
Abstract
The authors describe pathological and microbiological features of mortalities in a captive breeding colony of Lord Howe Island stick insects (Dryococelus australis) over a period of 18 months. There were 2 peaks of mortality in this period. In the first, insects presented dead with minimal premonitory signs of illness. In the second, affected insects were ataxic with contracted limbs and inability to climb or right themselves. Gross lesions were uncommon but included pigmented plaques on the gut and cloacal prolapse. Histological lesions in both outbreaks indicated a cellular innate immune response including nodulation characterized by Gram-negative bacterial bacilli entrapped within nodules of pigmented hemocytes, and melanization characterized by melanin within hemocyte nodules and around bacteria. Hemolymph culture findings varied and often yielded a mixed growth. Pure growth of Serratia marcescens was cultured in 44% of animals in Outbreak 1, while pure growth of Pseudomonas aeruginosa was cultured in 30% of animals in Outbreak 2. Cases with S. marcescens-positive culture often showed inflammation at the foregut-midgut junction. The frequency of mixed bacterial culture results did not allow firm conclusions about causality to be made, and may indicate primary bacterial infection or increased susceptibility to hemolymph colonization with an opportunistic pathogen. These findings highlight the utility of histopathology combined with ancillary testing when investigating mortality in captive insect colonies.
Keywords
Insects form a minor component of routine diagnostic veterinary pathology caseload. Selected insect species have increased perceived value due to their association with agriculture and research, striking physical features, endemicity, or endangered status. Colonies with conservation or display value require monitoring for health and disease. Knowledge of normal organ histology and tissue response to injury is essential for identification of genuine lesions, and interpretation of their significance. Informed examination of these species by diagnostic veterinary pathologists requires ongoing contributions of reference and case material to the scientific literature.
The Lord Howe Island stick insect (Dryococelus australis) is critically endangered and endemic to the Lord Howe Island group in the southwestern Pacific. 3 The population was decimated by black rats (Rattus rattus) arriving with European settlement, and was considered extinct by the 1920s. 6 However, 2 reports of dead insects on nearby Ball’s Pyramid were made in the 1960s by recreational rock-climbers. An expedition in 2001 lead to the discovery of a single colony of 24 individuals. 21 Two breeding pairs were removed in 2003 to establish captive breeding colonies at Melbourne Zoo (Parkville, Victoria, Australia) and a private insectary. 11 While the Melbourne Zoo colony thrived after initial difficulties, the second captive population died out. 4 Current wild population numbers are unknown, but totaled only 40 when surveyed in 2005. 4 The principal aim of founding a captive Dryococelus australis colony was to conserve this species, with the hope that a viable wild population could in the future be reestablished on Lord Howe Island.
A previous publication concerned with the husbandry management of Dryococelus australis at Melbourne Zoo briefly described pathological investigations on a small number of insects. 11 These yielded varied gross findings including desiccation, cuticular damage, poor body condition, and in 1 case a focus of green pigment embedded in the gut wall. Histopathological findings were reported as suspected cuticular fractures and 1 case of midgut ulceration associated with bacteria. Microbiological findings were not reported.
The present study describes the gross, histological, and microbiological findings of mortality investigations in this population over an 18-month period. The objective was to document, characterize, and compare the gross and histological lesions in relation to microbiological findings.
Materials and Methods
The captive colony of Dryococelus australis at Melbourne Zoo is managed in climate controlled glasshouses with individuals grouped by age class. Adult and subadult insects are housed together and juveniles separately. Temperature and humidity control are automated to maintain a target temperature of 22–26°C and target humidity of 70–80%, and measured daily. Insects are fed a variety of living plants raised on-site including species native to Lord Howe Island. Unfiltered mains water is provided in shallow plastic dishes and lighting is ambient. Adult glasshouses typically house 100–200 animals each. Enclosures and glasshouses are inspected daily for dead or moribund animals.
During the 18-month period 2 periods of higher than expected mortality were observed. The first (“Outbreak 1”) occurred in multiple glasshouses housing adults and subadults and in glasshouses housing juveniles. Outbreak 1 occurred between October 2013 and November 2014, with mortality slowly increasing to a peak in June 2014. The second period of increased mortality (“Outbreak 2”) occurred between December 2014 and March 2015, affected primarily adult and subadult insects in multiple glasshouses, with mortality increasing rapidly to peak in January 2015.
During Outbreak 1, 279 juvenile, adult, and subadult insects were examined and necropsied, and 337 in Outbreak 2. Examinations and postmortems were conducted on-site. Moribund animals were euthanized by placing the insect within an airtight container with a cotton ball saturated with isoflurane liquid (Isothesia®, Henry Schein®, North Gate, QLD, Australia) for a minimum of 30–60 minutes, until all signs of movement and responsiveness were absent and remained absent after removal from the chamber. Postmortem reports were available for 258/279 (93%) insects in Outbreak 1 and 202/337 (60%) in Outbreak 2. Given the large number of insects examined, gross findings were not consistently recorded. Where recorded, hemolymph volume and color, body condition, and gut fill were assessed subjectively. Normal hemolymph was considered to be clear to pale yellow. Hemolymph that was brown was considered discolored. Hemolymph volume was considered increased if upon incising the body cavity there was more than a small amount of fluid present, or if fluid noticeably distended the abdominal segments prior to incision.
A proportion of insects examined were selected for laboratory investigations. Hemolymph was sampled in 18 insects in Outbreak 1 and 37 insects in Outbreak 2. The ventral abdominal cuticle was swabbed with alcohol then incised using sterile instruments. The abdominal hemolymph was then swabbed with a sterile transport swab (COPAN Italia, Brescia, Italy). Hindgut was sampled in 7 insects in Outbreak 1 by incising the gut wall with a sterile scalpel and swabbing the intestinal contents. Hemolymph swabs were also submitted for 2 of these insects. A single swab of substrate crevices in 1 adult glasshouse was submitted. Swabs were individually streaked on prepoured plates (ThermoFisher Scientific, Australia) of horse blood Maconkey agar and horse blood agar with colistin and nalidixic acid (HBCNA) and incubated at 37°C in 5% CO2, on horse blood agar with neomycin (HBNEO) and on horse blood agar (HBA) without antibiotics and incubated at 37°C anaerobically, and onto Sabouraud agar and incubated at 37°C. Aerobic bacterial plates were read after overnight incubation. Anaerobic bacterial and fungal plates were read after 48 hours. Isolates were identified either by matrix assisted laser desorption ionization–time of flight (MALDI-TOF) identification using a Microflex Biotyper (Bruker, Germany), or by automated biochemical identification (VITEK® Compact, BioMérieuz, France). MALDI-TOF samples were prepared by direct cell profiling, where a single bacterial colony was picked, spotted directly onto the target plate and overlaid with 1 µl 70% formic acid solution prior to analysis. Some cases of suspected Pseudomonas aeruginosa growth were confirmed by subplating onto Cetrimide agar (ThermoFisher Scientific, Australia) and incubating at 42°C for 24 hours, with positive identification indicated by growth of bright green colonies. Antimicrobial disk diffusion susceptibility testing was performed according to the methods and standards of the Clinical Laboratories Standards Institute (CLSI).
