
Abstract
Stillbirth is a small and often cryptic fraction of neonatal mortality in mammals including pinnipeds. As part of an investigation into the poor reproductive success of the endangered New Zealand sea lion (Phocarctos hookeri), archived tissues from 37 stillborn pups born on Enderby Island between 1998 and 2012 were examined using histopathological techniques. Apart from bronchopneumonia with neutrophilic infiltration in 4 cases, few inflammatory conditions were identified in stillborn pups. However, 27/32 (84%) stillborn pups had aspirated squames present in the respiratory tract, without meconium. It is unclear if this finding represents fetal distress during parturition or whether it is a normal finding for this species. Three pups lacked histological evidence of hepatic glycogen storage, which may indicate placental defects or maternal undernutrition. No evidence of infectious disease was found on histopathological analysis, consistent with the low seroprevalence in New Zealand of infections known to cause reproductive failure in other pinniped species. This study forms an important baseline for further examination of stillborn New Zealand sea lion pups, as pup mortality is investigated as a contributor to the species’ decline.
Stillbirth, or delivery of a dead near-full-term fetus, is an uncommon but consistent cause of reproductive failure in mammalian species, including humans. While the exact etiology is often unknown, it is clear that a complex interplay between fetal, maternal, and placental factors combine to produce a viable offspring. In domestic animal species and even more so in wild populations, pathological investigation of stillborn fetuses is rare, perhaps due to the low yield of a definitive diagnosis, despite extensive diagnostic investigation. 26 Stillbirth, abortion and premature parturition have been regularly reported in routinely monitored pinniped species, 10,14 but in remote sub-Antarctic breeding sites utilized by the endangered New Zealand (NZ) sea lion (Phocarctos hookeri) little is known about the occurrence or extent of late gestational failure.
NZ sea lions are the only pinniped species endemic to New Zealand, with over 99% of breeding restricted to the NZ sub-Antarctic islands. The Sandy Bay colony at Enderby Island in the Auckland Islands archipelago is the species’ second largest breeding site and most intensively studied. In addition to restricted distribution, pup production is in decline, with almost half as many pups born in the most recent season (2014–2015), compared to 17 years prior when pup production was at an apparent peak. 5 Reproductive rate and average lifetime pup production estimates from Sandy Bay are low, with no clear reason identified for such poor reproductive success. 6,8 Although this site is the most well studied, outside of the austral summer field season (mid-December to mid-February) the island is uninhabited and early reproductive losses would be unnoticed. Reported stillbirth rates (proportion of stillbirths from total pup mortality) in otariids range from 2.6% to 9%. 18,23,27 –29 The stillbirth rate for NZ sea lions at Sandy Bay has been reported over 7 seasons, averaging 4.2% over this time. 4 This is likely to be an underestimate due to exclusion of early season abortions and premature parturitions. Long term trends in stillbirth rates have not been reported for NZ sea lions.
Whilst pup necropsies have been undertaken routinely at Sandy Bay since 1998, little to no investigation beyond gross examination has been performed. Accordingly, the aims of this study were to characterize the gross and histological lesions associated with stillborn NZ sea lions at Sandy Bay, Enderby Island, and to determine baseline epidemiological features of stillbirths at this site, including detection of possible trends over time.
Materials and Methods
Between 1998 and 2012, from mid-December to mid-February each year, all dead pups were collected at least once daily from the Sandy Bay colony, on Enderby Island, Auckland Islands (50°30’S, 166°17’E). Dead pups were identified by sex and flipper tag if present; and morphometric data were collected from 2001 onward including weight, body length (from nose to tail), axillary girth (circumference of the body just caudal to the pectoral flippers) and sternal blubber depth (measured ventrally at the level of the pectoral flippers). A full necropsy was undertaken as summarized by Castinel et al 4 by veterinarians or trained researchers. Tissue samples including lung, heart, liver, spleen, kidney, ovaries or testes, stomach, small and large intestine, pancreas, adrenal gland, thyroid, tongue, skeletal muscle, diaphragm, trachea, lymph nodes, cerebrum and cerebellum were preserved in 10% neutral buffered formalin for histopathology, with transport to and further processing at Massey University, Palmerston North, New Zealand. All samples were collected under permit from the New Zealand Department of Conservation (DoC). Approval for all work was obtained from the DoC Animal Ethics Committee: Approvals AEC52 (1 June 2002), AEC86 (31 December 2004), AEC158 (10 December 2007), and AEC200 (9 November 2009).
