 The strong breed predisposition, pedigree analysis of affected dogs, and the onset of the disease at a relatively young age suggest a hereditary influence in the disease pathogenesis. As the vast majority of studies on alopecia X have been done with Pomeranians, it is the author’s opinion that the term alopecia X should be reserved for the specific hair loss disorder in Pomeranians or at a maximum for dogs belonging to the European Spitz breed according to the Federation Cynologique Internationale (FCI https://www.fci.be/en/nomenclature/5-Spitz-and-primitive-types.html#s4). These include the German spitz in all size varieties (keeshound, giant spitz, medium-sized spitz, miniature spitz, and Pomeranian) and the Italian spitz, which is also known as the Italian volpino. For these dogs, it is known that they are closely related, which makes it more likely that the hereditary cause for the alopecia might be shared across these breeds. All other breeds reported to have alopecia X are genetically further apart, and thus, different genes may cause the alopecia. Therefore, this author would propose to call their alopecic disorder alopecia X-like.
Alopecia X affects young adult Pomeranians and may present in dogs younger than 1 year of age. However, some dogs do not develop alopecia until they are 4 to 5 years. Alopecia is seen more often, but not exclusively, in male dogs. Hair loss is noninflammatory, progressive, and bilateral symmetrical and spares the head, tail, and distal extremities (Fig. 8a). The skin is often hyperpigmented. 