Twenty-four insects were submitted for histology from Outbreak 1, comprising 20 subadults/adults and 4 juveniles. Forty-one insects were submitted from Outbreak 2, including 3 juveniles. All animals sampled for culture were submitted for histology. Insects were placed in 10% neutral buffered formalin after microbiological sampling. Standardized sections were developed throughout the outbreak to include a longitudinal section of the head and proximal hindlimb segment, longitudinal sections of the entire gastrointestinal system, and 2–3 transverse sections of the body cavity including thorax and abdomen. Cassettes containing exoskeleton were softened in 10% potassium hydroxide (Ajax Finechem, Taren Point, Australia) solution for 1 hour, and dilute hair conditioner (generic brands, 25 ml in 1 L of warm tap water) for 4 hours prior to processing. Tissues were paraffin-embedded for routine histological processing using 5 µm sections stained with hematoxylin and eosin. Gram stain (Brown-Hopps modification) was performed in 4 cases in Outbreak 1 and 2 cases in Outbreak 2. Periodic acid-Schiff (PAS), Grocott’s methenamine silver (GMS), and Ziehl-Neelsen (ZN) stains were performed on selected cases using routine methodology. All cases were examined by a single pathologist (C. Bayley). References for insect organ histology were available for some organs, and where no references could be found, gross organ identification was correlated with histology. 5,15,22 In a minority of cases some organs, generally the gonad or cloacal tissues, were not captured for examination. Histological lesions were scored using a semiquantitative system, where the presence of hemocyte nodules was scored from 0–3 and averaged for each organ to achieve a lesion score (LS), where 0 = no lesions, 1 = 1 hemocyte aggregate or nodule, 2 = 2 to 4 hemocyte nodules, and 3 = 5 or more hemocyte nodules, or large confluent nodules. Hemocyte aggregates were defined as closely aggregated clusters of hemocytes not forming a distinct nodule, whereas nodules comprised cohesive sheets and nodular aggregates.
Control animals were not submitted at the time of the outbreak. For the purposes of this study and to aid in population health monitoring, a control animal comprising a clinically healthy adult from the contemporary (2017) population was euthanized and submitted for gut and hemolymph culture and histology.
Results
The mean mortality rate for the subadult/adult population has been estimated as approximately 2.5% per week (C. Cheng, unpublished data). The weekly mortality rate in adults/subadults peaked at approximately 19% in the worst affected glasshouse in Outbreak 1 and approximately 21% in Outbreak 2. Dead juvenile insects were rapidly destroyed or removed by predation, precluding accurate estimation of their mortality rate.
Clinical History and Necropsy Findings
Clinical signs were minimal in Outbreak 1. Insects were either found dead or noted to have reduced movement immediately prior to death. In Outbreak 2, clinical signs were commonly seen for approximately 24 hours premortem. Affected adults and subadults were typically found on the floor showing ataxia, slow movement, and inability to right or climb. Dead insects were found in lateral or dorsal recumbency, often with the body and legs curled (Fig. 1).

There were few gross findings recorded in either outbreak (Table 1). Of the insects for which postmortem records were available (258 in Outbreak 1 and 202 in Outbreak 2), hemolymph volume was more frequently estimated as increased in Outbreak 1 (12%) than Outbreak 2 (2%). Hemolymph was discolored brown in 11/258 insects (4%) in Outbreak 1 but not in any insects in Outbreak 2. Pigmented plaques were observed in the gut tissues of 8/460 insects (2%) over both outbreaks. In Outbreak 1, 2/258 insects had dark brown to black 1–5 mm plaques at the junction of the foregut and midgut (Fig. 2). One insect in Outbreak 2 had a similar 3 mm plaque at the midgut/hindgut junction, and 3/258 insects in Outbreak 1 had similar, smaller pigmented plaques in the wall of the foregut and/or midgut. Two insects in Outbreak 1 had plaques on the hemocoel lining ventral to the foregut. Gas in the intestine was noted in 14/258 insects (5%) in Outbreak 1 and no insects in Outbreak 2. The foregut was noted to be empty in 9/202 insects (4%) in Outbreak 2, but only 3/258 (1%) of insects in Outbreak 1. In Outbreak 2, while dead animals were often anecdotally described as having a characteristic hunched posture with tightly contracted limbs which were difficult to extend, this finding was only recorded in 2/202 animals (1%) at postmortem, and was also recorded in 3/258 insects (1%) in Outbreak 1. Cloacal prolapse was noted in 7/258 animals (3%) in Outbreak 1 and 1/202 (<1%) in Outbreak 2. The prolapsed material was suggested to include either gut or reproductive tissues, however only 1 of these animals (from Outbreak 2) was submitted for culture and histology.
Gross Observations in Two Outbreaks of Mortality in Lord Howe Island Stick Insects (Dryococelus australis).
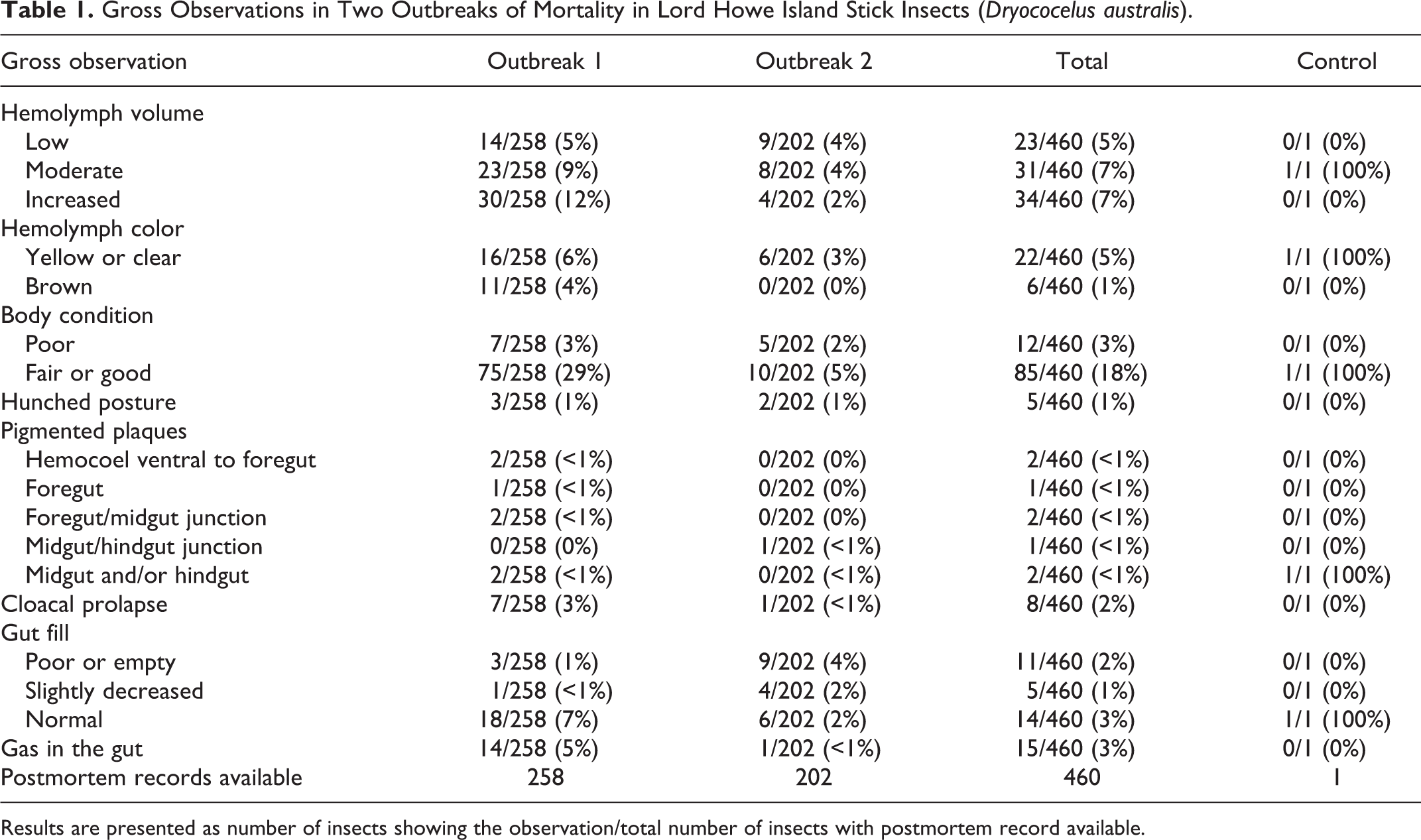
Results are presented as number of insects showing the observation/total number of insects with postmortem record available.
