
Abstract
Next generation sequencing (NGS) studies are revealing a diverse microbiota on the skin of dogs. The skin microbiota of canine sterile granulomatous and pyogranulomatous dermatitis (SGPD) has yet to be investigated using NGS techniques. NGS targeting the 16S rRNA and ITS-1 region of bacterial and fungal DNA, respectively, were used to investigate if bacterial and fungal DNA were associated with skin lesions in cases of canine SGPD. The study included 20 formalin-fixed paraffin-embedded (FFPE) skin samples and 12 fresh samples from SGPD-affected dogs, and 10 FFPE and 10 fresh samples from healthy dogs. DNA was extracted from deep dermis and panniculus, and microbial DNA was amplified using primers targeting the bacterial 16S rRNA V1-V3 and fungal ITS-1 regions. The amplified DNA was utilized for NGS on an Illumina MiSeq instrument. The sequences were processed using QIIME. No differences in fungal or bacterial alpha diversity were observed between the SGPD and control samples. Beta diversity analysis demonstrated differences in the bacterial communities between SGPD and control, but not in the fungal communities. Compared to controls, the family Erysipelotrichaceae and genus Staphylococcus were significantly more abundant in the SGPD FFPE samples, and genus Corynebacterium were more abundant in fresh samples. The bacteria found to be more abundant in SGPD are common inhabitants of skin surfaces, and likely secondary contaminants in SGPD cases. This study provides additional evidence that SGPD lesions are likely sterile.
Keywords
Sterile granulomatous and pyogranulomatous dermatitis (SGPD) are skin lesions uncommonly seen in dogs and rarely seen in cats. The etiology is unknown, but a possible cause is immune dysfunction associated with antigenic stimulation, as SGPD lesions often respond to immunosuppressive therapy. 31 Furthermore, it has been speculated that these lesions could be the result of low abundance of infectious organisms or the incomplete clearing of antigens associated with infectious organisms. 5,12,26 Clinically, SGPD lesions are characterized by multifocal nodules mostly affecting haired skin, which occasionally become ulcerated; and histologically, by a large infiltration of macrophages with or without multinucleated giant cells and neutrophils, with a multifocal to coalescing nodular pattern that extends from the dermis to the panniculus. 12,26 A complete work up to rule out infectious agents, including bacteria and fungi, must be performed before starting treatment with steroids. 12,26,27,31 Although the histological presentation is similar to lesions seen in cases of mycobacterial infections and with certain fungal infections, bacterial and fungal culture and special stains are often negative, suggesting these lesions are not associated with an infectious cause, and so, considered to be sterile. 26
A few studies have investigated the presence of infectious agents in SGPD lesions. 5,6,27 A study identified positive results through real-time PCR for Leishmania spp. and Serratia marcescens in skin samples of dogs diagnosed with sterile cutaneous nodular lesions. 5 In addition, Mycobacterium spp. and other partially acid-fast bacteria that have the potential to cause antigenic persistency, have also been investigated with PCR in cases of SGPD, but have not yielded positive results. 5,6
Molecular studies using sequencing of bacterial 16S rRNA gene and fungal ITS region, which is present in all bacteria and fungi, respectively, revealed that the skin surface is inhabited by a highly diverse and variable microbiome that had not been previously demonstrated using culture-based methods. 9,10,13,14 Other studies have further demonstrated that the skin microbiome extends beyond the surface; the dermis and subcutaneous tissues, once considered to be sterile environments, have been shown to have bacterial DNA, although whether this DNA is from living or active bacteria is still unknown. 21 A few studies have used next-generation sequencing (NGS) to describe the canine and feline skin microbiome and to investigate its possible role in skin diseases. 3,18,19,23,24 Through the characterization of these highly complex microbial communities, these studies revealed differences in the bacterial and fungal microbiota between healthy and allergic skin, and demonstrated the importance of NGS to describe a possible correlation between the microbiome and cutaneous allergies.
The development of NGS, which is capable of identifying very small amounts of bacterial or fungal DNA, brings a new opportunity to investigate if an infectious agent could be associated with the development of inflammatory skin lesions in canine SGPD. The presence of fragments of protozoa should also be investigated in canine SGPD lesions, however no fully curated protozoa databases are currently available to be used in NGS studies. The objectives of this study were to investigate if a potential underlying bacterial and/or fungal infection or DNA fragments of these organisms were associated with development of skin lesions in cases of canine SGPD. This is the first study investigating the role of bacterial or fungal pathogens, using NGS, as possible causes associated with SGPD lesions in dogs.
Materials and Methods
Patient Recruitment and Sample Collection
The study was approved by the Texas A&M University Institutional Animal Care and Use Committee. Samples were submitted for diagnostic purpose, and control samples were collected from a group of healthy dogs being euthanized for other purposes. A total of 20 healthy mixed-breed hound dogs with no history of skin diseases were enrolled in this study as control dogs. From these dogs, punch biopsies were collected;10 were stored in formalin and are referred to as formalin fixed paraffin embedded (FFPE) samples, and 10 were stored at –80°C and are referred to as fresh samples. Thirty-two dogs diagnosed with SGPD were recruited, and resulted in 20 FFPE samples and 12 fresh samples. The diagnosis of SGPD was made histologically by the presence of a deep granulomatous/pyogranulomatous inflammation involving the dermis and/or panniculus, with no evidence of any bacterial or fungal microorganisms on Gomori methenamine-silver (GMS), periodic acid–Schiff (PAS), acid fast and Gram stains. In addition, most samples had negative bacterial and fungal cultures, except for 2 fresh (Dogs 21 and 22) and 1 FFPE (Dog 16) samples of affected dogs that had positive cultures for Staphylococcus aureus.
