
Abstract
Ellis–van Creveld (EvC) syndrome is a human autosomal recessive disorder caused by a mutation in either the EVC or EVC2 gene, and presents with short limbs, polydactyly, and ectodermal and heart defects. The aim of this study was to understand the pathologic basis by which deletions in the EVC2 gene lead to chondrodysplastic dwarfism and to describe the morphologic, immunohistochemical, and molecular hallmarks of EvC syndrome in cattle. Five Grey Alpine calves, with a known mutation in the EVC2 gene, were autopsied. Immunohistochemistry was performed on bone using antibodies to collagen II, collagen X, sonic hedgehog, fibroblast growth factor 2, and Ki67. Reverse transcription polymerase chain reaction was performed to analyze EVC1 and EVC2 gene expression. Autopsy revealed long bones that were severely reduced in length, as well as genital and heart defects. Collagen II was detected in control calves in the resting, proliferative, and hypertrophic zones and in the primary and secondary spongiosa, with a loss of labeling in the resting zone of 2 dwarfs. Collagen X was expressed in hypertrophic zone in the controls but was absent in the EvC cases. In affected calves and controls, sonic hedgehog labeled hypertrophic chondrocytes and primary and secondary spongiosa similarly. FGF2 was expressed in chondrocytes of all growth plate zones in the control calves but was lost in most EvC cases. The Ki67 index was lower in cases compared with controls. EVC and EVC2 transcripts were detected. Our data suggest that EvC syndrome of Grey Alpine cattle is a disorder of chondrocyte differentiation, with accelerated differentiation and premature hypertrophy of chondrocytes, and could be a spontaneous model for the equivalent human disease.
Keywords
Ellis–van Creveld (EvC) syndrome was first described in children and is a complex clinical syndrome that presents with shortened limbs, retarded growth, postaxial polydactyly, and ectodermal and heart defects. 5 Epidemiologically, this syndrome is most frequent in the Amish community of Lancaster County (USA) due to endogamy. In a study of 52 Amish patients affected by EvC, 30 died under 6 months of age due to the syndrome. 15 EvC syndrome is an autosomal recessive disorder caused by a mutation in the EVC gene, located on human chromosome 4. 18 Takeda et al mapped the locus responsible for an autosomal recessive chondrodysplastic dwarfism in Japanese brown cattle to a region orthologous to human chromosome 4p16. 25 Two mutations in a gene contiguous to EVC, named limbin, were found in affected animals. 21,25 Mutations in the human ortholog of limbin, named EVC2, were later identified in human patients as responsible for a number of EvC phenotypes. 6 Loss of function mutations in the proteins coded by EVC and EVC2 lead to EvC syndrome, a recessive disorder. In contrast, negative function mutations in the terminal part of one of the proteins coded by these genes lead to Weyers syndrome, a dominant and generally less severe condition with incomplete penetrance. 20 These disorders are included in a group of diseases called ciliopathies, which are characterized by abnormalities in the primary cilia. 10 Mutations in the EVC or EVC2 gene can cause ciliary dysfunction, as these genes are involved in the development of cilia. 10 The EVC and EVC2 genes encode EVC protein and limbin, respectively, which are transmembrane proteins located in the basal bodies of the primary cilia. 10 Analysis of a murine model of EvC syndrome has demonstrated abnormal Hedgehog signaling. 19 Hedgehogs are a family of secretory peptides that have an important role in vertebrate development, controlling many critical steps of cell differentiation and patterning. 26 Genetic defects and anomalies of hedgehogs signaling pathways cause severe abnormalities in the appendicular, axial, and cranial skeleton in humans and other vertebrates. 26 Transgenic mice that overexpress sonic hedgehog (SHH) are affected by severe craniorachischisis and skeletal defects of the ribs, sternum, and long bones, with disarray of the growth plate in the tibia and absence of the femur and humerus. 26
Chondrocytes in growth cartilage are subjected to the influence of a plethora of extracellular factors, including systemic and local soluble factors, as well as components of cartilage extracellular matrix. 13 The major components of the cartilage extracellular matrix are the fibrillary cartilage-specific collagen II, with hyaluronan and proteoglycan aggrecan aggregates, giving the cartilage its special physical properties. Synthesis of collagen II is downregulated when chondrocyte hypertrophy occurs, at which stage synthesis of collagen type X is initiated. 13 Collagen X is primarily found in the hypertrophic zone, and its role is to facilitate the deposition of calcium within the matrix. 4 Therefore, collagen II can be used as a marker of cartilage matrix formation and collagen X as a marker of chondrocyte hypertrophy. 22
Fibroblast growth factor (FGF) signaling is essential for endochondral bone formation. Mutations cause achondroplasia, the most common human skeletal dysplasia.
12
Recently, a form of chondrodysplastic dwarfism was identified in Grey Alpine cattle (which we previously described as Tyrolean Grey cattle). A whole genome resequencing study revealed a single candidate causal mutation in EVC2, and Sanger sequencing confirmed the deletion of 2 base pairs in exon 19 as the cause of chondrodysplastic dwarfism in Grey Alpine cattle.
16
The aim of this study was to describe the gross and histologic lesions of EvC syndrome in Grey Alpine cattle; evaluate by immunohistochemistry the degree of differentiation and proliferation index of the physes to better elucidate the pathogenetic mechanisms of this EvC syndrome; and determine the level of expression of EVC and EVC2 mRNA in affected bones.