The control animal had grossly evident multifocal brown-black plaques up to 1 mm diameter around the midgut and hindgut, and normal gut fill.
Culture Findings
Serratia marcescens and Pseudomonas aeruginosa were the most commonly cultured bacteria in both outbreaks, with each showing a total prevalence of 20/55 (36%) overall (Tables 2 and 3). Serratia marcescens was identified in a majority of hemolymph samples from Outbreak 1 (15/18, 83%), including 8 as pure growth (44%), or with a single additional species cultured in 7/18 samples (39%). The additional species cultured were Pseudomonas aeruginosa (1/18), Klebsiella oxytoca (2/18), Raoultella ornitholytica (2/18), Morganella morganii (1/18), and an unidentified Gram-positive bacillus (1/18). Serratia marcescens was less common in Outbreak 2, where 5/37 insects (14%) had pure growth, and with either P. aeruginosa or mixed growth in 2 insects. Serratia marcescens cultures were all sensitive to trimethoprim/sulpha, enrofloxacin, marbofloxacin, and gentamicin, and were all resistant to doxycycline, tetracycline, amoxycillin/clavulanic acid, cephalexin, cephalothin, amoxycillin, and cefovecin.
Hemolymph Culture and Hemocyte Lesion Score (LS) in Mortality Outbreak 1 in Captive Lord Howe Island Stick Insects (Dryococelus australis).
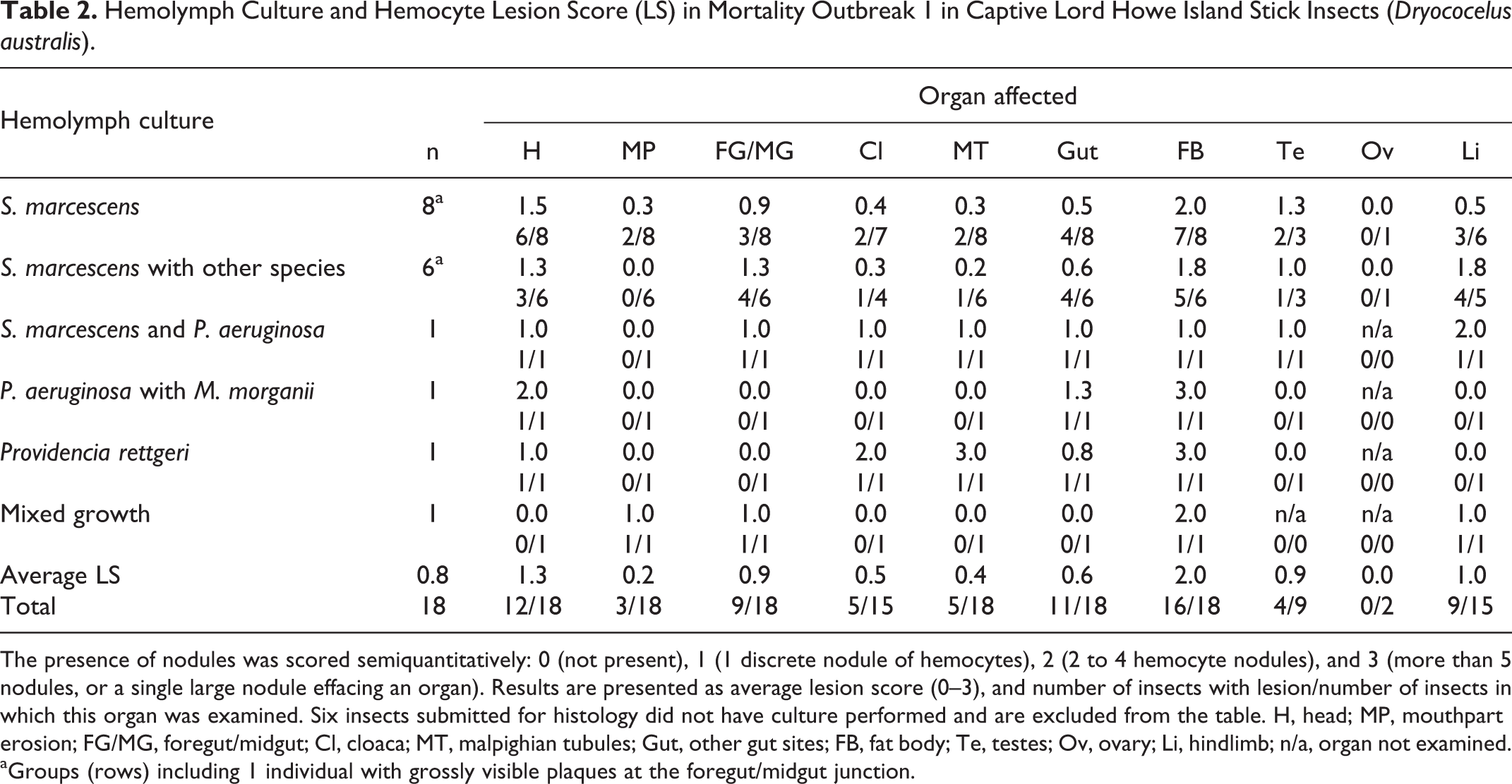
The presence of nodules was scored semiquantitatively: 0 (not present), 1 (1 discrete nodule of hemocytes), 2 (2 to 4 hemocyte nodules), and 3 (more than 5 nodules, or a single large nodule effacing an organ). Results are presented as average lesion score (0–3), and number of insects with lesion/number of insects in which this organ was examined. Six insects submitted for histology did not have culture performed and are excluded from the table. H, head; MP, mouthpart erosion; FG/MG, foregut/midgut; Cl, cloaca; MT, malpighian tubules; Gut, other gut sites; FB, fat body; Te, testes; Ov, ovary; Li, hindlimb; n/a, organ not examined.
aGroups (rows) including 1 individual with grossly visible plaques at the foregut/midgut junction.
