
Abstract
Breast cancer is the most common cancer and the second-leading cause of cancer mortality in women in the United States. A recent 2-year National Toxicology Program carcinogenicity study showed an increased incidence of proliferative mammary lesions (hyperplasia, fibroadenoma, adenocarcinoma) in F344/NTac rats exposed to bromodichloroacetic acid (BDCA), a disinfection by-product in finished drinking water with widespread human exposure. We hypothesized that the increase in mammary tumors observed in BDCA-exposed F344/NTac rats may be due to underlying molecular changes relevant for human breast cancer. The objective of the study was to compare (1) gene and protein expression and (2) mutation spectra of relevant human breast cancer genes between normal untreated mammary gland and mammary tumors from control and BDCA-exposed animals to identify molecular changes relevant for human cancer. Histologically, adenocarcinomas from control and BDCA-exposed animals were morphologically very similar, were estrogen/progesterone receptor positive, and displayed a mixed luminal/basal phenotype. Gene expression analysis showed a positive trend in the number of genes associated with human breast cancer, with proportionally more genes represented in the BDCA-treated tumor group. Additionally, a 5-gene signature representing possible Tgfβ pathway activation in BDCA-treated adenocarcinomas was observed, suggesting that this pathway may be involved in the increased incidence of mammary tumors in BDCA-exposed animals.
Keywords
Breast cancer is the most common cancer and the second-most common cause of cancer-related death in women in the United States, with an estimated 232 340 new cases and 39 620 deaths in 2013. 3,57 Up to 10% of breast cancers are characterized as hereditary, 37 while the remaining majority of tumors (90%–95%) are sporadic, involving complex and multifactorial etiologies, including dietary, hormonal, environmental, and lifestyle factors. 38 The complexity of this very heterogeneous disease is exemplified by the marked complexity of the molecular changes accompanying its pathogenesis. 30,64 Prognosis is typically based on a variety of factors, including hormone receptor status and luminal/basal immunophenotype of the tumor cells. Specifically, tumors that lack expression of hormone receptors (estrogen receptor alpha [ERα], progesterone receptor [PR]) and those that express predominantly basal markers (cytokeratin 5 [CK5], CK14, alpha smooth muscle actin [α-SMA]) are associated with a particularly poor prognosis, whereas those that retain hormone receptor function and have a luminal phenotype (CK8, CK18, CK19) have a more favorable prognosis. 11,25,26
Since the pathogenesis of the majority of breast cancers is multifactorial, the impact of environmental exposures that pose a potential human health hazard is critical. Environmental factors are an overwhelming contributor to the causation of cancer in humans. 36 A number of environmental contaminants and occupational exposures have been linked to an increased risk of mammary cancer in humans and/or rodent models. 24,66 To date, the National Toxicology Program (NTP) has identified 51 chemicals associated with mammary tumors in rodents, with 37 showing clear evidence of carcinogenicity in either the rat or mouse (http://ntp.niehs.nih.gov/go/SA-39). 47 Five chemicals were positive in both species (acrylamide, chloroprene, 1,2-dibromomethane, 1,2-dichloroethane, glycidol, and sulfallate). Chemically induced mammary gland tumors generally occurred more commonly in the rat, which is particularly relevant because many aspects of mammary carcinogenesis in the rat are similar to those in humans. 15,25,56 A recent 2-year NTP carcinogenicity study (TR-583) showed a dose-dependent increase in the incidence of proliferative mammary lesions (hyperplasia, fibroadenoma, adenocarcinoma) in F344/NTac rats exposed to bromodichloroacetic acid (BDCA) in drinking water. 50 The F344/NTac rat is a substrain of the F344/N rat and was used by the NTP in the BDCA chronic bioassay. We consider both the F344/N and the F344/NTac to be closely related, and the findings in this study apply to both rat strains. These strains have very similar genetic backgrounds since they are derived from the same founder strain. Following a workshop with input from several external experts, the NTP chose to continue the use of the F344/NTac rat in chronic bioassays of some chemicals, including the current BDCA study, because this rat strain is derived from the same parent stock and is genetically closest to the F344/N rat strain. 32 So, while there is a possibility that the spontaneous tumor incidence and underlying molecular mechanisms may differ between these 2 closely related rat strains, their very similar genetic backgrounds minimize this possibility.
BDCA is a disinfection by-product that is a contaminant of finished drinking water, formed by the reaction of oxidizing agents containing chlorine with naturally occurring organic material and bromide ions in source water. 54 It is also one of the most common disinfection by-products in surface drinking water supplies in the United States. 34 BDCA is therefore widespread in drinking water sources, and individuals are exposed to this compound in tap water through oral and dermal routes from oral consumption, cooking, bathing, and swimming. 50 A similar compound, bromochloroacetic acid, which is also a by-product in the disinfection of drinking water, was shown to induce mammary tumors in both male and female F344/N rats and B6C3F1 mice. 49 BDCA is not one of the haloacetic acids currently regulated by the Environmental Protection Agency, 50 and due to potential widespread human exposure, determination of whether they pose a possible human health risk is critical.
Sporadic breast cancer in humans follows a paradigm of progressive acquisition of various molecular alterations, including loss of tumor suppressor genes and overexpression of oncogenes and growth pathways in the progression from premalignant lesions to invasive cancer. 4,31 Fibroadenomas are the most common tumor in the mammary gland of humans and rats, and it is suspected that some mammary adenocarcinomas may arise from fibroadenomas in both species. 40 In NTP studies, the F344/N rat develops a background rate of mammary fibroadenomas and adenocarcinomas of 51.4% and 3.9%, respectively. 48 According to the Registry of Industrial Toxicology Animal database—an industry-sponsored database of peer-reviewed historical control tumor incidences—background mammary tumors (adenocarcinoma, adenocarcinoma arising in fibroadenoma, adenoma, fibroadenoma) are also fairly common in the Sprague-Dawley (58%) and Wistar rats (24%). 22 Separating spontaneous lesions from treatment-related lesions can be difficult because of these background incidences, particularly in studies in which a treatment-related tumor response is not obvious, since the level of spontaneous lesions present in treated groups is typically dictated by the number of spontaneous lesions in control animals. 39 Thus, identification of molecular markers that separate spontaneous from chemically induced tumors is of great importance.
The identification of genetic alterations in chemically induced rodent tumors that are similar to those observed in human cancer lends further support that they are of potential human health risk. 24 For example, overexpression and/or mutation of the epidermal growth factor receptor (EGFR) is associated with cell proliferation, angiogenesis, and tumor progression in human breast cancer. 16 Similarly, loss of function mutations in the phosphatase and tensin homologue (PTEN) tumor suppressor gene occur in a variety of human neoplasms, including breast cancer, and are associated with hormone resistance. 43 TP53 mutations are also common in triple-negative breast cancer and are associated with epithelial-mesenchymal transition. 9 The constitutive activation of the RAS signaling pathway is one of the most common alterations in human breast cancer, and while mutation of the HRAS gene is uncommon, overexpression of the protein is found in 60% to 70% of cases. 44 Finally, CTNNB1 plays an important role in normal development, as well as neoplastic transformation in the rodent and human mammary gland, and is often associated with hormone receptor–negative breast cancers in people. 27
Given the increased incidence of mammary tumors in F344/NTac rats exposed to BDCA and the widespread human exposure in drinking and bathing water, the objective of this study was to (1) immunophenotype mammary tumors from control and BDCA-exposed animals based on human criteria and (2) identify molecular changes in BDCA-exposed rats that are of relevance for human cancer risk. We hypothesized that the increased incidence of mammary tumors in BDCA-exposed animals may be associated with alterations in molecular pathways that are relevant for human cancer, including breast cancer. Identification of such relevant alterations would further strongly suggest that chronic exposure to this chemical poses a human cancer risk.