DNA Extraction
From all samples, including control and affected dogs, the epidermis and follicles were sectioned and separated from the deep dermis and panniculus with a sterile blade (Supplemental Fig. S1). The DNA extraction was performed on the sections from the deep dermis and panniculus to avoid amplification and sequencing of commensal microorganisms inhabiting the epidermis and hair follicles. Genomic DNA was isolated from FFPE tissues using the BiOstic FFPE Tissue DNA Isolation Kit (MoBio, Carlsbad, CA, USA), and from fresh tissues using the QIAamp DNA Kit (Qiagen, Gaithersburg, MD, USA), both were extracted following the manufacturer’s protocol. DNA extraction from negative samples (kit reagents and UltraPure DNase/RNase-Free water; Invitrogen, Carlsbad, CA, USA) was also performed and were included as controls. UltraPure water was used as a negative control because it was used in the bath after the FFPE samples had been cut on the microtome.
Sequencing and Analysis of 16S rRNA and ITS
Extracted DNA was submitted to MR DNA Laboratory (Shallowater, TX, USA), for amplification of the V1-V3 region of the 16S rRNA gene from bacteria, a region previously recommended for sequencing of skin samples, 20 and the ITS-1 region of the rRNA coding region from fungi. 8 DNA was amplified using 27Fmod (AGRGTTTGATCMTGGCTCAG) and 519Rmodbio (GWATTACCGCGGCKGCTG) primers targeting the 16S rRNA gene; and ITS1-F (CTTGGTCATTTAGAGGAAGTAA) and ITS2-R (GCTGCGTTCTTCATCGATGC) primers targeting the ITS-1 region of the fungal rRNA. The amplified DNA was utilized for NGS on an Illumina MiSeq instrument (Illumina Inc., San Diego, CA, USA). The raw sequences were processed using QIIME v1.9, where low quality reads (those with N’s or too short after trimming) were excluded during demultiplexing. 4 The operational taxonomic units (OTU) were picked using a 97% identity level and taxonomy was assigned using the Greengenes database for bacterial sequences 7,17,36 and Findley 8 database for fungal sequences. OTU tables were rarefied using the single_rarefaction.py command in Qiime, excluding samples with low sequence counts and achieving an even sequencing depth. Taxa with 1% or higher relative abundance in the negative control samples were considered suspected contaminants and were excluded from the analyses (Supplemental Table S1).
To calculate alpha diversity, which evaluates species richness and evenness within a sample, the Chao1, observed OTUs and Shannon metrics were used. To calculate beta diversity, which evaluates community structure similarity between different sample types, the unweighted and weighted UniFrac 16 (qualitative and quantitative phylogenetic based, respectively) metrics for the bacterial analysis, and the Jaccard and Pearson metrics for fungal analysis were used and the resulting plots viewed using EMPeror. 33 All of the original sequence files are available from the SRA database under the SRA study ID SRP101493, bioproject PRJNA369410.
Statistical Analysis
To evaluate differences in alpha diversity between FFPE and fresh samples from healthy and affected dogs, Kruskal-Wallis tests were performed in JMP Pro 12 (SAS Institute Inc., Cary, NC, USA). P values were corrected for false discovery rate, as previously described. 2 The analysis of similarities (ANOSIM) function in the statistical software package PRIMER 7 (PRIMER-E Ltd., Luton, UK) was used on distance matrices from beta diversity calculations to determine if control and affected dogs had significantly different bacterial and fungal community structures. The filtered relative abundance table was also formatted for linear discriminant analysis (LDA) effect size (LEfSe), 28 with the alpha value set to .05 and the LDA threshold set to 2.5, to identify significant differences in relative taxa abundance between control and affected samples.
Results
Clinical Data
The healthy mixed-breed hound dogs ranged from 1 to 13 years old (average 7.7 years old), and included 11 females and 9 males. None of the dogs presented with skin lesions. The SGPD dogs ranged from 3 to 13 years (average 6.9 years old), and included 21 females and 11 males (Table 1). There were 6 mixed breed dogs, 4 Dachshunds, 3 Labrador Retrievers, 2 Weimaraners, 2 Border Collies, 2 Malteses, 2 Australian Shepherds, 1 English Mastiff, 1 Bichon Frise, 1 Schnauzer, 1 German Shepherd, 1 Greyhound, 1 Boxer, 1 Beagle, 1 Pitbull, 1 Chihuahua, 1 Pekingese, and 1 Border Terrier. From the 32 affected dogs, 30 were from United States (14 from Texas, 4 from Indiana, 3 from New York, 2 from Massachusetts and Colorado, and 1 from Wisconsin, Vermont, Missouri, Illinois, and Connecticut), 1 was from Argentina, and 1 was from Italy. All control dogs were from Texas. The most commonly affected body site was the trunk (24), followed by head (12), neck (10), and limbs (9). The SGPD lesions presented as nodules (22), nodules with draining tracts (7), and plaques (3) (Table 1).
Clinical Information of Dogs With Sterile Granulomatous and Pyogranulomatous Dermatitis Enrolled in This Study.
