
Abstract
The aim of this work was to study the interstitial aggregates of immune cells observed in pulmonary parenchyma of calves preinfected with bovine viral diarrhea virus and challenged later with bovine herpesvirus 1. In addition, the intent of this research was to clarify the role of bovine viral diarrhea virus in local cell-mediated immunity and potentially in predisposing animals to bovine respiratory disease complex. Twelve Friesian calves, aged 8 to 9 months, were inoculated with noncytopathic bovine viral diarrhea virus genotype 1. Ten were subsequently challenged with bovine herpesvirus 1 and euthanized at 1, 2, 4, 7, or 14 days postinoculation. The other 2 calves were euthanized prior to the second inoculation. Another cohort of 10 calves was inoculated only with bovine herpesvirus 1 and then were euthanized at the same time points. Two calves were not inoculated with any agent and were used as negative controls. Pulmonary lesions were evaluated in all animals, while quantitative and biosynthetic changes in immune cells were concurrently examined immunohistochemically to compare coinfected calves and calves challenged only with bovine herpesvirus 1. Calves preinfected with bovine viral diarrhea virus demonstrated moderate respiratory clinical signs and histopathologic evidence of interstitial pneumonia with aggregates of mononuclear cells, which predominated at 4 days postinoculation. Furthermore, this group of animals was noted to have a suppression of interleukin-10 and associated alterations in the Th1-driven cytokine response in the lungs, as well as inhibition of the response of CD8+ and CD4+ T lymphocytes against bovine herpesvirus 1. These findings suggest that bovine viral diarrhea virus preinfection could affect the regulation of the immune response as modulated by regulatory T cells, as well as impair local cell-mediated immunity to secondary respiratory pathogens.
Keywords
Bovine viral diarrhea virus (BVDV) is an important pestiviral pathogen of cattle, causing considerable economic losses for the livestock industry. 25 The biology of BVDV is highly complex, depending on a variety of factors such as the viral genotype and biotype causing the infection, the immune status and age of the affected animals, and the gestational stage of cows in case of infection during pregnancy. 7,29,33,44 This leads to a broad spectrum of clinical manifestations and lesions, which are classified according to the type of BVDV infection—namely, acute infection, persistent infection, and mucosal disease. During acute infections, the pestivirus can induce severe reproductive, gastrointestinal, and respiratory disease in cattle. 21,38,49
The contribution of BVDV to the epidemiology of bovine pneumonias is a longstanding source of controversy. The virus has been isolated from the lungs in several studies of acute infection 1,9,17,20 as well as in persistently infected calves. 15,18,19,31,52 Likewise, certain publications have reported an increased susceptibility to bacterial pneumonia in BVDV infections, 10,23 as well as an exacerbation of the pathologic effects produced by other viruses in experimental coinfections. 8,14,28,40 The pulmonary lesions described in association with BVDV infection are typically mild, consisting of different degrees of bronchointerstitial pneumonia or bronchopneumonia with peribronchiolar, perivascular, or interstitial aggregates of lymphocytes and macrophages. 3,15,27 However, other studies have found no lung lesions in calves infected with BVDV or have observed pulmonary pathology that was attributed uniquely to the effects of the secondary infectious agent, 31,42,53,55 questioning the importance of BVDV as a cause of primary lung disease.
Recently, we carried out a study aimed at consistently reproducing the clinical signs and lesions associated with bovine respiratory disease complex (BRDC). The study consisted of a primary BVDV respiratory infection followed by a challenge with bovine herpesvirus 1 (BHV-1). This in vivo model has been extensively used to examine the specific mechanisms by which a primary BVDV infection favors the dissemination of BHV-1. In addition, the model has permitted the observation of an impairment of the systemic immune response based on a depletion of CD8+ T cells, alteration of their cytotoxic action, a delay in interferon γ (IFNγ) production, and low levels of interleukin (IL)–12. 35,48 Furthermore, calves preinfected with BVDV displayed significant tumor necrosis factor α (TNFα) secretion in serum and a reduced production of IL-10 following BHV-1 infection, leading to an exacerbation of the inflammatory response and to the development of intense clinical signs and inflammatory lesions. 48 These lesions have also been observed in the lung of BVDV-infected calves challenged with BHV-1, whose pulmonary parenchyma was affected by an interstitial pneumonia consisting of aggregates of mononuclear cells. 46
A variety of mechanisms have been proposed to elucidate the possible alterations in host responses that predispose to a bacterial pneumonia or favor viral synergy. Certain studies suggest that damage to airway epithelial cells, compromised local cell-mediated immunity, and alterations in the antimicrobial activity of pulmonary macrophages (MΦs) and neutrophils play important roles. 22,24,39,41,49 However, experimental studies of these immunomodulatory processes in the bovine lung have not been performed to date.