NZ sea lion necropsy records were reviewed and cases of stillbirth were identified by records of either visual confirmation of delivery of a dead pup or consistent necropsy findings including pulmonary atelectasis, a fresh umbilicus (Fig. 1.) and meconium in the colon (Fig. 2). Where formalin fixed tissues were available, they were trimmed, embedded in paraffin, sectioned at 4 µm and stained with hematoxylin and eosin for histopathological examination with grading (not present, mild, moderate or marked) of common findings as outlined in Supplemental Table 1. Special stains were used when required: Gram’s, Giemsa and Young’s fungal stains for further investigation of potential infectious agents and Periodic acid Schiff (PAS) for identification of mucins and mucopolysaccharides present in meconium.
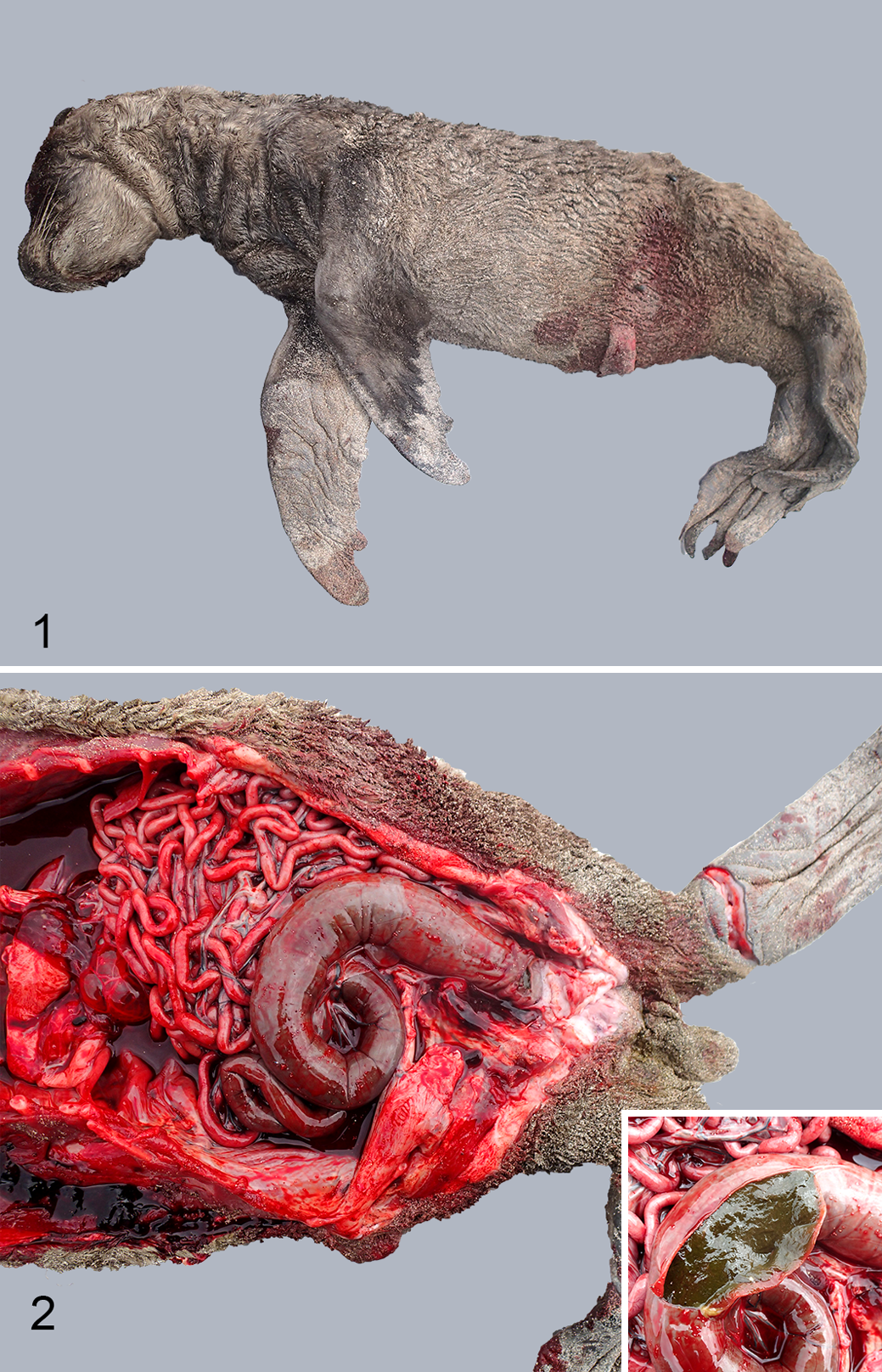
Stillborn pup, New Zealand sea lion (Phocarctos hookeri).