Hemolymph Culture and Hemocyte Lesion Score (LS) in Mortality Outbreak 2, and the Control Animal, in Captive Lord Howe Island Stick Insects (Dryococelus australis).
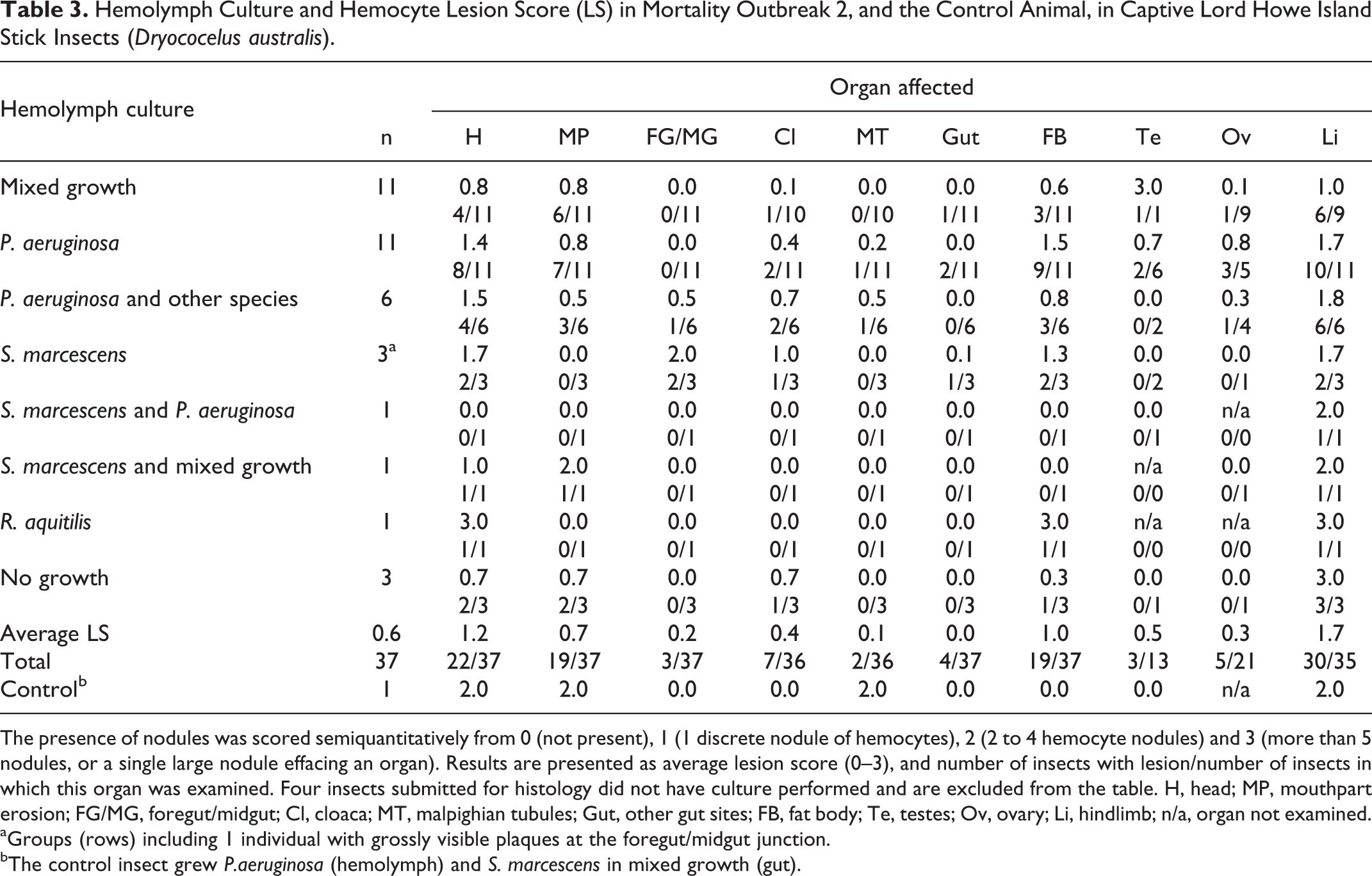
The presence of nodules was scored semiquantitatively from 0 (not present), 1 (1 discrete nodule of hemocytes), 2 (2 to 4 hemocyte nodules) and 3 (more than 5 nodules, or a single large nodule effacing an organ). Results are presented as average lesion score (0–3), and number of insects with lesion/number of insects in which this organ was examined. Four insects submitted for histology did not have culture performed and are excluded from the table. H, head; MP, mouthpart erosion; FG/MG, foregut/midgut; Cl, cloaca; MT, malpighian tubules; Gut, other gut sites; FB, fat body; Te, testes; Ov, ovary; Li, hindlimb; n/a, organ not examined.
aGroups (rows) including 1 individual with grossly visible plaques at the foregut/midgut junction.
bThe control insect grew P.aeruginosa (hemolymph) and S. marcescens in mixed growth (gut).
Pseudomonas aeruginosa was recovered in relatively few hemolymph cultures in Outbreak 1 (2/18, 11%), where it was cultured with S. marcescens or Morganella morganii. It occurred in a higher proportion of insects in Outbreak 2 (18/37, 49%), which included 11/37 (30%) as pure growth, and 7/37 (19%) with 1 (4/37) or 2 (1/37) other species, or with accompanying mixed growth of enteric flora (2/37). Other species cultured with P. aeruginosa in Outbreak 2 were S. marcescens (1/37), M. morganii (1/37), Lactococcus lactis (2/37), and the fungus Mucor spp. (1/37). Pseudomonas aeruginosa cultures were all sensitive to gentamicin, tobramycin, marbofloxacin, ceftazidime, ticarcillin/clavulanic acid, with either resistance or intermediate susceptibility to enrofloxacin.
Gut cultures were only available for Outbreak 1, and grew S. marcescens in 5/7 samples (71%), 4 in pure growth. Pseudomonas aeruginosa was cultured in 3/7 gut samples (43%) in Outbreak 1 with either S. marcescens, Kluyvera cryocrescens or M. morganii.
Fungal culture of hemolymph or gut was performed for all cases which had samples submitted for bacterial culture. Fungal culture was negative in all but 1 adult insect from Outbreak 2, which grew a fungus (Mucor sp.) in combination with P. aeruginosa, had typical hemocyte nodules containing bacteria, and no histological evidence of fungal infection. The Mucor sp. was therefore interpreted as postmortem contamination.
