
Abstract
As in many altricial species, rats are born with fused eyelids and markedly underdeveloped eyes. While the normal histology of the eyes of mature rats is known, the histomorphological changes occurring during postnatal eye development in this species remain incompletely characterized. This study was conducted to describe the postnatal development of ocular structures in Sprague-Dawley (SD) rats during the first month of age using histology and immunohistochemistry (IHC). Both eyes were collected from 51 SD rats at 13 time points between postnatal day (PND)1 and PND30. Histologic examination of hematoxylin and eosin-stained sections was performed, as well as IHC for cleaved-caspase-3 and terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) to evaluate apoptosis, and IHC for Ki-67 and phospho-histone-H3 to evaluate cell proliferation. Extensive ocular tissue remodeling occurred prior to the eyelid opening around PND14 and reflected the interplay between apoptosis and cell proliferation. Apoptosis was particularly remarkable in the maturing subcapsular anterior epithelium of the lens, the inner nuclear and ganglion cell layers of the developing retina, and the Harderian gland, and was involved in the regression of the hyaloid vasculature. Nuclear degradation in the newly formed secondary lens fibers was noteworthy after birth and was associated with TUNEL-positive nuclear remnants lining the lens organelle-free zone. Cell proliferation was marked in the developing retina, cornea, iris, ciliary body and Harderian gland. The rat eye reached histomorphological maturity at PND21 after a rapid phase of morphological changes characterized by the coexistence of cell death and proliferation.
Keywords
The use of neonatal and juvenile animals in preclinical drug toxicity studies is now considered crucial to adequately predict the potential toxic effects of new xenobiotics intended to be administered to children. 73,85 Indeed, it has been shown that pediatric humans and juvenile animals do not respond to drugs in the same manner as adults do, which is in part explained by the incomplete organogenesis in several systems at birth, and for variable timeframes postnatally. 3,24,40 Children and juvenile animals are more susceptible than adults to developing ocular complications, such as cataracts, after exposure to certain drugs. 46,47,91 Thus, the potential for drug toxicity in juvenile patients for a given drug should not be extrapolated from preclinical studies in adult animal models. Despite this knowledge, due to insufficient published data on juvenile drug toxicity, clinicians in the pediatric field must frequently make assumptions based on toxicology data collected from inappropriate life stages. 13,25
An important challenge in preclinical juvenile toxicity studies is the absence of age-matched controls when as unscheduled deaths include found dead and preterminal euthanasia unscheduled deaths occur prior to the end of the study. In addition to having to adequately identify xenobiotic-related lesions, pathologists evaluating juvenile tissues must also recognize developmental abnormalities and must not misinterpret physiological developmental changes as xenobiotic-associated lesions. Therefore, an understanding of the normal postnatal developmental histological features of various organs is required to avoid erroneous conclusions.
Evaluating organs as sophisticated and complex as the eye during juvenile preclinical toxicity studies can be particularly challenging without available age-matched controls. The eye is composed of highly specialized structures characterized by a development that begins during the gestational phase and continues postnatally for variable periods of time depending on the structure and the species. 49 Among laboratory animals, the rat (Rattus norvegicus) is the preferred species for preclinical juvenile studies. As in most other altricial species, the rat is born with closed eyelids and markedly underdeveloped eyes. 42,73 Although various studies have examined the development of specific ocular structures in rats, including the cornea, lens and retina, published information on the key histological features of postnatal development of the whole rat eye at closely spaced time points is sparse. 22,48,51,59,81,96
It is well recognized that organogenesis tightly depends on the interplay of cell division, cell differentiation and programmed cell death, mostly through apoptosis. 32 Despite several investigations of the distribution and the role of those cellular processes during the embryonic ocular morphogenesis in rodents, data about the occurrence of apoptosis and cellular proliferation during the postnatal development of the rat eye remain to be described in detail at different postnatal developmental stages. 10,52,55,58,81 Indeed, as the inhibition of mitosis or the induction of apoptosis are possible undesirable side effects of certain xenobiotics, including in the eye, it is crucial to thoroughly characterize histologically the postnatal occurrence and distribution of those essential cellular events of organogenesis within the different ocular structures to avoid misinterpretation. 26,36,57,74 Thus, this study aimed to provide detailed histological and immunohistochemical data on the postnatal ocular development of the Sprague Dawley rat during the first month of age, with a focus on the occurrence and distribution of cellular proliferation and programed cell death.
Materials and Methods
Animals
This study included 51 Sprague-Dawley (SD) rat pups from 6 time-mated Crl: CD(SD)IGS rat dams obtained from Charles River Laboratories Canada Inc. The pups were divided into 13 groups based on age as follows: postnatal day (PND)1, PND2, PND4, PND6, PND8, PND10, PND14, PND17, PND21, PND24, PND26, PND28, and PND30. Two females and two males were included in each group, except at PND24 (4 females), PND28 (1 female and 2 males), and PND30 (4 males). Rat dams were 9 to 11 weeks old at the start of mating and were housed individually with their pups in a controlled room to maintain a temperature of 21 ± 3°C; a relative humidity of 50 ± 20%; a 12-hour light/12 hour dark cycle; and a ventilation recirculation rate of 10 to 15 complete fresh-air changes hourly. A standard certified commercial chow (Harlan Teklad Certified Global Rodent Diet #2018C) and municipal tap water, treated by exposure to ultraviolet light and purified by reverse osmosis, were provided to the rat pups and dams ad libitum. Pups were separated from the dams at weaning time on PND21. Rat pups were euthanized via exsanguination (blood collection) under general anesthesia with isoflurane. Adult SD rats (n = 4) aged at least 6 weeks were used as a reference for histomorphologically mature ocular structures.
All animal procedures used in this study were performed at CiToxLAB North America Inc. and were approved by the Institutional Animal Care and Use Committee prior to conduct. The care and use of animals was conducted in accordance with the principles outlined in the current Guidelines published by the Canadian Council on Animal Care and the Guide for the Care and Use of Laboratory Animals. All experimental procedures followed standard operating procedures previously approved by the institutional animal care and use committee, and respect the Good Laboratory Practice principles.
Necropsy and General Histology
The eyes and optic nerves of 3 pups per time point ≤ PND10 were fixed in situ in neutral-buffered 10% formalin for 24–48 hours, while the eyes and optic nerves of the fourth pup per time point were removed and fixed in methanol carnoy for 24 hours. For pups aged > PND10 and for adult rat controls, the eyes and optic nerves were removed and fixed in Davidson’s solution for 18–24 hours, followed by fixation in neutral-buffered 10% formalin for 24–48 hours. For each pup, the eyes were collected and put in fixative within 10 minutes after euthanasia. All tissues were embedded in paraffin and sectioned sagittally at 5 μm thick, approximately at the level of the pupil center. Sections were mounted on SuperFrost Plus slides and the slides were dried at 37°C overnight except for slides containing eye tissues that were fixed in Davidson’s solution and intended to be used for the immunohistochemistry (IHC) staining; those slides were dried at 60°C overnight to enhance tissue adherence to the slides. Routine hematoxylin and eosin (HE) staining was used for the histological description.
Immunohistochemistry Staining Protocol
IHC staining was performed against Ki-67 and phospho-histone-H3 (PHH3) using one pup per PND time point or against cleaved-caspase-3 (activated) using two pups per PND time point. Only eyes fixed in neutral-buffered 10% formalin or Davidson’s solution were used. Briefly, sections were deparaffinized, rehydrated in decreasing concentrations of ethanol, and incubated in 0.5% H2O2 in methanol for 20 minutes to quench the endogenous peroxidase activity. Antigen retrieval was performed via heat-induced epitope retrieval using boiling citrate buffer (pH 6.0) for 15 minutes, followed by a cooling step in phosphate-buffered saline (PBS) 1X pH 7.2 for 20 minutes. Nonspecific binding sites were blocked with bovine serum albumin (BSA) (1% in PBS) for 30 minutes at room temperature. Slides were then incubated with the primary antibody, overnight at 4°C for the antibody against cleaved caspase-3 (applied at 0.315 m g/ml, #9661 from Cell Signaling Technology, Danvers, MA, USA), or for 1 h at room temperature for the following primary antibodies: PHH3 antibody Sc-8656-R (applied at 0.143 μg/ml, from Santa Cruz Biotechnology, Dallas, Texas, USA) or Ki-67 antibody ab15580 (applied at 0.909 μg/ml, from Abcam, Toronto, Ontario, Canada). Tissue sections were then incubated for 30 minutes at room temperature with a horseradish peroxidase-labelled polymer antirabbit antibody (K4002, from DAKO, Mississauga, Ontario, Canada), which was followed by color development using a commercial DAB substrate kit (DAB Quanto Chromogen and Substrate # TA-125-QHDX, Thermos Fisher Scientific). Counterstaining was performed with Mayer’s hematoxylin.
Immunohistochemistry Controls
IHC staining controls included a negative control performed by omitting the primary antibody and an isotype control. The latter consisted of substituting the primary antibody with the appropriate isotype-matched antibodies for all three primary antibodies; Jackson ImmunoResearch’s ChromPure Rabbit IgG catalog # 011-000-003 was used as the isotype control for caspase-3 and normal rabbit IgG Ig catalogue # sc-2027 from Santa Cruz Biotechnology was used as the isotype control for PHH3 and Ki-67. Positive tissue controls were performed on the appropriate rat tissues that had been fixed and processed following the same methods as for the rat pup eyes. Rat thymus was used as the positive tissue control for caspase-3, while small intestine containing Peyer’s patches and mesenteric lymph nodes were used as positive tissue controls for Ki-67 and PHH3. Negative and isotype controls resulted in a complete absence of labeling. The positive tissue control for caspase-3 resulted in positively stained apoptotic cells and apoptotic bodies in the thymus and the positive tissue controls for Ki-67 and PHH3 showed positively stained dividing cells in intestinal crypts and Peyer’s patches germinal centers.
TUNEL Histochemistry Protocol
TUNEL was performed using the ApopTag® Peroxidase In Situ Apoptosis Detection Kit (S7100, Millipore) following the manufacturer’s instructions on the same two pups per PND time point as the ones used for caspase-3 IHC staining. Only eyes fixed in 10% neutral-buffered formalin or Davidson’s solution were used. Briefly, sections were deparaffinized, rehydrated in decreasing concentrations of ethanol, and treated with 20 μg/ml proteinase K (P6556, Sigma-Aldrich, Oakville, Ontario, Canada) for 15 minutes at room temperature to allow a better accessibility to the DNA breaks in apoptotic cells or apoptotic bodies. Endogenous peroxidase was then quenched with 3% H2O2 (30% H2O2 diluted in PBS 1X pH 7.2) for 5 minutes at room temperature. After incubation with the equilibration buffer, tissue sections were incubated with freshly mixed digoxigenin-conjugated nucleotide triphosphates (NTPs) and Terminal deoxynucleotidyl transferase (TdT) enzymes at 37°C for 1 hour. The TdT enzyme reaction was terminated with the Stop/Wash buffer provided in the kit. Then, a peroxidase-conjugated antidigoxigenin antibody was added to the tissue sections for 30 minutes at room temperature and the reaction was visualized with the peroxidase chromogenic substrate 3,3-diaminobenzidine (DAB) (DAB Quanto Chromogen and Substrate # TA-125-QHDX, thermos scientific) for 4 minutes at room temperature. Counterstaining was performed with Mayer’s hematoxylin for 30 seconds at room temperature. Negative control was performed by substituting the TdT enzyme with PBS 1X pH 7.2 and resulted in a complete absence of staining. A positive tissue control was performed using rat thymus fixed in Davidson’s solution or 10% neutral buffered formalin, as appropriate.
