
Abstract
Thrombomodulin (TM) is a membrane glycoprotein expressed on endothelial cells, which plays a major role in the protein C anticoagulation pathway. In people with inflammation, TM expression can be down-regulated on endothelial cells and a soluble form released into circulation, resulting in increased risk of thrombosis and disseminated intravascular coagulation. TM is present in dogs; however, there has been minimal investigation of its expression in canine tissues, and the effects of inflammation on TM expression in canine tissues have not been investigated. The objective of this study was to evaluate endothelial TM expression in tissues from dogs with systemic inflammatory diseases. A retrospective evaluation of tissue samples of lung, spleen, and liver from dogs with and without systemic inflammatory diseases was performed using immunohistochemistry (IHC) and a modified manual IHC scoring system. TM expression was significantly reduced in all examined tissues in dogs diagnosed with septic peritonitis or acute pancreatitis.
Keywords
Thrombomodulin (TM) is a transmembrane glycoprotein primarily expressed on endothelial cells. 10,27 TM participates in various important biological processes, including hemostasis-thrombosis, inflammation, cancer, and embryogenesis. 10,13 The primary role of TM on endothelial cells is to activate the protein C anticoagulation pathway by binding to thrombin, which changes its substrate specificity from procoagulant factors to protein C. 3,14 This TM-dependent activation of protein C leads to potent anticoagulant effects primarily via the proteolytic inactivation of coagulation cofactors Va and VIIIa, thereby preventing unwanted blood clotting and thrombus formation. 11 For this reason, TM has been extensively studied in the context of systemic inflammatory diseases that can lead to thrombotic complications and potentially cause multiple organ failure. 4,10,15 In humans, TM is down-regulated on endothelial cells, and a soluble portion is released into circulation during inflammation, resulting in increased risk of thrombosis and disseminated intravascular coagulation (DIC). 12,15,32 Increased serum TM concentration in these conditions has been shown to be a biomarker of organ dysfunction and mortality in critically ill patients. 22,24 Similar severe inflammatory diseases occur in dogs. Although TM is expressed in dogs, there has been minimal investigation of TM expression in specific tissues, and the role of TM in inflammatory diseases has not been studied. 9,21,28 The objective of this study was to perform a retrospective evaluation of TM expression in paraffin-embedded formalin fixed tissues from dogs with and without systemic inflammatory diseases.
Materials and Methods
Normal tissue samples (lung, n = 7; spleen, n = 4; liver, n = 9) were retrieved from the archive of the Animal Health Laboratory at the University of Guelph from dogs that had been euthanized but had no gross or histopathological lesions of inflammatory or other disease. Hospital records from the Ontario Veterinary College Health Sciences Centre were searched between the years 2006 and 2011 to identify archived tissue samples from dogs diagnosed with septic peritonitis or acute pancreatitis. Patients were then selected based on whether they had clinical and laboratory findings supportive of systemic inflammatory response syndrome (SIRS). Patients diagnosed with sepsis due to bacterial peritonitis (lung, n = 11; spleen, n = 10; liver, n = 10) or with systemic inflammation due to acute pancreatitis (lung, n = 14; spleen, n = 13; liver, n = 14) were retrieved based on previously described criteria. 19,20,31 Archived samples from both normal and affected dogs were processed in the same histology laboratory and stored under identical conditions in the same facility. Immunohistochemistry (IHC) for thrombomodulin was performed on 4-µm tissue sections mounted on positively charged slides and using an automated stainer (Ventana BenchMark XT, Ventana Medical Systems Inc., Tucson, AZ, USA). Heat-induced epitope retrieval (HIER) at pH 8 was followed by incubation with mouse anti-human thrombomodulin (clone PBS-01; 1:200 dilution, Abcam Inc., Toronto, ON, Canada) for 1 hour at 37°C. An anti-mouse alkaline phosphatase-linked multimer detection system was used with Fast Red chromagen (Ventana Medical Systems). The sections were counterstained with modified Gill’s hematoxylin (Ventana Medical Systems). Lung from a separate healthy dog was used as a positive TM control (Fig. 1). For negative reagent controls, duplicate sections of each control and test tissue were subjected to the same IHC procedure with substitution of an irrelevant isotype-matched monoclonal antibody (in optimization assays) or antibody diluent alone (for routine assays).