Light to heavy growth of mixed organisms was more common in Outbreak 2 (11/37 insects, 30%) than Outbreak 1 (1/18 insect, 5%). There were 3 animals in Outbreak 2 with no growth on hemolymph culture. In the control animal, hemolymph culture yielded pure growth of P. aeruginosa, and gut culture yielded S. marcescens as a predominant organism in a mixed growth. The swab from crevices within the most affected adult/subadult enclosure yielded a heavy growth of enteric and environmental flora.
Histological Findings
Histological lesions of hemocoelomitis were found in 97% (63/65) of insects from both outbreaks submitted for histology. Lesions were typified by multifocal variably sized aggregates of hemocytes progressing to large confluent granuloma-like nodules which effaced organ architecture. Where they formed nodules, hemocytes were generally large, with pale eosinophilic, agranular cytoplasm. Hemocytes around the periphery of nodules were more often individualized, with cytoplasmic granules or vacuoles. Hemocyte nodules were either unpigmented or contained variable, often abundant brown homogenous pigment, consistent in appearance with melanin. Melanin formed either solid aggregates among hemocyte nuclei, or dense, partial, or complete capsules around central aggregates of short (approximately 1–2 µm length) Gram-negative bacilli. Hemocyte nodules and pigment remained clearly visible even in autolyzed specimens.
Hemocyte lesion score and the results of hemolymph culture are given in Table 2 (Outbreak 1) and Table 3 (Outbreak 2). In Outbreak 1, affected organs included the fat body (16/18), head tissues (12/18), gut (11/18), foregut/midgut junction (9/18), limb tissues (9/15), cloacal hemocoel (5/15), Malpighian tubules (5/18), testis (4/9), and mouthparts (3/18), with the ovary not affected in any case. In Outbreak 2, affected organs included the limb tissues (30/35), head tissues (22/37), fat body (19/37), mouthparts (19/37), cloacal tissues (7/36), ovary (5/21), gut (4/37), foregut/midgut junction (3/37), testis (3/13), and Malpighian tubules (2/36). The highest average lesion scores in Outbreak 1 in order of severity were in the fat body, head and limb (LS 2.0, 1.3, and 1.0, respectively), while in Outbreak 2 the order of severity from highest to lowest was the limb, head and fat body (LS 1.7, 1.2, and 1.0, respectively).
Nodules which occurred at the junction between the foregut and the glandular midgut were particularly striking in size, degree of pigmentation and the extent of tissue destruction (Figs. 3, 4). These lesions were more common and severe in Outbreak 1 (9/18, 50%, average LS 0.9) than Outbreak 2 (3/37, 8%, average LS 0.2). At these sites, the luminal epithelium was necrotic, deeply ulcerated and lined by mixed variably sized Gram-negative and Gram-positive bacilli. The submucosa and tunica muscularis were greatly expanded by coalescing sheets and nodules of pale hemocytes, necrotic debris, and lakes of melanin. Aggregates of short (approximately 1–2 µm length) Gram-negative bacilli were dispersed throughout the lesion, often encased by pigment. Inflammation and pigment deposition continued through the serosa to efface adjacent hemocoel organs, often with embedded aggregates of short Gram-negative bacilli. These lesions were more often detected in longitudinal rather than transverse sections.
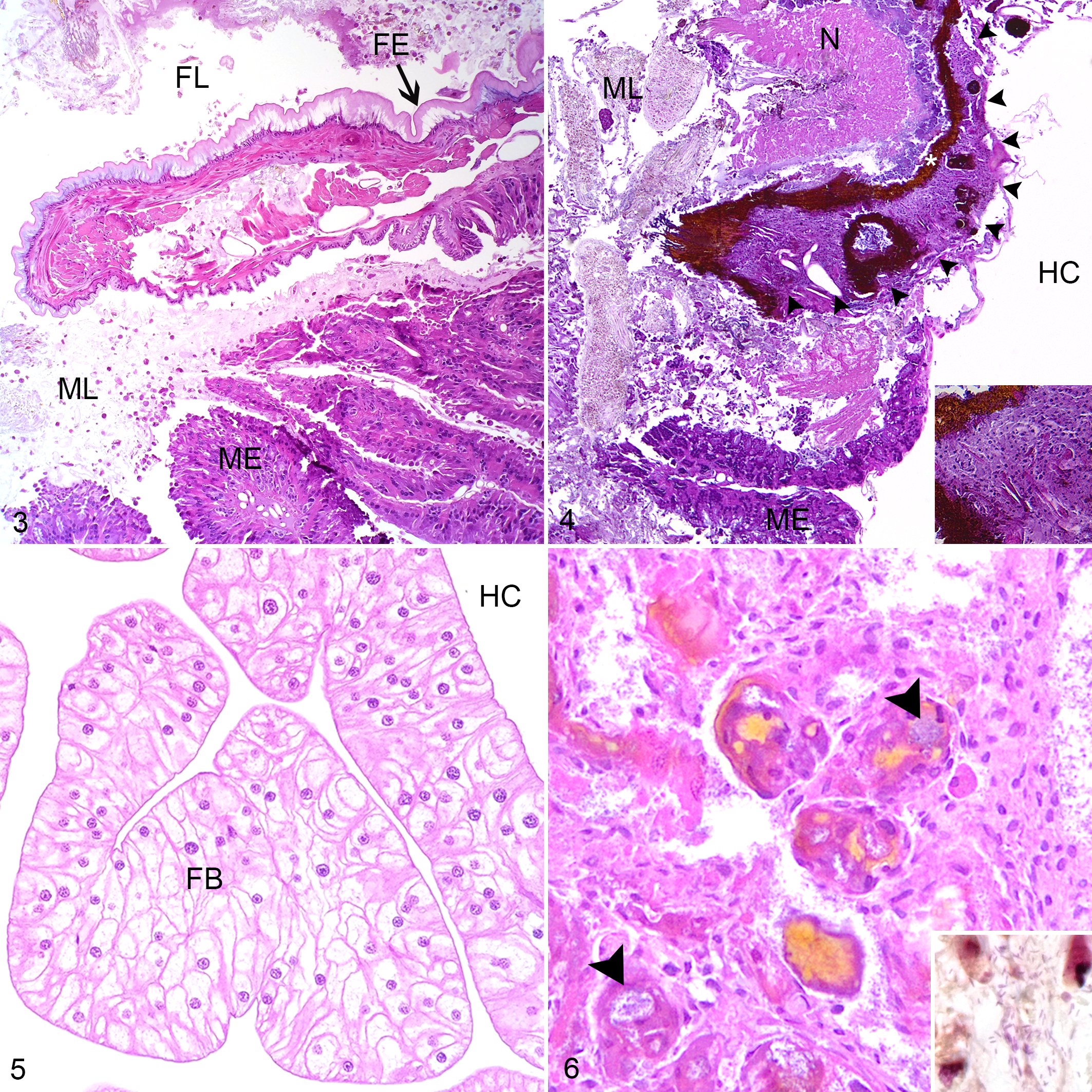
Fat body tissue (Fig. 5) was distributed throughout the abdomen, thorax and head. The fat body was more severely and frequently affected in Outbreak 1(16/18, 89%, average LS 2.0) than Outbreak 2 (19/37, 51%, average LS 1.0). The normal lobular architecture was effaced by hemocytes, with loss of adipocytes and fusion of lobules (Fig. 6). Limb nodules were more frequently seen in Outbreak 2 (30/35, 85%, average LS 1.7) than Outbreak 1 (9/15, 60%, average LS 1.0), and affected either the skeletal muscle fibers, nerves, fat body or appeared free in the limb hemocoel, often concentrated close to the site of attachment of the limb to the body (Figs. 7, 8). Reproductive tract lesions were more common in Outbreak 1, where they affected the testis only (4/9, 44%, average LS 0.9), than in Outbreak 2, where the testis (3/13, 23%, average LS 0.5) or ovary (2/21, 24%, average LS 0.3) were similarly affected. This presented as segmental effacement of tubular genitalia containing spermatozoa (assumed to be seminal vesicles) (Figs. 9, 10), or of the oviduct by pigmented nodules. Nodules found in the head were in the fat body, salivary tissue, and around or within the cranial ganglion (Figs. 11, 12), and were of similar severity between outbreaks (average LS 1.3 in Outbreak 1, 1.2 in Outbreak 2), but more frequently seen in Outbreak 1 (12/18, 67%) than Outbreak 2 (22/37, 59%).