Preliminary Standardization of Immunohistochemistry and Histochemistry
As rat pup eyes were fixed using different methods depending on the age, preliminary IHC and TUNEL histochemistry standardization assays were performed to establish if the IHC and TUNEL staining pattern was comparable between fixation methods. Therefore, for each IHC antibody and for TUNEL histochemistry, the appropriate positive tissue control was collected from a spare SD rat, divided in two portions and each portion was fixed either in neutral-buffered 10% formalin or in Davidson’s solution, following the same protocol as for the fixation of the rat pup’s eyes. Then, the two differently fixed positive tissue controls were simultaneously stained for IHC and TUNEL following the same protocols used for the rat pup’s eyes. TUNEL histochemistry and IHC staining against caspase-3 and PHH3 resulted in similar staining patterns regardless of the fixation method. However, Ki-67-positive cells appeared less numerous in Davidson’s-fixed positive control tissues (small intestine) compared to the same tissue fixed with neutral-buffered 10% formalin. Therefore, for rat pups aged >10 PND, IHC staining using the antibody against PHH3 was used as the main method to identify cell proliferation, in addition to HE-stained slides. For rat pups aged ≤ PND10, IHC against Ki-67 and PHH3 were used to evaluate cellular proliferation, in addition to the HE-stained slides.
Histology, IHC, and TUNEL Interpretation
The histological evaluation was performed qualitatively using both eyes from all 51 rat pups. For the evaluation of the cornea, the epithelium was divided into central, peripheral, and limbal histological regions as described elsewhere. 17 Furthermore, in this study, the term ora ciliaris retinae was used to refer to the junction between the retina and the ciliary body, as in rodents this region does not have a serrated appearance (as opposed to humans and primates, in which the term ora serrata is used). 35
IHC staining against Ki-67 and PHH3 were performed to detect, respectively, cells engaged in any steps of the cell cycle or cells exclusively in the mitotic phase. Ki-67 is a nuclear protein that is expressed in all stages of the cell cycle, except in the G0 phase when cells are quiescent. Its expression allows identification of not only mitotically active cells, but also cells in the interphase that are engaged in other cellular activities such as DNA synthesis and growth. 28,77 Furthermore, the localization of Ki-67 in the nucleus varies according to the cell cycle phase: during G1, Ki-67 is present diffusely in the nucleoplasm, while during the S phase, Ki-67 relocates to the nucleolus. During the prophase, Ki-67 becomes closely associated with the condensing chromatin and then remains at the periphery of the chromosomes during the different steps of mitosis. During telophase, Ki-67 can be found in the cytoplasm because of the fragmentation of the nuclear membrane occurring during mitosis. 28 The phosphorylation of histone H3, resulting in PHH3, occurs specifically during the mitotic chromatin condensation in the late G2 and the mitotic phases and, therefore, is detected exclusively in dividing cells. 38
IHC staining against cleaved-caspase-3 and TUNEL histochemistry were performed on consecutive sections to improve the sensitivity and specificity of detecting apoptosis. Caspase-3, when cleaved (activated), is the main executioner caspase in the apoptotic caspase cascade and is involved in the dismantling of most cellular substrates during apoptosis. 39 TUNEL histochemistry detects DNA fragmentation, one of the downstream cellular degradation events occurring during apoptosis. 27
In this study, cellular proliferation was considered present if mitotic figures were observed on HE-stained sections and if mitotically active cells immunopositive for Ki-67 and/or PHH3 were also noted. Apoptosis was considered to occur if apoptotic bodies or apoptotic cells were detected on HE-stained sections and if both cleaved-caspase-3 IHC staining and TUNEL histochemistry were positive.
Corneal Endothelium Scanning Electron Microscopy
Since primary cilia have been shown to be temporarily assembled on the apical surface of corneal endothelial cells during the first month of age in mice, we assessed with SEM the presence of such structures in rats. 6 As only paraffin blocks of ocular tissues were available, the blocks from one rat pup each at PND1, PND6, PND14, PND21, and PND30 were melted, the eyes were deparaffinized in xylene, the corneas were dissected under a binocular stereo-microscope, and the samples were kept in 100% ETOH until scanning electron microscopy (SEM) was performed. For sample preparation, ETOH was gradually substituted by amyl acetate, samples were dried in a Critical Point Dryer (Polaron E3000) and the samples were coated with gold (Edwards S150B) for 2 minutes at 1 kV, 20 mA. Evaluation of the limbal, peripheral and central regions of the corneal endothelium surface was conducted using a SU8010 Hitachi SEM.
Results
Detailed descriptions of the histological and immunohistochemical changes during postnatal ocular development of the SD rat are presented. Tables 1 and 2 summarize the PND intervals when cellular proliferation, characterized by the presence of immunopositive cells for Ki-67 and PHH3, and/or apoptosis, characterized by the presence of positive cells for caspase-3 and TUNEL histochemistry, are observed in specific ocular structures. The PND when the histomorphological maturity is reached for each ocular structure is shown in Table 3. Note that the eyes of SD rats are nonpigmented.
Age Ranges When Specific Ocular Structures of the Sprague-Dawley Rat Presented Cellular Proliferation and/or Apoptosis (Part 1).a
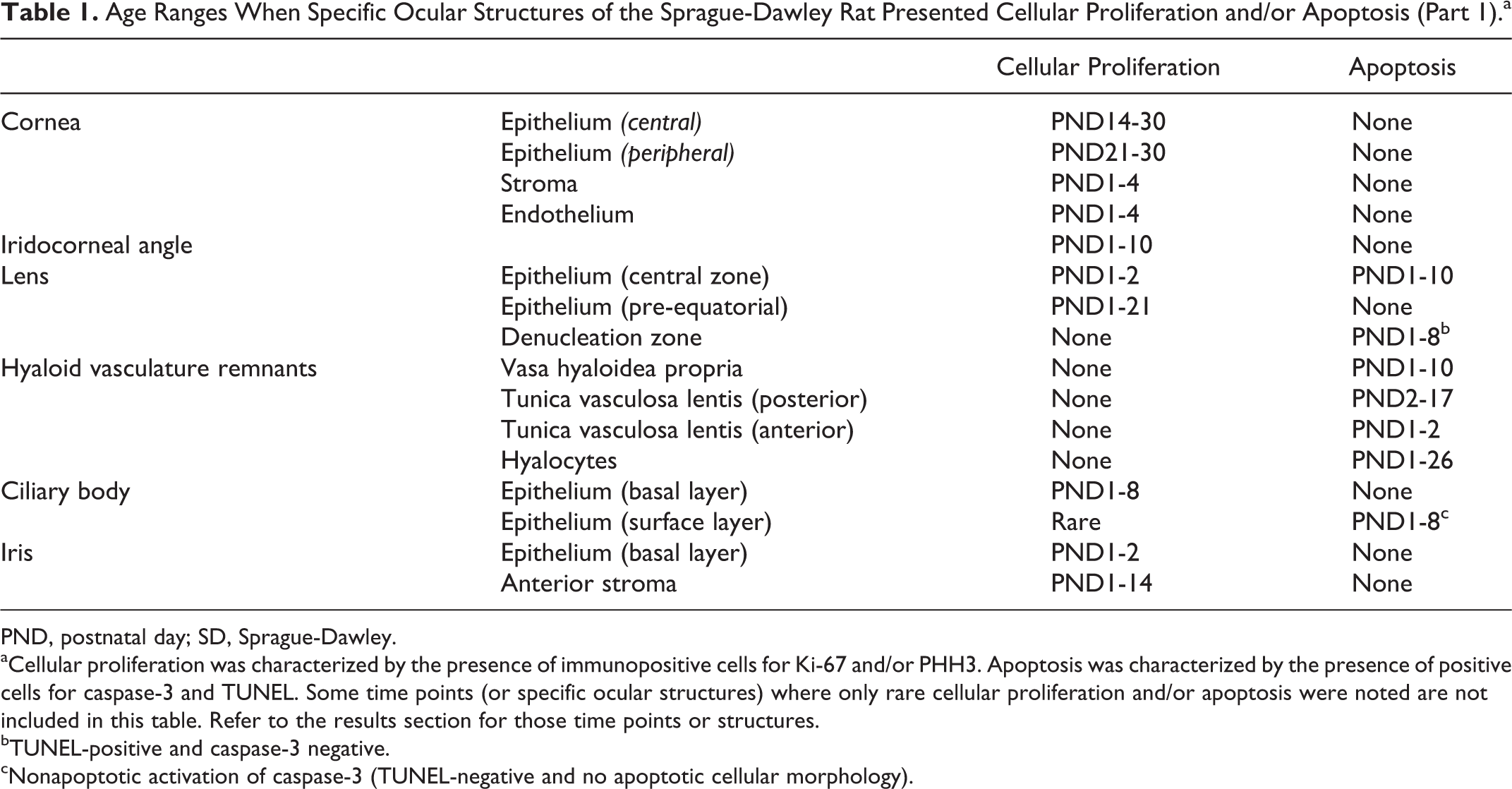
PND, postnatal day; SD, Sprague-Dawley.
aCellular proliferation was characterized by the presence of immunopositive cells for Ki-67 and/or PHH3. Apoptosis was characterized by the presence of positive cells for caspase-3 and TUNEL. Some time points (or specific ocular structures) where only rare cellular proliferation and/or apoptosis were noted are not included in this table. Refer to the results section for those time points or structures.
bTUNEL-positive and caspase-3 negative.
cNonapoptotic activation of caspase-3 (TUNEL-negative and no apoptotic cellular morphology).
Age Ranges When Specific Ocular Structures of the Sprague-Dawley Rat Presented Cellular Proliferation and/or Apoptosis (Part 2).a
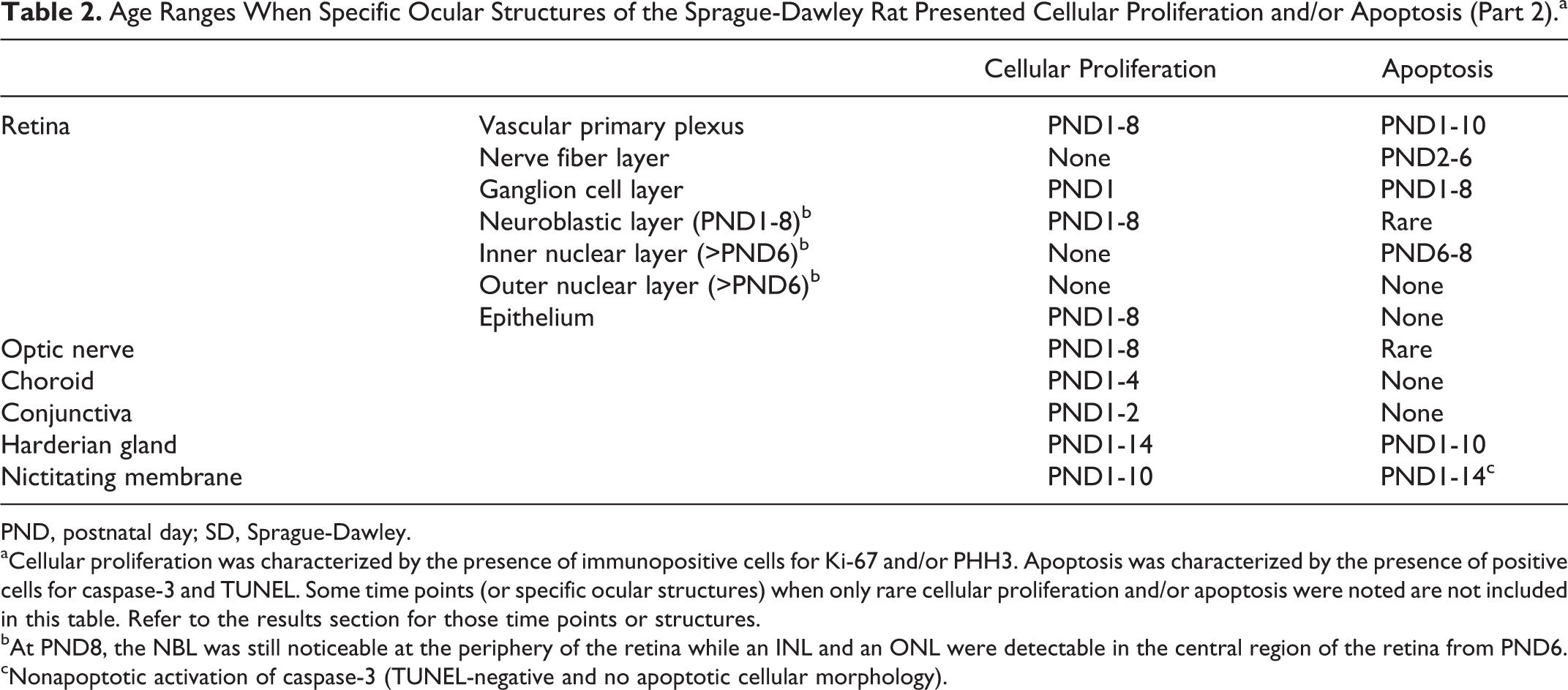
PND, postnatal day; SD, Sprague-Dawley.
aCellular proliferation was characterized by the presence of immunopositive cells for Ki-67 and/or PHH3. Apoptosis was characterized by the presence of positive cells for caspase-3 and TUNEL. Some time points (or specific ocular structures) when only rare cellular proliferation and/or apoptosis were noted are not included in this table. Refer to the results section for those time points or structures.
bAt PND8, the NBL was still noticeable at the periphery of the retina while an INL and an ONL were detectable in the central region of the retina from PND6.
cNonapoptotic activation of caspase-3 (TUNEL-negative and no apoptotic cellular morphology).