Material and Methods
Sample Collection
Five calves (3 females and 2 males, ranging from 2 months to 1 year of age) with a clinical diagnosis of chondrodysplasia were examined at necropsy. The diagnosis of EvC syndrome was confirmed by DNA testing, as the same animals were included in the whole genomic resequencing study that confirmed a deletion in the EVC2 gene. 15 Two Grey Alpine calves, 4 and 6 months old, that were not affected by deletion in the EVC2 gene were used as control subjects. Bones and soft tissues were routinely fixed in 10% formalin. Bone slabs 1 cm thick were decalcified in 10% EDTA solution. Sections 4 μm thick were cut from paraffin-embedded tissue and routinely stained with hematoxylin and eosin.
Immunohistochemistry
Five serial sections from decalcified bone samples of femur, humerus, rib, and vertebra of each animal underwent immunohistochemistry using antibodies to collagen II, collagen X, SHH, FGF2, and Ki67 (Supplemental Table 1). Sections were dewaxed and rehydrated. Endogenous peroxidase was blocked by immersion in 3% H2O2 in methanol for 30 minutes at room temperature. For detection of collagen II, collagen X, and SHH, enzymatic antigen retrieval was performed with 2.5% hyaluronidase from bovine testis (Type I-S, lyophilized powder, 400–1000 units/mg solid, Sigma-Aldrich, St Louis, MO, USA) in 0.01M phosphate-buffered saline (pH 7.4) containing Triton X-100 (0.025%) for 60 minutes at 37°C as described in the literature. 23 For detection of Ki67 and FGF2, antigen retrieval was performed by incubation in citrate buffer (pH 6.0) for 40 minutes in a heated bath at 95°C, followed by cooling at room temperature for 20 minutes. All antibodies were incubated with the tissue sections overnight at 4°C. Binding sites were revealed by use of secondary biotinylated antibody (dilution 1:200) and amplified using a commercial avidin-biotin-peroxidase kit (VECTASTAIN ABC Kits, Peterborough, UK). The chromogen DAB (3,3′diaminobenzidine; 0.05% for 3 minutes at room temperature) was used. Slides were counterstained with Papanicolaou hematoxylin. Primary antibody was replaced with an irrelevant, isotype-matched antibody as a negative control. For all antibodies, sections of humeral epiphysis, growth plate, and metaphysis of 2 unaffected healthy 4- and 6-month-old Grey Alpine calves were used as controls to assess the cross-reactivity with bovine tissues and the specificity of the immunohistochemical procedure. Controls underwent the same fixation and decalcification procedure. For each antibody, nondecalcified soft tissue of known immunohistochemical reactivity was used as a positive control to assess artifacts secondary to the decalcification procedure. Bovine urinary bladder was selected for SHH, 8 bovine lung and bronchial cartilage for collagen II, bovine small intestine for Ki67, and bovine granulation tissue for FGF2.
Labelling was first recorded as either positive or negative; then in each positive sample, within the growth plate, the location (resting, proliferative, and/or hypertrophic zone), distribution (diffuse, focal, multifocal and locally extensive), and intensity (strong, moderate, weak) of the labeling were evaluated. The Ki67 labeling index was obtained from the evaluation of 5 high-power fields, using an image analysis program (ImageJ, http://imagej.nih.gov/ij/index.html).
Molecular Analysis
Tissue samples from tibia, fibula, humerus, femur, tarsal bones, laminae of the hoof, rib, vertebra, heart, trachea, testicle, and tooth of 1 affected calf (No. 5) and 1 control calf (No. 6) were collected and conserved in RNAlater (Life Technologies, Gaithersburg, MD, USA) immediately after slaughter.
RNA was extracted using the RNeasy mini kit (Qiagen GmbH, Germany), per the manufacturer’s instructions, with the following modifications. The tissues (100 mg) were finely crushed in a 2-mL tube containing stainless-steel beads and 1 mL of TRIZOL (Ambion, Austin, TX, USA) using mechanical means (3 × 1-minute cycles of Tissue Lyser from Retsch, Qiagen). Then, 0.2 ml of chloroform was added and the RNA separated through centrifugation (12 000 g, 15 minutes, 4°C). Additional steps were carried out as described by the manufacturer.
Genomic DNA removal and synthesis of cDNA were performed using the Quantitect Reverse Transcription Kit (Qiagen GmbH, Germany), 200 ng of RNA, and oligodT mix, per the manufacturer’s instructions. The cDNA product was amplified using the primers EVC and EVC2 (Supplemental Table 3) designed using PRIMER3. 28 Polymerase chain reaction (PCR) was carried out using AmpliTaq Gold 360 Mastermix (Life Technologies, Carlsbad, CA, USA) with the following PCR conditions: 10 minutes at 95°C for preheating, then 1 minute at 95°C, 40 seconds at 60°C, and 1 minute at 72°C for 34 cycles. The resulting PCR products were separated on a 2% agarose gel with 0.5 µg/ml of ethidium bromide and visualized under ultraviolet light on a transilluminator.
Reverse transcription PCR (RT-PCR) products were purified using ExoSAP-IT PCR product cleanup (Affymetrix, Santa Clara, CA, USA) and directly sequenced using the PCR primers on an ABI 3730 capillary sequencer (PE Applied Biosystems, Foster City, CA, USA). Sequence data were analyzed using Sequencher 5.1 (Gene Codes Corporation).