Materials and Methods
Animals, Histology, and Immunohistochemistry
In the 2-year NTP bioassay, dose groups of 50 male and 50 female F344/NTac rats were exposed to 0, 250, 500, and 1000 mg/L of BDCA in drinking water per dose group. Routine tissues and tumors, including adenocarcinomas and fibroadenomas from BDCA-exposed female F344/NTac rats, were collected at the end of the bioassay. Mammary gland lesions were diagnosed per NTP standardized nomenclature and criteria. 5 Tumor incidence and statistics were performed as previously described. 50 Collection of paired frozen tissue from mammary tumors was performed according to criteria based on incidence, size (>0.5 cm in diameter), and viability (minimal hemorrhage and necrosis). At necropsy, tumors were sectioned in half: one half was flash frozen in liquid nitrogen for gene expression analysis, and the other half was fixed in 10% neutral buffered formalin for histology and immunohistochemistry. While there was adequate spontaneous fibroadenoma frozen tissue available, there was a lack of frozen spontaneous adenocarcinoma tissue available from this study due to the lower tumor incidence. Therefore, spontaneous adenocarcinomas were collected from 5 age-matched vehicle control female rats from other chronic (2-year) NTP studies in F344/N rats (nickel sulfate hexahydrate; D&C Yellow 11; 2,2-bis(bromomethyl)-1,3, propanedial; 3,3′,4,4′-tetrachloroazobenzene). In addition, frozen vehicle-control mammary gland was not available from the BDCA study, so mammary glandular units (epithelium, myoepithelium, stroma) were collected by laser microdissection from 5 age-matched female F344/N rats obtained from the National Institute of Aging. Since normal mammary gland is predominantly adipose tissue, this sampling was considered to more appropriately represent the predominant cellular populations in normal mammary tissue that would be comparable to those in mammary tumor tissue. Formalin-fixed, paraffin-embedded (FFPE) tumor samples were selected for immunohistochemistry based on the availability of paired collected frozen tissue such that each FFPE tumor could be matched to its respective frozen sample used for gene expression analysis. Furthermore, it was necessary to use paired FFPE samples for immunohistochemistry, as these tissues are processed immediately; FFPE samples not paired with frozen samples were subject to prolonged fixation (>4 weeks) and were thus not acceptable for immunohistochemistry. Seven FFPE normal mammary gland samples, 8 FFPE mammary adenocarcinomas (4 BDCA treated, 4 spontaneous), and 7 FFPE fibroadenomas (4 BDCA treated, 3 spontaneous) paired with frozen samples were used for immunohistochemistry based on these criteria. Samples collected for gene expression analysis, mutation analysis, and immunohistochemistry are listed in Supplemental Table 1.
Proliferative Mammary Gland Lesions in Control and BDCA-Exposed Female Rats in the 2-Year National Toxicology Program Bioassay.a
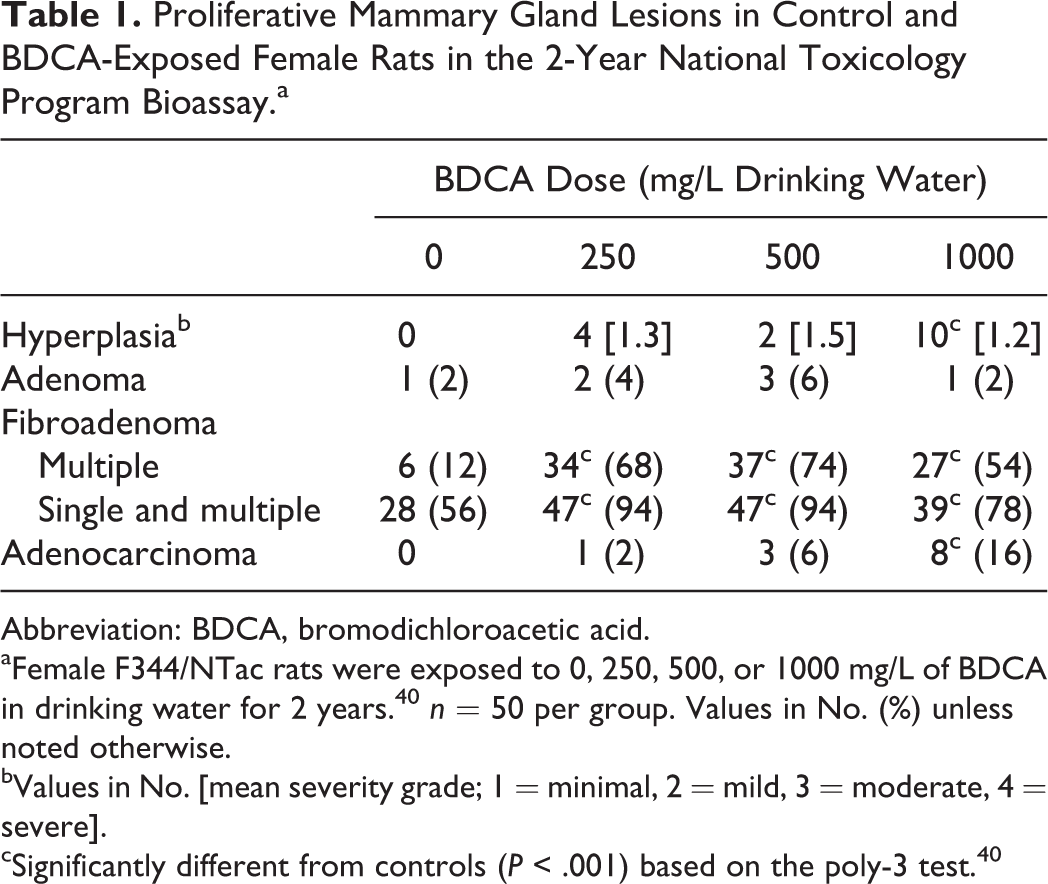
Abbreviation: BDCA, bromodichloroacetic acid.
aFemale F344/NTac rats were exposed to 0, 250, 500, or 1000 mg/L of BDCA in drinking water for 2 years. 40 n = 50 per group. Values in No. (%) unless noted otherwise.
bValues in No. [mean severity grade; 1 = minimal, 2 = mild, 3 = moderate, 4 = severe].
cSignificantly different from controls (P < .001) based on the poly-3 test. 40
Paraffin-embedded samples were sectioned at 5 µm and stained with hematoxylin and eosin or immunostained with the following antibodies using routine methods: mouse monoclonal anti-ERα (1:50, Beckman Coulter Inc., Brea CA), mouse monoclonal anti-PR (1:150, Beckman Coulter Inc), rabbit polyclonal anti-SMA (1:150, Abcam, Cambridge, MA), mouse monoclonal anti-CK14 (1:50, Abcam), and mouse monoclonal anti-CK18 (1:800, Santa Cruz Biotechnology, Santa Cruz, CA). Normal rat endometrium was used as a positive control for ERα, PR, SMA, and CK18, and normal rat skin was used as a positive control for CK14. Substitution of the primary antibody with normal serum from the species in which the primary antibody was raised was used for negative controls. Slides were washed, then labeled using the streptavidin ABC technique for detection of the primary antibody-specific protein. DAB (3,3-diaminobenzidine) was used to visualize all immune reactions, and slides were counterstained with Mayer’s hematoxylin. The sections were dehydrated through graded alcohols, immersed in xylene, and mounted with coverslips. Positive immunoreactivity to ERα and PR was defined as nuclear staining, and cytoplasmic and membrane immunoreactivity was considered positive for CK14, CK18, and SMA.