Data were tested for normality with the Kolmogorov-Smirnov test. Stillborn and live born morphometric comparisons were undertaken for normally distributed data with the Student’s 2–sided t-test, and for data that were not normally distributed, the Mann-Whitney test was used. Differences were considered significant when P < .05.
Results
Records showed that 812 pups had been necropsied at Sandy Bay, Enderby Island between 1998–1999 and 2011–2012. Forty-nine pups (26 male, 22 female, 1 unknown) were identified as stillborn from these records, of which 37 (20 male and 17 female) had archived samples that were processed for histopathology. The median date of birth for stillborn pups was 20 December (n = 48; range 5 December–30 January). This is approximately 1 week prior to the median birth date for live born pups (26–27 December). 7
In 2011–2012 postmortem examinations were only undertaken from 10 January onward, well after the median date of birth for stillborns. Therefore, data from this season should be interpreted with caution. When data from only entire seasons are included (1998–1999 to 2010–2011) this corresponds to a stillbirth rate (as a proportion of pup mortality) of 6%. Annual stillbirth rates over this period ranged from 1.7% in 2002–2003 to 28% in 2010–2011 (Fig. 3).
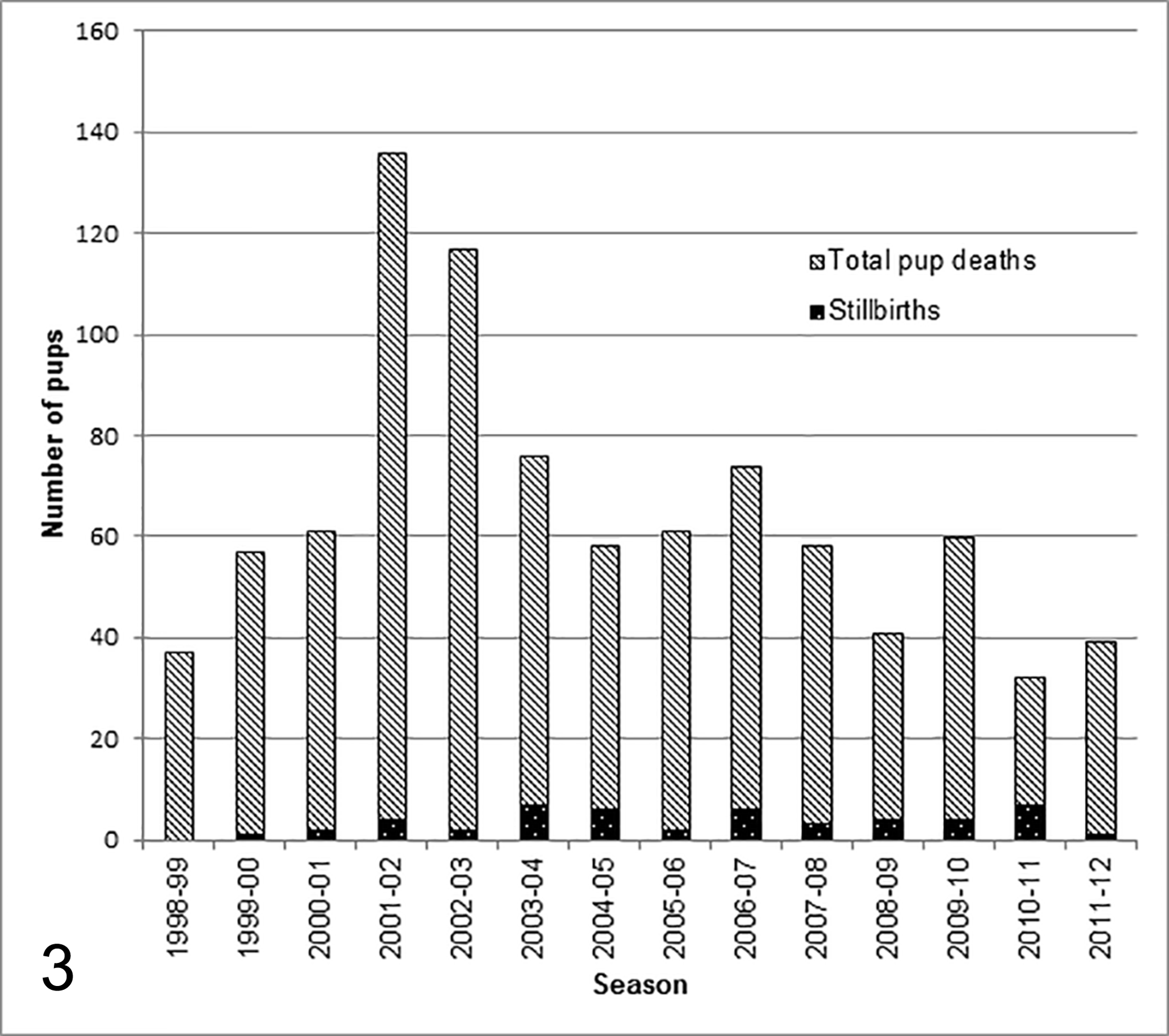
Comparison of stillborn New Zealand sea lions to total dead pups identified during monitored periods at Sandy Bay, Enderby Island. Note that necropsy schedule in 2011–2012 was incomplete.