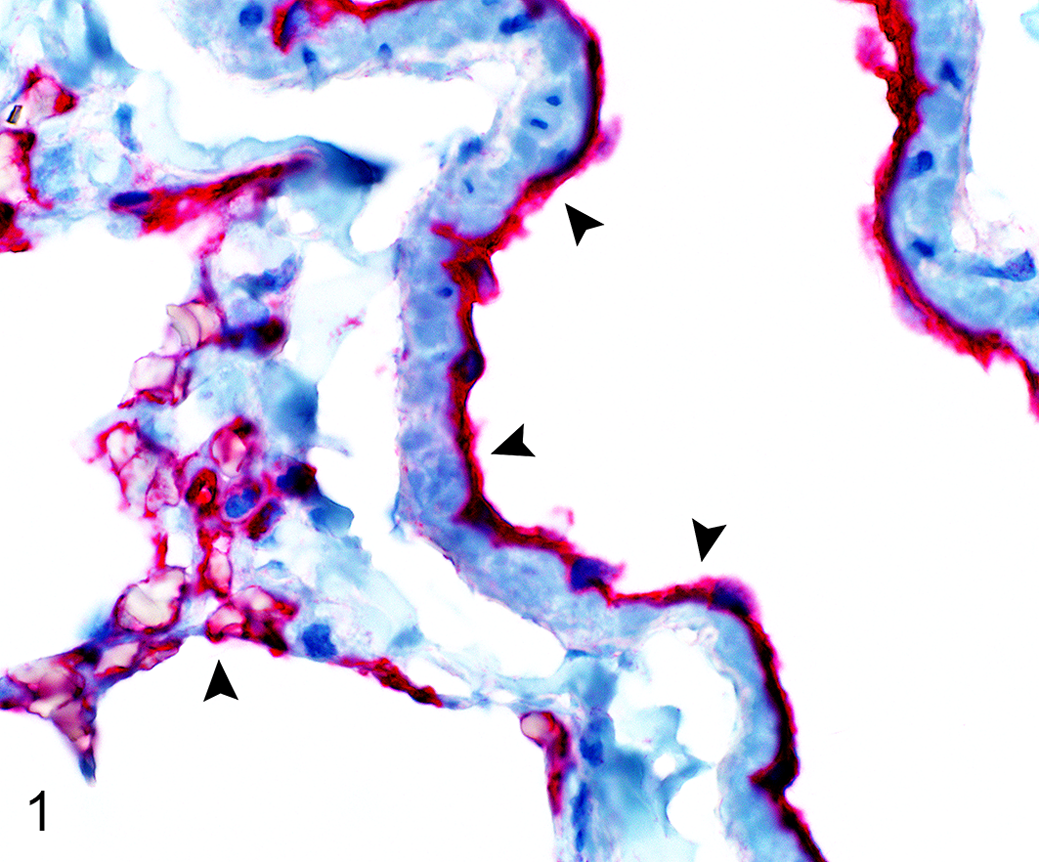
Positive thrombomodulin (TM) immunohistochemical staining, lung, normal dog. Arrowheads indicate strong endothelial TM immunoreactivity.