17-hydroxyprogesterone (17-OHP) is increased in dogs with alopecia X and was thought to be the result of an abnormal activity of the 21-hydroxylase enzyme. Simultaneously, baseline cortisol concentrations correlated with progesterone concentrations in Pomeranians.41,122 Treatment results for Pomeranians with alopecia X suggest that alterations in the steroid hormone metabolism are causative for the HC arrest.2,21,40,42 This is also supported by a study that performed whole transcriptome profiling of skin biopsies from Pomeranians with alopecia X, which provided evidence of abnormal cutaneous steroidogenesis characterized by the dysregulation of genes encoding enzymes in the steroid hormone metabolism. 13 However, relevant gene variants were not identified in affected dogs with either whole genome sequencing or selected sequencing of genes involved in steroid hormone metabolism or HF development (21-hydrolase gene, cathepsin L2 gene, patched homolog 2 gene).81,82,134 In a recent study, it was suggested that an increased number of mitochondrial gene mutations are associated with the HC arrest in Pomeranian dogs. 60
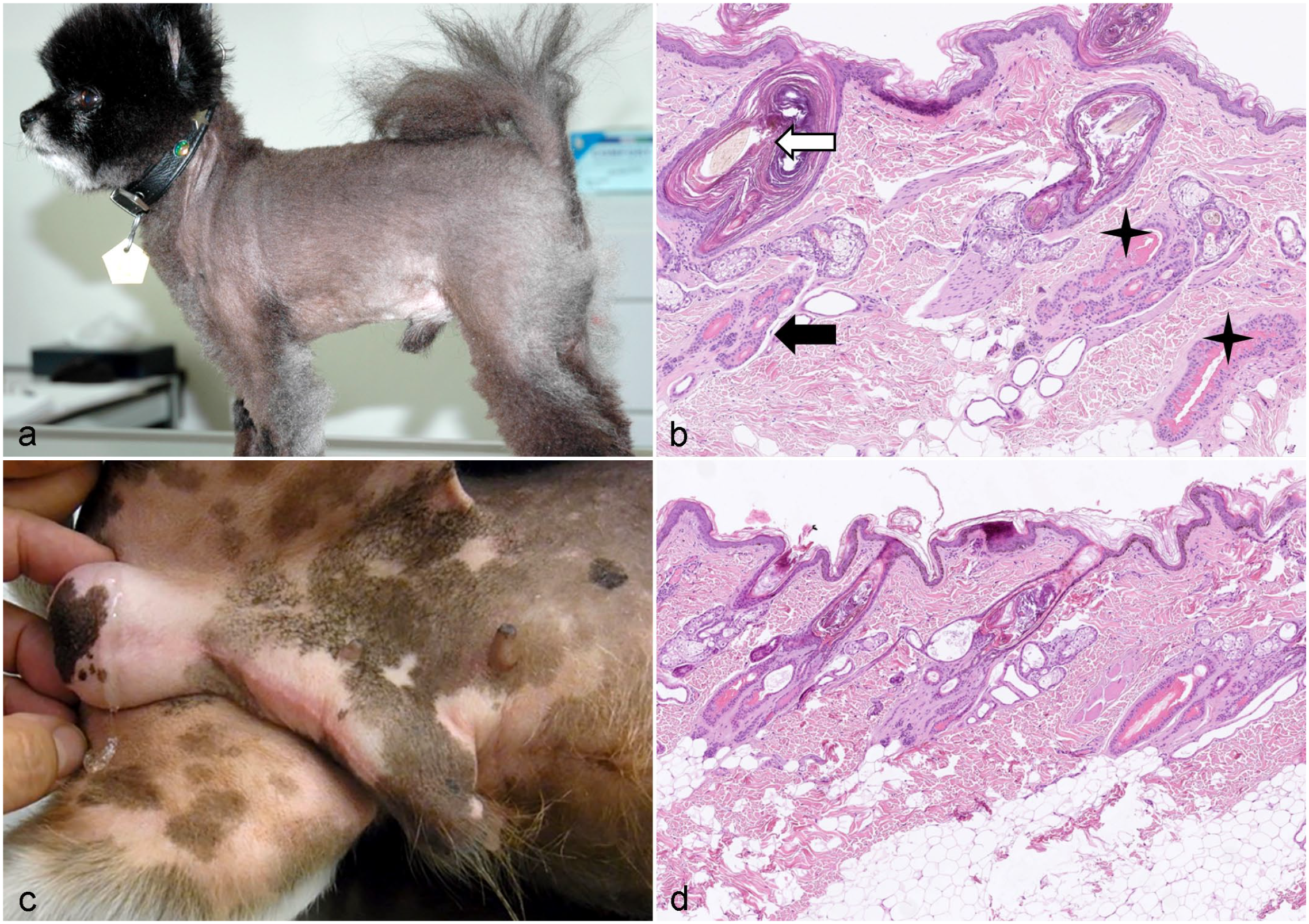
Histologically, alopecia X is characterized by a reduced number of anagen follicles and, compared with other HC disorders, a relatively high number of telogen follicles, as well as kenogen and atrophic follicles (Fig. 8b). These histologic findings resemble the findings in dogs with hyperestrogenism (Fig. 8d).13,81,98,82
Alopecia X-Like Disorders in Breeds Not Related to the European Spitz Breeds (Schipperke Dogs, Siberian Huskies, Alaskan Malamutes, Samoyeds, Chow Chows, Miniature, and Toy Poodles)