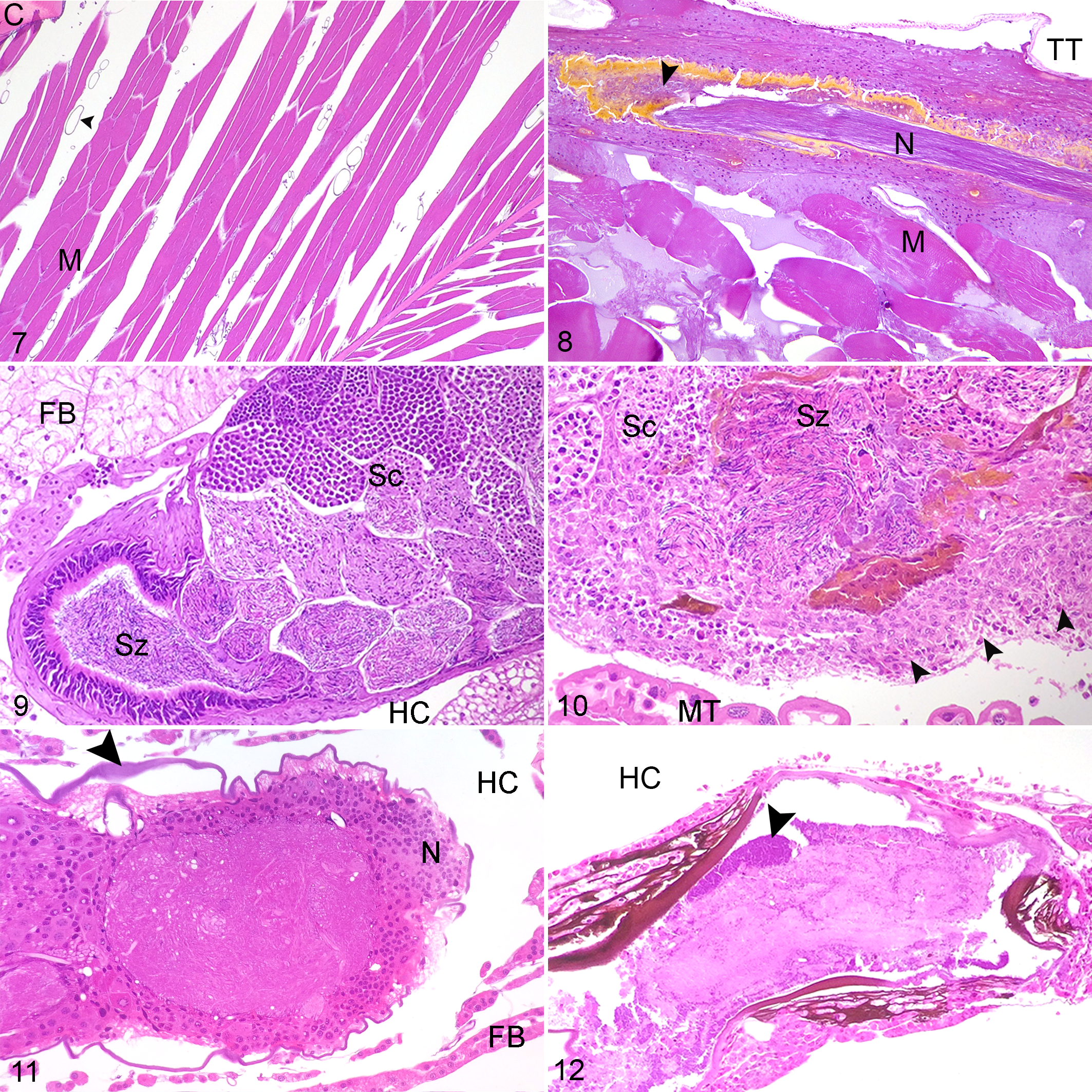
All insects in both outbreaks with Serratia-positive hemolymph cultures whether pure or mixed (20/55) had histological evidence of hemocoel inflammation with short (approximately 1–2 µm length) Gram-negative bacilli, in cases for which Gram stain was performed. The most high-grade lesions were consistently seen at the junction of the foregut and midgut (10/20, 50%, average LS 0.9–2.0), in the head tissues (12/20, 60%, average LS 1.0–1.7), and in the limb (12/20, 60%, average LS 0.5–2.0). Of the insects with grossly noted pigmented plaques at the foregut/midgut junction in both outbreaks, hemolymph culture and histology were available for 3, 2 from Outbreak 1 and 1 from Outbreak 2. Each had high grade hemocyte nodules at this site histologically, and positive hemolymph culture for S. marcescens, 2 in pure growth (1 from each outbreak) and 1 with Klebsiella oxytoca (from Outbreak 1). One insect with gut plaques in Outbreak 1 had only gut culture performed, which yielded a pure growth of S. marcescens.
All insects (20/55) in both outbreaks which cultured P. aeruginosa whether in pure or mixed growth had histological findings of hemocoelomitis with short (approximately 1–2 µm length) Gram-negative bacilli, in cases for which Gram stain was performed. Hemocyte nodules were most frequent and high grade in the limb (18/20, 90%, average LS 1.7–2.0), head (14/20, 70%, average LS 1.0–2.0), and fat body (14/20, 70%, average LS 0.8–3.0). The foregut-midgut junction was affected in 1 of these insects to a mild degree, which yielded S. marcescens in addition to P. aeruginosa from hemolymph culture. The foregut-midgut junction was not affected in any insect with a pure hemolymph growth of P. aeruginosa. The insect in Outbreak 2 noted to have cloacal prolapse had pure growth of P. aeruginosa from hemolymph. The prolapsed material comprised testis with high grade (LS 3.0) inflammation, accompanied by less severe inflammation in the head (LS 1.0) and no inflammation in the gut.