Results
Necropsy Findings
At necropsy, the limbs of all the calves were disproportionately short and bulky, rotated and arched in a “dumbbell-like” form, with an angular medial outward rotation of femur and tibia, suggestive of genu varum (Figs. 1–4). The long bones were severely reduced in length, with a short diaphysis (Figs. 3, 4). Morphometric results of chondrodysplastic bones compared with normal long bones (Figs. 1, 2) are presented in Supplemental Table 2. The humerus was most severely shortened and malformed, followed by the femur. The radius, ulna, tibia, fibula, metacarpal and metatarsal bones, proximal and distal phalanges, vertebrae and ribs were not altered in either shape or length. The articular cartilage of the distal femur from case No. 5 had a locally extensive area (9 × 6 cm) of cartilage loss, with yellow fibrillated cartilage around the edges of the lesion. No polydactyly or ectodermal dysplasia affecting hooves, hair, or teeth was observed. In calf No. 3, valvular dysplasia of the left atrioventricular valves, characterized by myxomatous valvular degeneration, with an associated atrial jet impact lesion was observed. An affected 2-month-old calf (No. 4) had cardiomegaly, dilatation of the common trunk of the pulmonary artery, and a patent foramen ovale (2–3 mm in diameter). Affected calf No. 5 was a unilateral right abdominal cryptorchid, with an undescended atrophic testicle (5.5 cm in diameter). The normal descended testicle was 8.5 cm in diameter.
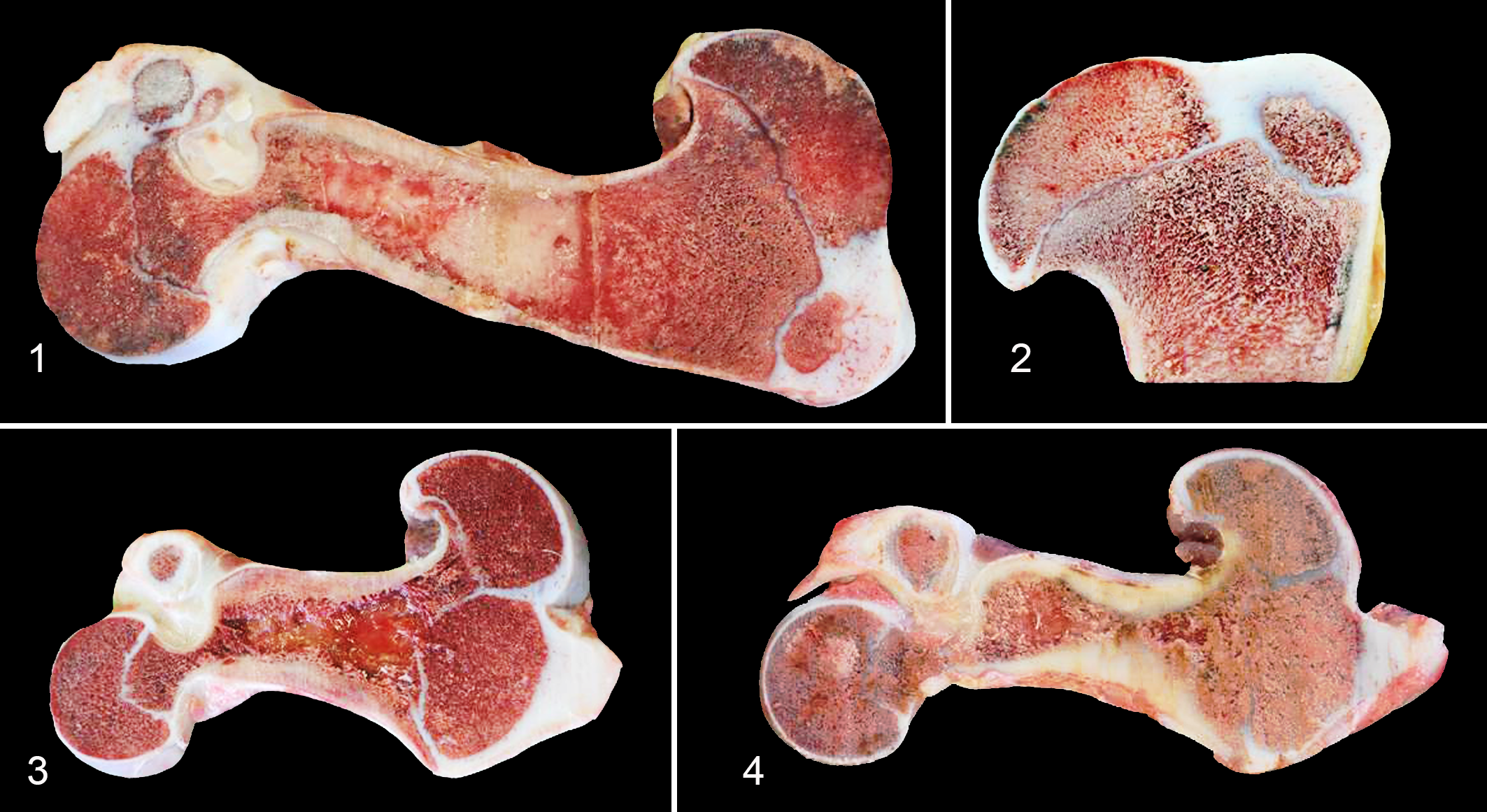
Ellis–van Creveld (EvC) syndrome, bone, calf. Sagittal sections of the femur from a control calf and calves affected by EvC syndrome.
Histopathologic Findings
The proximal humeral, proximal, and distal femoral physes, as well as the thoracic vertebra and the costochondral junctions, from 5 affected calves ranging in age from 2 to 12 months were examined (Figs. 5–10). The most consistent lesion, seen in all animals and all bones examined, was irregular thickening of the physis with tongues of physeal cartilage, consisting of both proliferative and hypertrophic chondrocytes, extending into the metaphysis. At times, these tongues of cartilage formed islands that were separated from the growth plate (Fig. 6). The cartilage matrix, particularly within cartilage islands but also within tongues of cartilage, was at times eosinophilic, suggesting matrix degeneration. Islands of cartilage deeper in the metaphysis were encased in a thin layer of osteoid. In the proximal femoral growth plate of calf No. 2, endochondral ossification had occurred within the physis, resulting in physeal duplication. In some areas of the physes, the cartilage matrix was rarefied, with exposure of collagen strands that extended across the space created by the chondrolysis. The epiphyseal-articular growth complex of the proximal humerus was similarly but more mildly affected, with small focal areas of chondrolysis and mild focal thickening of the articular cartilage. In the distal femur of calf No. 5, multiple islands of cartilage were encased in osteoid in the epiphysis, and a large tongue of cartilage extended from the articular-epiphyseal complex deep into the epiphysis.
