
Abstract

Keywords
Equine herpesviruses (EHV) 1 and 4 belong to the Herpesviridae, subfamily Alphaherpesvirinae, and are considered the most important equine viral respiratory pathogens along with equine influenza virus. Both viruses are endemic worldwide due to high incidence of infection early in life with establishment of latency in a high percentage of infected horses. 2 Following primary infection, reactivation of latent virus with subsequent shedding and transmission to naive hosts is common. Currently, it is estimated that as many as 80% of all horses are latently infected with EHV-1, and seroprevalence for EHV-4 is even higher. 12
Infections with EHV-1 and EHV-4 cause respiratory disease, and these are clinically indistinguishable unless the virus is identified in diagnostic samples. Respiratory disease in older or middle-aged horses is often mild. In contrast, it can be significant in young horses, in which it is characterized by a biphasic fever, depression, anorexia, coughing, and nasal and ocular discharge and can lead to secondary bacterial infection involving the lungs and lasting for 2 to 3 weeks.
Whereas EHV-4 typically stays restricted to the respiratory tract, EHV-1 also causes late-term (last-trimester) abortions and premature delivery of foals that die soon after birth. In addition, EHV-1 has been detected in the semen of infected stallions, although venereal transmission has not been reported. 14 Furthermore, EHV-1 is the cause of equine herpesvirus myeloencephalopathy (EHM) in up to 10% of infected horses. 1 Neurological signs include head pressing, ataxia, and paralysis with complete recumbency requiring euthanasia. Finally, EHV-1 infection can lead to chorioretinopathy, causing permanent lesions with “shotgun” morphology in a substantial proportion of infected horses (Fig. 1). 7 Lesions can be focal, multifocal, or, on rare occasions, diffuse, affecting the whole eye. Clinically, only diffuse lesions have a significant impact and cause loss of vision.
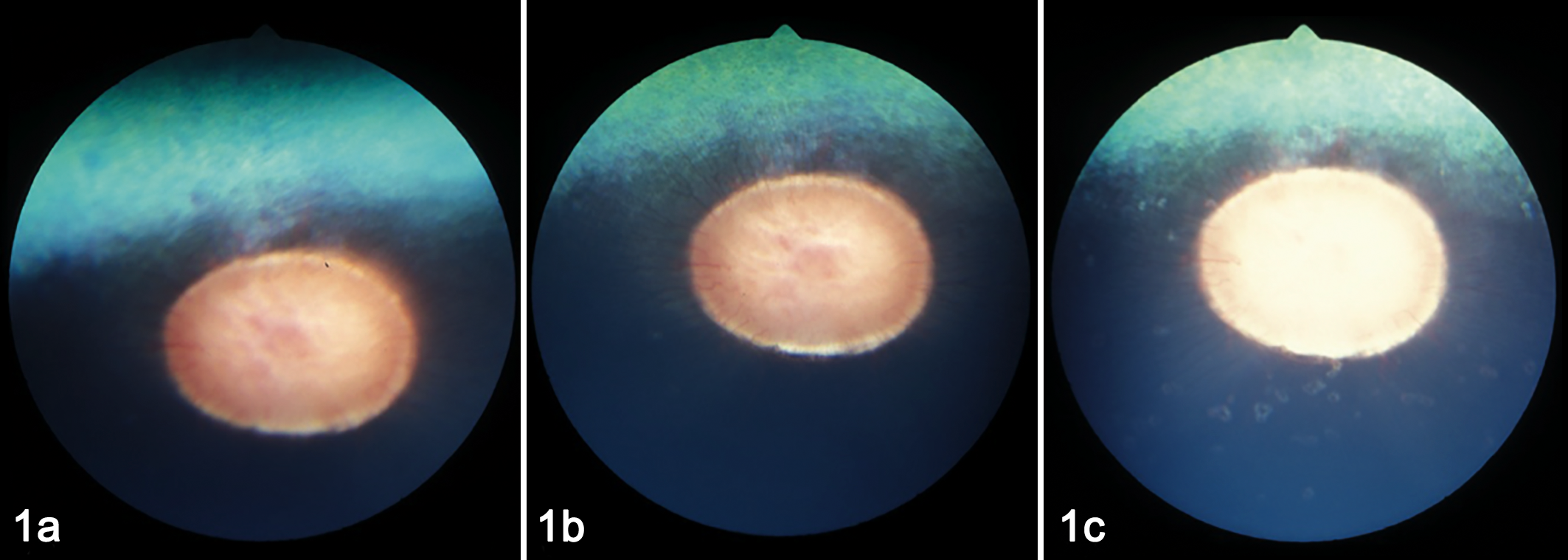
Development of ocular lesions following experimental infection of a yearling horse with equine herpesvirus 1. 7 Horses were infected intranasally with 5 × 107 plaque-forming units of a neuropathogenic strain of EHV-1 (Ab4). (a) Before infection. (b) One month after infection. (c) Fifty-four days after infection. Images were taken by fundus photography following sedation with detomidine (0.02–0.04 mg/kg, intravenous) and dilation of the pupil with 2 drops of 1% tropicamide and 2% phenylephrine. Images show the development of focal (b) and multifocal (c) small and irregularly circular lesions that occurred close to the optic disc usually within the limits of the retinal vasculature. The lesions presented as classical donut-shaped, depigmented lesions of the chorioretina with pigmented centers.