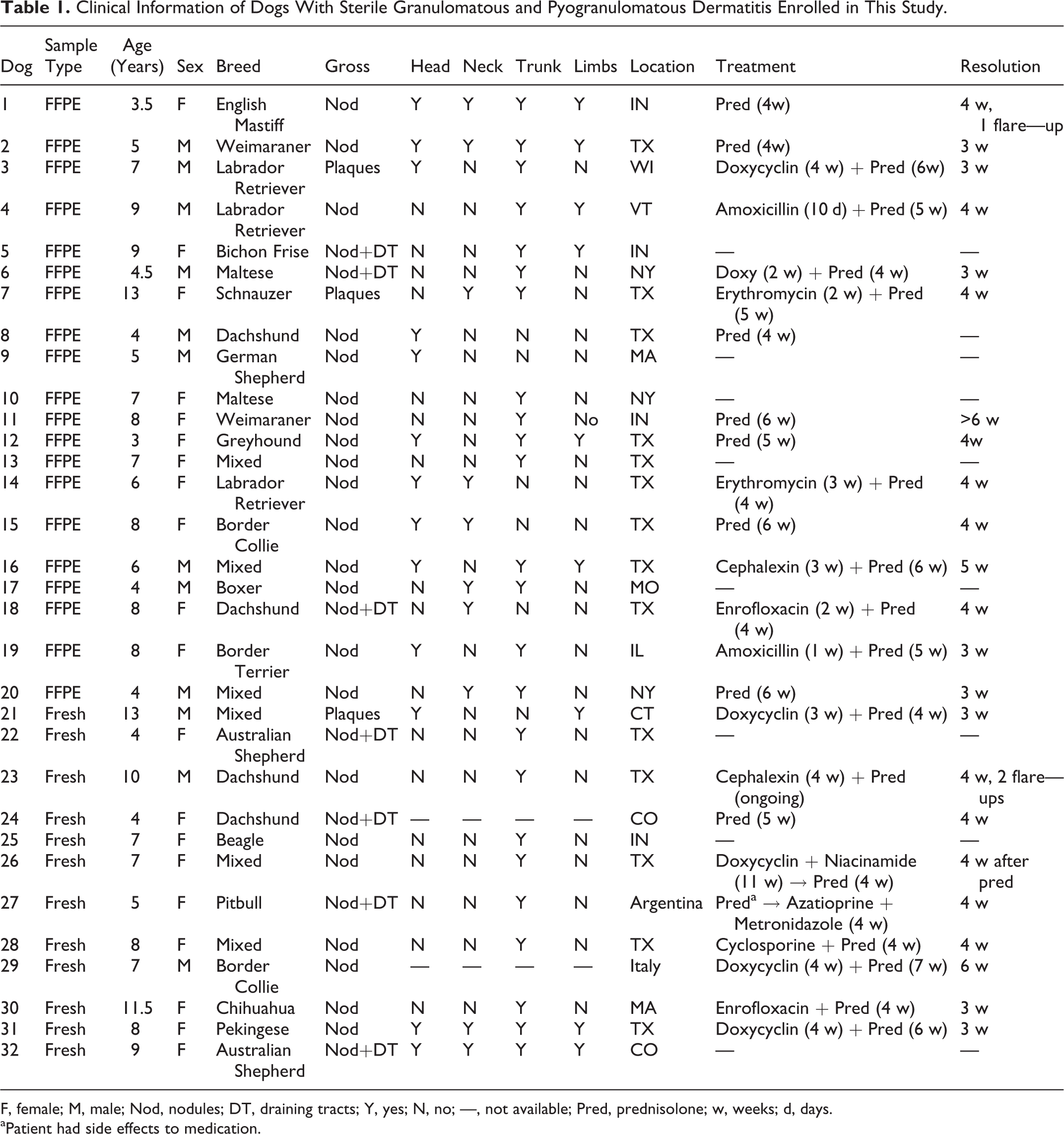
F, female; M, male; Nod, nodules; DT, draining tracts; Y, yes; N, no; —, not available; Pred, prednisolone; w, weeks; d, days. aPatient had side effects to medication.
Clinical follow-up was obtained for 24 SGPD dogs (Table 1). All dogs were treated with prednisolone; 9 dogs had no other therapy and 15 were also treated with doxycline (6), enrofloxacin (2), amoxicillin (2), cephalexin (2), erythromycin (2), or cyclosporine (1). One dog (case 27) developed side effects, and prednisolone was substituted by azathioprine and metronidazole. Lesions resolved in all dogs within 3 to 6 weeks. Relapse episodes occurred in 2 dogs, whereupon lesions were resolved with the reintroduction of steroid therapy.
Bacterial Diversity and Composition
From the 52 samples from healthy and SGPD dogs, from both FFPE and fresh samples, the average number of bacterial sequences amplified per sample was 39966, with a total of 1979 OTUs identified.
The FFPE samples were rarefied to 1400 sequences allowing us to compare 24 samples (10 control dermis, 14 SGPD), while the fresh samples were rarefied to 1100 sequences, allowing us to compare 12 samples (5 control dermis, 7 SGPD). The other samples were excluded from the analysis due to low number of sequences. The alpha diversity (diversity within a sample) was estimated using the observed OTUs metric (which measures the number of OTUs) and the Shannon diversity index (which measures the richness and evenness). There were no differences between control and SGPD (FFPE and fresh) samples with respect to richness and diversity on both alpha diversity metrics (Supplemental Fig. S2).