TM expression was assessed using a semi-quantitative scoring method described previously with modifications 5 such that scoring guidelines were established and applied to each type of tissue collected to account for variable structure and TM staining pattern (Supplemental Tables 1–3). The intensity of TM expression in lung and spleen was assessed semi-quantitatively as negative (0), weak (1), moderate (2), or strong (3) in 6 randomly selected high power fields (HPF) according to criteria described in Supplemental Tables 1 and 2. The scoring was performed independently by 2 of the authors (S. D. Kim, R. D. Wood ) using images obtained with a light microscope (Olympus BX43, objective, UplanFI 10x/0.30; Olympus, Hamburg, Germany). Image files were encoded to facilitate blinded analysis. The means of the respective results were calculated for further statistical analysis. Since previous studies reported poor TM staining in hepatocytes, 29,34,39 the total number of all vessels present in each section was instead counted along with the number of TM positive staining vessels (with score >0, Supplemental Table 3) to calculate the percentage of vessels stained positively. The mean of the respective results was calculated for further statistical analysis. All data were analyzed using the SAS 9.2 software package (SAS Institute, Inc., Cary, NC, USA). Numerical scores obtained for tissues from lung and spleen were analyzed using the Kruskal-Wallis test, whereas the percentage staining values obtained for liver tissues were analyzed using one-way ANOVA. Exact P values were calculated using the Monte Carlo method based on 1 million permutations. The Bland-Altman method was used to determine agreement between the 2 investigators. P values <.05 were considered statistically significant.
Results
Lung sections from normal dogs were characterized by strong positive TM staining with a mean score of 2.64 ± 0.078 (Fig. 2). The staining was continuous and homogeneous at the level of individual endothelial cells (Fig. 3A). The tissue sections from dogs with inflammation from peritonitis or pancreatitis had significantly reduced TM staining with a mean score ± SEM of 1.28 ± 0.20 and 0.91 ± 0.11 (P = .00002 and P = .003878 respectively, Fig. 2). TM staining pattern was discontinuous and not homogeneous, suggesting possible endothelial cell disruption and/or decreased expression of TM (Fig. 3B). Spleen from normal dogs was characterized by strong positive TM staining on the endothelium of most blood vessels and stroma within the spleen, with a mean score ± SEM of 2.41 ± 0.28 (Figs. 4A, 6). Spleen from dogs with inflammation from peritonitis or pancreatitis had significantly reduced TM staining with a mean score ± SEM of 1.18 ± 0.19 and 0.96 ± 1.35, respectively (P = .00408 and P = .023967, Figs. 4B, 6). No staining was visible in central arterioles, and weak to no staining was observed in the white pulp. The TM staining patterns were more diffuse and discontinuous throughout these sections (Fig. 4B). All liver sections had generally weak to no TM staining both in portal vessels and sinusoids, which is consistent with previous reports in mice and humans. 16,29 However, some moderate to strong staining in endothelium of hepatic arteries, portal veins, and central veins were present in the sections of normal dogs (Fig. 5A). The mean percentage of vessels labeled with TM antibody was 50.7% (95% CI, 42.1-59.3) for normal dogs, while dogs with septic peritonitis or acute pancreatitis had a significantly reduced mean of 7.0% (95% CI, 2.6-17.6) and 6.8% (95% CI, 3.3-13.5), respectively (P < .0001, Figs. 5B, 7).