Age of Histomorphological Maturity (or Complete Regression) of Various Ocular Structures in the Sprague-Dawley Rat.
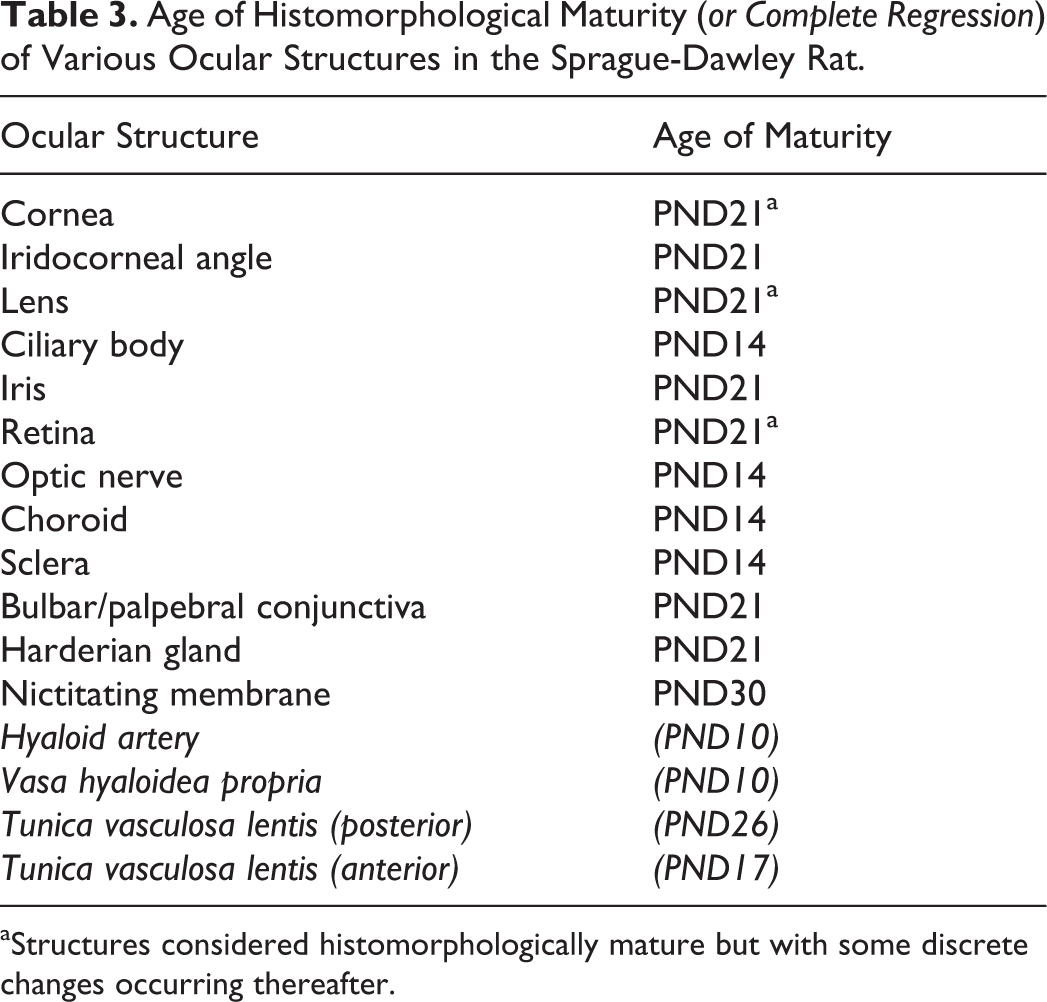
aStructures considered histomorphologically mature but with some discrete changes occurring thereafter.
Cornea
Epithelium
At PND1, the corneal epithelium was composed of a monolayer of flattened to oval cells in all the corneal regions (Fig. 1). Between PND2 and PND6, scattered squamous cells were occasionally forming a second cell layer and at PND8, a two-cell layer was almost completely present in all corneal regions. At this stage, basal cells began to differentiate into cuboidal epithelial cells in the central region and at PND10, cuboidal basal epithelial cells extended to the peripheral cornea while a third layer of rounded cells was present in the central region. Immediately following the opening of the eyelids at PND14, the corneal epithelium transformed to a nonkeratinized stratified squamous epithelium composed of 3–5 cell layers, the central and peripheral regions being more stratified (4–5 layers) than the limbal region (3 layers). From PND14 to PND30, the corneal epithelium gradually thickened to reach 6–7 cell layers in the central and peripheral regions, basal cells became columnar in those same regions, and superficial cells further flattened (Fig. 2).
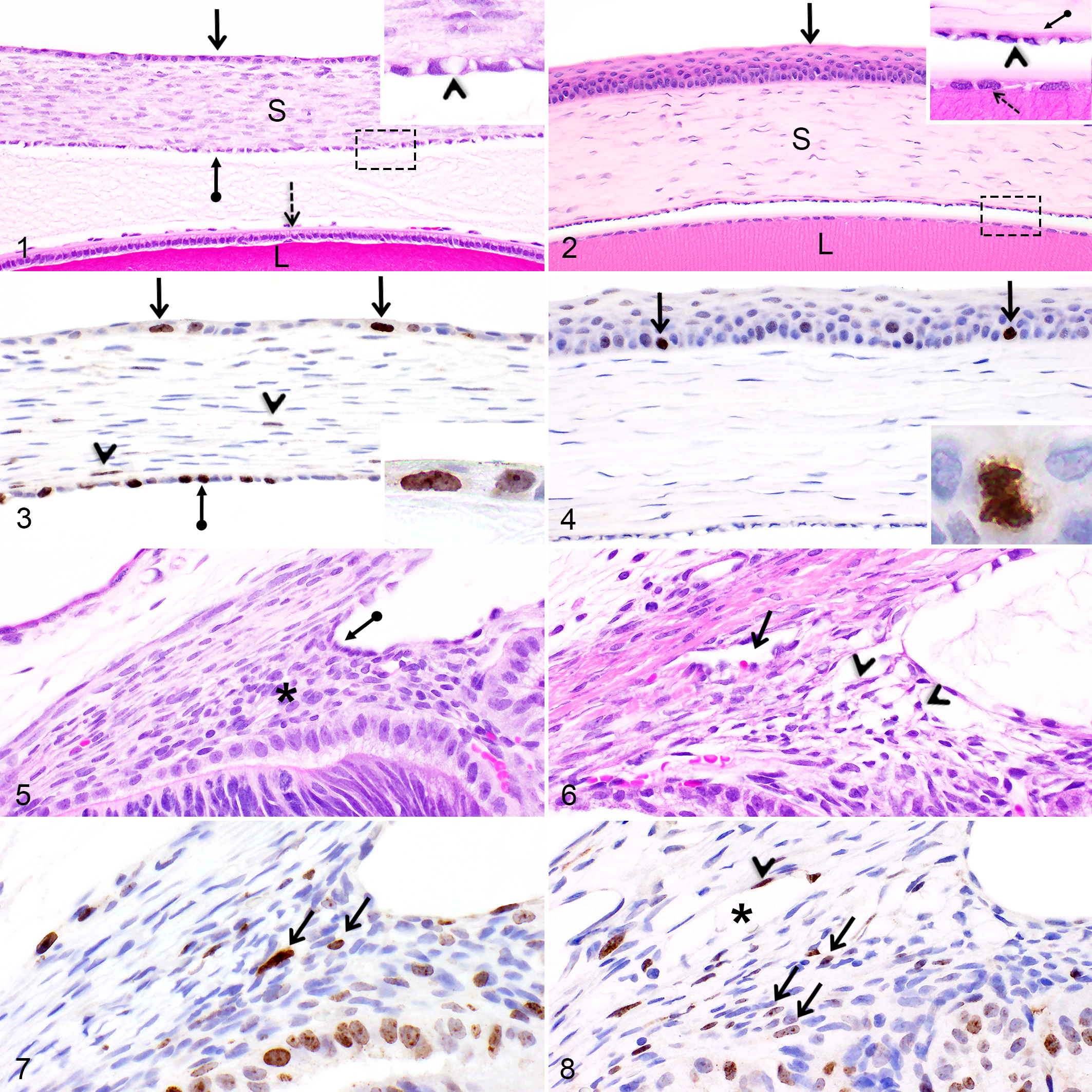
Before the opening of the eyelids, some basal epithelial cells in all 3 corneal regions showed intranuclear Ki-67 and PHH3 staining; some immunopositive cells corresponded to dividing cells (positively stained mitotic figures for both markers) while other epithelial cells were engaged in the cell cycle but were not in the mitotic phase, as shown by the diffuse nucleoplasmic Ki-67 labeling in the absence of associated mitotic figures (Fig. 3). The epithelial stratification process occurring between PND14 and PND30 was associated with numerous Ki-67 and PHH3 immunopositive mitotic figures in the basal epithelial cells; most dividing cells in the peripheral region were observed between PND21 and PND30 while dividing cells were always detectable in the central corneal region (Fig. 4). Throughout the first month of age, only rare mitotic cells were present in the corneal limbal region. Corneal epithelial cells were not positive for caspase-3. Intense labeling for TUNEL was occasionally noted solely in the corneal epithelial cells of some rat pups. This labeling was sometimes observed in only one eye (or a portion of the cornea of one eye) of some rat pups and was not noted in all the rat pups included in the same PND group. Furthermore, this staining pattern was repeatable and therefore not due to random nonspecific labeling.
Stroma
From PND1 to PND6, the corneal stroma was thin, hypercellular, and composed of plump stromal fibroblasts, termed keratocytes (Fig. 1). At this stage, numerous Ki-67 and PHH3 immunopositive stromal keratocytes were noticeable, especially at PND1 (Fig. 3) and PND2. From PND6 to PND10, keratocytes in the anterior half of the corneal stroma (closer to the epithelium) began to elongate and only rare stromal cells were positive for Ki-67 or PHH3. At PND14, immediately following the opening of the eyelids, the stroma drastically thickened while the cellularity drastically decreased. The nuclei of the stromal keratocytes became fully elongated and quiescent at PND21 (Fig. 2).