Real-Time Quantitative Polymerase Chain Reaction Arrays and Data Analysis
RNA was extracted from frozen samples from female rats (5 laser-captured samples of mammary epithelium from vehicle control, age-matched mammary gland, 3 spontaneous fibroadenomas from vehicle control rats, 4 fibroadenomas from BDCA-exposed rats, 5 spontaneous adenocarcinomas from vehicle control rats, and 4 adenocarcinomas from BDCA-exposed rats) using the Invitrogen TRIzol Kit (Supplemental Table 1). Fold increases and decreases in gene expression were determined by quantification of cDNA from spontaneous and BDCA-treated tumors relative to normal mammary epithelium from age-matched controls. The 18s RNA gene was used as the endogenous control for normalization of initial RNA levels. A rat polymerase chain reaction (PCR) array (PARN-131Z, SA Biosciences, Frederick, MD) representing 84 genes important in breast cancer pathogenesis was used to identify differential gene expression between spontaneous and BDCA-treated tumors. Quantitative differential gene expression levels were detected using arrays containing corresponding PCR primers and SABiosciences SYBR Green qPCR master mix, and the reactions were run on an ABI PRISM 7900HT Sequence Detection System (Foster City, CA) using the manufacturer’s protocols. Gene expression was normalized to Actb, and fold changes were calculated using the ΔΔCt method. 53 To examine the respective gene changes in the progression from normal to benign (fibroadenoma) to malignant (adenocarcinoma) tumor, a trend analysis was performed to evaluate for an increasing or decreasing trend in gene expression. In addition, pairwise comparisons were performed for spontaneous and BDCA-treated adenocarcinomas (controlling for directional errors), using a residual bootstrap-based methodology 17 to compute P values based on 10,000 bootstrap samples. The statistical significance was obtained by controlling the false discovery rate (FDR) at a nominal 5% level (FDR < 0.05). Trend tests and pairwise comparisons were performed using ORIOGEN 4.01. 19,51,52 Principal component analysis and unsupervised hierarchical cluster analysis were performed via Partek Genomics Suite 6.3 (St Louis, MO).
Gene Mutation Analysis
With a DNeasy Tissue Kit (Qiagen, Valencia CA), DNA was isolated from FFPE sections of 10 mammary fibroadenomas (5 spontaneous, 5 BDCA treated) and 13 adenocarcinomas (5 spontaneous, 8 BDCA treated) from BDCA-exposed female rats (Supplemental Table 1). Amplification reactions were carried out by seminested PCR via primer sets for rat Egfr (exons 18–21), Pten (exons 1–9), Tp53 (exons 5–8), Ctnnb1 (exon 2), and Hras (exon 2; Supplemental Table 2). Controls lacking DNA were run with all sets of reactions. PCR products were sequenced with an automatic sequencer.
Results of Immunohistochemical Analysis of Mammary Tumors From Control and BDCA-Exposed Rats.a
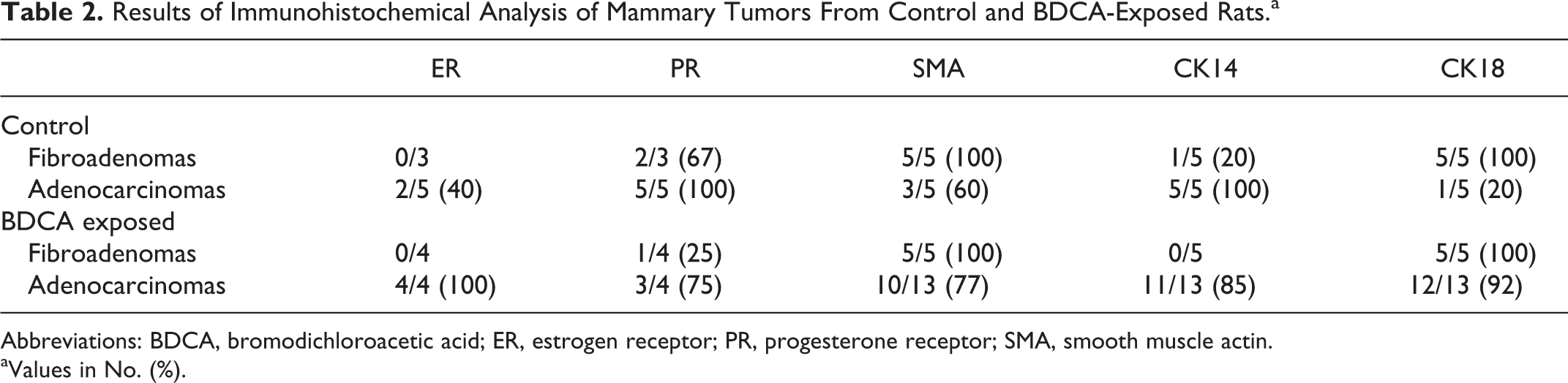
Abbreviations: BDCA, bromodichloroacetic acid; ER, estrogen receptor; PR, progesterone receptor; SMA, smooth muscle actin.
aValues in No. (%).
Results
Exposure of F344/NTac Rats to BDCA in Drinking Water for 2 Years Is Associated With Induction of Mammary Hyperplasia, Fibroadenoma, and Adenocarcinoma
In the original 2-year NTP bioassay, exposure of 50 female F344/NTac rats to BDCA at doses of 250, 500, and 1000 mg/L in drinking water per dose group resulted in a treatment-related increase in the incidence of mammary gland hyperplasia, fibroadenoma, and carcinoma 50 (Table 1). Approximately 56% of vehicle control female rats developed spontaneous fibroadenomas. In contrast to the normal vehicle control mammary gland (Fig. 1), mammary hyperplasia was characterized by proliferation of glandular acini resulting in irregular nodules separated by fine fibrovascular stroma (lobular) and/or proliferation of ductal epithelial cells into multiple layers or papillary projections (ductal; Fig. 2). Mammary fibroadenomas were expansile, well-demarcated proliferations of tubules and acini of well-differentiated glandular epithelial cells separated by abundant fibrous stroma (Fig. 3). Mammary adenocarcinomas were characterized by infiltrative, generally poorly demarcated proliferations of solid lobules and irregular tubules and acini composed of poorly differentiated glandular epithelial cells within scant fibrovascular stroma (Fig. 4), often associated with variable necrosis and inflammatory cells.