Morphometric data were available for 33 stillborn pups (Table 1). The general trend was that stillborns were lower in weight and longer, with a smaller axillary girth than live born pups. This was significant in males for length (P = .001, Student’s t-test, n = 17) and girth (P < .0001, Mann-Whitney test, n = 17) and in females for mass (P = .026, Student’s t-test, n = 16) and girth (P < .0001, Mann-Whitney test, n = 16). Average sternal blubber depth was 9.6 mm and 9.4 mm for male and female stillborn pups respectively. Blubber depths of live pups were not determined.
Comparison of Morphometric Parameters of a Sample of Live Born and Stillborn NZ Sea Lion (Phocarctos hookeri) Pups at Sandy Bay, Enderby Island.

Mean ± SD
aSignificantly different (P < .05). bSignificantly different (P < .01).
Considering the circumstances of collection in the field and prolonged storage of samples, preservation was generally good, however tissues from 4 animals were severely autolyzed, making histopathological interpretation difficult such that these data were excluded. One of these was the only sample on which evidence of postmortem bacterial invasion (with a Gram positive rod) was evident on microscopic examination in all tissues examined including heart, liver, lungs, tonsil, spleen, intestinal tract and muscle.
Stillborn Pups
Tissues from 33 pups were sufficiently well preserved to allow histological analysis, however 1 tissue set was missing lung and 4 were missing liver. Common findings are summarized in Table 2. Lung sections varied from completely atelectic to mostly collapsed with slight peripheral aeration. In the majority of pups examined, there was some degree of foreign material present in the terminal bronchioles including homogenous eosinophilic material (likely amniotic fluid), sloughed squamous epithelial cells, cellular debris and alveolar macrophages. A typical case is seen in Fig. 4.
Summary of Histopathologic Findings and Severity in Stillborn NZ Sea Lion (Phocarctos hookeri) Pups.