Endothelium
Throughout the first month of age, the corneal endothelium remained a monolayer, however endothelial cells underwent morphologic changes during this time. At PND1, immature endothelial cells were rounded and presented a large clear intracytoplasmic vacuole (Fig. 1). From PND1 to PND10, the morphology of the endothelial cells transitioned from rounded to oval. At PND8 and PND10, a discrete discontinuous Descemet’s membrane was occasionally noticeable at the basal pole of the endothelial cells. At PND14, the endothelial cells flattened, their nuclei condensed and the intracytoplasmic clear vacuoles were smaller. From PND14 to PND30, the endothelial cells continued to flatten and Descemet’s membrane thickened (Fig. 2). Nevertheless, at PND30, the endothelial cells were not as elongated and Descemet’s membrane not as thick as the adult rat cornea. Of note, between PND1 and PND4, numerous Ki-67 and PHH3 immunopositive proliferating corneal endothelial cells (Fig. 3) were noticeable, while from PND6 to PND14, only rare immunopositive mitotic endothelial cells were present.
Since we observed in one animal rare structures resembling cilia on the apical surface (the pole facing the anterior chamber) of corneal endothelial cells, we assessed with SEM the presence of such structures. However, primary cilia were not visualized on the apical surface of the corneal endothelial cells, in any of the corneal regions, in rat pups aged PND1, PND6, PND14, PND21, or PND30 (data not shown).
Iridocorneal Angle and Trabecular Meshwork
At PND1 and PND2, a small iridocorneal angle recess was present and behind it, the developing iridocorneal angle (termed angle mesenchyme) was densely packed with plump mesenchymal cells (Fig. 5). In some rat pups, a discrete trabecular beam with minimal intertrabecular spaces was present solely in the anterior portion of the iridocorneal angle. At PND4, the angle recess was larger and additional intertrabecular spaces were formed in the anterior portion of the trabecular meshwork, while the posterior cells remained closely packed. At PND6, Schlemm’s canals were first noticeable next to the developing trabecular meshwork (Fig. 6). From PND6 to PND8, the trabecular meshwork progressed posteriorly (Fig. 6) and at PND10, the iridocorneal angle’s mesenchymal cells were differentiated into elongated cells in the anterior portion. Compared to the adult rat eye, the trabecular meshwork was more cellular at PND10. After PND14, the trabecular meshwork could only be partially evaluated because the iris was sometimes collapsed on the cornea. Nevertheless, the mesenchymal cells forming the trabecular meshwork appeared fully differentiated into flattened cells throughout the angle around PND21. Some Ki-67 and PHH3 immunopositive mesenchymal cells were seen in the developing iridocorneal angle between PND1 and PND10 (Figs. 7, 8). The postnatal development of the trabecular meshwork was not associated with caspase-3- or TUNEL-immunopositive mesenchymal cells at any of the time points.
Lens
Lens fibers
From PND1 to PND10, newly formed secondary lens fiber cells were numerous, resulting in a high density of nuclei in the bow region (Fig. 9). As the secondary fiber cells differentiated and internalized, their nuclei gradually disintegrated to form the organelle-free zone (OFZ) (Fig. 9). From PND1 to PND8, the shape of the nuclei in the differentiating secondary fiber cells transitioned from basophilic and elongated nuclei in the bow region to eosinophilic and rounded nuclear remnants near the developing OFZ (Fig. 9). Furthermore, from PND1 to PND8, prominent nucleoli (1–3 per nucleus) were distinguishable in the nuclei of secondary lens fiber cells (Fig. 9, inset). From PND6, the eosinophilic rounded nuclear remnants bordering the OFZ became less numerous and by PND14, these structures had almost completely disappeared. After the disintegration of the rounded nuclear remnants, only small basophilic nuclear fragments remained noticeable at the edge of the OFZ and this cellular debris was more abundant from PND14 to PND24, but some was noted until PND30. During the first month of age, secondary lens fiber cells gradually accumulated and the OFZ widened. In addition, from PND21 to PND30, the nuclear density in the bow region decreased, but remained higher compared to the adult rat lens.
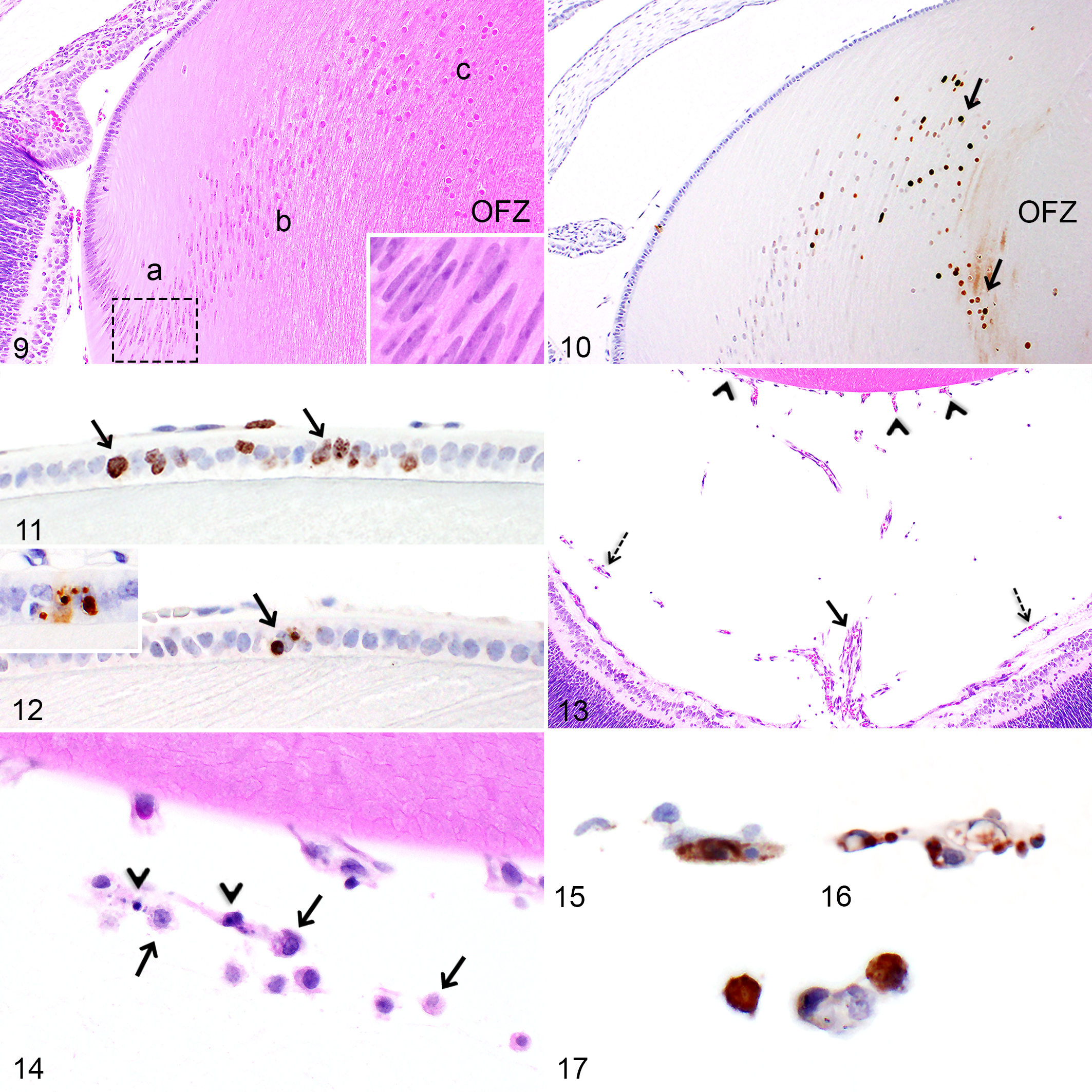
Importantly, nuclear remnants from the secondary lens fiber cells, including the eosinophilic rounded nuclear remnants and the small basophilic nuclear fragments lining the OFZ, were strongly positive for TUNEL histochemistry from PND1 (Fig. 10) to PND8, however those nuclear structures were not immunopositive for caspase-3.
Lens subcapsular anterior epithelium
At PND1, the lens anterior epithelium was simple cuboidal in the central region (Fig. 1) and simple columnar in the pre-equatorial region. At one week of age, the epithelial cells in the central region were oval and at three weeks, they were elongated (Fig. 2). The anterior epithelial cells continued to elongate until PND30 but they were still not as flat as in the adult rat lens.
At PND1 and PND2, numerous epithelial cells immunopositive for Ki-67 (Fig. 11) and PHH3 were noticeable in the central region of the subcapsular epithelium. Immunopositive dividing epithelial cells were mainly present in the pre-equatorial region and were more abundant during the first three weeks of age. From PND1 to PND10, scattered apoptotic epithelial cells, immunopositive for caspase-3 and TUNEL (Fig. 12) were observed throughout the entire length of the subcapsular anterior epithelial, except at the pre-equatorial regions.
Capsule
The lens capsule was undetectable at the posterior pole of the lens until PND6. Conversely, at the anterior pole, a thin capsule was detectable from birth. During the first month of age, the lens capsule gradually thickened, however, it did not attain the same thickness as in the adult rat lens.
Hyaloid Vasculature System
Various remnants of the hyaloid vasculature system were visible at birth and persisted for variable lengths of time (Fig. 13). From PND1 to PND8, remnants of the hyaloid artery were present at the caudal aspect of the vitreous and remnants of the vasa hyaloidea propria were noticeable alongside the inner limiting membrane of the retina. The vasa hyaloidea propria capillaries gradually became more distant from the retina as the pups aged and completely regressed at PND10. At birth, frequent remnants of the tunica vasculosa lentis were observed alongside the anterior, lateral and posterior aspects of the lens and they regressed completely first at the anterior pole of the lens at PND17 and subsequently at the lens posterior pole at PND26. Furthermore, from PND1 to PND21, macrophages (termed hyalocytes) occupying the vitreous and the anterior and posterior chambers were frequently observed and were often located adjacent to the regressing hyaloid capillaries (Fig. 14). Some hyalocytes could be seen until PND30.
Endothelial cells composing the capillaries from the tunica vasculosa lentis and the vasa hyaloidea propria, but not the hyaloid artery, were positive for caspase-3 (Fig. 15) and TUNEL (Fig. 16) at different PNDs (refer to Table 1). From PND1 to PND26, some hyalocytes were positive for caspase-3 and TUNEL (Fig. 17) or contained engulfed caspase-3- or TUNEL-positive apoptotic bodies.
Ciliary Body
The ciliary body epithelium, termed pars ciliaris retinae, is composed of two epithelial cell layers, the surface (inner) epithelial layer which consists in an extension of the sensory retina and the basal (outer) epithelial layer which is the continuation of the retinal pigmented epithelium (not pigmented in SD rats) (Fig. 18).