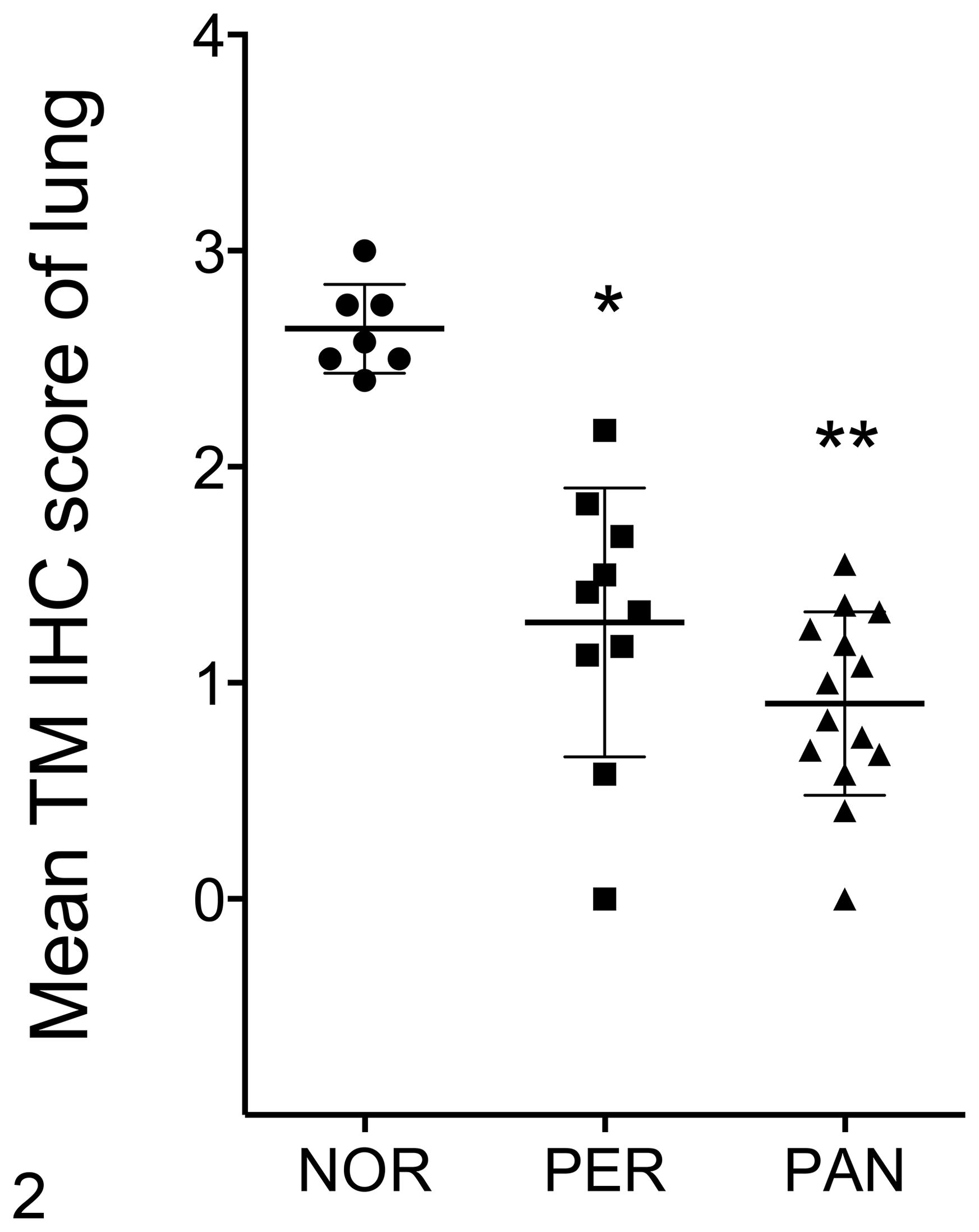
Thrombomodulin expression based on immunohistochemical (IHC) score in normal lung (NOR), or lung from dogs with septic peritonitis (PER) or acute pancreatitis (PAN). The data points show the mean IHC score of 6 high power fields (HPF) for each case. The mean ± SEM are shown. *P < .00002, **P < .004 (Kruskal-Wallis test, exact using Monte Carlo method).