As outlined in the paragraph about alopecia X, the author prefers to use the term alopecia X-like disorder for the alopecic conditions of unknown cause in the other double-coated breeds and the miniature poodle. However, the clinical presentation and the histological findings reported for Schipperke dogs are similar to those reported for Pomeranians with alopecia X.13,82,83 The histological findings in the other dog breeds affected with an HC disorder of unknown cause have, to the best of the author’s knowledge, not been described in detail and thus cannot be compared with alopecia X.2,21,23,39,74
Color Dilution Alopecia
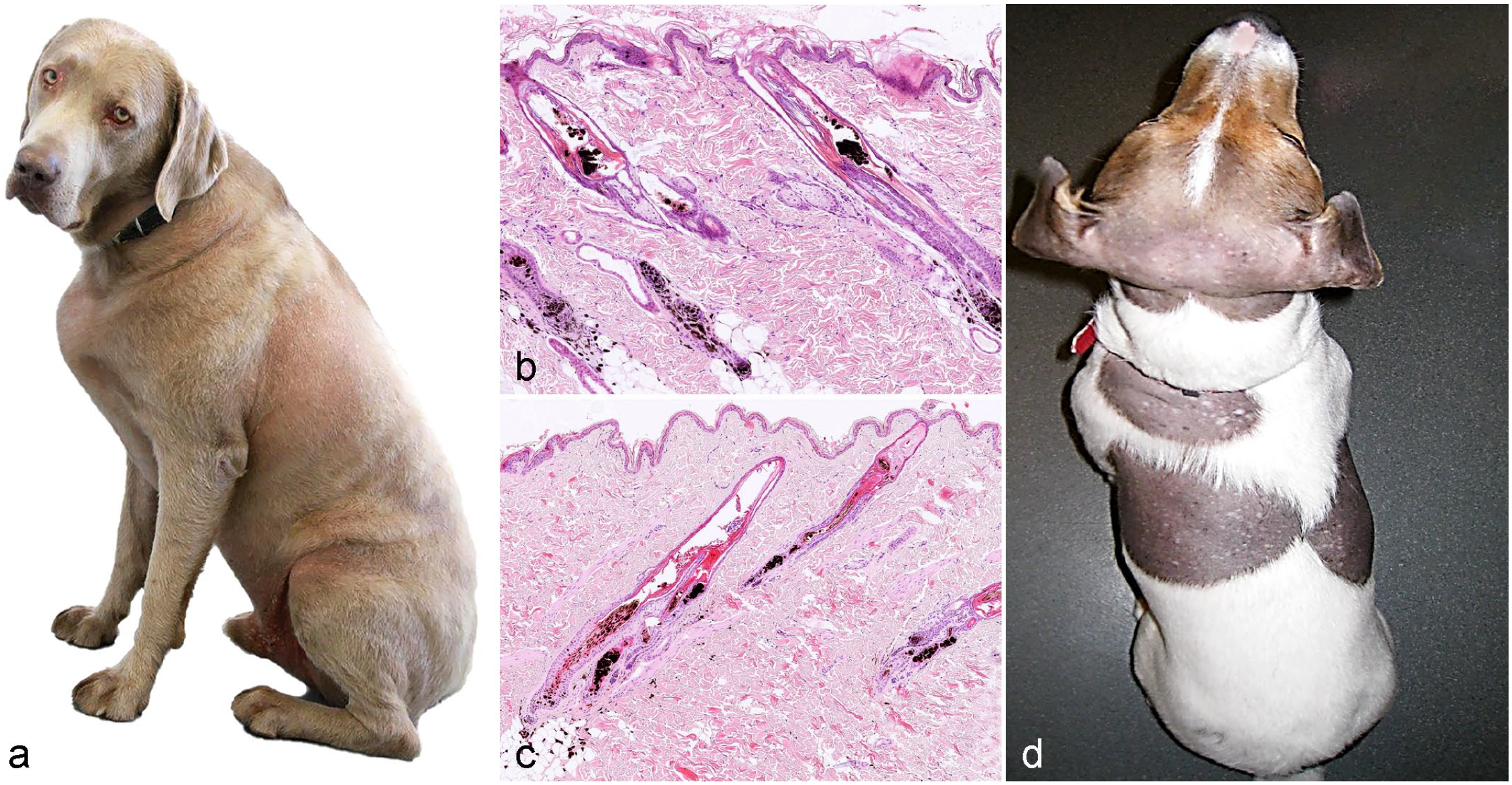
Color dilution alopecia can occur in all breeds with dilute coat colors, including mongrels, but is seen more commonly in blue Doberman pinschers, silver Labradors, Weimaraners, Yorkshire terriers, and Irish setters. The disease is also known as blue Doberman syndrome, fawn Irish setter syndrome, and blue dog disease. In dogs, coat color dilution, leading to the specific pigmentation phenotypes, blue, gray, fawn, or red, is caused by a defective transfer of melanosomes to the matrix cells in the HF bulb. Up to now, 3 variants in the melanophilin gene (MLPH) have been identified as causes of the dilute phenotype in dogs.8,34,138 The defect in melanosome transfer results in large aggregates of melanosomes, which appear histologically as large pigment clumps in the HS, the follicular lumen, and the outer root sheath. Despite the fact that MLPH gene variants cause coat color dilution in many dog breeds, not all dogs with a dilute coat color develop color dilution alopecia. Thus, yet unidentified factors appear to be required for the development of alopecia. These are most likely hereditary factors associated with 1 or several stages of HS pigmentation, including melanosome formation, melanin production, and transfer of melanosomes to the matrical cells of the hair bulb.