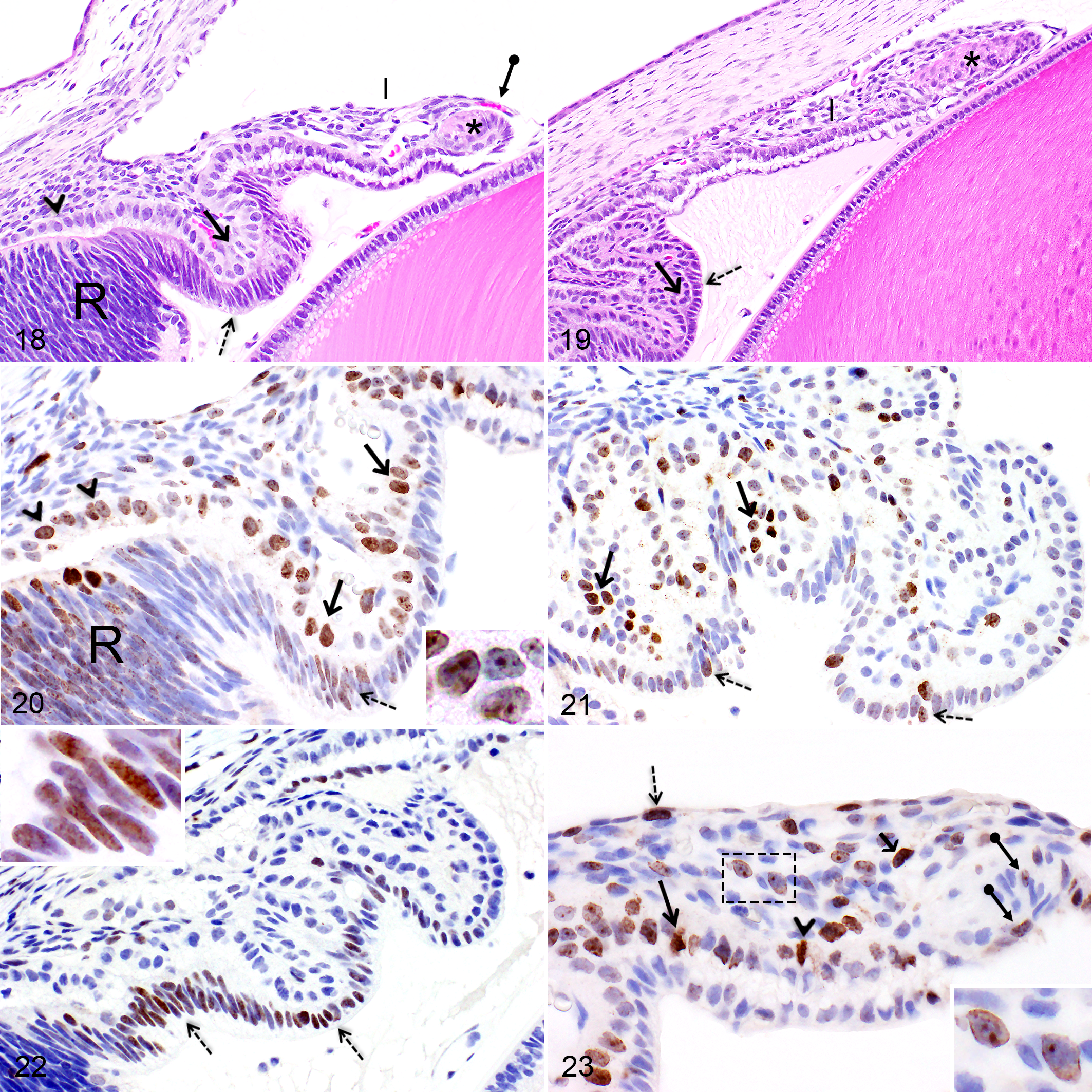
At birth, the ciliary body was markedly underdeveloped in SD rat pups as it corresponded to one large immature ciliary process presenting a thick pars ciliaris retinae (Fig. 18). The surface epithelial layer was composed of columnar cells which presented in some areas a pseudostratified organization. In addition, the surface epithelial cells in the region of the ora ciliaris retinae were markedly tall and disorganized (Fig. 18). The basal epithelial cell layer included large cuboidal cells with a voluminous nucleus and a pale amphophilic cytoplasm (Fig. 18). At PND4, both layers of the pars ciliaris retinae formed 2–3 immature large and short ciliary processes and the surface epithelial cells varied from columnar, near the ora ciliaris retinae, to cuboidal near the pars plicata (base of the iris) (Fig. 19). At this stage, the basal epithelial cells presented a darker, more condensed nucleus and an eosinophilic cytoplasm (Fig. 19). At PND6 and PND8, both layers of the pars ciliaris retinae were composed of cuboidal epithelial cells and the cytoplasm of basal epithelial cells became strongly eosinophilic. At PND14, the number of ciliary processes and their morphology were similar to the adult rat eye.
From PND1 to PND8, numerous basal cells of the pars ciliaris retinae showed diffuse nucleoplasmic or intense nucleolar staining for Ki-67 (Figs. 20, 21) and some were positive for PHH3. Concomitantly, the retinal epithelium in the region of the ora ciliaris retinae (which becomes the basal layer of the pars ciliaris retinae) also presented numerous Ki-67 (Fig. 20) and PHH3 immunopositive epithelial cells. From PND1 to PND10, occasional surface epithelial cells were immunopositive for Ki-67 (Figs. 20, 21) and PHH3. Between PND10 and PND24, only rare basal epithelial cells were immunopositive for Ki-67 or PHH3. Interestingly, from PND1 to PND8, the surface layer of the pars ciliaris retinae, mainly the portion near the ora ciliaris retinae, showed diffuse nucleoplasmic staining for caspase-3 (Fig. 22), however, those immunopositive epithelial cells did not display an apoptotic phenotype and were negative with TUNEL histochemistry.
Iris
At PND1 and PND2, the iris was markedly underdeveloped (Fig. 18). The iris was short with a poorly developed iridal sphincter muscle, a hypercellular stroma and an undifferentiated pars iridica retinae (posterior epithelium) (Fig. 18). Furthermore, until PND14, a delicate vascular membrane, likely compatible with a remnant of the pupillary membrane, was visible at the extremity of the iris, extending from a layer of fibroblastic cells lining the anterior edge of the iris (Fig. 18). At PND4 and PND6, the iris began to elongate, the cellularity of the anterior stroma decreased and the iridal sphincter muscle enlarged (Fig. 19). At PND6, the basal pole of the basal epithelial cells began to display a discrete eosinophilic coloration compatible with the development of myofilaments that later generate the dilator muscle of the iris. The basal epithelial cells completely differentiated into myoepithelial cells at PND21. The amount of smooth muscle cells composing the iridal sphincter muscle progressively increased and this muscle became fully developed between PND10 and PND14.
At PND1 and PND2, some basal cells of the pars iridica retinae were positive for Ki-67 (Fig. 23) and PHH3 while only rare basal cells were immunopositive from PND4 to PND10. Some of the fibroblastic cells lining the anterior edge of the iris and several mesenchymal cells present in the anterior stroma were also immunopositive for Ki-67 and PHH3 between PND1–PND8 and PND1–PND14, respectively (Fig. 23). Between PND1 and PND8, occasional myocytes composing the developing iridal sphincter muscle were positive for Ki-67 (Fig. 23) and PHH3. Caspase-3- or TUNEL-positive cells were not observed in the developing iris nor in the regressing pupillary membrane.
Retina
The retina of the neonatal rat is poorly differentiated and remarkable remodeling occurs during the first two weeks of age. The development of the peripheral retina (anterior region) was delayed by approximately 2 days compared to the development of the central retina (posterior region). When not specified, the PND associated with the histological or IHC findings described below corresponds to the time point when the changes occurred in the central region of the retina.
Nerve fiber layer and ganglion cell layer
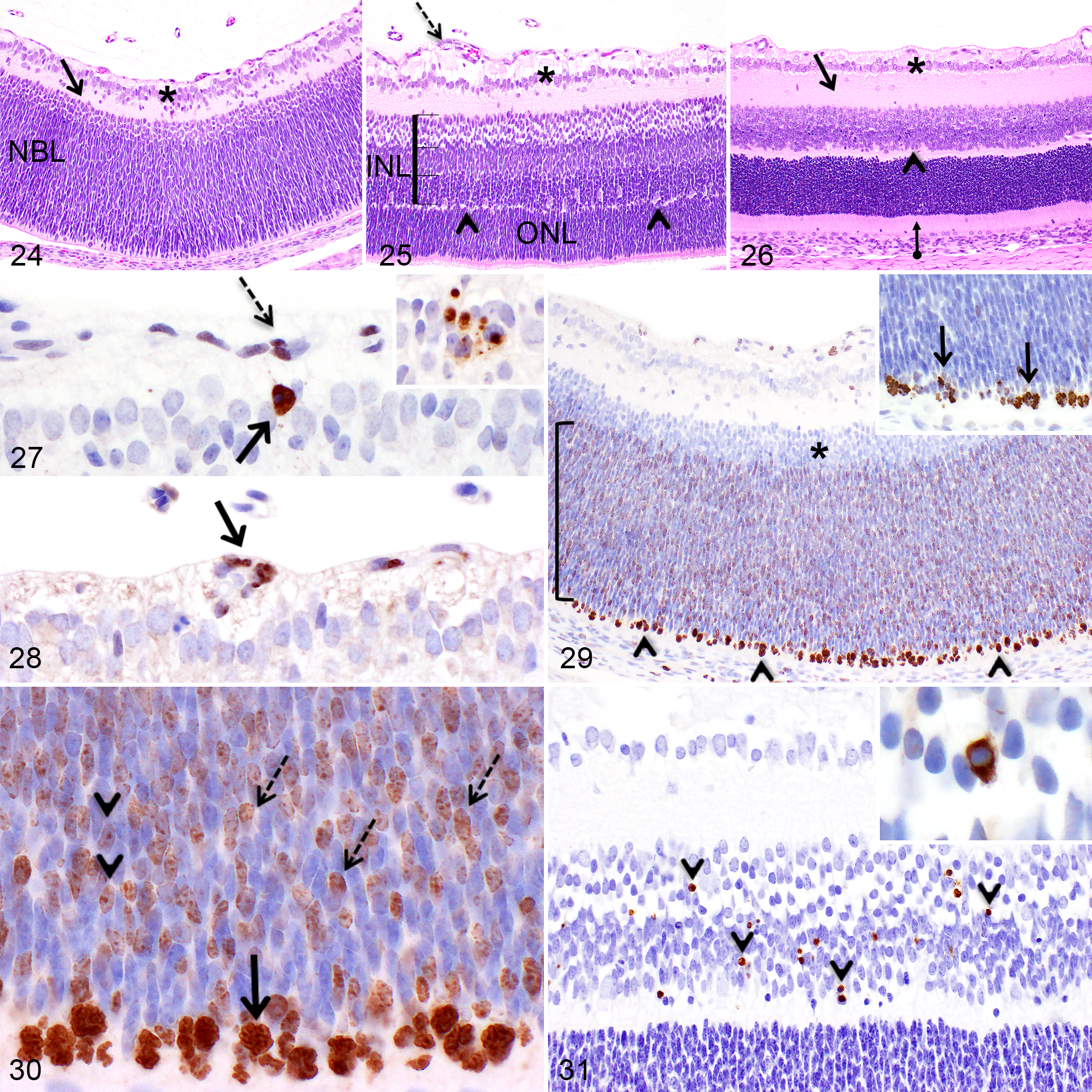
At PND1 and PND2, the ganglion cell layer was hypercellular and presented a multilayered (3–4) arrangement (Fig. 24) composed of densely packed large ganglion cells and fewer cells with small rounded nuclei. The hypercellularity of the ganglion cell layer was associated with a markedly thicker nerve fiber layer in the central region of the retina compared to the adult eye, but this layer was almost indistinguishable at the periphery of the retina until PND4. From PND4 to PND8, the ganglion cell layer drastically decreased to 1–2 cells thick centrally and the nonganglion cells observed within this layer became scarce (Fig. 25). Moreover, at PND8, the thickness of the nerve fiber layer at the periphery was almost equivalent to that of the center. The ganglion cell layer was mostly a monolayer of ganglion cells at PND14 (Fig. 26) while the nerve fiber layer was comparable to the adult rat retina at PND21. Until PND17, a heterogeneous population of cells was noticeable in the entire length of the nerve fiber layer. Until PND30, the cell density of the ganglion cell layer gradually decreased, however it remained higher than in the adult eye. Notably, at all time points, the ganglion cell layer was less cellular at the periphery of the retina compared to the central region. Ki67 and PHH3 immunopositive nonganglion cells were present in the ganglion cell layer solely at PND1. Some caspase-3- or TUNEL-positive apoptotic bodies and apoptotic cells were noticeable in the ganglion cell layer from PND1 (Fig. 27) to PND8 and in the nerve fiber layer from PND2 to PND6.
Primary vascular plexus of the retina
At PND1 and PND2, a heterogeneous cellular and vascular network, composed of numerous rounded and fusiform cells as well as capillaries lined by immature plump endothelial cells, emerged from the optic nerve head and extended in the nerve fiber layer lining the inner surface of the retina bilaterally. This vascular network corresponds to the primary plexus of the retina’s vasculature and occupied only the central portion of the retina at this stage. Between PND6 and PND10, the capillaries present in the central region of the retina were gradually replaced by small arterioles and venules, while some immature capillaries extended peripherally (Fig. 25) to reach the ora ciliaris retinae. At PND14, the primary vascular plexus was similar to the adult eye (Fig. 26).