The insect from Outbreak 1 with Providencia rettgeri on hemolymph culture had high grade lesions in the Malpighian tubules (LS 3.0), cloacal tissues (LS 2.0) and fat body (LS 3.0). One juvenile insect from Outbreak 2 cultured Rahnella aquatilis from hemolymph and had high grade hemocyte nodules in the head (LS 3.0), fat body (LS 3.0), and limb (LS 3.0), which contained numerous 4–8 µm diameter round to oval yeast-like bodies, often present within hemocyte cytoplasm, and often heavily melanized. Yeasts stained PAS and GMS positive, and ZN negative. Hemolymph fungal culture was negative.
In insects with only light to heavy mixed growth from hemolymph (12/55, 22%), histological lesions were present in a similar distribution to other insects, with the limb (7/12, 58%, average LS 1.0), fat body (4/12, 33%, average LS 0.6–2.0), and head (4/12, 33%, average LS 0.8) most frequently affected. High grade inflammation was seen in the testis of 1 insect in Outbreak 2 (LS 3.0). A single insect in Outbreak 2 with mixed growth from hemolymph had no histological evidence of hemocoelomitis, and another had only minimal shallow mouthpart erosions without associated hemocoelomitis.
The few insects with no growth from hemolymph culture (3/55, 5%) each had histological evidence of hemocyte nodules containing Gram-negative bacilli, with high grade inflammation in the limb (3/3, 100%, average LS 3.0), and less severe inflammation in the head (2/3, 67%, average LS 0.7), mouthparts (2/3, 67%, average LS 0.7), cloacal tissues (1/3, 33%, average LS 0.7) and fat body (1/3, 33%, average LS 0.3).
The control insect had histological evidence of relatively high grade hemocoelomitis with short (approximately 1–2 µm length) Gram-negative bacilli, with lesions seen in the head (LS 2.0), limb (LS 2.0), mouthparts (LS 2.0), and Malpighian tubules (LS 2.0).
Discussion
This study aimed to document, characterize, and compare the gross, histologic, and microbiological findings in a captive population of critically endangered stick insects investigated for increased mortality. We found that while significant histological and microbiological findings were readily obtained by routine diagnostic methods, there was limited scope to make firm causal associations between specific bacteria and histological lesions.
Comparison of pathologic and microbiologic findings was hindered by the limited recording of gross observations, leading to likely underestimation of the proportion of animals showing gross lesions. Only a minority of the grossly examined insects were submitted for culture and histology, further limiting interpretation of gross lesions. Cloacal prolapse was seen more commonly in Outbreak 1, and could reflect the higher frequency of enteric nodules, possibly indicating straining to defecate or increased gut motility. However, prolapse was also seen in an insect in which there were no gut lesions, and cloacal prolapse could also be a nonspecific finding in a debilitated insect. Hemolymph volume was more often increased in Outbreak 1, but the relatively few animals in which this was assessed in both outbreaks means this could be underestimated. As virtually all insects submitted showed hemocoelomitis, increased hemolymph volume would seem an unreliable gross indicator of inflammation. Similarly, body condition was rarely described as poor despite the presence of often widespread inflammatory disease; this could reflect the difficulty in assessing body condition in insects, but also suggests a relatively short period of clinically significant illness. The gross lesion of pigmented plaques at the foregut-midgut junction was a reliable indicator of high grade inflammation at this site in affected animals.
Serratia marcescens and Pseudomonas aeruginosa were the most commonly recovered bacteria from hemolymph. Both are Gram-negative bacteria considered ubiquitous in the environment and potential opportunistic pathogens of humans, associated with nosocomial infections leading to sepsis, with multiple antibiotic resistance a common feature. 17 Serratia marcescens can be isolated from the intestinal tract of humans and many insect species, and is a known potential pathogen of captive insects including silkworms and Drosophila melanogaster. 12,14,17,18 Likewise, P. aeruginosa is a known potential pathogen of captive insects. 2,9,14 Serratia marcescens has been associated with sepsis potentiated by thermal stress in another captive insect species, while P. aeruginosa-associated insect mortality outbreaks have been associated with high temperature and humidity. 1,14 However, S. marcescens has been identified as normal flora in several phasmid species, and was considered to potentially aid in detoxification of dietary monoterpenes in phasmids consuming Eucalyptus spp. leaves. 22 The normal gut flora of D. australis remains unknown, and the isolation and extremely low population size of the wild population has prevented such investigations to date. Further studies may allow comparisons between wild and captive gut microbiota, and explore their roles as potential opportunists in correlation with husbandry factors.
The majority of insects examined showed histological evidence of hemocyte nodules with entrapped bacteria, consistent with bacterial hemocoelomitis. However, our results indicate that hemolymph culture findings and histological examination alone do not allow confident diagnosis of the cause of bacterial infection in D. australis, given the frequency of mixed hemolymph bacterial culture, the presence of both bacteria in hindgut cultures in Outbreak 1, and the presence of both bacteria in the clinically healthy control insect. While bacteria were clearly seen within hemocyte nodules, the Gram-negative species cultured have a broadly similar size range which precludes histological differentiation based on size. Additional diagnostics to conclusively identify the entrapped bacteria might include laser-capture microdissection from the paraffin blocks, fluorescence in-situ hybridization or immunohistochemistry to localize particular bacterial species within nodules or within hemocyte cytoplasm. In a previous study of bacterial hemocoelomitis, nodules were dissected, washed in insect-buffer saline, crushed and directly streaked on agar to identify the causal bacteria. 8 Lesions were most commonly found in the head tissues, limb and fat body, regardless of culture findings, suggesting these sites as useful samples for histology or ancillary testing in cases of suspected bacterial hemocoelomitis.
As insects were often found dead, it is possible that postmortem leakage and proliferation of gut bacteria within the hemolymph contributed to the bacteria recovered. The recovery of mixed growth from hemolymph in the 2 insects without histological evidence of hemocoelomitis suggests such contamination had occurred. Distal leg joint hemolymph has been suggested as a site that is less likely to be compromised by gut contamination in arachnids, and aspiration of hemolymph from this site should be considered in future investigations. 20 Mixed growth was more common in Outbreak 2. The clinical course was anecdotally longer than in Outbreak 2, with signs observed for up to 24 hours premortem. A longer period of debilitation may have allowed higher frequency of premortem hemocoel bacterial colonization, and could explain the higher proportion of mixed bacterial growths.
Negative hemolymph culture was seen in 3 insects with hemocyte nodules containing bacteria, so cannot be taken as evidence of infection-free status. Negative culture results may be explained by entrapment of bacteria within hemocyte nodules; immunological isolation may have prevented wider hemolymph colonization. This might also in part explain the presence of nodules in the clinically healthy control, and may indicate some degree of tolerance to subclinical bacterial hemocoelomitis in this population.