The beta diversity (the difference in community structure between samples) was estimated using weighted UniFrac (which accounts for relative abundance and phylogenetic relationships of observed organisms) and unweighted UniFrac (which accounts for phylogenetic relationships of observed organisms) phylogenetic metrics. Significant differences in dissimilarity were observed in unweighted UniFrac analysis between fresh SGPD and control samples (R = .241, P = .043), but not FFPE samples (R = –.064, P = .863) (Fig. 1).
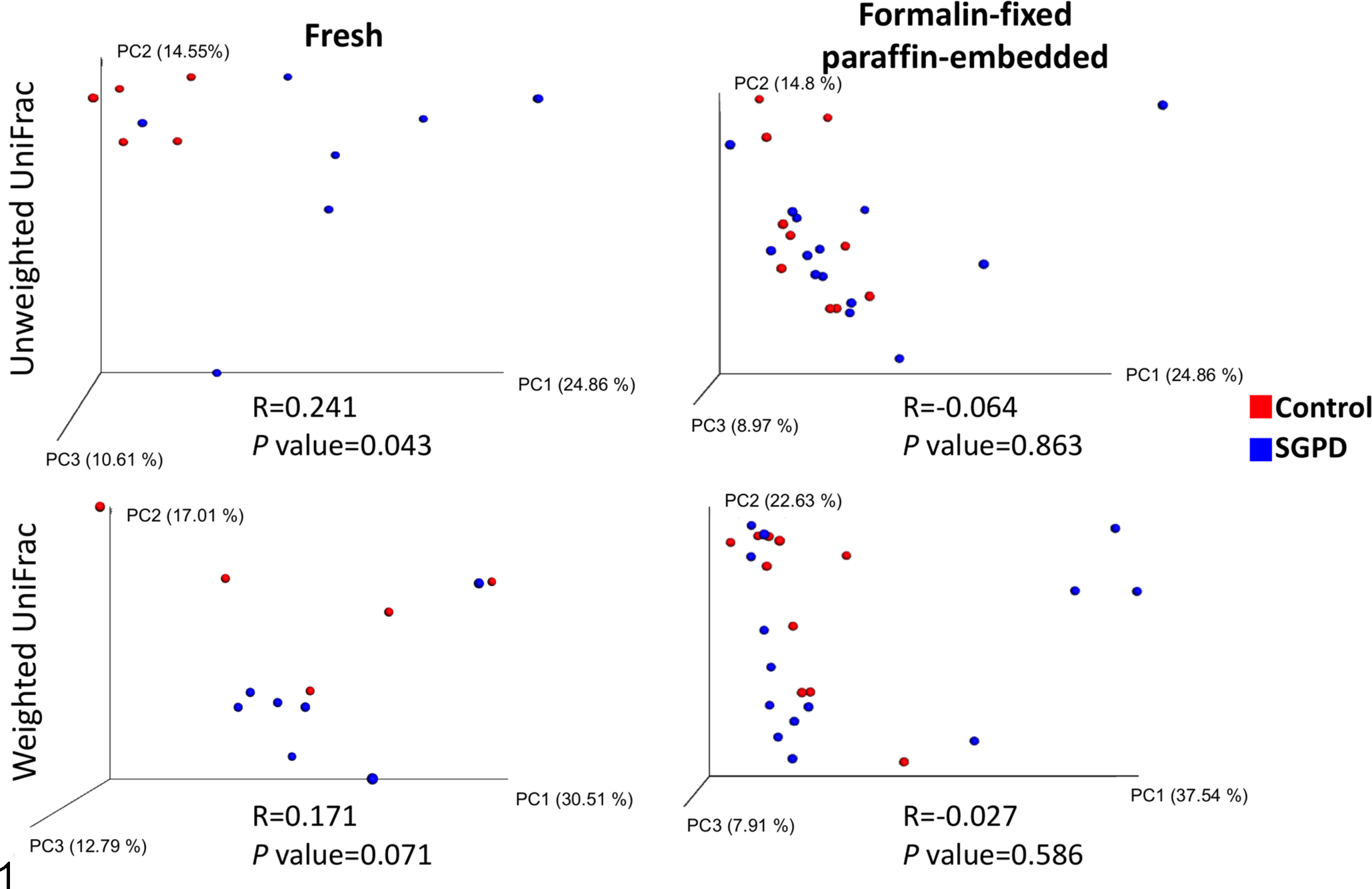
Principal coordinate analysis (PCoA) 3D plots comparing bacterial communities in samples of dermis and panniculus from control dogs (red) and dogs with sterile granulomatous and pyogranulomatous dermatitis (SGPD; blue). PCoA plots allows visualization of the beta diversity results, which reveals patterns of clustering between sample types. Each dot on a PCoA plot represents a single sample. Clustering of dots by sample type (ie, SGPD vs Control) would suggest that community structure is correlated with sample type. A statistical test, analysis of similarities (ANOSIM), is also performed to describe the beta diversity results. R-values closer to 0 indicate that the communities of the different tested sample types are similar whereas R-values closer to 1 indicate dissimilarity. In addition to the R-values, the P values are also important by demonstrating the significance of the R-value. There are significant differences between the bacterial communities in fresh control and SGPD samples based on unweighted UniFrac, a qualitative phylogenetic beta diversity metric (R = .241, P = .043), but not in the quantitative version of this metric, weighted UniFrac (R = .171, P = .071).