From PND1 to PND8, capillary endothelial cells and nonendothelial cells present in the primary vascular plexus were immunopositive for Ki-67 (Fig. 28), while less numerous dividing PHH3-immunopositive cells were noticed. These immunopositive cells were present centrally as well as at the edge of the growing vascular network. Concurrently, from PND1 to PND10, numerous capillary endothelial cells and nonendothelial cells included in the primary vascular plexus throughout its entire length presented diffuse intranuclear labeling for capsase-3 (Fig. 27); however, these immunopositive cells only rarely presented an apoptotic phenotype and were only occasionally TUNEL-positive.
Inner plexiform layer
From PND1 to PND6, the inner plexiform layer appeared thin compared to the adult retina and it included several scattered nuclei (Figs. 24, 25). At PND14, immediately following the opening of the eyelids, the inner plexiform layer drastically thickened, reaching a similar thickness as in the adult retina (Fig. 26).
Neuroblastic layer (PND1–PND8)
From PND1 to PND6, the immature retina included a prominent undifferentiated neuroblastic layer, instead of an inner and outer nuclear layer. At PND1, this thick layer of undifferentiated neuroblastic cells was present over the entire length of the retina and was composed of densely packed fusiform nuclei; and the inner 1/5 band of the neuroblastic layer was markedly less cellular compared to the rest of this layer (Fig. 24). Furthermore, at PND1 and PND2, a prominent band of numerous mitotic multipotential neuroblastic cells was easily noticeable bordering the outermost edge of the neuroblastic layer on its entire length (Fig. 29). At PND4, a discrete outer plexiform layer (OPL) dividing the neuroblastic layer into a primitive inner nuclear layer and outer nuclear layer became noticeable solely in the central region of the retina. At this stage, the primitive inner nuclear layer in the central region appeared subdivided into three morphologically different zones; the inner zone contained less densely packed, rounded nuclei, the middle zone contained slightly paler basophilic fusiform nuclei, while the outer zone (anterior to the thin OPL) contained densely packed, dark fusiform nuclei with scattered large nuclei (compatible with horizontal cells). The newly formed outer nuclear layer was composed of densely packed fusiform nuclei and, at this stage, less numerous mitotic neuroblastic cells were noticeable at the outermost edge of this layer in the central region. At PND6, the three morphologically different zones composing the primitive inner nuclear layer became further distinct (Fig. 25) and, at this stage, the OPL, and therefore the inner nuclear layer and outer nuclear layer, reached half the length between the optic nerve and the ora ciliaris retinae bilaterally. A thin OPL extended to the ora ciliaris retinae at PND8 and at this stage, the last (and rare) mitotically active neuroblastic cells were present at the periphery of the retina bilaterally.
In addition to the prominent band of Ki-67- and PHH3-positive mitotic neuroblastic cells present at the outermost edge of the neuroblastic layer during the first 6–8 PNDs, numerous postmitotic neuroblastic cells migrating anteriorly displayed Ki-67 staining characterized either by diffuse granular nucleoplasmic staining or intense nucleolar staining (Figs. 29, 30). Importantly, those Ki-67-positive neuroblastic cells were negative for PHH3 (Fig. 29, inset), confirming their postmitotic stage. Furthermore, nuclei located in the inner 1/5 less densely packed band of the neuroblastic layer described previously did not stain positively for Ki-67 nor PHH3 (Fig. 29). At PND6, the Ki-67-immunopositive postmitotic cells present in the neuroblastic layer were solely noticeable toward the periphery of the retina (data not shown). At PND8, rare Ki-67- and PHH3-positive mitotic neuroblastic cells located at the outermost edge of the retina were present only near the ora ciliaris retinae, where the neuroblastic layer was still noticeable. Occasional apoptotic cells and apoptotic bodies immunopositive for caspase-3 and TUNEL were observed in the anterior half of the neuroblastic layer or in the newly formed inner nuclear layer, but not the outer nuclear layer, between PND1 and PND8.
Inner nuclear layer, outer nuclear layer, and outer plexiform layer (PND6–PND30)
At PND8, the cells previously composing the three well-defined morphological zones of the inner nuclear layer were blended together, resulting in a heterogeneous inner nuclear layer, and the nuclei present in the outer nuclear layer (photoreceptor nuclei) became oval with condensed chromatin. At PND10, the anterior half of the inner nuclear layer was less densely packed than the posterior half. At PND14, the inner nuclear layer was thinner than the outer nuclear layer and in the latter (Fig. 26), the chromatin of the nuclei formed several clumps within each nucleus. At PND21, both the inner nuclear layer and outer nuclear layer had become slightly thinner.
Apoptosis was frequently observed in the developing inner nuclear layer but never in the outer nuclear layer. Particularly at PND6 and PND8, the full thickness of the inner nuclear layer included numerous caspase-3-and TUNEL-positive apoptotic cells and apoptotic bodies (Fig. 31). From PND10 to PND17, some immunopositive apoptotic cells and apoptotic bodies were observed mainly in the posterior half of the inner nuclear layer.
Photoreceptor cell layer and the retinal epithelium
The photoreceptor layer was poorly developed at birth (Figs. 24, 32) and became distinguishable over the entire length of the retina at PND8. Following the opening of the eyelids at PND14, the photoreceptor layer drastically thickened. This layer reached a similar thickness as in the adult retina at PND21 (Figs. 26, 33). Before PND4, the retinal epithelium was composed of one layer of rounded to oval cells (Fig. 32). After PND4, occasional binucleated retinal epithelial cells became present in the central region and this phenotype became more frequent after PND10 (Fig. 33). In addition, from PND6 and thereafter, retinal epithelial cells became more rounded. Proliferating retinal epithelial cells immunopositive for Ki-67 and PHH3 were observed between PND1 and PND8 (Fig. 32, inset).
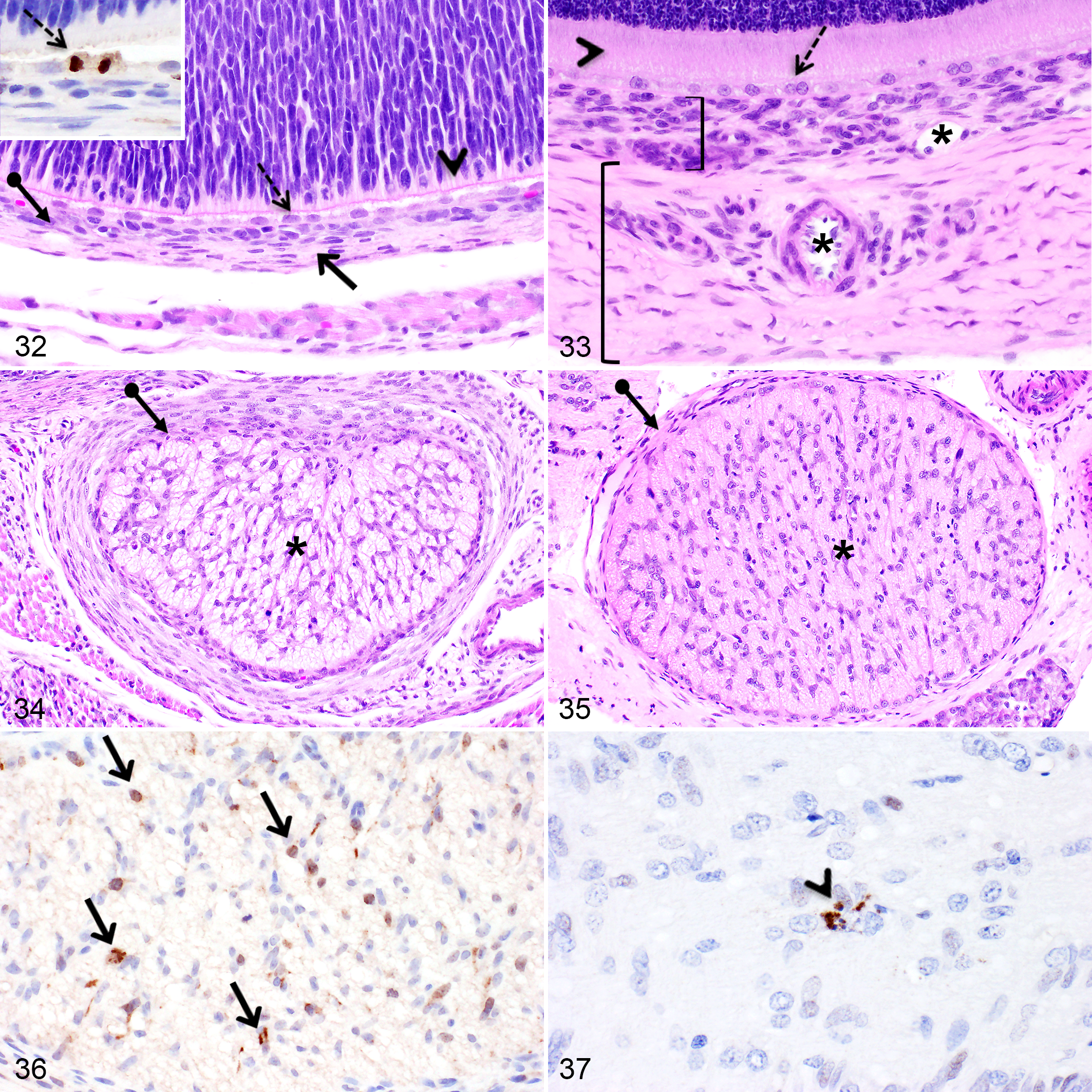
Optic Nerve
At PND1, the optic nerve was composed of randomly arranged glial cells, compatible with astrocytes, numerous plump glial progenitor cells and oligodendrocytes, as well as nonmyelinated nerve fibers (Fig. 34). By PND10, mature oligodendrocytes were more numerous and arranged linearly. Axon myelination was completed at PND14 (Fig. 35). Furthermore, at PND1, the meninges surrounding the optic nerve were thin, cellular and included rounded to oval meningeal cells (Fig. 34). After PND8, meningeal cells elongated, the meninges thickened and contained blood vessels (Fig. 35).
Proliferative glial cells immunopositive for Ki-67 and PHH3 were occasionally noticeable between PND1 and PND8 (Fig. 36), while they became rare at PND10 and PND14. The meninges surrounding the optic nerve occasionally included Ki-67 and PHH3 immunopositive meningeal cells between PND4 and PND8 (data not shown). Throughout the first month of age, occasional caspase-3- and TUNEL-positive glial cells were detectable in the optic nerve (Fig. 37).
Choroid
Minimal changes occurred in the choroid after birth except a thickening in the first 2 weeks of age as well as the development of small arterioles and venules in the vascular layer of the choroid during the first week (Figs. 32, 33). Before PND4, only capillaries from the choriocapillary layer were present. From PND1 to PND4, numerous oval cells within the choroid were immunopositive for Ki-67 and PHH3, while less numerous immunopositive proliferative cells were detectable until PND14.
Sclera
From PND1 to PND4, the sclera was thin and hypercellular (Fig. 32). From PND6 to PND10, the sclera slightly thickened through the addition of new collagen fibers, scleral cells began to elongate and the cellularity decreased. At the time of eyelid opening at PND14, a marked thickening of the sclera occurred, which was associated with the development of larger veins and arteries within the sclera (Fig. 33). Ki-67- and PHH3-immunopositive scleral cells were occasionally observed between PND1 to PND10.