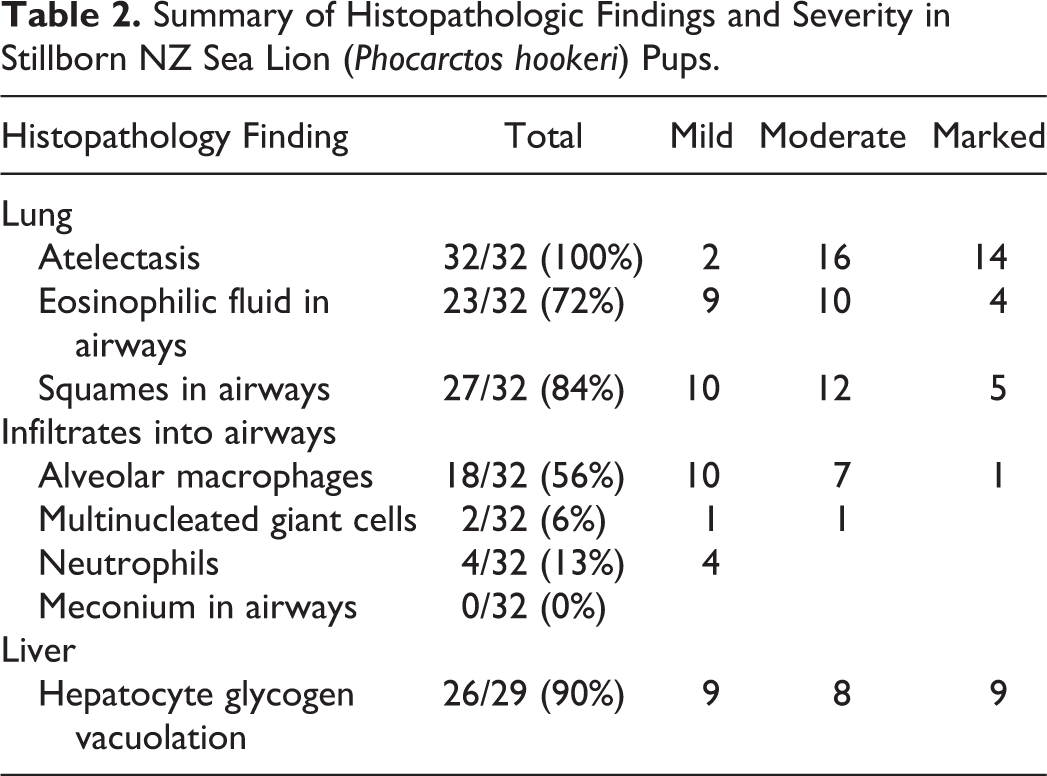
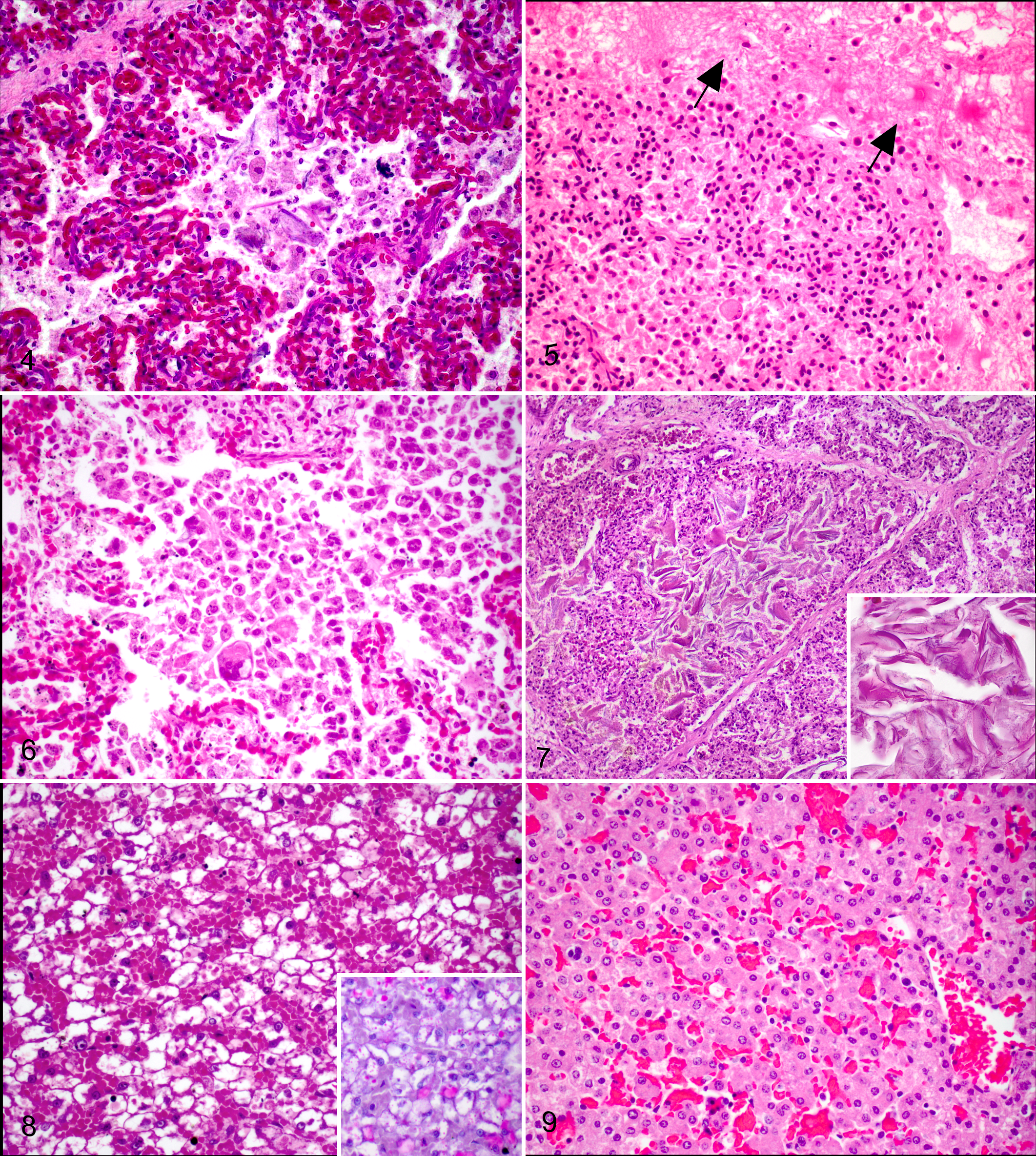
Four pups (13%) were diagnosed with aspiration pneumonia characterized by moderate to severe infiltration of alveolar spaces with neutrophils and macrophages around foci of squames. In 1 case this was also associated with proteinaceous fluid (amniotic fluid) and expansion of the interstitium by fibrin (Fig. 5) and in 2 cases with multinucleated giant cells (Fig. 6). These pups were not temporally associated.
In total, 27 pups had squames present within the alveoli, but 11/27 (41%) were not associated with any alveolar macrophage or other inflammatory cell infiltration (Fig. 7). There was no significant association between presence of squames and pup birth weight to indicate fetal distress and dystocia as the cause of aspiration (P = .64, Student’s t-test). There was subjectively a positive association between amount of aspirated material and intensity of histiocytic response. There was variation in the distribution of squames when present, usually appearing diffusely throughout the airways, in conjunction with amniotic fluid, cellular debris and varying levels of inflammatory response. In some cases however, the accumulation of squames was multifocally severe (Fig. 7), completely filling an entire airway, while other areas were unaffected. In these dense squame accretions there was no associated inflammatory response identified. Small round pink-orange globules were seen occasionally within airways but were not consistent with meconium on PAS staining, and were thought to be proteinaceous fluid.
The majority of pups (26/29 [90%]) exhibited diffuse vacuolation of hepatocyte cytoplasm, interpreted as storage of glycogen, in most cases obscuring the lobular structure at low power (Fig. 8). Hepatic extramedullary hematopoiesis was common, surrounding both portal tracts and sinusoids. In 3 animals however, there was markedly reduced vacuolation, resembling the histologic structure of an adult liver (Fig. 9).
No significant histological abnormalities were seen in any other tissues examined.
Discussion
Investigation of stillbirth beyond gross examination is often overlooked to the detriment of a comprehensive survey of neonatal mortality. This study attempts to address this knowledge gap in NZ sea lions by investigating archived tissue samples of stillborn pups over a 14 year period. Although historical tissue collection of stillborn pups has been sporadic, it is anticipated that this study can form a baseline that can be elaborated on in the future to form a wider view of late gestational reproductive failure in the species.