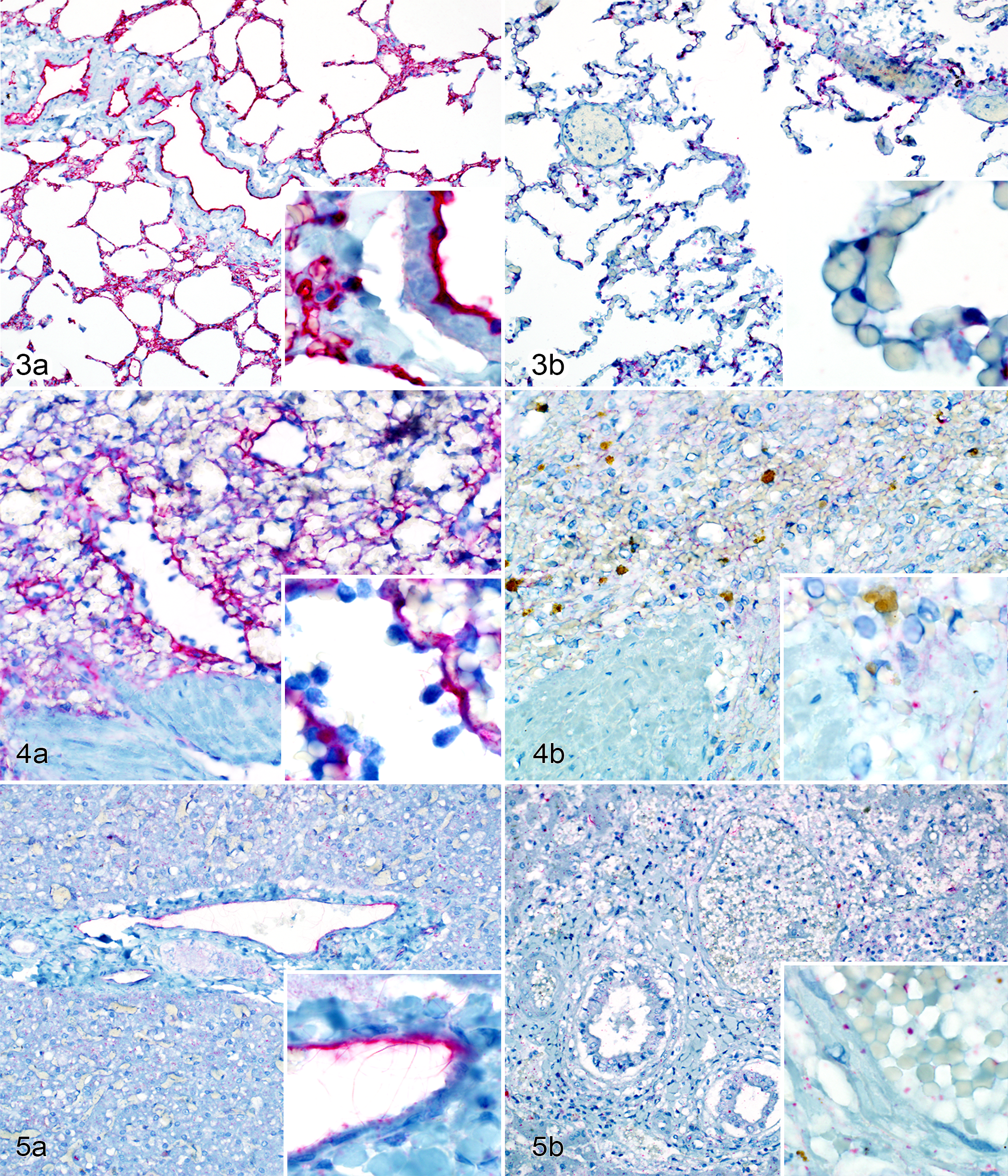
Thrombomodulin (TM) immunohistochemical labeling, dog.