Conjunctiva
The bulbar and palpebral conjunctival epithelium consisted of a monolayer of oval epithelial cells at PND1 and became a 2-cell layer at PND4. At PND6 and PND8, the second layer occasionally resembled a transitional epithelium and at PND14, the conjunctival epithelium was stratified squamous (4–5 layers) to transitional in some areas. Numerous conjunctival epithelial cells were immunopositive for Ki-67 and PHH3 at PND1 and PND2, while less numerous immunopositive basal epithelial cells were observed during the stratification processes from PND4 to PND10. After PND14, only occasional Ki-67- or PHH3-positive basal cells were noticeable.
Extraocular Structures
Harderian gland
At birth, the Harderian gland was markedly underdeveloped and composed of tubules lined by 1–2 layers of undifferentiated epithelial cells (Fig. 38). At this stage, the cytoplasmic to nuclear ratio was low compared to the mature gland. The cytoplasm of the epithelial cells was basophilic and rare intracytoplasmic vacuoles were present. After the first week of age, the glandular portion further developed and the tubules began to present lumens. At PND21, the cytoplasm of the epithelial cells was eosinophilic, contained numerous clear vacuoles and porphyrins were first noticeable in the gland lumina (Fig. 39).
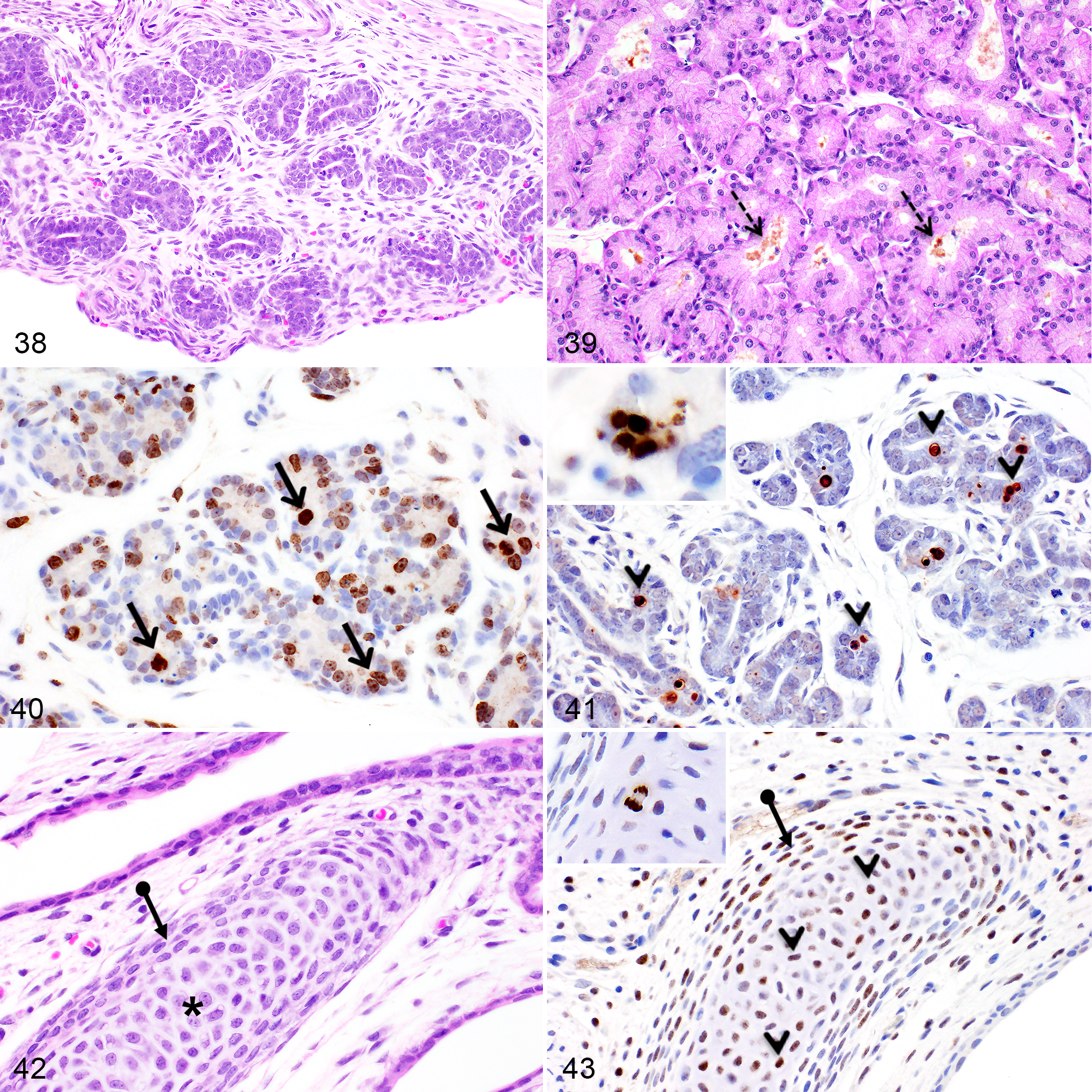
Numerous Ki-67- and PHH3-immunopositive epithelial cells were present during the first two weeks of age, while occasional immunopositive cells were detectable until PND21 (Fig. 40). Furthermore, abundant apoptotic epithelial cells positive for caspase-3 and TUNEL were noticeable between PND1 and PND10 (Fig. 41).
Nictitating membrane
From PND1 to PND6, the nictitating hyaline cartilage was short and composed of densely packed, immature, round, pale basophilic chondrocytes forming numerous isogenous groups while the cartilage matrix was pale and eosinophilic (Fig. 42). At this stage, a thick hypercellular perichondrium composed of plump fibroblasts was present and, after PND8, perichondrial cells became spindle-shaped. Furthermore, at PND8 the cartilage matrix became basophilic. At PND14 and PND17, some chondrocytes present in the center of the cartilage began to flatten, whereas chondrocytes found at the extremities remained rounded. From PND21 to PND30, chondrocytes continued to flatten, while the perichondrial cells further elongated. At PND30, the nictitating cartilage was similar to the adult, although it remained slightly more cellular.
Between PND1 and PND10, some chondrocytes were immunopositive for Ki-67 and PHH3, mainly toward the extremities of the growing nictitating cartilage (Fig. 43, inset). Simultaneously, from PND1 to PND14, numerous immature chondrocytes presented diffuse intranuclear IHC staining for caspase-3 (Fig. 43), while immunopositive cells were less numerous between PND17 and PND30. The caspase-3-immunopositive cells did not present an apoptotic phenotype and they were negative for TUNEL.
Discussion
Distinguishing the physiologic histomorphological changes in the normal postnatal organ development from drug-related lesions or diseases-related lesions in juvenile animals can be challenging, particularly when evaluating a structure as sophisticated and complex as the eye. As in most other altricial species, the rat is born with closed eyelids and markedly underdeveloped eyes. 42,73 The present study showed that rapid and drastic tissue remodeling events occur in several ocular structures, especially prior to the eyelid opening, around two weeks of age. The postnatal phase of the ocular development in the SD rat not only implies cellular proliferation in most structures, but also apoptosis, which was particularly remarkable in the developing lens subcapsular anterior epithelium, the retina’s developing inner nuclear layer and ganglion cell layer, and the Harderian gland. Apoptosis was also noted in the regressing hyaloid vasculature. Nuclear degradation in the newly formed secondary lens fiber cells was noteworthy after birth and was associated with intense and numerous TUNEL-positive nuclear remnants lining the lens organelle-free zone. Nonapoptotic activation of caspase-3 was observed in the basal cell layer of the immature epithelium of the ciliary body as well as in the developing nictitating cartilage. Cellular proliferation, highlighted by the presence of Ki-67- and PHH3-positive cells, was marked in the developing retina, lens, cornea, iris, ciliary body and Harderian gland. The rat eye reached an overall histomorphological maturity at PND21.
Cornea
The present study showed that the cornea of neonatal rats is markedly underdeveloped at birth and major histomorphological changes occur in association with the opening of the eyelids, notably the stratification of the epithelium and the drastic thickening of the stroma. The corneal developmental features reported in this study correlate with previous studies in rodents and were similarly observed in humans at the time of the eyelid opening, which occurs around 24 weeks of pregnancy. 17,29,78,86,96 It is noteworthy that the maturation of the corneal epithelium, including the stratification process and the basal cells shape differentiation, commenced in the central region and progressed toward the limbal region, as previously described. 17 Unlike what has been described in developing mice or in adult rabbits, primary cilia were not detected on the apical surface (pole facing the anterior chamber) of the corneal endothelial cells in rat pups included in our study. 6,23,34 However, since we only assessed 5 time points (PND1, 6, 14, 21 and 30), it is possible that we missed the time window when the assembly of primary cilia occurs in rats. Indeed, in mice, primary cilia were only temporarily assembled during the postnatal development of the cornea as they were shown to be involved in the morphogenesis of immature endothelial cells into mature hexagonal-shaped endothelial cells. 6 It is also a possibility that the cilia were lost during the processing and handling of the corneal tissues. Although we confirmed with SEM that corneal endothelial cells in developing SD rats adopt a hexagonal shape during the first month of life, it is possible that primary cilia are not required for the normal morphogenesis of corneal endothelial cells in rats.
In this study, Ki-67- and PHH3-positive proliferating epithelial basal cells and endothelial cells were observed before the opening of the eyelids, which correlates with previous studies in rats showing that corneal basal epithelial cells present in all corneal regions correspond to highly proliferative stem cells during the first 1.5 weeks of age and that 12% of corneal endothelial cells at birth are undergoing cellular proliferation. 17,29,48,97 Last, stromal keratocytes were only proliferating shortly after birth and they became fully elongated at PND21, which is required to allow an adequate transmission of light through the cornea. This marks the histomorphological maturity of the cornea. After PND21, only subtle changes were observed such as a discrete thickening of the corneal epithelium and Descemet’s membrane, and thinning of the endothelium.
Iridocorneal Angle
The sequence of morphological changes occurring in the developing trabecular meshwork of the iridocorneal angle in SD rats is similar as in mice and human, but with different developmental windows and rates; the development of the trabecular meshwork in rats seems to occur at a slightly faster rate compared to mice. 20,63,71,84 Similarly to what is published in other species, including mice, dogs, monkeys, and humans, the postnatal development of the trabecular meshwork of SD rats in the present study did not involve cell death. 63,64,75,82,84 However, our results contradict a previous study in SD rats that reported the occurrence of necrotic mesenchymal cells during the development of the trabecular meshwork. 72 This previous report in rats has initiated a debate on the developmental mechanisms of the iridocorneal angle in mammals; however, our data, together with several other studies in different mammalian species, suggest that cell death is not required to create the intertrabecular spaces in the iridocorneal angle in mammals. 63,64,75,82,84
Lens
The development of the lens is complex and requires different sophisticated cellular differentiation events to reach the transparency needed to adequately transmit light to the retina. 94 In humans, most lens developmental processes are completed during the late gestational stages, 50 while the lens of newborn rats is markedly immature and major morphological changes occur during the first month of age, especially the denucleation of secondary lens fibers. Interestingly, during the first 8 PNDs, the rounded lens fibers nuclei bordering the OFZ were positive for TUNEL histochemistry, demonstrating the presence of DNA breaks and suggesting that DNA fragmentation is one of the last nuclear degradation events occurring to form the OFZ. A similar TUNEL histochemistry staining pattern was reported in the bovine and chicken developing lens. 5,21 Moreover, in the present study, nuclear degradation in secondary lens fibers was not associated with the activation of caspase-3, suggesting that this apoptotic executioner caspase is not required during this differentiation phenomenon. The role of different caspases and apoptotic pathways during lens fibers differentiation has been widely debated as contradictory results exist. 61,92 Nevertheless, using knockout mice, executioner caspases including caspase-3, have been shown to be unnecessary for the lens fiber organelle loss, supporting our findings in rats. 94 On the other hand, apoptosis of lens anterior epithelial cells, shown with caspase-3 and TUNEL histochemistry, was noticeable as a physiological finding during the first 10 days postnatally. Apoptotic anterior epithelial cells were also observed in the developing lens of human, between gestational weeks 6 to 9 and postnatally in mice. 8,94 Some level of cell death in the immature lens cuboidal anterior epithelium may be required to generate the space needed for those cells to become adequately elongated to allow an optimal transmission of the light through the lens. 94 Furthermore, the inhibition of apoptosis in the differentiating lens anterior epithelium has been shown to lead to the development of anterior cataracts. 94 Although new secondary fibers continue to be added throughout life, the lens of SD rats reached similar histomorphological features to adults around 3 weeks of age postnatally.