The bacterial taxonomic composition was also determined (Fig. 2; Supplemental Table S2). In the fresh control samples, the most abundant phyla were Firmicutes (41.6%), followed by Tenericutes (22.4%), Proteobacteria (16.3%), and Bacteroidetes (15.4%), and the most abundant families were Mycoplasmataceae (22.4%), Planococcaceae (15.9%), and Porphyromonadaceae (10.4%). In the fresh SGPD samples, the most abundant phyla were Firmicutes (65%), Actinobacteria (17%), and Proteobacteria (13.1%), and the most abundant families were Streptococcaceae (21.4%), Enterococcaceae (14%), and Bacillaceae (13.9%). In the control FFPE samples, the most abundant phyla were Proteobacteria (52.4%), Firmicutes (32.7%), and Actinobacteria (8.2%), and the most abundant families were Enterobacteriaceae (10.7%), Xanthomonadaceae (10%), and Planococcaceae (8.7%). The most abundant phyla in SGPD FFPE samples were Firmicutes (47.2%), Proteobacteria (35.8%), Bacteroidetes (16.1%), and the most abundant families were Planococcaceae (16%), Prevotellaceae (15.1%), and Bacillaceae (8.9%) (Fig. 2).
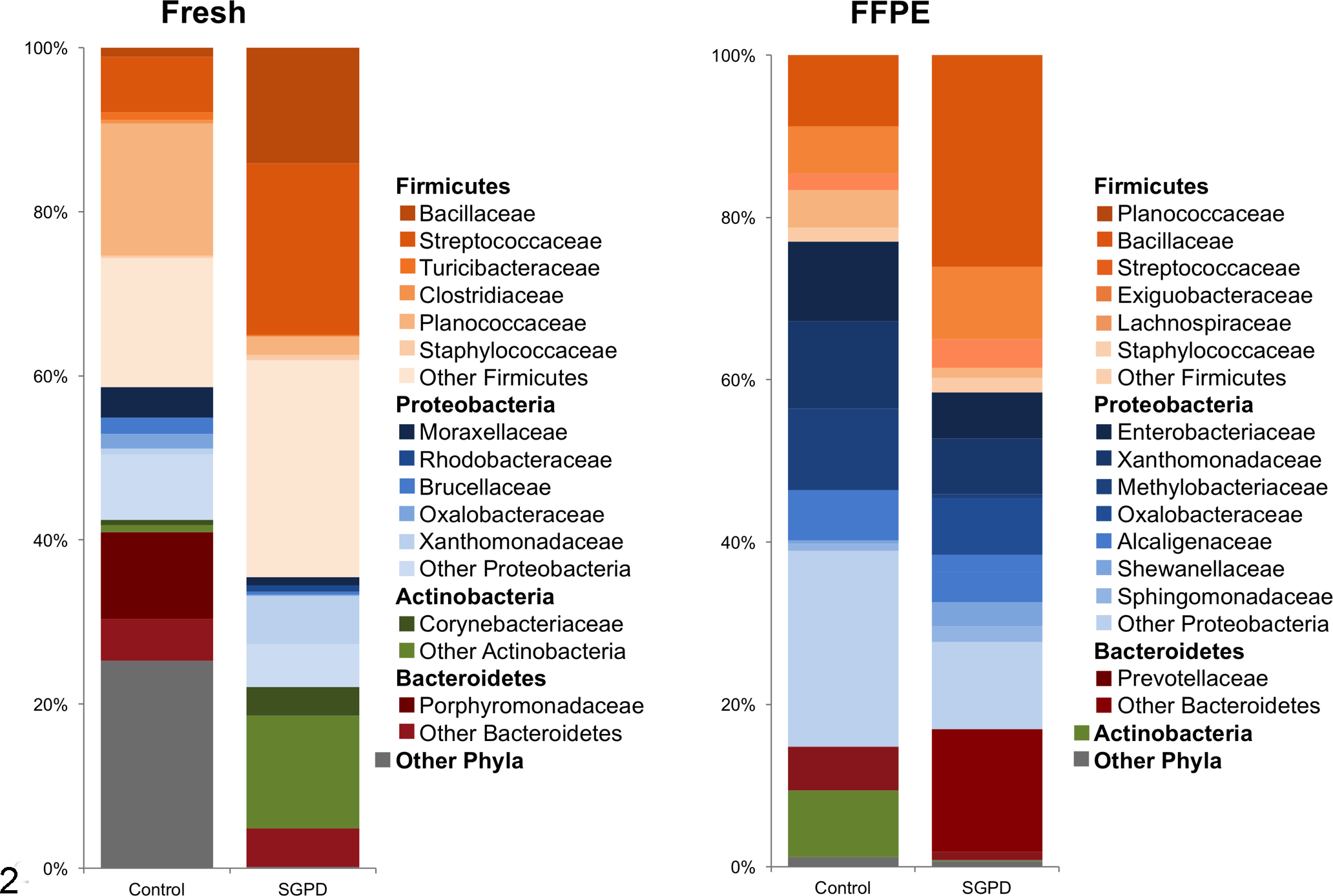
Bacterial phyla and families in samples of dermis and panniculus from control dogs and dogs with sterile granulomatous and pyogranulomatous dermatitis (SGPD). The data show the average relative abundances of the most common bacterial taxa identified in fresh and formalin-fixed paraffin-embedded (FFPE) samples.