A consistent histological finding in 84% of stillborn NZ sea lion pups was the presence of sloughed squamous epithelial cells within the small airways and alveoli of the lungs, often with alveolar macrophages present. In 4 cases this was accompanied by neutrophilic infiltration and aspiration pneumonia. These findings are not uncommon in the human and domestic animal literature but the pathophysiology of periparturient fetal aspiration has not been fully established. 2,13 Large volumes of squamous cells within the lungs are commonly associated with fetal respiratory distress and anoxia in utero, with consequent aspiration of exfoliated squames in amniotic fluid, and often meconium. 9,11 Keratin within the airways is well known to invoke a dose dependent histiocytic inflammatory response, but in experimental studies on neonatal rats, meconium invoked a significantly more severe neutrophilic inflammatory response. 15,16 To our knowledge, there are no published reports of meconium aspiration in fetal pinniped pups, however periparturient aspiration of squamous epithelial cells has been noted sporadically. Moderate to severe aspiration of squames and other debris was identified in 30/59 (56%) of premature California sea lion pups (Zalophus californianus) examined histologically. 10 In some cases this was suggested to compromise initial expansion of the lungs, however associated inflammation was not reported. Lynch et al 14 also found a high prevalence (25/29 [86%]) of squamous cells in the lungs of aborted Australian fur seal pups (Arctocephalus pusillus doriferus), of which 6 had concurrent lymphoplasmacytic pneumonia. Of the stillborn northern fur seals (Callorhinus ursinus) examined histologically by Spraker and Lander, 29 only 2 pups (2/63 [3%]) were diagnosed with pneumonia (one with concurrent placentitis), however squame aspiration was not reported. Although all 4 NZ sea lion pups diagnosed with aspiration pneumonia had moderate to severe inflammation associated with aspirated squames, the majority of pups with squame aspiration had little to no inflammatory response. While the full significance of this finding is unclear, this may reflect a short survival time after squame aspiration. Without the finding of meconium aspiration in these animals, it is difficult to conclusively demonstrate that these pups experienced periparturient respiratory distress.
Dystocia was recognized as a common cause of perinatal mortality in northern fur seals, especially in small females giving birth to large pups 29 and could give rise to respiratory distress of the fetus during parturition. Although morphometric data are not available for pregnant females, the findings presented here suggest that dystocia also occurs in NZ sea lions, as some particularly large pups (up to 13.2 kg) were amongst the stillborn cohort. These cases would be consistent with fetal-pelvic disproportion described in domestic cattle, which is more common in male offspring, with high calf birth weight often being a more important factor than small maternal pelvic size. 19,20 Abnormal presentation of the fetus at delivery is also a contributing factor to dystocia in domestic animals but in Antarctic fur seals (Arctocephalus gazella) at least, breech presentation was more common than cephalic and although parturition duration of the former was longer, it was not reported to be associated with fetal death. 1 Fetal orientation at parturition does not appear to be a risk factor for dystocia in NZ sea lions with approximately 50% of pups born in breech position but correlation with risk of stillbirth has not been assessed (B. L. Chilvers, unpublished data).