Respiratory disease caused by EHV-1 and EHV-4 results in inflammation, congestion, and sometimes necrosis of the upper respiratory tract, and the lungs may become involved in later stages. One can find typical herpesvirus inclusion bodies in the nuclei of the respiratory epithelium. Abortions can be a consequence of EHV-1 infection of the fetus with widespread hemorrhages and edema, splenomegaly, and inclusion bodies in fetal organs. Alternatively, abortion can result from infection of the vasculature of the endometrium without infection of the fetus. In this case, the lesions in the uterine vasculature largely resemble the vasculitis observed in the neurological form of the disease. For the neurological form, the consistent histopathological changes are vasculitis and thrombosis of small blood vessels in the spinal cord or brain, myelitis with axonal degeneration, perivascular cuffing, and lymphocytic and histiocytic infiltrates. However, gross pathological and histopathological alterations are sometimes not obvious, and more sensitive methods are needed for a definite diagnosis of EHV-1 abortion or EHM, such as immunohistochemistry, in situ hybridization, virus isolation, polymerase chain reaction (PCR), or quantitative PCR (qPCR). Interestingly, histopathological changes are also common in the chorioretinal vasculature and the testis following EHV-1 infection and mirror those observed in the central nervous system and uterus (Figs. 2–6). The relevance of these findings for fertility and long-term vision remains to be further investigated.
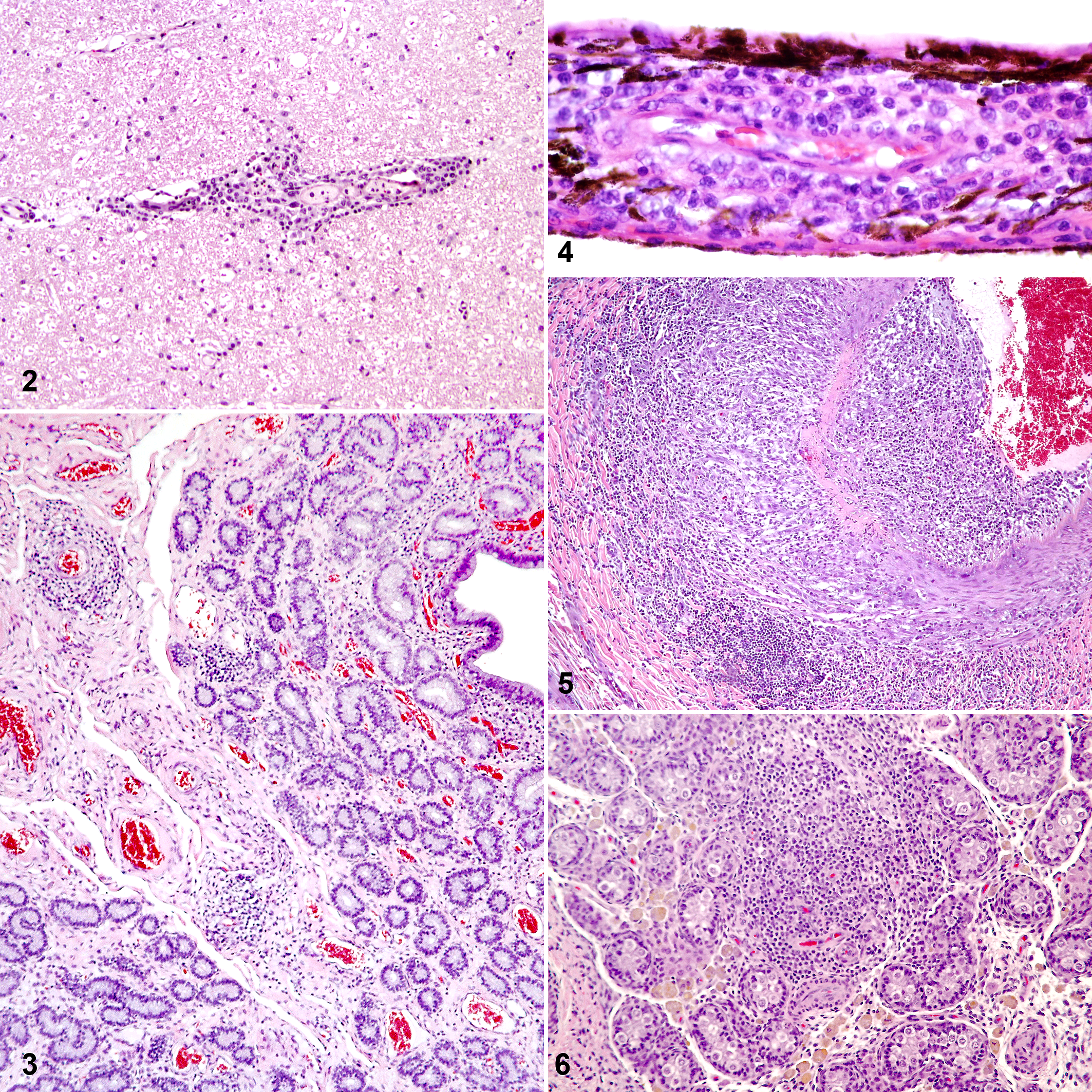
Equine herpesvirus 1 infection, 1-year-old horse, 9 days after infection intranasally with 5 × 107 plaque-forming units of a neuropathogenic strain of EHV-1 (Ab4). The horse showed clinical signs of equine herpesvirus myeloencephalopathy following the infection.