Clinically, the alopecia primarily affects the trunk, and histologically, large melanin aggregates are present within the HS and the outer root sheath (Fig. 9a, b). It has been suggested that the large melanosome aggregates damage the HS cuticle and fracture of the HS. As large melanosome aggregates are also present in normally haired dogs with a dilute hair coat (eg, Weimaraners or Doberman pinschers without alopecia), it is difficult to make a definite diagnosis based on histology alone and clinical information is needed. A ruptured HS cuticle, fractured HSs, and abundant free pigment may be histological keys to decide whether it is color dilution alopecia or not. If the HS cuticle in most HSs is intact, it is likely that the color dilution is not the cause for the alopecia. 150
Black Hair Follicular Dysplasia
Black hair follicular dysplasia is seen in bi- or tricolored dogs and has been reported in bearded collies, Jack Russell terriers, large Münsterländer dogs, salukis, and a New Zealand huntaway dog.66,75,96,123,143 Black hair follicular dysplasia is inherited as an autosomal recessive trait and is most likely related to color dilution alopecia. Within the first year of life, the black hair coat gets dull and brittle and the hairs break easily, finally resulting in complete alopecia of the black-haired areas of the head, pinnae, neck, and back (Fig. 9d). Like in color dilution alopecia, large melanin aggregates are present within the HS, the follicular lumen, and the outer root sheath; thus, abnormal processing and transport of melanin may play a role in the pathogenesis. Follicles are distorted and contain fragmented HSs (Fig. 9c). Scanning electron microscopy has shown that the HS cuticle is absent over large areas of the hair fiber. The white-haired areas are histologically normal. 143
Follicular Dysplasia in Weimaraners
Follicular dysplasia in Weimaraners, also a color dilute dog breed, affects young adults and presents with progressive alopecia of the trunk associated with recurrent folliculitis/furunculosis. The head and limbs are not affected. The HS abnormalities, the histopathological lesions, and scanning electron microscopy findings are similar, but less pronounced, to findings in color dilution alopecia. However, this form of alopecia occurs rarely in Weimaraners, and only few cases have been reported. 72
Alopecia in the Frisian Waterdog (Wetterhoun)
Alopecia in the Frisian waterdog has not yet been reported in the literature, but has been investigated within the frame of a doctoral thesis from the Vetsuisse Faculty of Bern. Clinically, dogs present with nonpruritic, noninflammatory bilateral truncal hair loss affecting only the pigmented coat, similar to the Pont Audemer spaniel. Less curly, thinner, and dull HSs are present in border areas. Histologic findings in alopecic skin consist of dilated keratin-filled infundibula, an increased number of kenogen HFs, a decreased number of anagen HFs, and decreased diameters of HFs and HSs. Small numbers of apoptotic keratinocytes were infrequently seen in the isthmus region. Small pigment aggregates in the HSs and the infundibular lumen were also present. Trichography indicates less curly hair and more HSs without medulla in border areas (Anja Ebner and Petra Roosje, personal communication).
Alopecia in Labrador Retrievers
An alopecic condition with early onset is seen in Labrador retrievers in the United States (Candace Souza, personal communication). The dogs have no underlying endocrinopathy and no coat color dilution. This condition has not been described in the literature, and histological findings are unknown.
Alopecia Associated With Miniaturization of the Hair Follicles (Canine Pattern Alopecia)
Canine pattern alopecia (pattern baldness) affects predisposed breeds with short smooth hair coats, such as dachshunds, Boston terriers, and Chihuahuas.47,78 Breed predilections suggest a genetic basis. Canine pattern alopecia develops within the 1 year of life, and the dogs develop a bilaterally symmetric thin coat on the pinnae; the skin caudal to the pinnae; the caudal thighs; the perineal skin; or the ventral neck, chest, and abdomen (Fig. 10a). HF miniaturization is a proposed key feature of the syndrome, but histology and trichograms, analyzing the HSs quality, have not been performed in most cases (Fig. 10b). 149 Thus, it cannot be excluded that some cases are rather associated with follicular dysplasia resulting in structurally impaired HSs, and the term pattern alopecia has been applied wrongly. This has been shown recently for sighthounds. 14
Follicular miniaturization is also seen in human female and male pattern hair loss. In humans, it has been proposed that follicular miniaturization is caused by a shortened anagen phase. As initiation of the next anagen phase is not more effective, the telogen phase is prolonged. The telogen follicles eventually lose their HS, which leads to an increase in kenogen follicles, which over time may get more and more atrophic, possibly ending in follicular deletion. 116
Follicular Lipidosis in Rottweiler Dogs
Follicular lipidosis is rare and has been only reported in 4 Rottweiler dogs.48,51 The affected dogs presented with partial alopecia on the mahogany or tan points of the face and feet within the first year of age. In neighboring areas, hairs turned mouse gray. In biopsies of affected skin, ballooning of the matrical cells due to lipid deposition was seen. Mainly hair bulbs of primary anagen HFs were affected. Rare vacuoles were also seen in HSs. Some HSs presented with an irregular contour and changes in the cuticle.