Significant differences in the relative abundance of specific taxa between the control dermis and SGPD samples were identified using Kruskal-Wallis tests and LEfSe. Using LEfSe, the family Erysipelotrichaceae and the genus Staphylococcus were significantly more abundant in the FFPE SGPD samples compared to the FFPE control samples. The genus Corynebacterium was significantly more abundant in fresh SGPD samples compared to fresh control samples (Fig. 3).
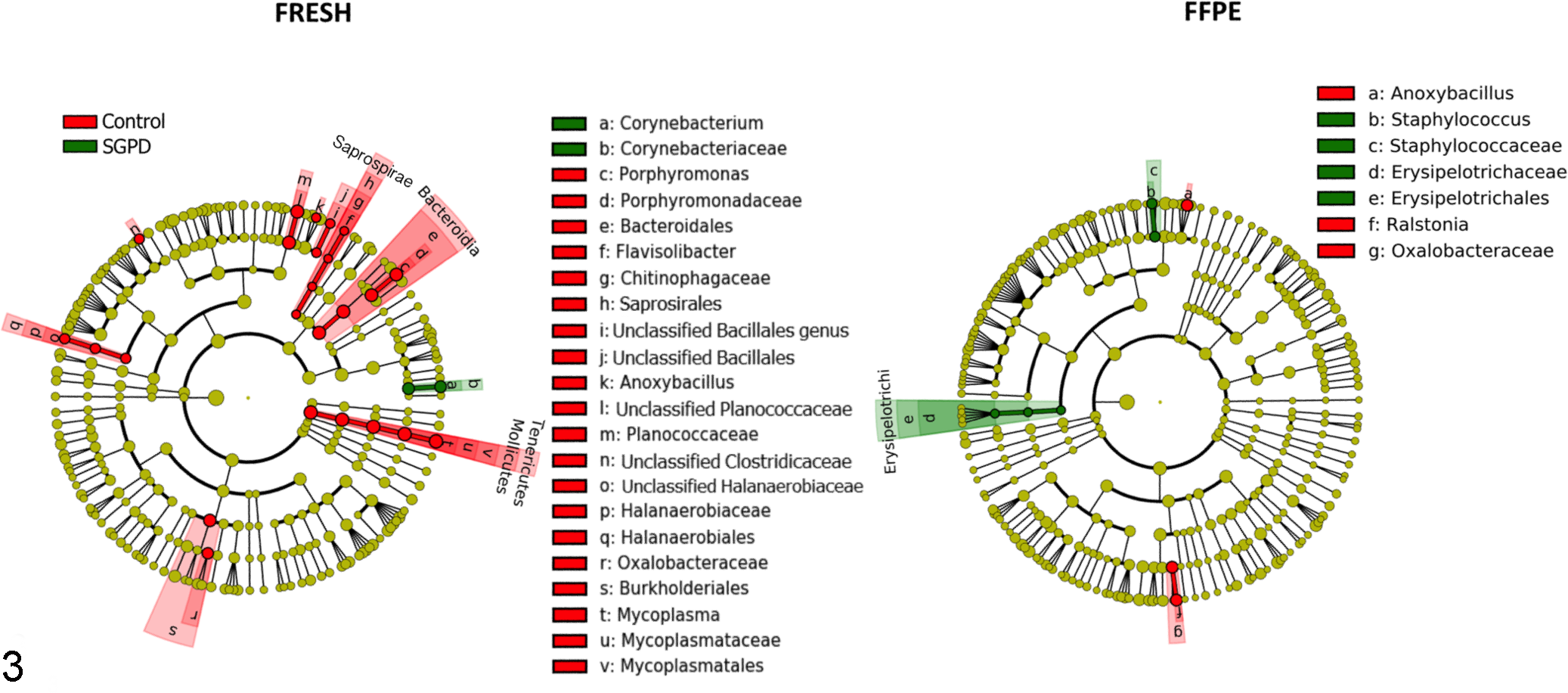
Comparison of bacterial taxa between dermal samples of control dogs and dogs with sterile granulomatous and pyogranulomatous dermatitis (SGPD), in both fresh and formalin-fixed paraffin-embedded (FFPE) samples. Cladograms plotted from linear discriminant analysis effect size (LEfSe) show the taxa that are differentially abundant, arranged by taxonomic levels with phyla in the innermost ring and genera in the outermost ring; each circle is a member within that level. The FFPE SGPD samples had relatively higher numbers of Erysipelotrichaceae and Staphylococcus spp. compared to FFPE controls, and the fresh SGPD samples had relatively higher numbers of Corynebacterium spp. compared to fresh controls.
Fungal Diversity and Composition
From the 52 samples from healthy and SGPD dogs, the average number of fungal sequences amplified per sample was 39223 with a total of 123 OTUs identified. The FFPE samples were rarefied to 1700 sequences allowing us to compare 15 samples (4 control dermis, 11 SGPD). The fresh samples were rarefied to 1700 sequences, allowing us to compare 13 samples (7 control dermis, 6 SGPD). Samples with low number of sequences were excluded from the analysis as aforementioned. Alpha diversity was estimated using the observed OTUs metric and Shannon diversity index; and beta diversity was estimated using Jaccard estimator (which uses the presence and absence of shared fungal taxa between samples) and Pearson distance metric (which accounts for differences in relative abundances of fungal taxa between samples). No significant differences were observed in fungal alpha or beta diversity between control and SGPD samples (FFPE and fresh) (Supplemental Figs. S3 and S4).