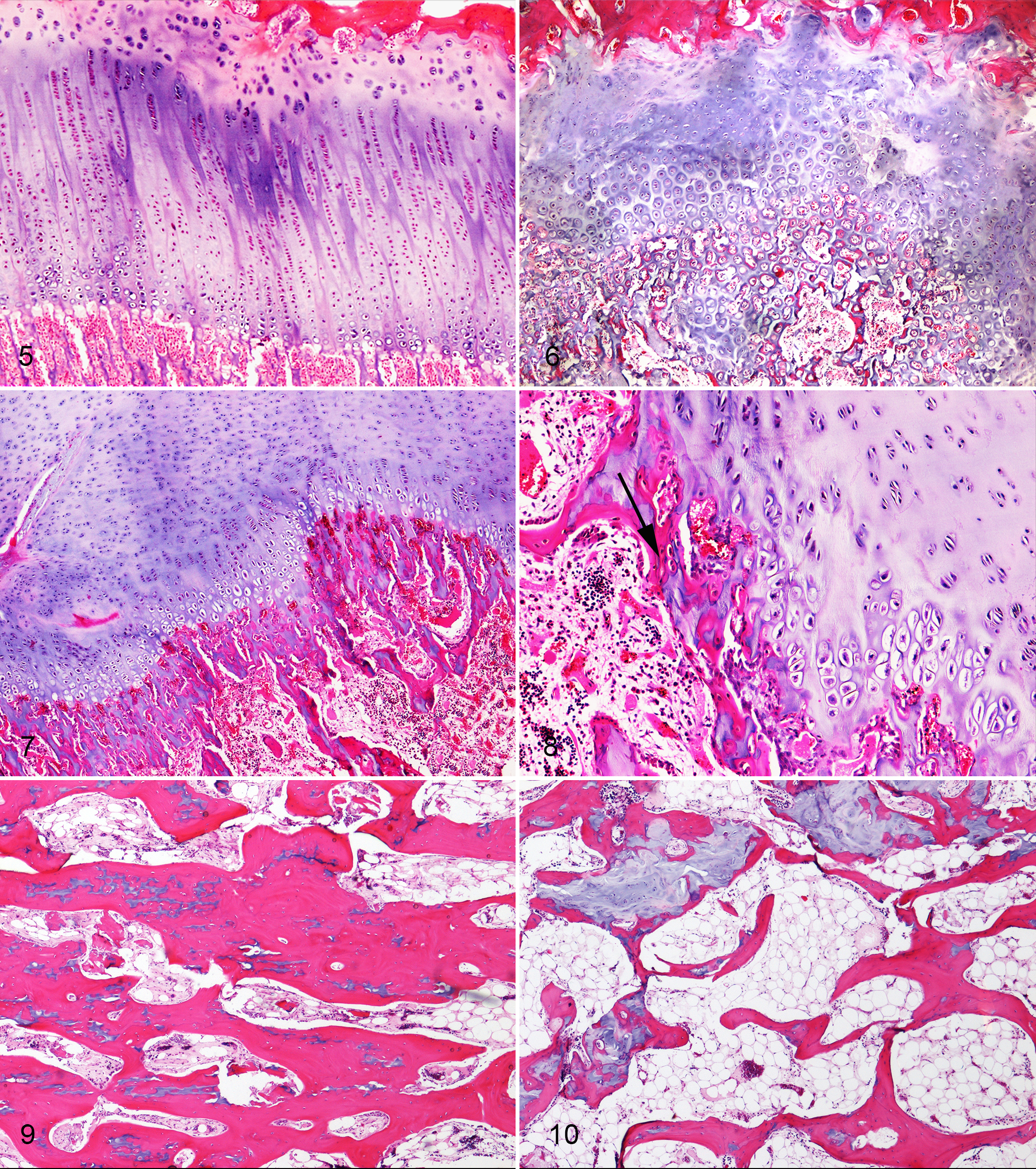
Ellis–van Creveld (EvC), bone, calf. Physes from a control calf and calves affected by EVC syndrome. Hematoxylin and eosin.
Changes to the physeal proliferative and hypertrophic chondrocyte zones were variable. In the proximal humerus of calf No. 1, there was expansion of resting zone chondrocytes; the proliferative and hypertrophic zones were not clearly demarcated; and both zones appeared thinner than normal (Fig. 7). While in some animals, there was decreased thickness of the proliferative zone, in the other animals, the thickness of these zones was normal (when compared to those of a normal control; Fig. 5); however, the chondrocytes were often arranged haphazardly and sometimes in small clusters rather than in columns (Fig. 6).
In the primary spongiosa, the most common lesion was truncation of metaphyseal trabeculae (Fig. 6) and mild disorganization of the primary spongiosa. While calcified cartilage spicules were lined by osteoid, many were truncated immediately below the growth plate and did not fuse into larger trabeculae in the deeper primary spongiosa (Fig. 7). Often trabeculae extended transversely below the growth plate (Fig. 8), linking the short stubby trabeculae. When compared to the control bone (Fig. 9), in the secondary spongiosa of affected calves, remodeling was minimal, and thin trabeculae of bone were linked in a honeycomb-like pattern without aligning along the lines of the forces applied to the bone (Fig. 10).