Pituitary Dwarfism in German Shepherd Dogs, Saarloos and Czechoslovakian Wolfdogs, and Tibetian Terriers
Pituitary dwarfism in German Shepherd dogs, Saarloos and Czechoslovakian wolfdogs, and Tibetian terriers is an autosomal recessively inherited disease, which is associated with mutations in the LIM homeobox3 gene in all 4 dog breeds.65,135,144,145 This gene variant leads to a defect in the organogenesis of the adenohypophysis, resulting in a combined deficiency of growth hormone, thyroid stimulating hormone (TSH), and prolactin, whereas the production of adrenocorticotropic hormone is normal.68,145 Besides growth retardation, affected dogs lack primary hairs and have prominent secondary hairs resulting in a puppy coat. Over time, bilateral symmetrical alopecia progresses and the skin becomes hyperpigmented.65,87
Acquired Alopecia Associated With an Impaired Hair Cycle
Hair Cycle Disorders of Endocrine Origin
Endocrinopathies (eg, hyperadrenocorticism, hyperestrogenism, and hypothyroidism) are due to imbalances in hormones and manifest as nonpruritic, bilateral symmetric alopecia or hypotrichosis resulting from a deregulation of the HC. The remaining hair coat is dull and dry, can be easily epilated, and fails to regrow after clipping. Whereas the cause for the endocrinopathy is known, in most cases the specific action of the deregulated hormones on the canine HF has not yet been studied. A recent review summarized the effect of various hormones on the HF. 52 As outlined above (Fig. 2), different hormones have a similar impact on the different HC phases. Thus, although the different endocrinopathies may have the tendency to present with specific histological features at some stage of the disease, histologic findings are similar and a definite diagnosis has to be based on the appropriate combination of clinical history, clinical features, clinical pathology testing, specific endocrine testing, and sometimes imaging.
In general, it can be said that, in addition to the lack/reduction in anagen HFs, typical histological features of associated with different endocrinopathies are:
Alopecia X and hyperestrogenism—mainly telogen and kenogen follicles (Fig.8b, d).
Hyperadrenocorticism—abundant kenogen follicles and severe follicular atrophy as well as dermal atrophy and a thin epidermis. Comedone formation is typical (Fig. 11a).
Hypothyroidism—anagen or telogen, normal sized primary follicles, and kenogen or atrophic secondary follicles. In addition, epidermal hyperplasia may develop if concurrent infections are present. Dermal mucinosis may be seen, but is rare (Fig. 11b).
Hair cycle disorders of endocrine origin
However, as stated above, these “typical” features may change during the disease course. For example, a dog with hyperestrogenism may present with severe follicular atrophy.
Hypothyroidism
Hypothyroidism is a commonly seen endocrinopathy in mainly older dogs of medium to large breeds. Clinical signs are associated with the deficiency of triiodothyronine (T3) and thyroxine (T4).90,105 In dogs, like in human beings, the HF is considered to be an important target for thyroid hormones. Deficiency of thyroid hormones is most often caused by idiopathic thyroid atrophy or lymphocytic thyroiditis. Clinical lesions of hypothyroidism start with a dull, dry, easily epilated hair coat that fails to regrow after clipping. Subsequently, alopecia develops in areas of hair coat friction, including the tail, elbows, hips, around the neck, and on the dorsal surface of the nose. Symmetric truncal alopecia is not as common as once thought (Fig. 11d). 38
Thyroid hormones affect the HF in various ways. From other species, the following data are available in the literature:
Thyroid hormones promote the anagen HC phase: Thyroid hormones (T3, T4) downregulate apoptosis of human matrix keratinocytes, and T4 upregulates their proliferation.
137
T4 prolongs the duration of anagen of human HFs in vitro.
137
In mice and rats, topically applied T3 enhances both skin and hair growth.
119
In female human scalp HF organ culture, TSH failed to have a significant effect on human HS elongation, anagen duration, or matrical cell proliferation. Thus, it was concluded that despite the presence of functional TSH receptors in human scalp HFs, the elevated levels of TSH seen in hypothyroid patients may result from thyroid hormone effects.