The fungal taxonomic composition in fresh and FFPE SGPD and control samples were determined (Fig. 4; Supplemental Table S2). In the fresh control samples the most abundant fungal phylum was Ascomycota (83.3%), and the most abundant families were Capnodiales (55.3%), Polyporales (16.1%), and Saccharomycetaceae (14.6%). In the fresh SGPD samples, the most abundant fungal phylum was Ascomycota (90%), and the most abundant families were Capnodiales (36.1%), Phyllachoraceae (16.2%), Hypocreales (10.8%). The most abundant fungal phylum in FFPE control samples was Ascomycota (58.7%), and the most abundant families were Psathyrellaceae (30.1%) and Pleosporaceae (25.6%). In the FFPE SGPD samples, the most abundant fungal phylum was Ascomycota (79.9%), and the most abundant families were Pleosporaceae (28.7%), Trichocomaceae (24.5%).
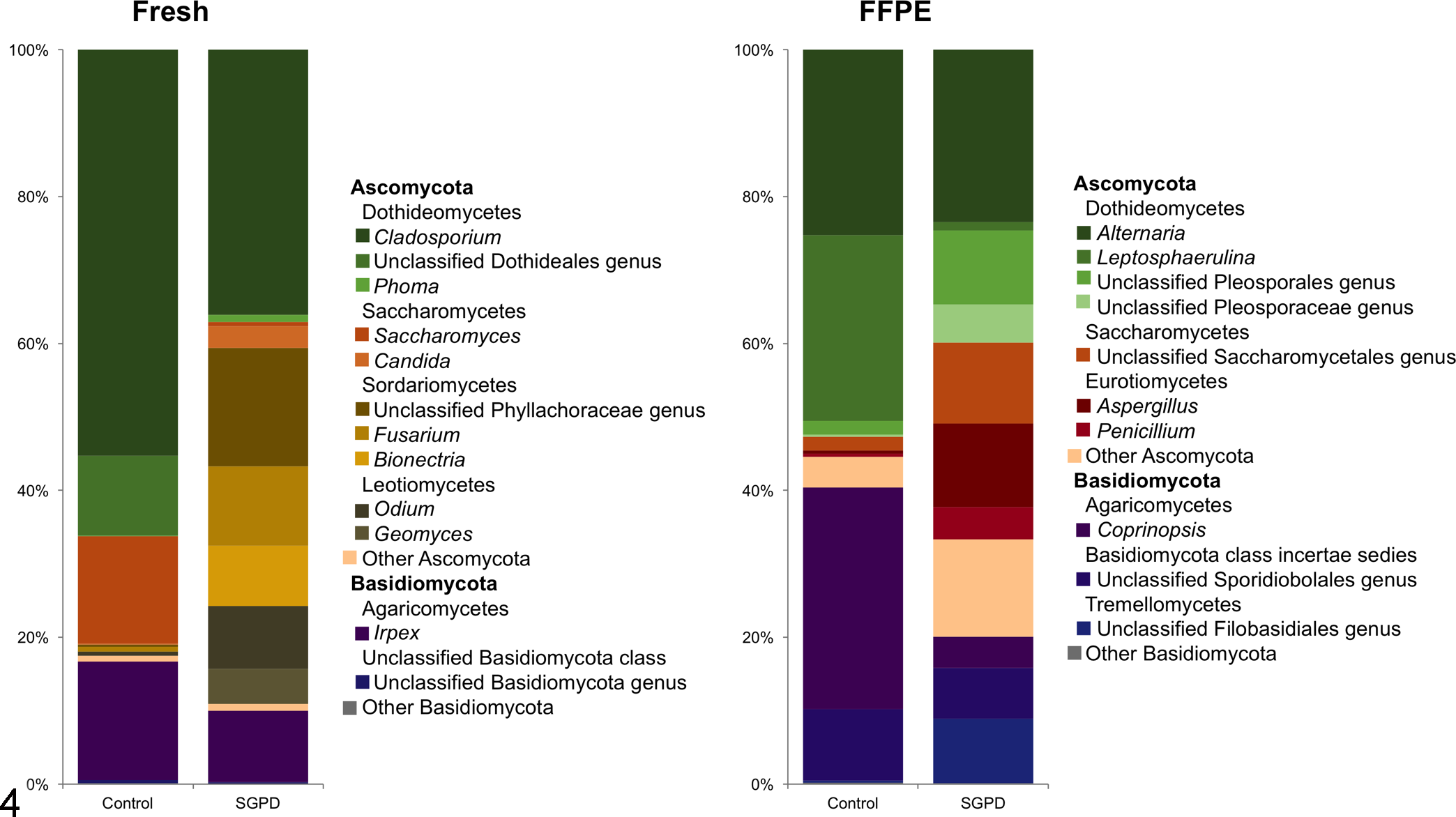
Fungal phyla and families in samples of dermis and panniculus from control dogs and dogs with sterile granulomatous and pyogranulomatous dermatitis (SGPD). The data show the average relative abundances of the most common fungal taxa identified in fresh and formalin-fixed paraffin-embedded (FFPE) samples.
To determine whether specific taxa were differentially abundant between control dermis and SGPD, Kruskal-Wallis tests and LEfSe were used. Neither statistical tests revealed any taxa that were significantly different in relative abundance between control and SGPD (FFPE and fresh) samples.