In the cortex of the distal femur of calf No. 3, the laminar cortical bone had not undergone compaction. Bone directly beneath the periosteum was lined by plump osteoblasts; however, laminar bone deep to this was separated by loose connective tissue and not obviously lined by osteoblasts.
Immunohistochemical Findings
In bone sections of unaffected breed-matched calves, collagen II was expressed uniformly, with a diffuse distribution and strong intensity in the cartilage matrix of all 3 zones of the growth plate and in the cartilage matrix of the primary and secondary spongiosa (Fig. 11). Diffuse, moderate to strong reactivity to collagen II was detected in the resting, proliferative, and hypertrophic zones; in the primary spongiosa; and in the cartilage spicules retained in the trabeculae of the secondary spongiosa in case Nos. 1, 2, and 3 and in the controls. In case No. 4, strong, uniform, diffuse reactivity to collagen II was detected in only the physis of the rib. In the humerus and femur of case No. 4 and the humerus, femur, and vertebra of case No. 5, reactivity was not seen in the resting zone of the physis; the intensity of labeling was weak in the proliferative and hypertrophic zones (Fig. 12); but there was strong reactivity in cartilage spicules retained in trabecular bone of the primary spongiosa. In case No. 5, collagen II showed 2 patterns of reactivity in the hypertrophic zone of the physis: when collagen II was expressed in the matrix, it formed a perichondrocyte ring, and the chondrocyte cytoplasm was unlabeled; but when collagen II was expressed in the cytoplasm of chondrocytes, a perichondrocyte ring did not label (Fig. 13).
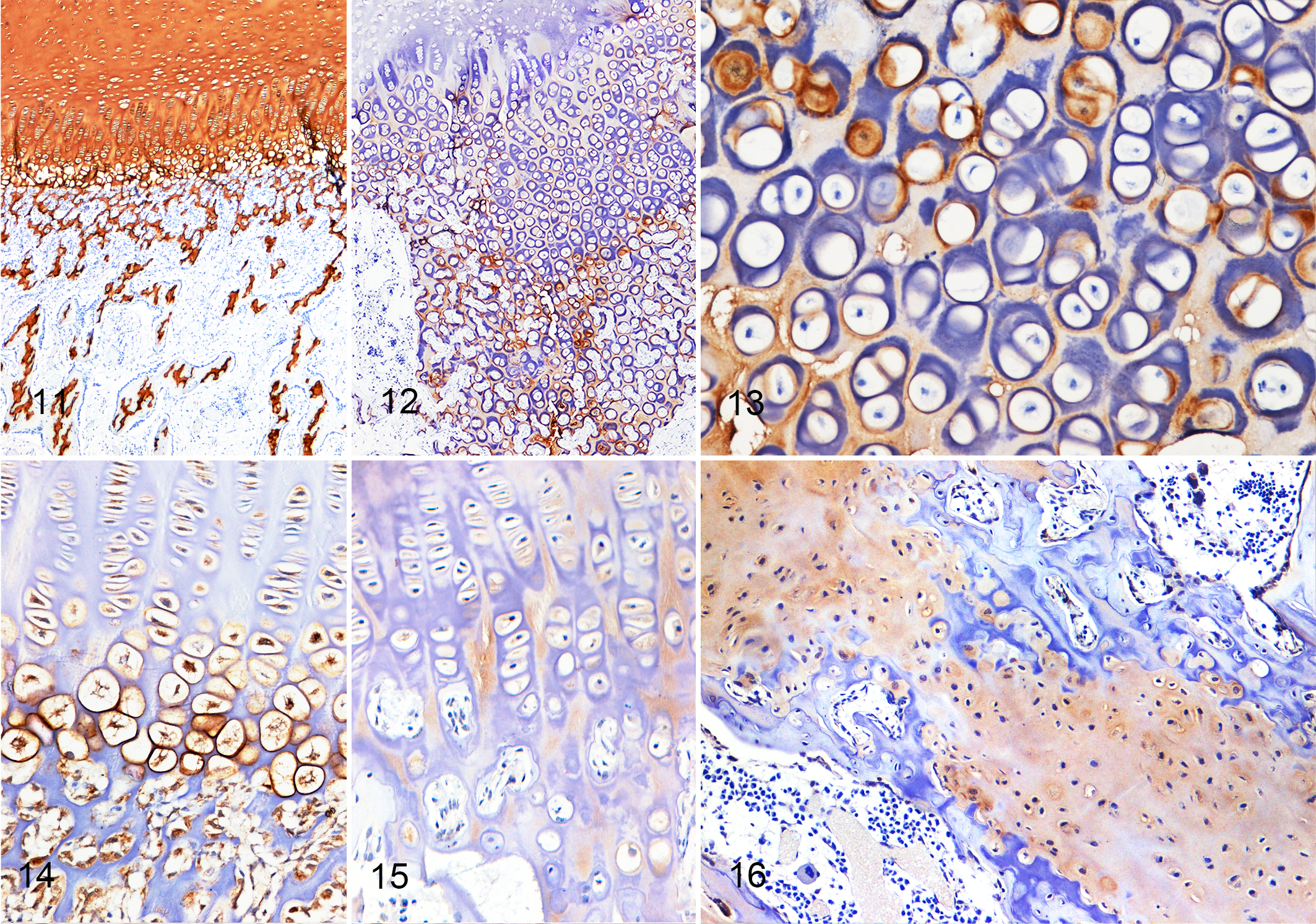
Ellis–van Creveld, bone, calf; immunolabeling for collagen II, DAB chromogen.
In bone sections of unaffected breed-matched calves, collagen X was restricted to the hypertrophic zone of the physis, where it was expressed in the matrix around hypertrophic chondrocytes, forming strong perichondrocyte rings (Fig. 14). It also was diffusely expressed in the cytoplasm of prehypertrophic and hypertrophic chondrocytes.