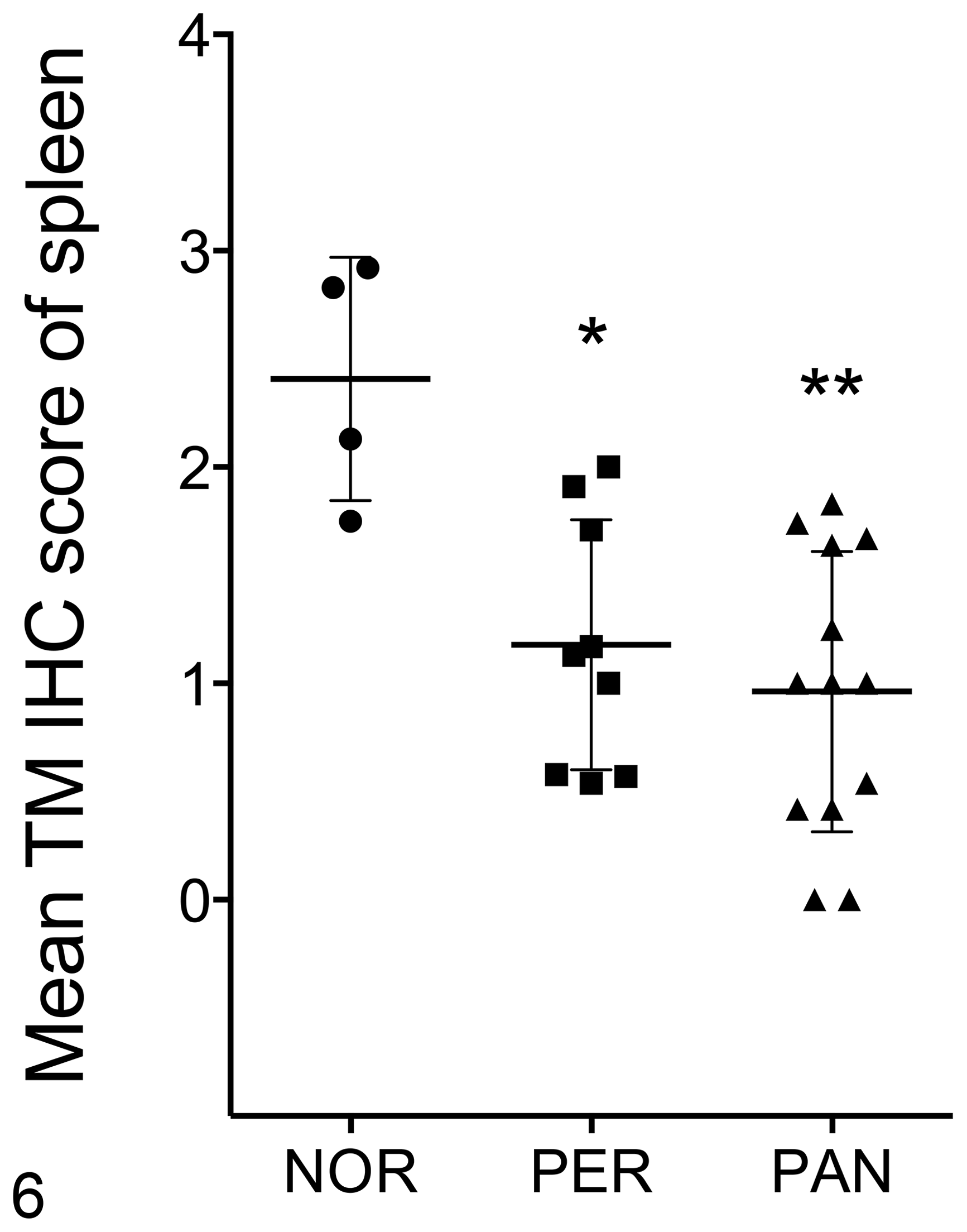
Thrombomodulin expression based on immunohistochemical (IHC) score in endothelial cells of normal spleen (NOR), or spleen from dogs with septic peritonitis (PER) or acute pancreatitis (PAN). The data points show the mean IHC score of 6 high power fields (HPF) for each case. The mean ± SEM are shown. *P = .004, **P = .024 (Kruskal-Wallis test, exact using Monte Carlo method).