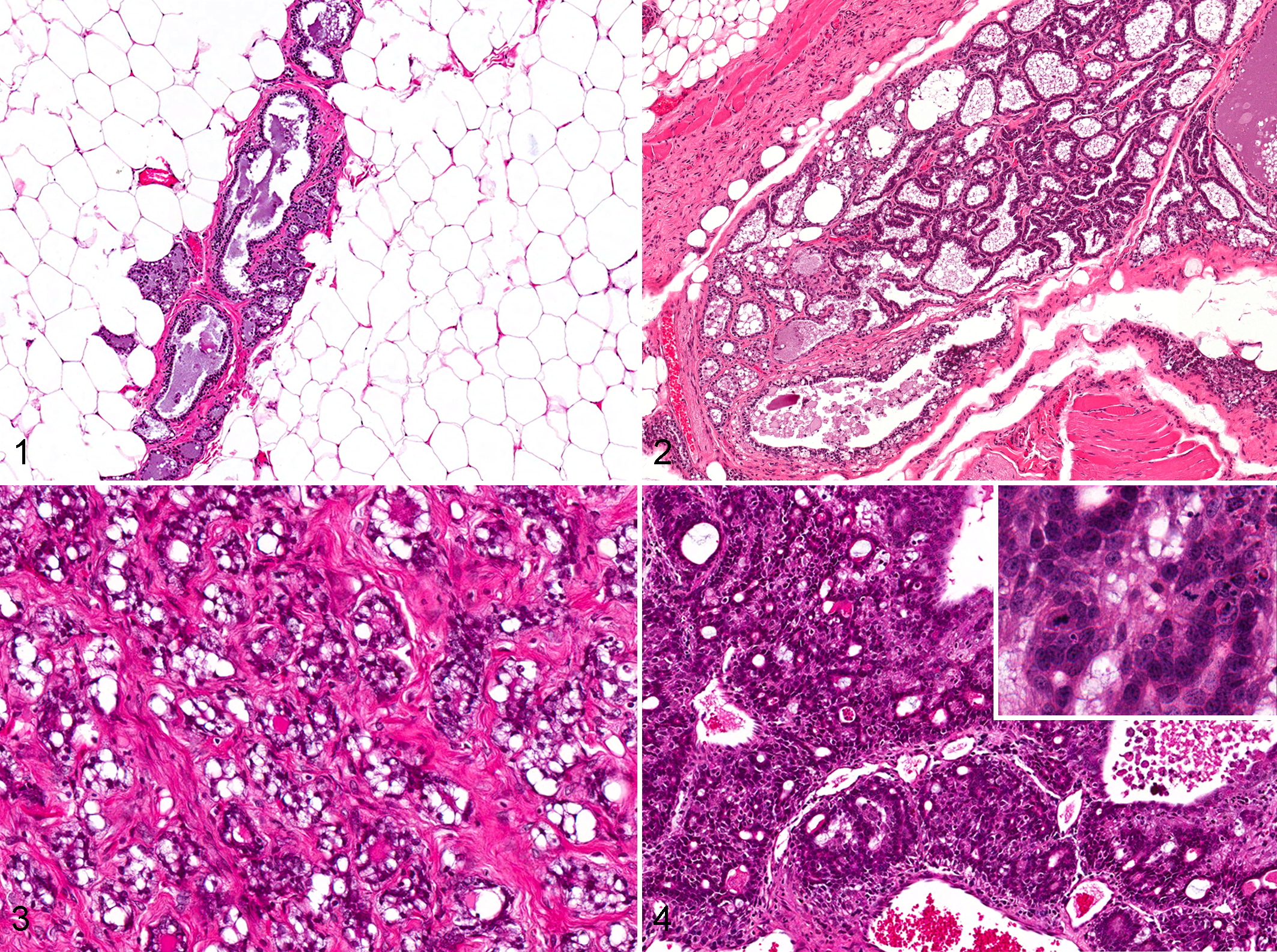
Mammary gland, bromodichloroacetic acid–exposed rat.
Mammary Adenocarcinomas in F344/NTac Rats Harbor a Mixed Luminal/Basal and Hormone Receptor Immunophenotype and Mutations Observed in Human Breast Cancer
Spontaneous and BDCA-treated mammary adenocarcinomas were hormone receptor (ERα/PR) positive and immunoreactive to both luminal (CK18) and basal (CK14, α-SMA) markers (Table 2). Normal mammary gland (Fig. 5a) and BDCA-treated fibroadenomas (Fig. 6a) were negative for ERα, while 4 of 4 (100%) of BDCA-treated adenocarcinomas were positive (Fig. 7a). Spontaneous fibroadenomas were negative for ERα, while 3 of 5 (40%) spontaneous adenocarcinomas were positive for ERα. Normal mammary gland shows a few positive cells for PR (Fig. 5b), and 3 of 4 BDCA-treated fibroadenomas (Fig. 6b) were negative for PR. In contrast, 3 of 4 BDCA-treated adenocarcinomas were positive for PR (Fig. 7b). Two of 3 spontaneous fibroadenomas and 5 of 5 spontaneous adenocarcinomas were PR positive. In general, the number of cells immunoreactive for PR in each tumor was fewer than that for ERα in both spontaneous and BDCA-treated fibroadenomas and adenocarcinomas.
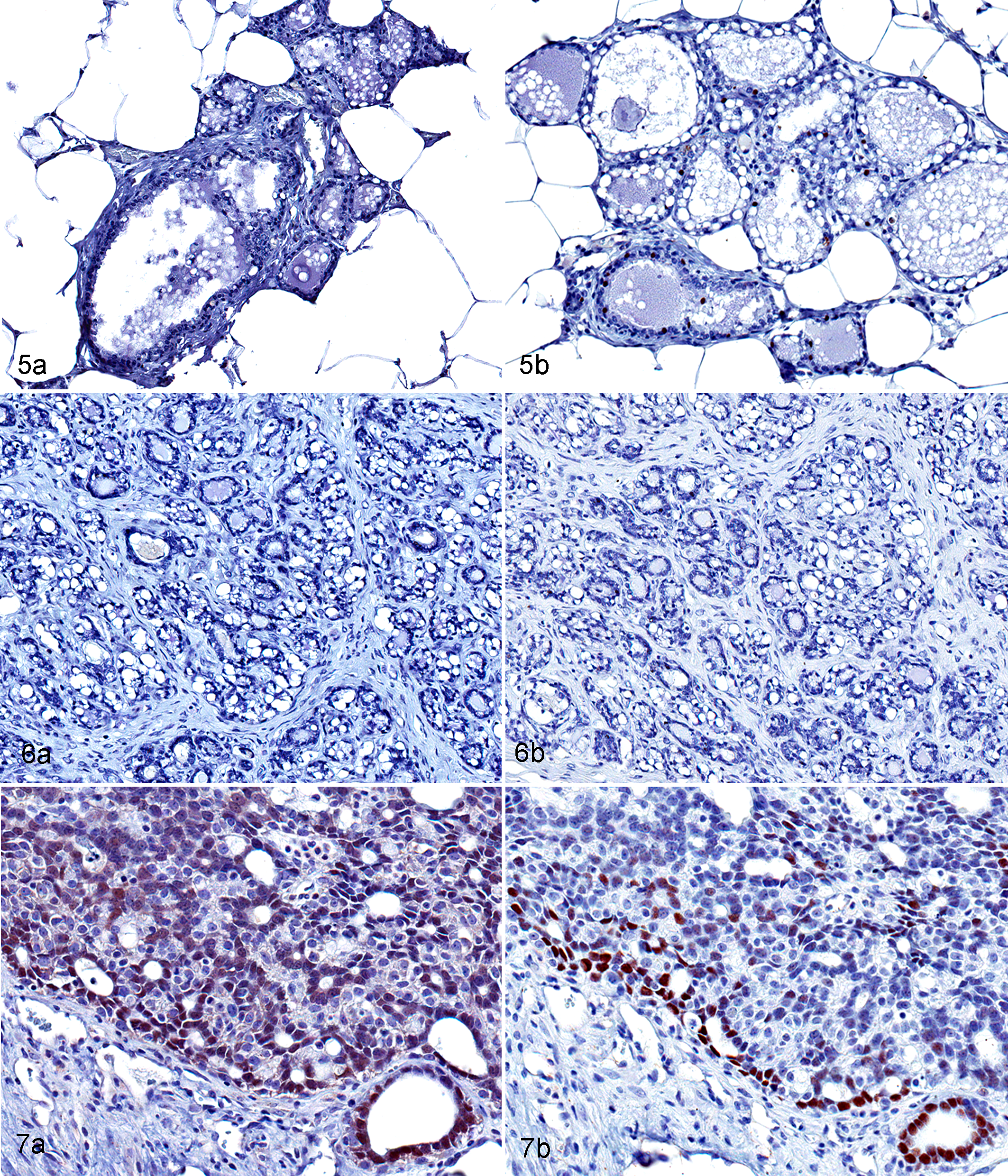
Mammary gland, bromodichloroacetic acid–exposed rat. Immunohistochemistry.
In control and BDCA-exposed animals, myoepithelium of normal mammary gland (Fig. 8) and fibroadenomas (Fig. 9) were positive for SMA. Glandular epithelial cells showed very faint immunoreactivity to SMA. In contrast, the basal epithelial layers in a majority of spontaneous (3 of 5) and BDCA adenocarcinomas (10 of 13) were strongly positive (Fig. 10) for SMA. Normal mammary gland (Fig. 11) and spontaneous fibroadenomas (Fig. 12) were negative for CK14, and 1 of 5 fibroadenomas from BDCA-exposed animals showed focal expression for CK14. In contrast, 5 of 5 spontaneous adenocarcinomas and 11 of 13 adenocarcinomas from BDCA-exposed animals were diffusely positive (Fig. 13). Luminal epithelial cells of normal mammary gland (Fig. 14), all spontaneous and BDCA fibroadenomas (Fig. 15), 1 of 5 spontaneous adenocarcinomas, and 12 of 13 adenocarcinomas from BDCA-exposed animals (Fig. 16) were positive for CK18.