Discussion
In the present study, NGS targeting the 16S rRNA and ITS-1 region of bacterial and fungal DNA, respectively, were used to investigate if bacterial and fungal infections were associated with development of skin lesions in cases of canine SGPD. We hypothesized that the NGS data would reveal a significant difference in the bacterial or fungal microbiota between control dogs and dogs with SGPD lesions. However, significant differences between the samples of the different groups were only found for organisms commonly identified on skin surfaces of dogs, and no significant differences in abundance of known invasive or novel pathogens were identified in skin lesions of SGPD. Therefore, this study provides evidence that SGPD lesions are likely sterile, as has been previously suggested. 26,31
We found that the genus Corynebacterium was significantly more abundant in fresh SGPD samples compared to fresh controls, whereas the family Erysipelotrichaceae and the genus Staphylococcus were more abundant in FFPE SGPD samples compared to FFPE controls. Bacteria of the genera Corynebacterium and Staphylococcus are commonly present on the skin surfaces of healthy dogs 3,24,34 and humans, 10 where it is found mostly in moist environments. In one study, the genus Corynebacterium was considered to be transient in the skin of dogs, 11 and were also commonly found in the environment including soil, water, and sediment. In addition, Staphylococcus spp. are the most common bacteria isolated from canine hair and hair follicles. 11 Bacteria of the family Erysipelotrichaceae (members of the phylum Firmicutes) are common inhabitants of the skin and mucosal surfaces of healthy dogs, 24 and are commonly found in the canine fecal microbiota. 30 Thus, the bacteria more abundant in SGPD samples are common inhabitants on the skin of dogs. They are presumed to be secondary contaminants in SGPD lesions due to alterations in the epidermal barrier or ulcerations that allow penetration of these organisms into deeper skin layers, and are not likely to be primary causes of SGPD. No significant differences were identified between the fungal organisms amplified with ITS-1 primers from SGPD and control fresh and FFPE samples. Although therapy is a factor that could affect the microbiota, given the low number of cases identified to be enrolled in this study, it was not possible to exclude those patients who had received steroids and/or antibiotics prior to collection of skin biopsies.
One of the limitations of this study was that only bacteria and fungi were investigated as possible underlying causes of SGPD, but other pathogens, such as protozoa, were not investigated in this study. Previous studies identified the presence of Leishmania sp. by immunohistochemistry and PCR in samples of SGPD, 5,6 showing the importance of looking for Leishmania spp. in areas where this disease is endemic. Although the south-central region of Texas is considered to be endemic for Leishmania mexicana, which causes cutaneous lesions in dogs 22 and cats, 32 other Leishmania spp. infections in the United States are considered uncommon, with most cases identified in animals that have travelled to endemic areas in the world. 1,29 The present study aimed to utilize NGS to evaluate only the bacterial and fungal composition in these lesions. At this point, there are no fully available curated protozoa databases that could be used to identify protozoal organisms amplified with universal primers and using NGS, as have been developed for bacteria and fungi. Certainly, these infectious agents should be evaluated in future studies, as they can mimic the clinical and histopathological findings of SGPD.
A challenge in this study was the presence of numerous microbial sequences in the negative and reagents-only control samples. Any taxa with 1% or higher abundance in the negative control samples were excluded from all sample analyses to avoid misinterpretation of the data. The presence of contaminating DNA is a well-known problem and challenge in microbiome studies. Problems with contamination of reagents used in DNA extraction and next-generation sequencing library preparation; 25,35 and contamination with the investigators’ skin, oral, and respiratory microbiota 15 have been well-described; however standardized methods to eliminate these contaminants from the data analysis are not yet well established. One recommendation to overcome problems with low biomass samples is to add negative controls to sequencing runs, which allow identification of sequences originating from contamination, and to distinguish them from those derived from actual constituent taxa. 15,25,35 In this study, sample preparations were performed inside laminar airflow hoods using only sterile and DNA-free reagents, with investigators wearing gowns to avoid DNA contamination. In addition, DNA extractions were performed in a biosafety cabinet. Despite our efforts to minimize contamination in the samples, high numbers of shared OTUs between tissue samples and negative controls, which were extracted and submitted to sequencing concurrently with all the samples, were found.
Another challenge in this study was the use of FFPE samples, which by themselves can be contaminated with microbial DNA during histological processing, or by the process of separating epidermis and hair follicles from the dermis and panniculus. Furthermore, a different DNA extraction kit had to be used for the FFPE samples, which could have resulted in the different microbial abundances identified in FFPE samples versus fresh samples. Despite our efforts to reduce contamination, numerous microbial sequences identified in the control and SGPD skin samples could have come from these sources.
This study revealed that the bacteria (Erysipelotrichaceae, Staphylococcus and Corynebacterium) found to be more abundant in SGPD samples are common inhabitants of the skin surface and unlikely to be the underlying infectious agents of SGPD. Thus, the microbiota analysis performed in this study, together with clinical response of lesions to steroids or a combination of steroids and antibiotics in all dogs, provide further support that SGPD lesions are likely sterile, as has been previously hypothesized. Nevertheless, this is the first study utilizing next-generation sequencing to identify potential bacterial and fungal pathogens in the skin, and additional studies should be developed utilizing molecular techniques targeting specific pathogens to confirm these findings, and to rule out other pathogens not evaluated in this study.
Footnotes
Acknowledgements
The authors would like to acknowledge the dermatologists, general practitioners, and pathologists who contributed to this research study by submitting skin specimens.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
Fabio B. Rosa’s visitor scholar fellowship was supported by the Coordination for the Improvement of Higher Education Personnel CAPES-Brazil, project no. 99999.006281/2015-00.
Supplementary material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.