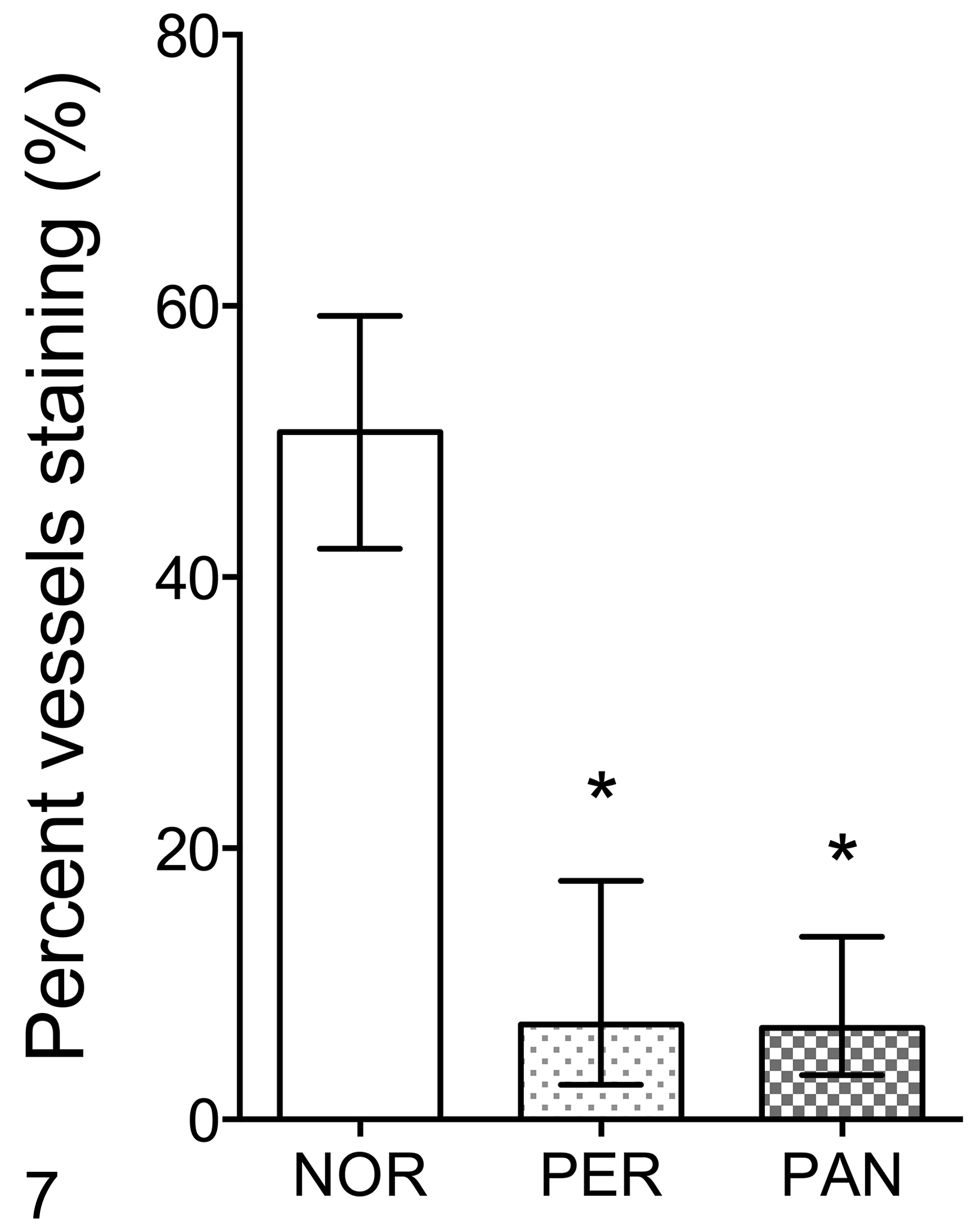
Extent of vascular expression of thrombomodulin in the liver of normal dogs (NOR) and those with septic peritonitis (PER) or acute pancreatitis (PAN). The data show the mean (± SEM) percentage of blood vessels that were labelled by immunohistochemistry for thrombomodulin. *P < 0.0001, 1-way ANOVA.