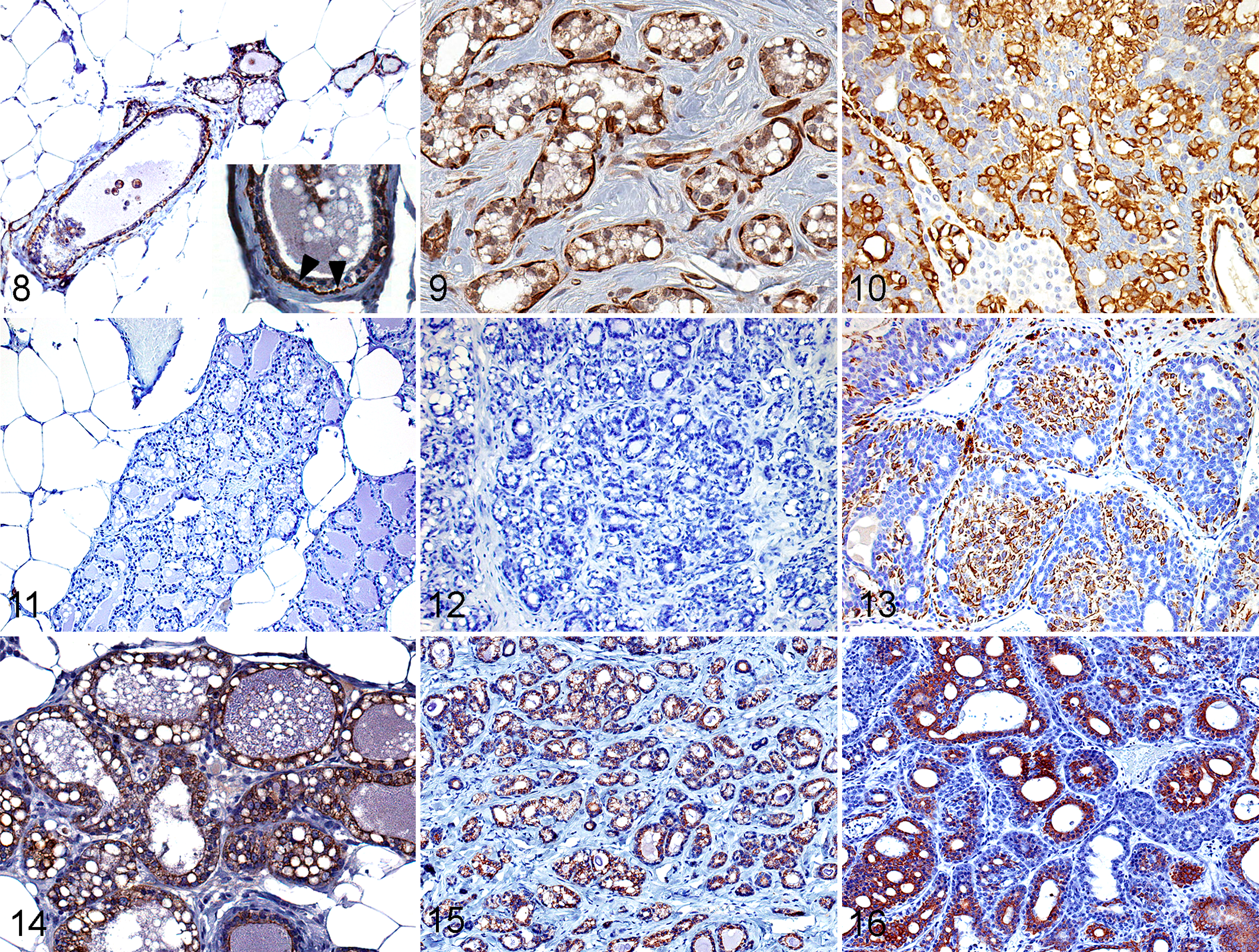
Mammary gland, bromodichloroacetic acid–exposed rat. IHC.
Mammary Adenocarcinomas in F344/Ntac Rats Harbor Low Incidence of Mutations Observed in Human Breast Cancer
There were no mutations observed in Tp53, Pten, or Egfr in any of the spontaneous fibroadenomas evaluated. Interestingly, there were Tp53 mutations in 4 of 5 BDCA-treated fibroadenomas evaluated; however, these were synonymous mutations, in which a single base pair change did not result in a change in the resulting amino acid sequence (data not shown). Of the 5 spontaneous adenocarcinomas evaluated by mutation analysis, 2 harbored Tp53 mutations in exon 5 (Table 3), and of the 8 BDCA-treated adenocarcinomas, 1 mid-dose animal harbored a mutation in exon 8 of Tp53 and also in exon 19 of Egfr, and 2 high-dose animals each had mutations in exon 7 of Pten and exon 20 of Egfr.
Gene Mutations in Mammary Adenocarcinomas From Control and BDCA-Exposed Rats.a
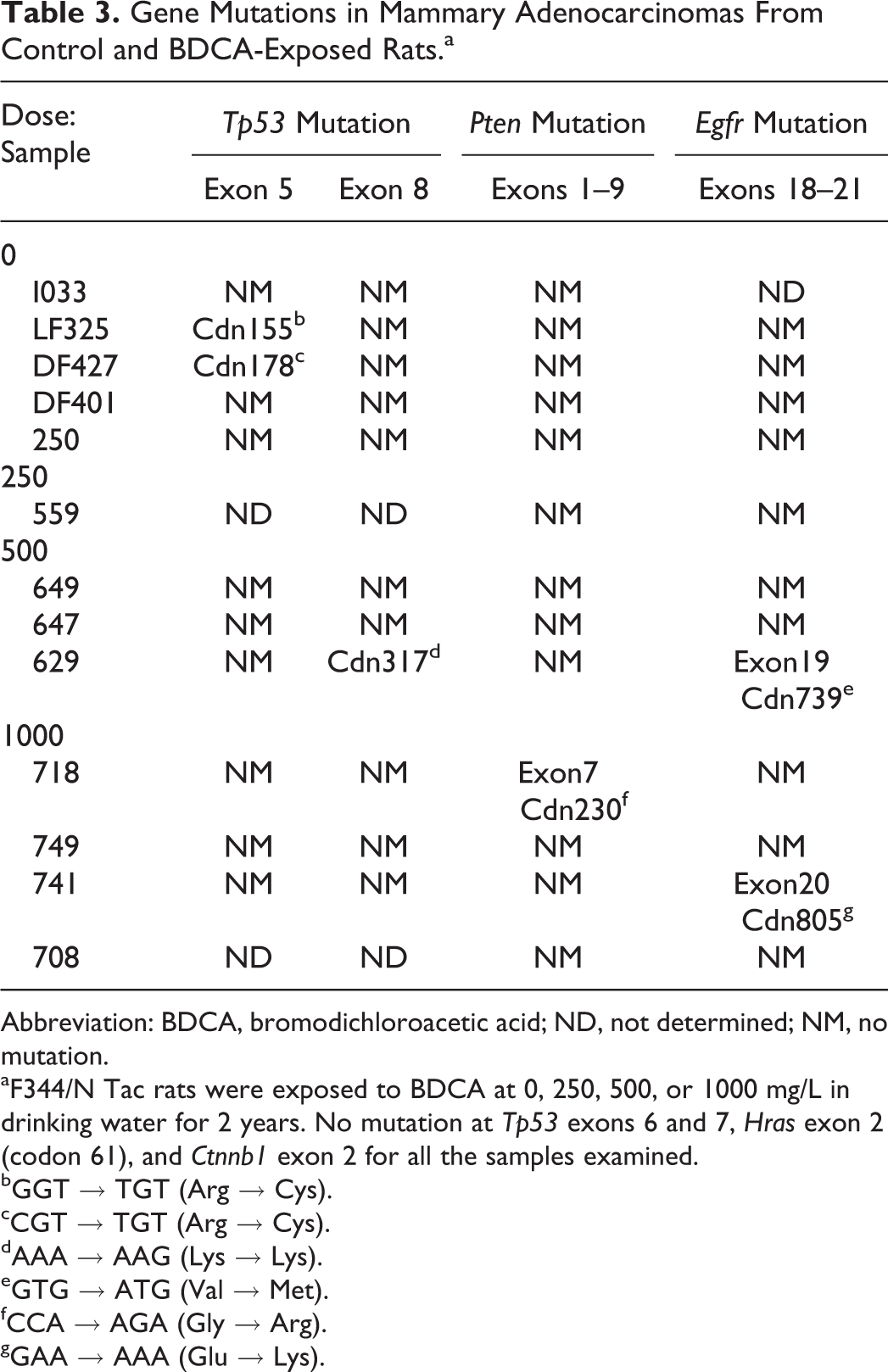
Abbreviation: BDCA, bromodichloroacetic acid; ND, not determined; NM, no mutation.