To our knowledge, this is the first report that aims to evaluate and characterize the interstitial aggregates of immune cells in the pulmonary parenchyma of calves preinfected with BVDV and subsequently challenged with BHV-1, as well as to clarify the immunomodulatory role of BVDV on these cells in the local cell-mediated immunity and its possible alteration induced by BVDV in favoring the appearance of BRDC.
Materials and Methods
Animals and Experimental Design
The experimental design and collection of samples applied to this experimental study have been described by Risalde et al.
48
Briefly, 30 Frisian calves (8–9 months old), BVDV and BHV-1 antigen and antibody (Ab) free, were obtained from a herd free of tuberculosis, brucellosis, and bovine leukemia virus. Animals were housed in the Animal Experimental Centre of Cordoba University (Spain) and separated in 2 groups based on the inoculation protocol: The BVDV/BHV-1 group (14 calves) was inoculated intranasally with 10 ml of a suspension of noncytopathic BVDV-1 strain 7443 with a concentration of 105 tissue culture infective dose 50% (TCID50)/ml. Twelve days later, when the calves did not demonstrate any clinical signs or viremia, they were challenged with 2 ml of BHV-1.1 strain Iowa containing 107 TCID50/ml. Two of these calves were not inoculated with BHV-1.1 and were used as BVDV infection controls. The BHV-1 group (12 calves) was inoculated only with the BHV-1.1 strain. As a negative control for this group, 2 uninfected (UI) calves received an innoculum of 2 ml of tissue culture fluid free of viruses.
The time point of BHV-1.1 inoculation was defined as 0 days postinoculation (dpi). After the viral challenge, clinical examination was performed daily until euthanasia. At staged time points and in groups of 2 animals, the infected calves were sedated with xylazine hydrochloride (Rompun 2%; Bayer Healthcare, Kiel, Germany) and euthanized via intravenous injection with thiopental sodium (Thiovet; Vet Ltd, Leyland, Lancashire, England) at 1, 2, 4, 7, and 14 dpi. The 2 BVDV-infected but BHV-1–negative control calves were similarly sedated and euthanized the same day in which the other animals of their group were inoculated with BHV-1. On the other hand, the 2 uninfected animals that comprised the negative control group were euthanized at the end of the study. The experimental procedure was carried out in accordance with the Code of Practice for Housing and Care of Animals Used in Scientific Procedures, approved by the European Economic Community in 1986 (86/609/EEC amended by the directive 2003/65/EC).
Pulmonary Histopathology
Samples from the lung (cranial and caudal lobes of the lung) were collected during necropsies and fixed in 10% buffered formalin, Bouin’s solution, and zinc salts fixatives. These samples were routinely processed, embedded in paraffin wax, sectioned (3 μm), and stained with hematoxylin and eosin (HE) for structural studies. The types of histopathologic lesions were graded as absent or 0% of pulmonary parenchyma affected (–), mild or 10% to 30% affected (+), moderate or 30% to 70% affected (++), and severe or 70% to 100% affected (+++). For graphical representation, these lesions were scored from absent to severe (0–3).