Discussion
The results of this study demonstrate that TM expression is significantly reduced in lung, spleen, and liver of dogs with systemic inflammation when evaluated by IHC. Previous reports suggest that down-regulation of TM expression on endothelium during inflammation may involve transcriptional and posttranslational regulation. Recent studies 23,32 showed that endothelial TM protein and mRNA levels were significantly decreased in response to TNF-α via the mechanism of NF-κB activation in bovine endothelial cells and human umbilical vein endothelial cells (HUVECs). This TNF-α dependent suppression of endothelial TM expression by infiltrating inflammatory cells may account for the potential transcriptional change in endothelial TM expression in lung, spleen, and liver of dogs with severe inflammatory diseases. Interestingly, however, the TM promoter lacks a NF-κB response element, and it has been shown that NF-κB mediates cytokine-induced down-regulation of TM expression by competing for binding to p300, a co-activator required for full expression of TM. 32
While inflammatory cells were not observed in some sections, all selected cases satisfied the inclusion criteria for systemic inflammation. Inflammatory mediators derived from neutrophils at the site of inflammation can have distant effects if present in systemic circulation. Previous reports suggest that neutrophil-induced oxidation or proteolysis of the extracellular domain of TM by inflammatory cells can occur. 1,2 TM is structurally organized into 5 distinctive domains: an N-terminal C-type lectin domain, a chain of extracellular EGF-like repeats, an extracellular serine/threonine-rich region, a transmembrane region, and a short cytoplasmic tail. 10,27 It was demonstrated that endothelial TM could be inactivated up to 90% by oxidation of methionine-388 located within the fifth EGF-like domain and later confirmed by others. 18,35 –37 It is suggested that the sources of oxidation are probably neutrophil-derived NADPH oxidase and reactive oxygen species (ROS) generated from inflammatory responses. 2,18 The production of proteases such as elastase and cathepsin G by activated neutrophils also appear to be involved in down-regulation of TM expression. Incubation of HUVECs with either granulocyte elastase or cathepsin G decreased cellular TM activity by 90% and 80%, respectively. 1 This study also demonstrated that the decrease in TM activity was accompanied by increase in soluble TM variants in the HUVECs cultured media. 1 The role of neutrophil-derived elastase and cathepsin G in the reduction of endothelial cell surface TM activity and increase in soluble TM was confirmed by later studies. 6,7,26,30
Furthermore, recent studies suggest other proteases such as matrix metalloproteinases (MMPs) and rhomboid may be involved. A bioactive lipid mediator present in biological fluids during endothelial damage, lysophosphatidic acid, induced shedding of the TM lectin-like domain in HUVECs, and it was MMP-dependent. 38 A family of evolutionarily conserved intramembrane serine proteases, rhomboid, was also shown to cleave the extracellular domain of TM on endothelial cells, leading to an increase in soluble TM. 8,25 Therefore, although the mechanism for down-regulation of TM expression on endothelial cells from the tissues of dogs with severe inflammatory diseases has not been investigated, the evidence to suggests that both transcriptional and posttranslational regulation are involved and that soluble TM may therefore be increased in plasma.
Lastly, while there tended to be different patterns and intensity of staining between tissues, even in healthy dogs, we documented decreased positive staining in all cases of inflammation. The differences in TM expression patterns observed in lung, spleen, and liver are likely due to the differences in tissue organization, relative number of endothelial cells per organ, and various types of endothelium comprising the organs. 17,33,34
There are a few limitations to this study. The small sample sizes and lack of matched controls for breed, sex, or age of the patients in the present study could be improved in future studies to more accurately estimate the TM expression. Furthermore, the use of a canine-specific TM antibody may be useful to overcome the challenges associated with evaluating TM expression in tissues, particularly in liver.
In summary, we demonstrated reduced expression of TM by immunohistochemistry in tissues of dogs with diseases that satisfied SIRS criteria. Future studies will evaluate soluble TM concentration in blood samples as a next step for determining if the protein can serve as a biomarker or prognostic indicator in dogs with systemic inflammatory conditions.
Footnotes
Acknowledgements
Sincere thanks to William Sears for assistance with statistical analysis, Dr Marie Holowaychuk for advice with study design, and to the staff of the histology section of the Animal Health Laboratory for assisting with immunostaining.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was generously funded by the Ontario Veterinary College Pet Trust and a University of Guelph Undergraduate Research Assistantship.