aF344/N Tac rats were exposed to BDCA at 0, 250, 500, or 1000 mg/L in drinking water for 2 years. No mutation at Tp53 exons 6 and 7, Hras exon 2 (codon 61), and Ctnnb1 exon 2 for all the samples examined.
bGGT → TGT (Arg → Cys).
cCGT → TGT (Arg → Cys).
dAAA → AAG (Lys → Lys).
eGTG → ATG (Val → Met).
fCCA → AGA (Gly → Arg).
gGAA → AAA (Glu → Lys).
Spontaneous and BDCA-Treated Adenocarcinomas Show Gene Expression Alterations Related to Pathways of Tumorigenesis in Human Breast Cancer
When normal mammary gland, spontaneous fibroadenoma, and spontaneous adenocarcinoma were compared, trend analysis reported a statistically significant upward (positive) trend in 37 of 84 genes, and a statistically significant downward (negative) trend in 15 of 84 genes (FDR < 0.05; Table 4). In BDCA-exposed animals, trend analysis reported 49 of 84 genes with an upward trend and 10 of 84 genes with a downward trend, for the same comparisons. Spontaneous and BDCA-treated mammary tumors shared 34 of the upward-trending genes and 9 of the downward-trending genes. Three upward-trending genes on the array were exclusive to spontaneous tumors, while 15 upward-trending genes associated with breast cancer were exclusive to BDCA-treated tumors. Conversely, there were 6 genes exclusively trending downward in spontaneous tumors, while only 1 gene (Brca2) was exclusively trending downward in BDCA-treated tumors. While spontaneous and BDCA-treated tumors shared a majority of genes in terms of directionality of trend, BDCA-treated tumors had proportionally more genes that were exclusively upward trending; conversely, spontaneous tumors had proportionally more genes exclusively trending downward.
Significantly Altereda Genes by Trend Analysis in Fibroadenomas and Adenocarcinomas From Control and BDCA-Exposed Rats.
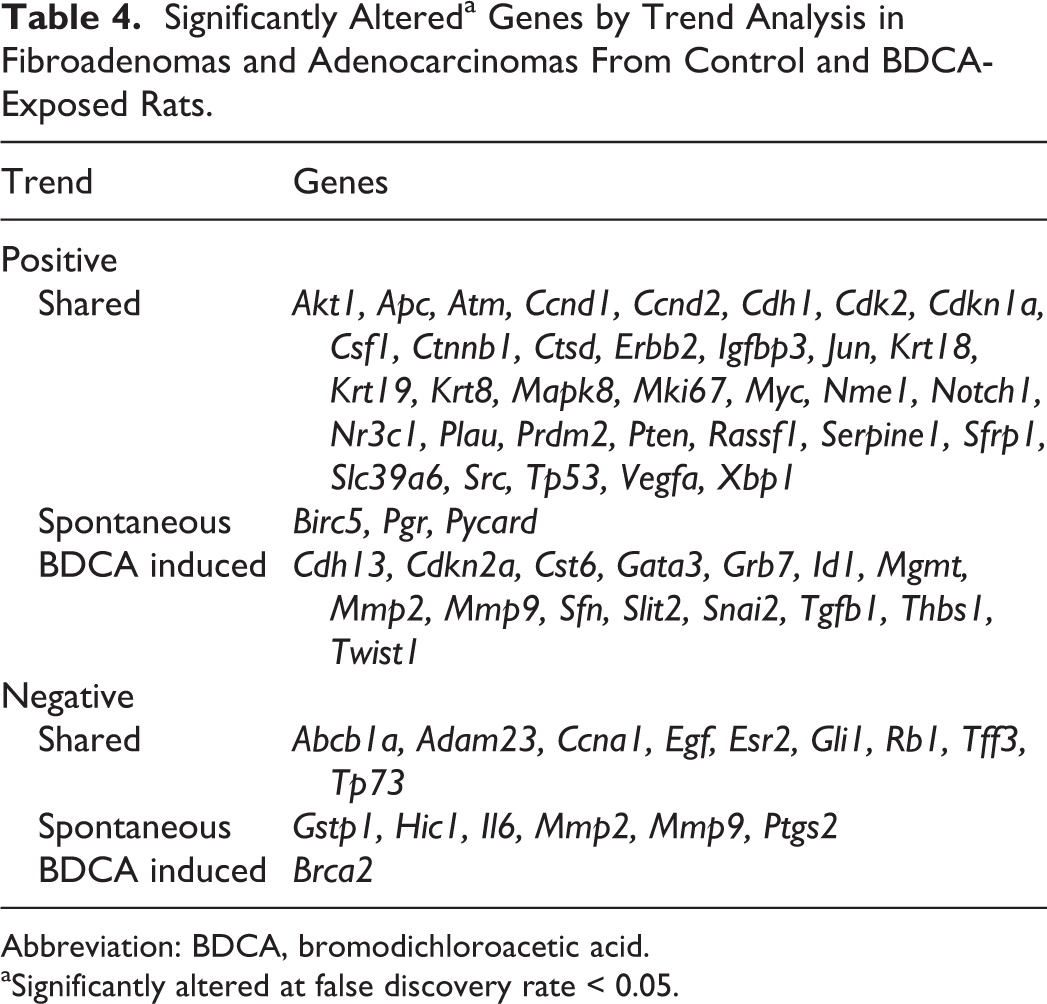
Abbreviation: BDCA, bromodichloroacetic acid.
aSignificantly altered at false discovery rate < 0.05.
Genes that showed a statistically significant upward trend that were shared by both spontaneous and BDCA-treated mammary lesions (Table 4) included those involved in signal transduction, including the PI3k/Akt, Wnt, Notch, and Mapk pathways (Akt, Apc, Ctnnb1, Sfrp1, Notch1, Mapk8); glucocorticoid signaling (Igfbp3, Nme1, Nr3c1); cell cycle (Ccnd1, Ccnd2, Cdk2, Cdkn1a, Ki67, Myc, Pten, Rassf1, Tp53); cell adhesion (Cdh1, Csf1); DNA damage (Atm); matrix remodeling (Ctsd, Plau); epithelial-mesenchymal transition (Src, Notch1); angiogenesis (Vegfa); and luminal tumor classification markers (Krt8, Krt18, Krt19, Erbb2, Slc39a6, Xbp1). Genes that were found to trend upward exclusively in spontaneous tumors included Birc5 (antiapoptosis, basal marker), Pgr (hormone receptor), and Asc (matrix remodeling). Conversely, there were several more genes exclusively trending upward in BDCA-treated tumors, including genes associated with cell cycle (Cdkn2a, Sfn); matrix remodeling (Mmp2, Mmp9, Cst6); adhesion, angiogenesis, and epithelial-mesenchymal transition (Cdh13, Id1, Slit2, Tgfb1, Thbs2, Twist); hedgehog signaling (Snai2); and both luminal (Gata3) and basal (Grb7) tumor markers.