10
Hyperadrenocorticism (Cushing’s disease/syndrome)
Hyperadrenocorticism is a common cause of alopecia in dogs and the result of a prolonged exposure to inappropriately high plasma concentrations of (free) cortisol. Naturally occurring disease is usually caused by bilateral adrenal cortical hyperplasia secondary to a functional pituitary neoplasm, and less often by a functional adrenal cortical neoplasm.9,100,110 Iatrogenic hyperadrenocorticism is also common in dogs and is due to administration of exogenous glucocorticoids or treatment with trilostane.64,67 Rarely, accidental topical contact with glucocorticoids used on the skin of human beings may cause lesions, particularly in small dogs. The alopecia in hyperadrenocorticism spares the head and extremities and is accompanied by comedone formation and thin skin resulting in the risk of increased bruising, poor wound healing, and increased susceptibility to infections (Fig. 11c). Calcification of the dermis (calcinosis cutis) on the dorsal head, neck, and back, and sometimes also in inguinal and abdominal areas and extremities, can occur in dogs. It is seen most often with iatrogenic hyperadrenocorticism. 38
Glucocorticoids affect the HF in various ways. From other species, the following data are available in the literature:
Impaired anagen induction Steroids result in the reduction in DP cells in mice. The DP plays a crucial role in the activation of follicular stem cells and when the number of DP cells is too low the HF fails to enter anagen.
26
Shortened anagen and premature catagen induction In mice, glucocorticoids reduce the mitotic activity of hair matrix cells and stimulate their differentiation. This finding could be in line with a shortened anagen cycle phase resulting in premature catagen induction.
107
Dexamethasone treatment reduces the number of human DP cells in vitro and downregulates the expression of the KI-67 protein, vascular endothelial growth factor (VEGF), and hepatocyte growth factor (HGF).
28
It is known that the DP is essential for the growth and the differentiation of follicular keratinocytes. Elevated levels of plasma glucocorticoids in chronic stress also inhibit the proliferation of DP cells and suppress the expression of VEGF.
80
Dexamethasone increases the expression of dickkopf-1 (DKK1), a known catagen inducer, in DP cells and a neutralizing DKK1 antibody attenuates the Dexamethasone-induced inhibition of HS elongation and delays Dexamethasone-induced catagen in mice.
71
Dexamethasone induces the activity of the androgen receptor in human and murine DP cells, which might result in a shortened anagen phase.
70
Hyperestrogenism
Hyperestrogenism can develop in male and female dogs. In females, higher estrogen levels originate from ovarian cysts, and rarely ovarian neoplasm. In males, elevated serum estrogen concentrations are mostly derived from functional sertoli cell tumors or less commonly testicular interstitial cell tumors. Estrogen administration for the treatment of estrogen-responsive urinary incontinence, vaginitis, or prostate hypertrophy and hyperplasia has also caused hyperestrogenism in dogs.47,128 Rarely, accidental topical or oral contact with estrogens used on the skin of human beings may cause lesions due to skin contact or licking, particularly in small dogs. 152 The bilateral symmetrical alopecia often begins in the perineal–genital region before it progresses to the abdomen, caudal and medial thighs, chest, flanks, and neck (Fig. 8c). Hyperpigmentation may be seen. In addition to endocrine alopecia, female dogs have enlarged vulvas and abnormalities of the estrus cycle. Male dogs can develop gynecomastia, a pendulous prepuce, a linear preputial erythema (Fig. 8c), or an enlarged prostate because of squamous metaplasia of prostatic ducts. 38
Estrogens affect the HF in various ways. From other species, the following data are available in the literature:
Impaired anagen onset: In mice, it has been shown that estrogen induces apoptosis of precortex cells and causes premature catagen induction by the upregulation of transforming growth factor β2 (TGF-β2). Immediately after the HF enters premature catagen, the expression of bone morphogenic protein 4 (BMP4) is increased. It is speculated that the upregulation of BMP4 prevents the transition of HFs from telogen to anagen, thus resulting in an increased number of telogen follicles.
56
Premature catagen induction Estrogens alter gene expression of several HC-relevant genes and signaling pathway members, such as the mitogen-activated protein kinase pathway and the TGF/BMP family, and are thus involved in the initiation and promotion of catagen by the induction of apoptosis.
103
In isolated human HFs, it has been shown that estrogen inhibited matrix keratinocyte proliferation and enhanced their entry into apoptosis.