Processing of Lung Tissue Samples for Immunohistochemistry (IHC)
Fixed lung samples collected at the different euthanasia time points were embedded in paraffin wax and processed for immunohistochemistry (IHC) using the avidin-biotin-peroxidase complex (ABC) method. Tissue sections (3 μm) were dewaxed and rehydrated. Endogenous peroxidase activity was exhausted by incubation with 0.3% hydrogen peroxide in methanol for 30 minutes at room temperature. The samples were subjected to different pretreatments for antigen retrieval depending on the Abs used (Table 1). Subsequently, sections were rinsed 3 times in phosphate-buffered saline (PBS; pH 7.2) for 10 minutes, covered with 1% normal horse serum (Pierce-Endogen, Woburn, MA) in 0.05 M Tris buffered saline (TBS; pH 7.6) for 30 minutes at room temperature, and incubated with the primary mouse Abs at 4°C overnight. For the rat and rabbit primary Abs, 1% normal rabbit serum or 20% normal goat serum, respectively, was used to replace normal horse serum. After primary incubation, slides were washed in PBS (3 times for 5 minutes each) and then incubated with the secondary Abs for 30 minutes at room temperature. Biotinylated horse anti–mouse IgG secondary Ab (Pierce-Endogen) diluted 1:200 in Tris buffer containing normal horse serum 1% was used for the primary monoclonal Abs (mAbs), and biotinylated goat anti–rabbit IgG secondary Ab (Vector Laboratories, Burlingame, CA) diluted 1:200 in TBS containing normal goat or rabbit serum 1.5% was used for the primary polyclonal Abs (pAbs). After 3 further 5-minute washes in PBS, samples were incubated with the ABC complex (Vectastain ABC Elite Kit; Vector Laboratories) for 1 hour at room temperature. All tissue sections were rinsed in TBS and incubated with the chromogen solution (NovaRED Substrate Kit; Vector Laboratories). Finally, slides were counterstained with Harris’s hematoxylin.
Details of the Immunohistochemical Primary Antibodies.
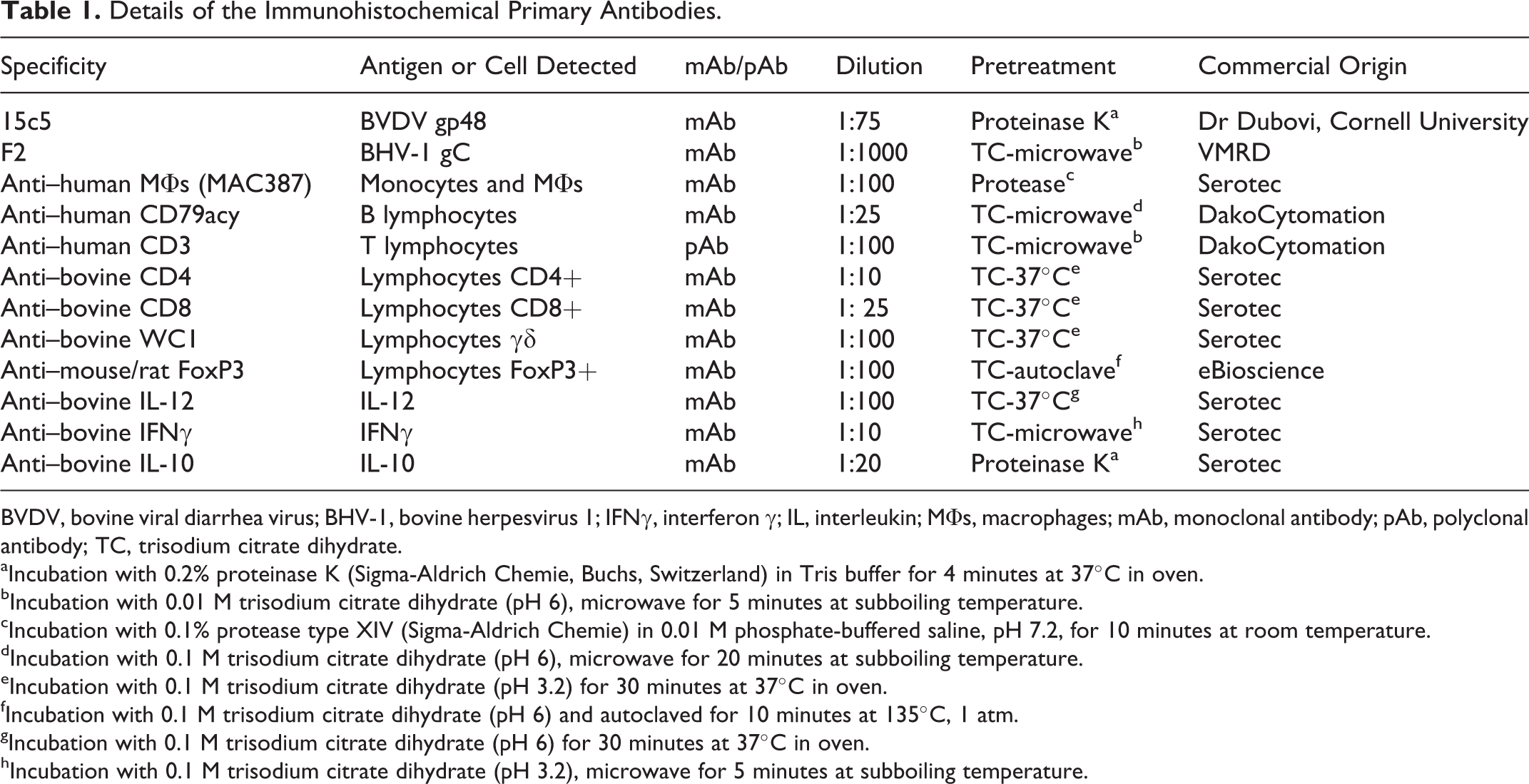
BVDV, bovine viral diarrhea virus; BHV-1, bovine herpesvirus 1; IFNγ, interferon γ; IL, interleukin; MΦs, macrophages; mAb, monoclonal antibody; pAb, polyclonal antibody; TC, trisodium citrate dihydrate.