Hyaloid Vasculature and Pupillary Membrane
The hyaloid vasculature is crucial for the nourishment of the developing lens and other ocular structures. 15 While the involution of the entire hyaloid system is normally completed by 35–36 weeks of gestation in humans, remnants of the pupillary membrane and of the embryonic hyaloid vasculature were present in neonatal rats and progressively regressed at variable rates within the first 3 postnatal weeks. These results are consistent with previous reports in rodents. 12,43,45,62,65 It is noteworthy that the complete regression of the hyaloid vasculature occurs at approximately the same time as the lens histomorphological maturity. In the present study, apoptotic endothelial cells were noted at various time points in the regressing vasa hyaloidea propria and the tunica vasculosa lentis capillaries, but not the regressing hyaloid artery and pupillary membrane, as opposed to previous studies reporting cell death in these two structures. It is possible that the occurrence of apoptotic cells in these structures was missed in our study due to the extremely short delay existing between the onset of apoptosis and the phagocytosis of apoptotic bodies, which can be less than 30 minutes, and the small number of cells composing the vestigial structures. 88 Furthermore, consistent with previous studies in rodents and humans, numerous macrophages (hyalocytes) were noted during the first 3 weeks after birth and some were apoptotic or contained apoptotic bodies, suggesting a role for these macrophages in the removal of regressing capillaries. 65,95 In addition, some studies have provided evidence that hyalocytes may induce apoptosis in hyaloid endothelial cells. 7,37,56,95
Iris and Ciliary Body
The iris and ciliary body of neonatal SD rats were markedly underdeveloped at birth, likely because the adjustment of the lens and the miosis or mydriasis of the pupil are not needed while the eyelids are fused. Those two structures developed rapidly prior to the opening of the eyelids at PND14 through cellular proliferation. The iris sphincter muscle became fully developed by PND14 while myoepithelial cells composing the iris pupillary dilator muscle only became fully differentiated at PND21, which correlates with previously described electron microscopy findings in albino rats. 53 Interestingly, in humans, there is also a delay between the development of the dilator muscle compared to the development of the sphincter muscle. 70 Notably, this study revealed the occurrence of intranuclear activation of caspase-3 in the epithelial cells of the surface layer of the ciliary body epithelium during the first 8 PND. This caspase activation was not accompanied by an apoptotic phenotype nor a positive staining for TUNEL histochemistry and thus suggests a nonapoptotic activation of caspase-3. The activation of caspase-3 without the induction of apoptosis has been associated with different crucial cellular events involved during organ development or tissue remodeling including terminal cellular differentiation of several cell types, cell migration and cell proliferation. 18,54,79 Therefore, it is possible that the activation of caspase-3 plays similar nonapoptotic functions during the development of the ciliary body epithelium, however its precise role needs to be further investigated. The ciliary body and the iris were histomorphologically mature at PND14 and PND21, respectively.
Retina and Optic Nerve
The retina of neonatal rats is markedly underdeveloped and represents the ocular structure that undergoes the most drastic morphological changes during the postnatal developmental stage, particularly during the first two weeks of age, when abundant cellular proliferation and programed cell death coexist. The sequence of histomorphological changes occurring during the development of the retina and the optic nerve in the SD rat is similar to other mammals, including humans, nonhuman primates, mice, and dogs, but with different time windows. 62,76,80,83 The major morphological developmental changes described postnatally in rats take place prenatally in humans. 62 The retina of the SD rat is considered histomorphologically mature at PND21, albeit subtle changes occur thereafter, such as the thinning of the outer and inner nuclear layers and a gradual reduction of cell density in the ganglion cell layer.
Cellular proliferation in the developing retina and optic nerve
During the first 8 PNDs, the outer edge of the thick neuroblastic layer contained abundant dividing multipotent neuroblastic cells, immunopositive for Ki-67 and PHH3, responsible for the generation of all cell types composing the mature retina. 9 The numerous Ki-67-positive and PHH3-negative nondividing neuroblastic cells present in the neuroblastic layer, anterior to the outer edge mitotic band, correspond to neuroblastic cells that have committed to a specific cell type and are migrating anteriorly to the right position where they will subsequently further mature. 2,4 The positive Ki-67 staining in the absence of PHH3 staining indicates that those postmitotic neuroblastic cells are still engaged in the cell cycle for a certain period of time. More precisely, the diffuse granular nucleoplasmic staining or the nucleolar staining suggest that those neuroblastic cells are engaged in the G1 or S phase, respectively, before exiting the cell cycle. 28 As cones and most of amacrine and horizontal cells are believed to be formed at birth in rodents, the proliferating and migrating neuroblastic cells noted in the present study most likely correspond to developing rods, bipolar cells and the remaining amacrine cells and Muller cells. 4,9,93 Furthermore, the Ki-67/PHH-3 negatively stained innermost band of the neuroblastic layer described in this study likely corresponds to postmitotic amacrine cells that have migrated into place prior to birth and have exited the cell cycle. 33,41,58 Thus, the three temporary morphologically different zones subdividing the newly formed inner nuclear layer between PND 4–8 are composed of amacrine cells (inner zone), bipolar cells (middle zone), and Muller cells with scattered horizontal cells (outer zone). 9,93 Similar to what has been reported in mice and other strains of rats, some ganglion cells and retinal epithelial cells were proliferating in rats for variable lengths of time after birth. 1,4 The proliferating phase of the retinal epithelium correlates with the appearance of binucleated cells, which are normally seen in rodents 1,87 It is not clear if Ki-67-positive epithelial cells are undergoing cellular division or only nuclear division to become binucleated. The proliferating cells noticeable within the developing rat optic nerve during the first 2 weeks of age are compatible with developing astrocytes and oligodendrocytes. 11,66
Apoptosis in the developing retina
It has been previously reported in rats that approximately 50% of the generated bipolar, amacrine, horizontal and Muller cells, but not the rods, die during the normal development of the retina, mostly during the postnatal phase. 41,89 This correlates with our findings as apoptotic cells were observed in the anterior half of the neuroblastic layer and in the newly formed inner nuclear layer, but not in the outer nuclear layer, which is mostly composed of rods in primarily nocturnal animals. 14 Moreover, the distribution of the apoptotic cells in the inner nuclear layer described in the present study suggests that bipolar cells undergo cell death for a longer period than amacrine cells as, between PND10 and PND17, apoptotic cells were present almost only in the posterior half of the inner nuclear layer. This finding is consistent with previous studies in rats. 41 Apoptosis in the ganglion cell layer, as observed in the present study during the first 8 PNDs, has been reported before as one of the mechanisms associated with the diminution of the cellular density of this developing layer during the postnatal stage. 19,68,69,89 Furthermore, in early postnatal days, the ganglion cell layer of rats has been shown to include both ganglion cells as well as displaced or migrating amacrine cells, which correlates with the population of nonganglion cells noted in the ganglion cell layer in the present study. Both cell populations were shown to display cell death in rats. 41,68
Primary Retinal Vasculature Plexus
Similarly to what was described in mice, the regression of the hyaloid vessels is rats occurred simultaneously with the emergence of a capillary network from the optic nerve head which progressively spread in the nerve fiber layer across the inner surface of the retina. 30 This primitive capillary network corresponds to the retinal primary vascular plexus, which is subsequently remodeled into three vascular networks located in the nerve fiber layer and the plexiform layers. This study highlights that the remodeling of the primary vascular plexus into a mature hierarchical vascular tree requires both marked cell proliferation and apoptosis. Indeed, apoptosis was previously shown to be needed for the pruning of some capillaries while larger blood vessels proliferate in the retina. 30,44 In addition, it has been established that the development of the retinal primary vasculature is preceded by an invasion of proliferating astrocytes emerging from the optic nerve head and spreading in a centrifugal fashion across the innermost aspect of the retina. 30,90 These astrocytes could contribute to some of the Ki-67- and PHH3-positive labeling noted in the developing retinal primary vascular plexus in rats. 31,60
Harderian Gland
The postnatal development of the Harderian gland in rats was associated with prominent cell proliferation and cell death through apoptosis. Interestingly, to our knowledge, the abundant cell death observed in the rat developing Harderian gland has only been reported recently and the nature of the apoptotic cells remain to be characterized. 67
Limitations of the Fixation Methods
Due to the small size of the rat pups in the first days of life, the eyes and optic nerves of 3 pups per time point ≤ PND10 were fixed in situ in neutral-buffered 10% formalin, while the eyes and optic nerves of the fourth pup per time point were removed and fixed in methanol Carnoy for comparison of fixation methods. As the preferred fixative for optimal tissue preservation of whole eyes is Davidson’s solution, the eyes and optic nerves of rat pups aged > PND10 were removed and fixed in Davidson’s solution. 16 While Davidson’s fixation resulted in the best morphology preservation, especially for the retina, in our study, methanol carnoy provided the worst histological quality as it caused marked shrinkage of all ocular structures. Formalin was considered largely satisfactory for the histological description and caused less issues for immunohistochemistry compared to Davidson’s solution. Indeed, the first issue with Davidson’s solution consisted of poor tissue adherence to the slides during the heat-induced epitope retrieval (HIER) step with citrate buffer despite the use of highly adherent slides. Similar to what has been reported by others, the HIER step frequently resulted in partial detachment of the ocular tissue section, especially the cornea, lens and the sclera. 16 Drying the slides overnight at 60°C after sectioning markedly improved the retention of sections on slides and did not alter the IHC staining when tested with positive tissue controls that were dried either at 37°C or 60°C overnight. Furthermore, we noted that the sensitivity of the IHC staining using the Ki-67 antibody seemed slightly less (fewer number of positive cells) in Davidson’s-fixed tissues compared to 10% formalin-fixed tissue when optimization tests were performed with positive tissue controls (intestine and lymph nodes). However, IHC staining using the capase-3 or PHH3 antibodies as well as TUNEL histochemistry were comparable with both fixation methods. Decreased IHC staining intensity and specificity in rat ocular tissues fixed with Davidson’s solution compared to ocular tissues fixed in 10% formalin has been reported for several antibodies. 16 Last, with both formalin and Davidson’s fixation methods, we occasionally observed intense nonspecific labeling for TUNEL solely in the corneal epithelial cells, which we suspect is a consequence of the unintentional and occasional drying of the cornea of some rat pups during the necropsy procedures. As the corneal epithelium is directly in contact with air during the necropsy procedures, we believe this is the most likely cause for the repeatable nonspecific staining observed only in the cornea of some rat pups.
Conclusion
The present study demonstrated that, within the first month of age, the eye of the SD rat undergoes dramatic morphological changes, especially associated with the eyelid opening at PND14, which require both cellular proliferation and apoptosis. Cellular proliferation was observed in most structures during the first 2 weeks of age, including the cornea, retina, iris, ciliary body, lens, sclera, choroid, and Harderian gland. Cell death through apoptosis was marked in the developing retina, the regressing hyaloid vasculature, the lens anterior epithelium and the Harderian gland. Altogether, the eye of the SD rat reached a histomorphological maturity around PND21. The data from this study will serve as a reference database useful in pediatric drug development as well as in other fields studying juvenile ocular pathology.
Footnotes
Acknowledgements
The study was supported through grants from the program Mitacs Accelerate, the «Fonds du Centenaire» of the University of Montreal and from CiToxLAB North America. We are thankful to Ancuta Apreutese for her technical help, Eiko Kawamura from the Western College of Veterinary Medicine of the University of Saskatchewan for her technical support with the SEM, as well as Cedric Gordon, study director, for his general support.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.