11
Miscellaneous Noninflammatory Acquired Alopecic Disorders
Post-Clipping Alopecia
Post-clipping alopecia is characterized by a lack of hair regrowth after clipping. The underlying cause is unclear, but it seems to occur more often in dogs with a long telogen HC phase (eg, Siberian huskies and other breeds with a thick undercoat). 127 Thus, it may well be that many cases of post-clipping alopecia are due to the fact that hair regrowth after clipping takes longer because the dogs have been clipped in an early telogen stage and hair will regrow only when the long telogen phase in these dogs is finished and the HFs reach the next late anagen stage. In addition, vascular perfusion changes in response to cutaneous temperature changes after clipping and underlying preclinical endocrinopathies have been discussed as possible causes. 78
Alopecia Associated With Excessive Hair Loss—Telogen Effluvium
Telogen effluvium is rare in dogs and is the result of excessive premature shedding of the HSs (exogen). Thereafter, synchronized initiation of new anagen occurs. Thus, the major histological finding is that all HFs are in telogen or more often already in anagen. Dogs are nevertheless bald since the HSs have not yet reached the skin surface. The underlying cause for the synchronization of the HC in dogs is not known. 47 In humans, various causes of telogen effluvium have been identified, such as stress, illness, medications with all-trans retinoic acid and salicylic acid, excessive ultraviolet light, nutritional deficiencies, lymphocytotoxic (autoimmune) activity, and long-term use of estrogen-containing contraceptives. 27 It is likely that similar causes induce excessive shedding in animals.
Anagen Effluvium/Defluxion
Anagen effluvium is due to interruption of the mitotic activity of the rapidly proliferating anagen hair bulb matrix cells. It is most often related to chemotherapy and is best documented in humans. It is rarely seen in dogs and mostly affects breeds with an anagen-dominated HC, but other breeds may be affected as well. In dogs, chemotherapy-associated anagen effluvium has been most commonly associated with doxorubicin and 5-fluorouracil therapy. However, other chemotherapeutic agents result in alopecia as well.36,43 The abrupt cessation of mitotic activity of the matrix cells leads to a narrowing of the proximal portion of the HF and the production of fragile HSs. When these fragile HSs reach the skin surface, they break and anagen effluvium occurs. In 1 study, the onset of alopecia after the beginning of the drug administration ranged from 7 to 120 days, and the time to resolution ranged from 42 to 150 days. 36 Diagnosis is based on clinical history, physical examination, and microscopic examination of pulled hairs (trichograms). In the early course of the disease, the pulled hairs have irregularly narrowed or pointed ends. In the late course, only telogen hairs, which are not mitotically active, can be pulled since all anagen hairs have been lost.
Alopecia Associated With Ischemic Dermatopathy
Ischemic dermatopathy is caused by impaired vascular perfusion and is associated with an underlying vasculopathy or vasculitis. In a recent study, miniature poodles, Chihuahuas, Maltese, Yorkshire terriers, and Jack Russell terriers were significantly over-represented, and about 50% of the cases were associated with a vaccine. 6 Clinically, focal or multifocal alopecia is the most common lesion. Lesions are found mostly on the pinnae, vaccination sites, periocular regions or in the face, and on the tail (Fig. 10c). Besides alopecia crusts, scale, erythema, erosions or ulcers, and hyperpigmentation are seen commonly. Pruritus may be present. Systemic signs have been seen in about a third of the dogs.6,47 Histologically, primary and secondary HFs are severely atrophic and may be absent. HFs are described as “fading” follicles. They are devoid of HSs, and only a thin, short epithelial strand remains in the HF. Prominence and homogenization of the connective tissue surrounding the HF is a frequent finding (Fig. 10d). Vascular lesions are seen in the capillaries and small arterioles and are characterized by loss of vascular distinction or endothelial cells. Inflammation of the vessel walls is seen rarely in dogs. The dermal collagen is pale and is described to have a smudged appearance. Lymphocytes and plasma cells are distributed randomly (“confetti-like”) throughout the dermis and eventually the panniculus. Dermal edema and/or mucin deposition is seen frequently. Vacuolation of epidermal basal keratinocytes and rare apoptotic cells, described as cell poor interface dermatitis, may be seen.47,142 The detailed pathogenic mechanisms of how ischemia affects the HFs are unknown. In 1 study, it has been shown that under hypoxic culture conditions the capacity of follicular stem cells to form clonal colonies is impaired. 113
Traction Alopecia