In all 5 chondrodysplastic cases, expression of collagen X was lost in the pericellular rings around chondrocytes of the hypertrophic zone, where reactivity was only rarely detected. Similar to controls, expression of collagen X was multifocal in the cytoplasm of prehypertrophic and hypertrophic chondrocytes, but inconsistent weak expression in the matrix of the hypertrophic zone of the physis was found in affected animals (Fig. 15). Strong and diffuse labeling of the cartilage matrix of primary spongiosa and cartilage spicules was detected in trabecular bone of the secondary spongiosa of the chondrodysplastic cases (Fig. 16).
In bone sections of unaffected breed-matched calves, SHH was diffusely expressed in the cytoplasm of prehypertrophic and hypertrophic chondrocytes and had weak expression in the matrix of the primary spongiosa and cartilage spicules of the secondary spongiosa. In calves affected by chondrodysplasia, expression of SHH in the cytoplasm of prehypertrophic and hypertrophic chondrocytes was multifocal to diffuse, with variably intense expression in the cartilage matrix of the primary and secondary spongiosa. Reactivity in the cytoplasm of proliferative chondrocytes was detected only in the rib of calf No. 4, where it was weak and multifocal.
In the normal growth plate, in both control calves, FGF2 was diffusely expressed in the cytoplasm of resting, proliferative, prehypertrophic, and hypertrophic chondrocytes. In affected calves, FGF2 expression was lost in the majority of the sections. In calf No. 4, multifocal labeling was observed in the cytoplasm of resting and proliferative chondrocytes, and calf No. 1 showed positive reactivity in the cytoplasm of hypertrophic chondrocytes but not resting or proliferative chondrocytes.
Ki67 showed multifocal nuclear labeling mainly in the chondrocytes of the proliferative zone and occasionally in the resting chondrocytes of the control calves. The proliferative indices of the 2 control calves were 19.7% and 12.1%. Ki67 was expressed at a lower level in the proliferative zone of the affected calves with chondrodysplasia, and the average proliferative index of the chondrodysplastic calves was 7.4%. The immunohistochemical results are summarized in Table 1.
Immunohistochemical Results by Histologic Region of the Physis and Metaphysis.

Abbreviations: +, positive; –, negative; +/–, variably positive; –/+, mostly negative but rarely positive; EVC2, affected animals with EVC2 mutation; FGF, fibroblast growth factor; MHZ, matrix hypertrophic zone.
Expression of EVC and EVC2 Transcripts
RNA was extracted from several tissues of an affected calf (No. 5) and 1 healthy control (No. 6). RT-PCR was performed with primers targeting a region that included exons 17 to 19 in bovine EVC and exons 18 to 20 for EVC2 (targeting the region containing the c.2993_2994ACdel mutation). In samples obtained from the affected calf, RT-PCR revealed the presence, as expected, of the EVC transcript, but interestingly the EVC2 transcript also was detected (Fig. 17), which was confirmed by Sanger sequencing. We confirmed the presence of both EVC and EVC2 transcripts, detected through RT-PCR, in the samples obtained from the control animal (Fig. 18).
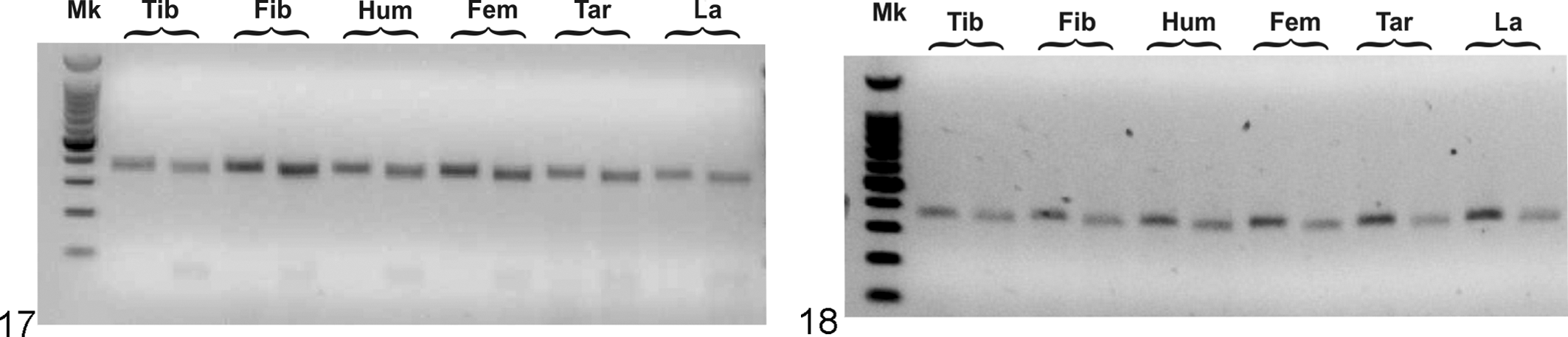
Tissue-specific gene expression in the case of Ellis–van Creveld (EvC; Fig. 17) and the control calf (Fig. 18), showing polymerase chain reaction–amplified fragments of cDNA prepared from several tissues. For each organ, the product from the polymerase chain reaction targeting the EVC1 cDNA sequence (352 base pairs) is shown on left, and the one targeting the EVC2 sequence (342 base pairs) is on the right. Fem, femur; Fib, fibula; Hum, humerus; La, laminae of the hoof; Mk, 100–base pair ladder; Tar, tarsus; Tib, tibia.