Those genes shared between spontaneous and BDCA-treated tumors with a statistically significant downward trend (Table 4) included xenobiotic transporters (Abcb1a), matrix remodeling (Adam23), cell cycle (Ccna1, Rb1), angiogenesis (Egf), Hedgehog signaling (Gli1), DNA damage (Tp73), and luminal tumor markers (Tff3).
BDCA-Treated Mammary Adenocarcinomas Are Associated With Upregulation of Genes Associated With the Tgfβ Pathway, Tumor Progression, and Invasion
Pairwise comparison of spontaneous and BDCA-treated mammary adenocarcinomas was performed to identify differentially expressed genes relevant for human breast cancer. This analysis showed that of the 84 genes included on the PCR array, 8 genes were significantly (P < .05) upregulated in BDCA-treated mammary adenocarcinomas compared to spontaneous tumors, 5 of which are associated with increased matrix remodeling, tumor progression, invasion, metastasis, and Tgfβ pathway signaling (Mmp2, Mmp9, Id1, Vegfa, Thbs1; Table 5).
Significantly Upregulateda Genes Associated With Tgf-β Signaling, Breast Cancer Growth, Invasion, and Metastasis in BDCA-Treated Mammary Adenocarcinomas.b
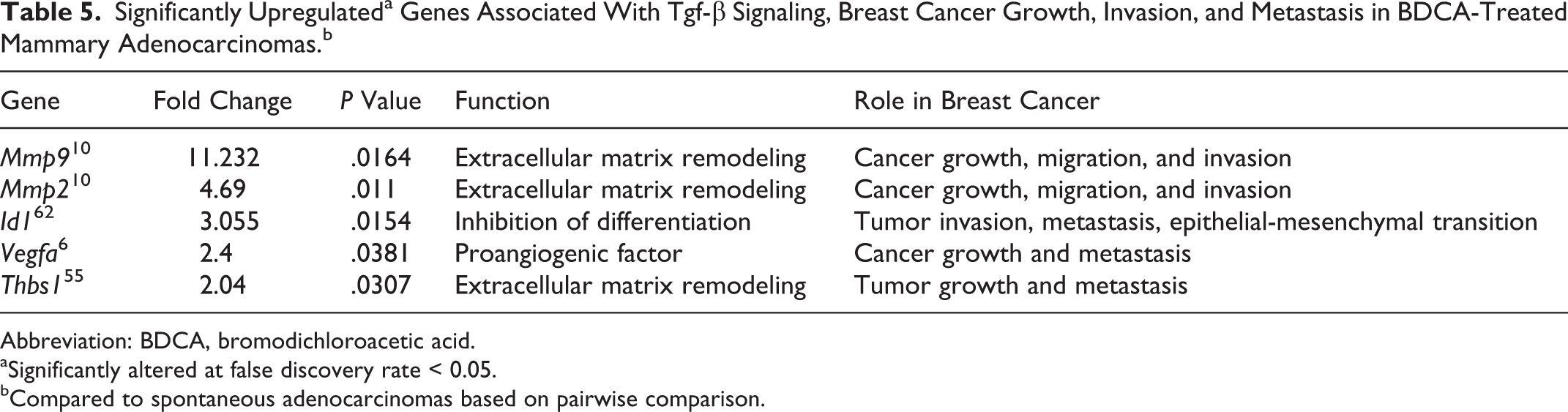
Abbreviation: BDCA, bromodichloroacetic acid.
aSignificantly altered at false discovery rate < 0.05.
bCompared to spontaneous adenocarcinomas based on pairwise comparison.
Discussion
Studies have shown that a majority of human cancer is due environmental factors. 36 Whether environmental contaminants, occupational exposures, or nutritional/lifestyle factors, an overwhelming number of environmental exposures have been linked to an increased risk of mammary cancer in humans and/or rodent models. 24,66 These environmental exposures pose a significant risk to human populations, not only because of their detrimental effects on human health, but also because numerous compounds are unregulated. The current 2-year chronic rat bioassay confirmed the carcinogenic effects of the environmental contaminant BDCA, with increased incidence of mammary hyperplasias, fibroadenomas, and adenocarcinomas. These findings resulted in a conclusion of clear evidence of carcinogenic activity of BDCA in female F344/NTac rats. 50 Additional findings supporting clear evidence of carcinogenicity in F344/NTac rats included increased incidences of malignant mesothelioma and cutaneous epithelial tumors. 50 There are no studies in the current literature which report carcinogenic activity of BDCA; however, a variety of reports describe carcinogenic activity of other haloacetic acids. 12 –14,42 One related haloacetic acid, bromochloroacetic acid, caused increased incidence of fibroadenoma of the mammary gland of rats. 49 No studies report carcinogenic effects of BDCA in humans; however, several have shown a link between bladder, rectal, and brain cancer risk and exposure to water disinfection by-products. 7,41,46 This is the first report to assess the molecular phenotype of BDCA-induced mammary tumors in rats in the context of human breast cancer, and it suggests that BDCA may be an environmental cause of human breast cancer. This is important because BDCA is not currently regulated by the Environmental Protection Agency, 50 and due to potential widespread human exposure, determination of the possible human health risk is critical.
By comparing the gene and protein expression of spontaneous adenocarcinomas from other NTP studies to that of BDCA-treated lesions, we were able to perform a focused analysis to (1) characterize differences in luminal/basal phenotype, (2) examine hormone status, and (3) evaluate differential gene expression, of these lesions in BDCA-exposed F344/NTac rats. Histologically, mammary tumors from control animals and BDCA-exposed animals were morphologically very similar. These tumors had similar rates of metastasis, and there were no clear differences in local invasion or cellular atypia. Immunohistochemical analysis showed that adenocarcinomas from control and BDCA-exposed animals in general possessed the same immunophenotype. Both showed an ERα/PR-positive phenotype and a mixed luminal/basal phenotype. Human breast cancers are generally classified as luminal or basal based on immunohistochemistry or gene expression data, and this classification generally informs on biological behavior and prognosis, with luminal tumors harboring a more favorable prognosis and less aggressive clinical course than basal tumors. 58 However, not all human breast cancers necessarily fall into mutually exclusive groups based on luminal and basal markers; in fact, the classification of breast cancers based on basal (CK5, 14, 17) and luminal (CK8, 18, 19) markers remains controversial. 25 For example, some breast cancers in humans may express both basal and luminal markers, and it has been shown that even in the normal breast, basal markers may be expressed in some of the duct or acinar luminal epithelial cells, in addition to basal epithelial or myoepithelial cells. 20,21 In a study examining basal and luminal cytokeratin expression in 1,944 human breast carcinomas, 27.4% of tumors expressed both luminal and basal cytokeratins, 1 making the determination of biological behavior difficult based on the relative expression of these markers.
Mutation analysis of spontaneous and BDCA-treated mammary tumors illustrated possible molecular alterations associated with phenotype and exposure. For example, untreated fibroadenomas did not harbor mutations in any of the genes analyzed, whereas a majority of BDCA-treated fibroadenomas harbored synonymous Tp53 mutations. Only treatment-related tumors exhibited Pten mutations (1 fibroadenoma and 1 adenocarcinoma) and Egfr mutations (1 fibroadenoma and 2 adenocarcinomas), whereas spontaneous tumors did not harbor mutations in these genes. The mutation spectra observed in treated fibroadenomas and adenocarcinomas and the increased variety of mutations in BDCA-treated adenocarcinomas may suggest increased DNA insult and damage leading to progression of tumorigenesis or an increased incidence of neoplastic transformation in the mammary gland overall. However, a conclusion based on a small incidence of mutations in experimental groups of relatively small size such as this would be difficult. Ideally, a larger sample number should be evaluated for additional mutations in these 3 genes, as well as a broader number of genes to gain a better understanding of how these alterations relate to the increased incidence of tumors in this study.