Although statistically significant associations between bacterial species and lesions were not investigated, some commonalities in lesion pattern were noted. Large, destructive hemocyte plaques at the junction of the foregut and midgut were a common lesion in cases where S. marcescens was cultured, but this site was rarely affected in cases where P. aeruginosa or other bacteria were cultured. All insects from both outbreaks that had grossly visible gut plaques grew S. marcescens. The foregut-midgut junction may be a point of bacterial entry associated with S. marcescens infection, though further investigation would be needed to confirm this. Chitinous cuticle lines the lumen of the fore- and hindgut, but is absent in the midgut, which is lined by a matrix of secreted protein fibrils, proteoglycans and glycoproteins offering physical and immunological protective properties, known as the peritrophic membrane. 16 Serratia marcescens subtypes with increased lethality to insect hosts have been shown to have proteolytic and chitinolytic virulence factors that degrade the peritrophic membrane. 1 The zone of transition from cuticular lining to peritrophic membrane may present a vulnerability to infection which can be exploited by certain bacteria, perhaps potentiated by other as yet unknown factors.
In P. aeruginosa-positive cases, inflammation was common in the limb tissues, perhaps explaining the clinical presentation of ataxia, incoordination and limb contracture. While individual clinical course was not recorded for each insect, it is possible that the anecdotally longer time course in the case of P. aeruginosa infection may have allowed for clinical expression of the limb lesions. However, as gross findings including limb contraction were inconsistently recorded, and muscular lesions were also observed in S. marcescens-positive cases, it is not possible to confirm a difference in clinical presentation attributable to these specific bacteria.
Erosions on the mouthparts were seen more frequently in cases with P. aeruginosa or mixed cultures, most frequently recovered from Outbreak 2. The significance of the mouthpart erosions is unknown, as their occurrence was not associated with increased frequency or severity of hemocyte nodules in the head. Mouthpart damage might suggest difficulty with prehension of food as part of the clinical syndrome, or some effect of insects being stranded on the enclosure floor, perhaps in contract with damp substrate.
Other bacteria were recovered from hemolymph culture in both outbreaks. Providencia rettgeri, Morganella morganii and Rahnella aquatilis are considered ubiquitous Gram-negative bacteria with the potential to cause rare opportunistic infections in a range of species. 19 Providencia rettgeri has been identified as a pathogen of insects, and it is notable that it was recovered in this study in pure growth from an insect with histological lesions of hemocoelomitis. 7 Lesions in that case were located in the head and fat body as in other cases, but also within the cloacal tissues and Malpighian tubules, sites infrequently affected in other insects. If this represents genuine infection, this might indicate different tissue tropism or infection route for P. rettgeri. The Gram-positive coccus Lactococcus lactis was cultured from hemolymph in 2 cases in Outbreak 2, however no Gram-positive cocci were noted in hemocyte nodules, suggesting this was a contaminant.
The histological findings illustrate several features of the insect immune system and suggest a common immune response to bacterial infections. Insects do not have adaptive immunity with inducible antigen-specific defense mechanisms. Rather, they possess complex innate humoral defenses including hemolymph antimicrobial peptides and lysozymes, and cell-mediated defenses including hemocyte phagocytosis and nodulation. 13 Hemocytes are the primary effector cell and several functionally distinct subtypes have been identified which vary between insect groups. 10 Adhesion of hemocytes to a pathogen results in nodulation (entrapment of organisms) and encapsulation (formation of a larger multilayered nodule). To the vertebrate pathologist, these nodules appear analogous in form and function to the vertebrate granuloma, though this analogy is not commonly made in the invertebrate literature. Nodulation is known to involve the action of conserved signaling molecules such as eicosanoids, and also at least 1 novel insect protein known as Noduler. 8 Melanization is induced by activation of a hemolymph prophenyloxidase system by contact with bacterial and fungal components. Melanin creates a physical barrier to invasion and allows pathogen isolation for hemocyte attack, and forms a vital part of the innate immune response in insects. 13 Melanization was a prominent feature in hemocyte nodules in the present study, and was useful in identifying inflammation in more autolyzed specimens.
Hemocyte nodules have been shown to develop within 18 hrs of E. coli inoculation, therefore estimating the age of the lesions in the present study is challenging. 8 The sudden death with a full foregut presentation in Outbreak 1 would suggest a short illness, whereas the presence of observable clinical signs, and poor gut fill in some insects, suggests a more extended illness in Outbreak 2. However, insects in both outbreaks had similar histologic lesion severities, and similar lesions were also present in an apparently healthy control insect. Clinical signs are therefore an unreliable proxy for the presence of hemocoelomitis in this species.
Whether these infections represent opportunistic infection by host-associated bacteria or exposure to pathogenic strains, susceptibility to infection is likely multifactorial, involving host and nonhost variables. This insect population has suffered an extreme genetic bottleneck, being descended from a single breeding pair. Genetic drift and inbreeding depression can lead to loss of heterozygosity which can reduce genetic fitness, though these deleterious effects can be buffered by rapid breeding and larger population sizes. 23 While the breeding program is structured to encourage genetic diversity, further studies are needed to identify any reduction in genetic or immunological diversity in this population compared to the wild insect population or archived specimens. 11 Furthermore, captive stick insects experience potential stressors of captivity, including population densities that may vary from the natural state, human contact including direct handling, pollutants in the urban environment, and a diet and environmental parameters that can only approximate natural conditions. Ambient temperature and humidity variations are known to influence the function of the insect immune response, and have been associated with mortality in laboratory-reared insects. 9,14 In this population, long term studies to identify any effect of temperature on mortality rates are currently being undertaken. The role of environmental conditions in precipitating opportunistic bacterial infection, and the presence or absence of S. marcescens and P. aeruginosa in the wild population, have yet to be determined.
This report presents the clinical, gross, microbiological and histological findings in 2 significant mortality outbreaks in a high conservation-value captive insect population. Insect disease investigation is an unexplored field for most diagnostic veterinary pathologists. The frequency of readily detectable histological lesions makes such diseases appropriate for routine diagnostic laboratory investigation. However, culture results should be interpreted with caution, and analysis of nondiseased control insects is recommended when possible. Adjunct diagnostic methods to identify bacteria within hemocyte nodules should be considered. Further study may provide additional information regarding bacterial pathogenicity and virulence factors, environmental and host factors contributing to infection, and appropriate captive management measures to treat and prevent such mortalities in the future.
Footnotes
Acknowledgements
We thank Melbourne Zoo veterinarians Drs Sarah Frith, Kate Bodley, Helen McCracken, and Megan Curnick for their substantial clinical and postmortem investigations into the mortality events. The dedicated histological processing and microbiological teams at Gribbles Veterinary Pathology provided valuable support and expertise. We are grateful to the invertebrate team at Melbourne Zoo for assistance with colony records and clinical observations.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.