Discussion
Chondrodysplasia caused by an autosomal recessive mutation of the Limbin gene (orthologous to human EVC2) was described for the first time in the Japanese brown breed. 25 A genetic study of inherited chondrodysplasia caused by a deletion in the EVC2 gene was recently reported in Grey Alpine cattle, 16 and in the present article, we describe the gross, histologic, immunohistochemical, and molecular hallmarks of EvC in this breed.
The necropsy findings of calves with deletion in the EVC2 led to a morphologic diagnosis of disproportionate dwarfism (chondrodysplasia). The main changes are related to the skeletal system, and the long bones were particularly affected; they appeared short, bulky, rotated, arched in a “dumbbell-like” form, and severely reduced in length, with a very short diaphysis, compared to normal calves.
Several types of inherited chondrodysplasia occur in cattle, including “bulldog” chondrodysplasia, Telemark, brachycephalic, dolichocephalic, and rhizomelic types. 27 Based on the necropsy findings, the chondrodysplasia in Grey Alpine calves is not similar to the brachycephalic type of chondrodysplasia, because of the absence of brachycephaly and less severe involvement of metacarpal bones. Our cases are most similar to the rhizomelic type of chondrodysplasia because the lesions are confined to the long bones of the limbs, with prominent shortening of the forelimbs, as described in Japanese Brown cattle with chondrodysplasia. 27
In humans, EvC involves multiple organs and is characterized by cardiovascular malformation in 60% of patients, 24 the most common of which are atrioventricular septal defects and common atrium. 20 Similarly, 2 calves of our case series were affected by cardiac lesions, including left atrioventricular valvular dysplasia in a 6-month-old calf and an atrial septal defect (patent foramen ovale) in a 2-month-old calf. The prognosis of human EvC syndrome is related to the severity of the cardiovascular malformation. 20 Cryptorchidism (as seen also in calf No. 5), hypospadias, and mucometra also have been reported in humans with EvC, but such genital lesions are rare. 2
The mammalian growth plate, also known as the physis, is a highly specialized mesoderm-derived cartilaginous structure. Each growth plate is a sandwich-like, multilayer structure divided into 4 functional zones: reserve, proliferative, hypertrophic, and calcified. 4 The last zone is then converted into the primary spongiosa, which is remodeled into the secondary spongiosa. 4 Chondrocytes are metabolically active cells that synthesize various elements of the extracellular matrix, 9 including collagen II, which is used as a marker of cartilage differentiation because it is expressed in cartilage matrix. 7 Fibrillar cartilage-specific collagen type II is the major component of the cartilage extracellular matrix. 14
Human fetal and postnatal (18-month-old child) articular cartilage has homogenous labeling for collagen II. 7 Immunohistochemical examination of the physis in Japanese brown cattle with chondrodysplasia showed type II collagen immunoreactivity in the cartilage matrix throughout the epiphyseal growth plate and within the primary bone trabeculae. 23 Similarly, in 3 of our cases, there was diffuse, moderate to strong labeling in the resting, proliferative, and hypertrophic zones; in the primary spongiosa; and in cartilage spicules retained in the trabeculae of the secondary spongiosa. The remaining 2 cases, however, lacked immunolabeling in the resting zones and had a weak intensity of labeling in the proliferative and hypertrophic zones, with strong intensity in cartilage spicules retained in trabecular bone. During endochondral ossification, the aggrecan- and type II–collagen rich cartilage is resorbed. 13 As chondrocyte hypertrophy occurs, synthesis of type II collagen is downregulated, and synthesis of the nonfibrillar type X collagen is initiated. A peptide derived from proteolytic degradation of type II collagen induces chondrocyte hypertrophy, as assessed by expression of collagen type X. 14 Our data suggest that in some animals with EVC2 chondrodysplasia, collagen II may be prematurely degraded; its expression was decreased in the resting zone of both a 2-month-old calf and a 12-month-old calf, suggesting that the decrease in expression is not age related but instead may be due to accelerated differentiation of physeal chondrocytes, early hypertrophy, and endochondral ossification. A larger number of cases would be necessary to further confirm these hypotheses.
Type X collagen is a marker of cartilage differentiation, a nonfibrillar component of the matrix specific for hypertrophic chondrocytes. 17 Maturation of chondrocytes occurs due to physical and biochemical changes that occur in a spatial and temporal pattern. 4 The role of type X collagen is to facilitate the deposition of calcium within the matrix. 1 As hypertrophic chondrocytes become terminally differentiated, the gene Col10a1, which encodes for collagen X, ceases to be expressed in chondrocytes, leading to activation of a group of genes that induce extracellular matrix mineralization and cellular apoptosis. 4 Collagen X and cellular hypertrophy together are indicators of terminally differentiated growth plate chondrocytes. 4
Collagen X is distributed in the hypertrophic zone of the fetal physis, around blood vessels, and in the perichondrium. 7 In Japanese brown chondrodysplastic cattle, collagen X is expressed in hypertrophic chondrocytes of the physis. 23 In our cases, a general loss of collagen X expression was noted in the perichondrocyte rings in the hypertrophic zone, with collagen X immunolabeling limited to the cytoplasm of prehypertrophic and hypertrophic chondrocytes and the cartilage matrix of the primary and secondary spongiosa. Our data suggest that EvC-affected calves have a disorderof chondrocyte differentiation, which may indicate an accelerated differentiation and premature hypertrophy.