6
Although EHV-1 and EHV-4 respiratory infections are clinically indistinguishable, their pathogenesis is different, and because of that, secondary manifestations of EHV-4 are rare. Primary infection with both viruses occurs via the respiratory tract. Following replication in the airway epithelium, EHV-1 is detected in the local lymph nodes by 24 to 48 hours postinfection, and a cell-associated viremia is established between days 4 and 10 postinfection. This cell-associated viremia transports the virus to sites of secondary infection, where contact between infected leukocytes and the endothelium leads to endothelial cell infection, inflammation, thrombosis, and tissue necrosis, as well as secondary disease manifestations directly following viremia. While a positive correlation between viremia and incidence of EHM exists, only a small percentage (∼10%) of viremic horses subsequently develops EHM. A combination of host and viral factors is likely associated with the incidence of EHM, including age, breed, sex, and season. In addition, a single-nucleotide polymorphism identified in the viral polymerase gene results in a coding change (D752 vs N752) and is strongly associated with neuropathogenicity (D752) and high levels of viremia. 12 However, both D752 and N752 viral strains can cause EHM, and biosecurity should be the same regardless of the viral strain isolated.
In contrast, a viremia is not typically established with EHV-4 infection, and this is the main reason that nonrespiratory disease manifestations are rare following infection with EHV-4. The basis of the differences in the pathogenic potential between these genetically closely related viruses may be related to differences in viral entry. 9 Generally speaking, attachment and entry into host cells for most herpesviruses is facilitated by complex interactions between host cell receptors and herpesviral glycoproteins, including gB, gC, gD, and gH/gL. For EHV-1, Frampton et al 4 showed that EHV-1 gD uses equine major histocompatibility complex-1 (MHC-1) as an alternative receptor for viral entry into equine cells. Furthermore, recent studies showed that while MHC-I binding contributes to entry into fibroblasts, equine brain vascular endothelial cells, and cells of epithelial origin, it is not required for entry into other cells. 8,13 This indicates that EHV-1 uses different entry pathways to infect different target cell populations of its natural host, and glycoprotein D is in an important viral determinant in this process. 11 Interestingly, swapping glycoprotein D between EHV-1 and EHV-4 using bacterial artificial chromosome (BAC) mutagenesis transferred resistance/permissiveness of different cells to EHV-1/4 respectively, providing direct evidence that the cell tropism of EHV-1 and EHV-4 in vitro depends on gD. 3
To follow up on these in vitro data, an in vivo study by Holz et al 6 tested the hypothesis that replacing gD of EHV-1 with gD of EHV-4 (EHV-1gD4) would prevent establishment of viremia and thus EHM in horses. While no EHM was observed and respiratory disease was attenuated, viremia was reduced but not prevented, and the onset of viremia was delayed following infection with EHV-1gD4 compared to wild-type infection. In addition, histological changes were similar in the spinal cord, testis, and the endometrium and more prevalent in the eyes of horses infected with EHV-1gD4, 5 suggesting that additional viral determinants contribute to the establishment of viremia and secondary EHV-1 disease manifestations. Similarly, a current study by Minato et al 10 in this issue of Veterinary Pathology generated transgenic mice expressing equine MHC-I and showed more severe EHV-1 infection and necrosis in the bronchiolar epithelium of the transgenic mice expressing a low level of equine MHC-I, but neither histological lesions nor viral antigen were detected in the liver, spleen, kidneys, heart, or brain of transgenic or wild-type mice inoculated with EHV-1. While generation of transgenic mice with more widespread overexpression of equine MHC-I may provide a suitable model for the study of EHV-1/EHM pathogenesis, it is important to remember that multiple EHV-1 proteins are likely involved in each step of the pathogenesis, as are differing host factors/receptors and host genetic backgrounds.
Utilization of technology to generate alternative in vivo models like the murine model proposed by Minato et al, 10 as well as taking advantage of the unique genetic relatedness of EHV-1/4 and the ability to selectively alter individual viral genes using BAC mutagenesis or CRISRP/Cas9, is likely to significantly advance our understanding in the future and inform natural host studies necessary to fully understand EHV-1/4 pathogenesis and find solutions for protection.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.