Normal neonatal hepatocytes contain autophagosomes filled with glycogen, visible microscopically as distended “water clear” cytoplasm, 22 a common finding in stillborn NZ sea lion pups. Glycogen provides the newborn with an energy supply in the immediate postnatal period before suckling begins. Glycogen autophagy is upregulated under the influence of glucagon immediately after birth and maintained at high levels for around 12 hours, after which time the vacuolated microscopic structure of the hepatocytes has all but disappeared. 22 It has been well described that humans and mice that are deficient in hepatic glycogen at birth have poorer chances of survival and often die due to the effects of hypoglycemia and energy depletion. 12 Three (10%) of the examined stillborn NZ sea lion pups had no evidence of hepatic glycogen vacuolation on histopathology, a finding not previously reported in pinnipeds. One did not have recorded morphometric data, but the remaining 2 were born well below the mean stillborn weights for their sex, with scant (1–4 mm) blubber coverage, the least fat stores of all pups examined. For 2 of these pups, data were available regarding the mother, who in both cases had produced live pups in the years before and after the stillbirth, however information was not available on body condition of the live pups. This may correspond with death in utero before metabolic stores were established or fetal malnutrition. The latter, termed intrauterine growth restriction in human babies is characterized by a delivery weight that is small for gestational age, with minimal subcutaneous fat, small lungs, liver and thymus, and low liver glycogen levels. 3,31 This syndrome in humans has been suggested to be a consequence of either impairment of maternal-fetal transfer of nutrients due to placental pathology, fetal factors such as genetic abnormalities or inadequate maternal nutrition. 3 The latter likely occurs in NZ sea lions and nutritional stress has been proposed as a contributor to species decline. 24 On the other hand, these emaciated pups accounted for a small proportion of the sample, with average stillborn NZ sea lion sternal blubber depth of almost 10 mm. Assessment of the role of maternal undernutrition in NZ sea lion perinatal mortality is warranted, and should include routine investigation of placentas, examination of hepatic glycogen stores in early neonatal deaths, and assessment of maternal body condition.
The low prevalence of inflammatory lesions in NZ sea lion stillborn pups is in contrast to other otariid studies that have found relatively high prevalence on both gross and histopathologic examination. Spraker and Lander 29 undertook the most extensive study of mortality in a pinniped species to date, with full necropsies of over 3000 northern fur seals of all age classes over a period of 20 years. Gross lesions were identified in 191/252 (76%) of stillborn pups, but only 17/63 (27%) of the pups examined histologically had inflammatory lesions. The most common finding was placentitis (14/17 [82%]) with pneumonia, enteritis, pulmonary edema, or mild hepatitis occurring only in single cases. In premature California sea lion pups from San Miguel Island, inflammatory lesions were also present in the majority of cases (50/59 [85%]). 10 Although 9 of these pups were concurrently suffering domoic acid toxicity, bacterial infections (31/59 [53%]) affecting pup organs or the placenta and inflammation of unknown etiology (26/59 [44%]) predominantly in the lung and brain were well represented. Lynch et al 14 identified inflammatory infiltrates in lungs (8/29 [28%]) and hearts (8/30 [27%]) of aborted Australian fur seal fetuses as well as placentitis in 1 case (1/11 [9%]). These reports are in contrast to other pup mortality studies in which only cursory gross necropsies were performed and consequently large proportions (up to 49%) of animals were not able to be designated a cause of death, with a substantial risk of misdiagnosis for those animals with an assigned etiology. 17,18,23 Placentas have not been routinely collected from NZ sea lions to date, but this should be considered for future studies.
Although contrary to reports in other otariid species, in which disease has been a prominent feature of stillborn pathology, for the disease agents of interest it is perhaps not surprising that evidence was not found in the NZ sea lion pups examined. Previous work has shown that sub-Antarctic NZ sea lion populations have less than 1% seroprevalence to the agents Brucella abortus and Leptospira interrogans serovar pomona 25 and no detectable antibodies to T. gondii 21 indicating that the adult females sampled had not been exposed to these agents. Similarly, mainland New Zealand is one of the few places in the world free of Coxiella burnetii 30 and this status likely also applies to its outlying sub-Antarctic islands. Additional screening in future studies for other disease agents including influenza, distemper, Campylobacter and algal toxins may be useful, to determine presence in the NZ region. Future investigations should incorporate collection of placental tissue and ancillary investigations of placental and pup tissues designed to detect the presence of infectious agents.
Footnotes
Acknowledgements
We would like to thank Evelyn Lupton and Saritha Gils for their help with histological processing; Mike Hogan and Craig Thomas for postmortem sample assistance; and all those that contributed to New Zealand sea lion postmortem examinations in the field, including Jacinda Amey, Amelie Augé, Laura Boren, Kelly Buckle, Aurélie Castinel, Simon Childerhouse, Nick Gales, Katja Geschke, Nadine Gibbs, Baukje Lenting, Andy Maloney, Kate McInnes, and Laureline Meynier.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The project was in part funded by the Marion Cunningham Memorial Fund, of the New Zealand Veterinary Association Wildlife Society.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.