Traction alopecia is the results of chronic partial ischemia that is not associated with an underlying vascular pathology. It occurs when rubber bands are applied tightly to retain the hair and is the result of a repetitive or continuous impaired blood supply to HFs and adjacent skin. The impaired blood supply is due to shearing and compression of the smaller diameter blood vessels due to prolonged forces of traction. It most often presents as focal alopecia on the top of the head or on the ears in longhaired dogs. Once developed, the alopecia is long term and generally permanent. Histologically, depending on alopecia duration, HFs are either in kenogen or are atrophic, and may be partially or completely lost and replaced by dermal fibrosis. Adnexal glands are also usually atrophic and may be absent. 47
“Postinflammatory Alopecia” and Hair Cycle Arrest of Unknown Cause
In addition to the disorders described, noninflammatory alopecia, associated histologically with an impaired HC, may be seen in cases for which the causes and/or the diagnoses remains unclear. As outlined in the review on HF structure, morphogenesis, and regeneration in the same issue of this journal, the HC is a tightly regulated and highly conserved process that involves numerous signals derived from epithelial, neuroendocrine, and mesenchymal cells. These signals converge on HF stem cells, transient amplifying cells, and also during cytodifferentiation. Many of these signals have not yet been identified in rodents or dogs. Nevertheless, these finely balanced activators and inhibitors have an impact on the normal function or dysfunction of the HC. A detailed review on the impact of inflammation on the HC and epidermal regeneration is provided in a recent review. 91
Examples of possible mechanisms resulting in an impaired HC are as follows:
In mice, sonic stress results in the recruitment of substance P and nerve growth factor, which results in perifollicular inflammation, HF keratinocyte apoptosis, premature HF regression (catagen induction), and inhibition of HF keratinocyte proliferation. 108
In systemic stress conditions, plasma glucocorticoids are elevated through activation of the hypothalamo-pituitary-adrenal axis. 4 The effects of steroids have been discussed earlier.
Chronic restraint stress increases the number of substance P immunoreactive nerve fibers and activates mast cells. 80
Chronic restraint stress inhibits hair growth by prolonging the telogen stage and delaying the subsequent anagen and catagen stage. 80
Stress is associated with premature catagen development in mice. 5
In humans, examination stress results in shift the immune response to a TH1 response and transiently hampers hair growth. However, the changes stay within a physiological range. 109
Mast cell degranulation triggers catagen induction. 5
Skin-resident macrophages undergo apoptosis prior to SC activation at anagen onset. This process is linked to distinct gene expression, including Wnt transcription. Experimental induction of macrophage apoptosis during early telogen results in stem cell activation in vivo. A macrophage-specific pharmacological inhibition of wingless / integrated (WNT) production delays HF growth.19,97
A subset of dermal macrophages secretes oncostatin M, maintains HF stem cell growth, and inhibits hair growth. 147
Data from mice suggest that interleukin (IL)-6 family members inhibit hair growth in a dose-dependent manner. As increased levels of IL-6 are present in dermal inflammation, IL-6 may play a partial role hair growth inhibition seen in inflammatory disease. 154
These data at least suggest that also in dogs, stress or inflammatory mediators released by mast cells, or other cells of the innate immune system, may result in HC arrest disorders.
Summary
Noninflammatory alopecia in dogs may be present at birth, develop early at life with a presumed hereditary cause, or is clearly acquired. The underlying gene variants or the detailed underlying pathogenesis is unknown for most cases. Since the HF has only limited possibilities to respond to an impaired regulation and since histopathology may change during the course of a disease, histologic findings can be similar in different diseases. Thus, histology may suggest an underlying cause, but is not reliable. A detailed clinical history, a thorough clinical examination including blood work, the appropriate selection of the biopsy site, and the detailed histological findings need to be combined to achieve a final diagnosis.
Footnotes
Acknowledgements
The author would like to thank Keith Linder for helpful discussions about the HF during the writing process of the chapter about the integument for the seventh edition of Pathologic Basis of Veterinary Disease. I am grateful to all dermatologists that were offering clinical images for this review and my colleagues that were working with me on different HF projects over the past years. A great thank you also to the editors and guest editors of this journal that invited me to write this review and Hannah Bender for her great support in creating the figure panels.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.