Gene expression analysis showed that both spontaneous and BDCA-treated tumors expressed mostly luminal markers (CK8, 18, 19, Gata3, Slc39a6, Xbp1) but also Notch1, which is upregulated in basal-type breast cancer and regulates a basal phenotype. 23 The relevance of the dual expression of basal and luminal markers observed in mammary adenocarcinomas in BDCA-exposed F344/N rats is uncertain. It may point to a more aggressive phenotype, but it has also been shown in human breast cancer that more than half of ERα+/PR+ breast cancers contain a small population of neoplastic basal cells negative for hormone receptors, which can expand on the basis of hormone therapy and Notch signaling, 23 as well as account for a mixed luminal/basal phenotype. Alternatively, given that these tumors arise late in life and are not widely invasive or metastatic, their biological behavior may reflect a less aggressive phenotype unrelated to an immunophenotype as seen in humans.
BDCA-treated and spontaneous mammary tumors shared a number of molecular alterations in gene expression related to cancer progression and growth. A number of genes that play a role in cell proliferation, growth, and survival (Ccnd1, Ccnd2, Cdk2, Erbb2, Jun, Mki67, Myc, Akt1, Ctnnb1, Vegfa, Slc39a6) showed a positive increasing trend from normal mammary gland to fibroadenoma to adenocarcinoma in both groups (spontaneous and BDCA-treated), suggesting that these changes are common to the process of mammary carcinogenesis in rats and humans. However, importantly, there was a large proportion of genes related to cell cycle progression, matrix remodeling, cellular adhesion, and angiogenesis that were trending upward exclusively in BDCA-treated lesions. Specifically, additional cell cycle mediators (Cdkn2a), matrix remodeling enzymes (Mmp2, Mmp9), and genes associated with cell adhesion, angiogenesis, invasion, and epithelial-mesenchymal transition (Id1, Tgfb1, Thbs2, Twist)—important features of more aggressive human breast cancer—were overrepresented compared to spontaneous lesions. In addition, BDCA-treated tumors showed a decreasing trend in the expression of Brca2, an important DNA repair gene that, when mutated in humans, confers a 45% to 85% increased lifetime risk for breast cancer. 45 Trend analysis comparing patterns of gene expression between spontaneous and BDCA-treated neoplastic mammary lesions suggests that BDCA-treated lesions represent a more aggressive phenotype, including increased cell cycle signaling, matrix remodeling, invasion, and epithelial-to-mesenchymal transition.
Last, results of a pairwise comparison between spontaneous and BDCA-treated adenocarcinomas identified a 5-gene signature of particular relevance to more aggressive breast cancer (Id1, Vegfa, Mmp2, Mmp9, Thbs1), which was significantly upregulated in BDCA-treated compared to spontaneous adenocarcinomas. Of particular interest is the fact that these genes are regulated in part by Tgfβ1, which showed a significantly increasing trend in BDCA-treated mammary carcinomas. TGFβ has been shown to act as a tumor suppressor in early stages of breast cancer development but stimulate invasion and metastasis in later stages. 2,61 It is commonly overexpressed in human breast cancer; it is associated with angiogenesis, epithelial-mesenchymal transition, tumor cell invasion, proliferation, and antiapoptosis 45 ; and it correlates with cancer progression and a poor prognosis. 28 There is a paucity of data in the literature reporting involvement of this pathway in the development of spontaneous mammary tumors in the F344/NTac rat; however, some studies have shown that stimulation of rat mammary adenocarcinoma cells by Tgfβ in experimental syngeneic models enhanced their tumorigenicity 33 and metastatic behavior. 63 While it is possible that BDCA is acting on preexisting molecular pathways to promote a background spontaneous mammary tumor rate in the F344/NTac rat, these data suggest that BDCA could alter function of the Tgfβ pathway separately from background alterations, through chemical-specific mechanisms.
TGFβ regulates the expression of MMP2 and MMP9, 65 matrix metalloproteinases that function to degrade extracellular matrix, including type IV collagen, 60 the main constituent of the basal lamina. Conversely, MMP2 and MMP9 also cleave adhesion molecules and activate growth factors, including TGFβ. 18 In line with this, MMP2 and MMP9 have been shown to be associated with breast cancer cell growth, migration, and invasion. 10 Inhibitor of differentiation 1 (Id1)—a member of the helix-loop-helix family of genes found to be significantly upregulated in BDCA-treated adenocarcinomas—is also regulated by Tgfβ. Overexpression of ID1 in human breast cancer is associated with a more invasive phenotype, as well as epithelial-mesenchymal transition, 62 a phenomenon associated with the acquisition of a more invasive and metastatic phenotype. While there are conflicting reports on the regulation of ID1 by TGFβ in various epithelial and fibroblast cell lines, 8,29 ID1 has been shown to be strongly, albeit transiently, upregulated by TGFβ1 in mammary epithelial cells. 35 Although the upregulation of Id1 in relationship to Tgfβ signaling in these tumors is uncertain, it may provide additional evidence of a Tgfβ-mediated mechanism in BDCA-treated adenocarcinomas. Thbs1 is overexpressed in many different types of human cancer and in some forms of breast cancer (lobular carcinoma). 55 The protein product of this gene is secreted in response to TGFβ stimulation and plays a role in activation of a variety of proteases as well as latent TGFβ. Members of the vascular endothelial growth factor (Vegf) family play a central role in the regulation of angiogenesis, which plays an important role in the process of neoplastic growth. Angiogenesis is also regulated by a number of secreted growth factors, including members of the Tgfβ family. 59 In fact, the induction of Vegf during tumor angiogenesis is mediated in part through Tgfβ stimulation. 6
While adenocarcinomas in BDCA-exposed animals did not differ substantially in terms of morphologic features correlating with increased invasion and there were no differences in rates of metastasis, changes in gene expression suggest that adenocarcinomas from BDCA-exposed animals are molecularly different from spontaneous tumors. The overrepresentation of Tgfβ mediators in adenocarcinomas from BDCA-exposed animals may suggest a correlation between activation of this pathway and the increased incidence of mammary tumors observed in BDCA-exposed animals. This pathway does not appear to be an important pathway in spontaneous mammary adenocarcinoma development in this strain; therefore, these data show that, in part, the process of mammary tumorigenesis in BDCA-exposed F344/NTac rats involves Tgfβ-dependent mechanisms, which are relevant for human breast cancer. Since this pathway plays such an important role in many human breast cancers, its involvement in mammary carcinogenesis in BDCA-exposed F344/NTac rats supports the hypothesis that BDCA exposure in humans may pose a significant health risk. The findings of this study are particularly important in terms of hazard identification for the general public, since there is not only the increased tumor incidence in rats exposed to this chemical, which has a wide human exposure, but also the presence of molecular alterations that are very relevant for development of human cancer. These data show that chronic exposure to this chemical should be considered a potential environmental human health hazard; the molecular findings strengthen the results of the carcinogenicity bioassay, help further define this hazard, and provide additional data for the regulatory community to consider. Additional studies on molecular mechanisms of carcinogenesis in animals exposed to BDCA and other water disinfection by-products are warranted on the basis of our identification of molecular alterations relevant for human cancer.
Footnotes
Acknowledgements
We thank the Cellular and Molecular Pathology Branch Necropsy Core Laboratory personnel; Cellular and Molecular Pathology Branch Histology, Immunohistochemistry, and Special Techniques Core Laboratories; and the National Institute of Environmental Health Sciences DNA Sequencing Core for their excellent technical expertise.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Division of the National Toxicology Program, National Institutes of Environmental Health Sciences, National Institutes of Health.