SHH plays a role in differentiation and growth plate organization by indirectly activating the functional receptor Smo (transmembrane protein smoothened). The latter recruits EVC/EVC2 to activate Gli protein transcription factor. 3,30 In the control calf, SHH immunolabeling was in the cytoplasm of prehypertrophic and hypertrophic chondrocytes but weak in the matrix of the primary spongiosa and cartilage spicules of the secondary spongiosa. No significant differences between control and dwarf calves were found. Detailed analysis of the long bones of transgenic mice that overexpress SHH showed that chondrocyte differentiation was blocked at prehypertrophic stages, hindering endochondral ossification and trabecular bone formation. 26 Chondrocytes of transgenic mice also had increased expression of the SHH receptor (Patched-1), PTHrP, and SOX9. 26 Tavella et al. demonstrated that continuous expression of SHH in chondrocytes of transgenic mice interferes with cell differentiation and growth plate organization. 26 However, further studies are necessary to better elucidate the precise role of EVC and limbin in modulating the response to hedgehog ligands. 20
Ki67 is a marker of cell proliferation and can be used as a measure of the proliferative activity of the growth plate. The growth plate of EvC-affected calves had a proliferation index markedly lower than the control, suggestive of reduced chondrocyte proliferation, paralleling the shortening of the proliferative zone observed histologically.
Basic FGF2 is a prototype member of the FGF family and is found in the resting, 11 proliferative, and hypertrophic zones of the growth plate. 17 It is a potent ligand for all 4 FGFRs. Overexpression of FGF2 in mice causes shortening and flattening of long bones and widening of the growth plate, with decreased hypertrophic chondrocytes. 17 FGF2 has been shown to be one of the main FGF family members expressed by chondrocytes of the physis. 12 FGF2 was detected in the resting zone, proliferative zone, and prehypertrophic and hypertrophic zones of our control calves, paralleling what has been reported in normal human and animal model physes. 11,17
In calf No. 4, multifocal expression of FGF2 was found in the resting and proliferative zones, and in calf No. 1, there was multifocal labeling of the cytoplasm of hypertrophic chondrocytes. In the majority of affected calves, there was a loss of expression of FGF2 in the growth plate zones, compared to control subjects. In the literature, it has been reported that one of the roles of FGF2 is to induce the proliferation of chondrocytes. 17 Rats treated intravenously with FGF2 showed a substantial increase in the number of proliferating cell nuclear antigen–positive chondrocytes. 17 In our study, the combination of decreased Ki67 proliferation index and loss of expression of FGF2 suggests that downregulation of FGF2 could be responsible for the reduced proliferation index, which was additionally visible as a shorter proliferative zone microscopically. Further studies on FGF signaling are needed to elucidate this possible role of FGF2.
We designed RT-PCR experiments using primers aimed at the EVC and EVC2 transcripts, which we were able to detect. Comparing the bands in Figure 17, we can observe a comparable intensity for both transcripts (EVC and EVC2) in all the analyzed tissues. With the same PCR conditions (and roughly the same starting amount of RNA for cDNA synthesis), we do not observe dramatic differences between affected case and control (RT-PCR control in Fig. 18). The very low number of biological replicates prevents a thorough semiquantitative comparison; nonetheless, we can assume that in the affected tissues, the EVC2 transcript is expressed at detectable and comparable levels with EVC, as well as with the control. This was a somewhat surprising result because we did not expect to find any detectable EVC2 transcript in the affected animals. In fact, it has been suggested that mutations EVC and EVC2 causing EvC would lead to nonsense-mediated decay and ultimately to the destruction of the mutant mRNA transcript. 24 This would explain the recessive inheritance for EvC: the nonfunctional transcripts would be destroyed, and only the functional ones would be translated. In the case of 2 faulty transcripts (homozygosity), the pathologic condition would arise. 24 This is very different from the dominantly inherited Weyer syndrome, in which the one mutant allele is supposed to result in production of a defective protein that has a dominant negative function on the signaling complex (the defective protein disrupts normal binding even of wild-type proteins) and thus triggers the syndrome. 29 We suggested this explanation as the most likely cause for the recessive nature of the dwarfism in Grey Alpine calves in our previous paper. 16 However, our current data show the presence of EVC2 transcript at detectable levels in many tissues of an affected animal (Fig. 17), comparable to the control (Fig. 18). This suggests that the mutant mRNA could be translated (Fig. 17), thereby making our initial theory of nonsense-mediated decay and ultimate destruction of the transcript unlikely. Since we have evidence of the recessive nature of the dwarfism in Grey Alpine calves, perhaps the most likely scenario is that the protein produced by the c.2993_2994ACdel mutation is not able to act in a dominant negative manner. Nonetheless, the product could be heavily misfolded, resulting in its targeting by the proteasome and subsequent destruction; thus, it may be unable to negatively affect signaling in the cell. 24 In other words, we speculate that if the transcript that we detect is translated, a posttranslational regulatory mechanism could intervene to “exclude” the faulty protein from signaling.
The reduced expression of collagen II in resting areas suggests that it is prematurely degraded, leading to the secretion of peptides required for induction of chondrocyte hypertrophy. The lack of expression of collagen X in the matrix around the hypertrophic chondrocytes is suggestive of fully differentiated, adult cartilage. The low proliferation index in affected calves further reinforces this hypothesis.
In conclusion, the histologic features indicative of terminal differentiation of the physis—associated with premature collagen II degradation, abnormal collagen X expression, a low proliferation index, with a loss of expression of FGF—are all findings suggesting that the pathogenesis of EvC syndrome may involve a reduced proliferation, an early hypertrophy, and accelerated differentiation of physeal chondrocytes leading to early ossification. Our detection of EVC2 transcripts shows that, contrary to what has been suggested, the EVC2 mutant transcript is present in affected tissues and is not destroyed by nonsense-mediated decay. Thus, it is likely that other mechanisms (perhaps mutant protein degradation) may explain the lack of a functional protein. Furthermore, the similarities identified with the human EvC condition suggest that bovine EvC syndrome would be a useful spontaneous model for humans with EvC syndrome.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.