aIncubation with 0.2% proteinase K (Sigma-Aldrich Chemie, Buchs, Switzerland) in Tris buffer for 4 minutes at 37°C in oven.
bIncubation with 0.01 M trisodium citrate dihydrate (pH 6), microwave for 5 minutes at subboiling temperature.
cIncubation with 0.1% protease type XIV (Sigma-Aldrich Chemie) in 0.01 M phosphate-buffered saline, pH 7.2, for 10 minutes at room temperature.
dIncubation with 0.1 M trisodium citrate dihydrate (pH 6), microwave for 20 minutes at subboiling temperature.
eIncubation with 0.1 M trisodium citrate dihydrate (pH 3.2) for 30 minutes at 37°C in oven.
fIncubation with 0.1 M trisodium citrate dihydrate (pH 6) and autoclaved for 10 minutes at 135°C, 1 atm.
gIncubation with 0.1 M trisodium citrate dihydrate (pH 6) for 30 minutes at 37°C in oven.
hIncubation with 0.1 M trisodium citrate dihydrate (pH 3.2), microwave for 5 minutes at subboiling temperature.
Details of the primary mAbs and pAbs are summarized in Table 1. For the negative controls, specific primary Abs were replaced by rabbit or mouse nonimmune sera (DakoCytomation, Glostrup, Denmark). For gp48 detection by IHC, positive control tissues were from calves persistently infected with BVDV (kindly provided by Dr Dubovi, Cornell University, NY), and for gC detection, positive control samples were from several aborted fetuses positive to BHV-1.1 (courtesy of Moredum Research Institute, Scotland, UK), while negative control tissues were from specific pathogen-free calves not exposed to these viruses.
Cell Counting
To evaluate the number of immunolabeled cells and to compare the results obtained using different Abs, 2 paraffin wax blocks from the lung of each animal were selected. On tissue sections from these blocks, cell counts were carried out by 2 experienced observers in 50 fields of 0.2 mm2 chosen randomly and were blinded to which group was being analyzed. The results were given as the number of positive cells per 0.2 mm2. Cellular identification was based on morphologic features, location, and size of the cells.
Results
Pulmonary Pathology
The morphologic changes observed in the respiratory tract were previously described in Risalde et al. 46,47 Briefly, no remarkable pulmonary lesions were observed in negative control calves. However, in the BVDV/BHV-1 group, significant histologic changes were observed by 1 dpi. The most severe changes were observed in the BVDV/BHV-1 group between 2 and 7 dpi, with a peak at 4 dpi occurring in both dual- and single-infection groups. Furthermore, both groups demonstrated a decrease in lesions by 14 dpi (Fig. 1). The specific histopathologic findings consisted of both inoculated groups being affected by an interstitial pneumonia with alveolar septal thickening produced by interstitial aggregates of mononuclear cells. In addition, fibrin microthrombi and clusters of platelets in some pulmonary venules were observed. These lesions were noted earlier and with greater severity in coinfected calves, most significantly at 4 dpi, and were associated with occasional alveolar edema and hemorrhage (Figs. 1 –3).
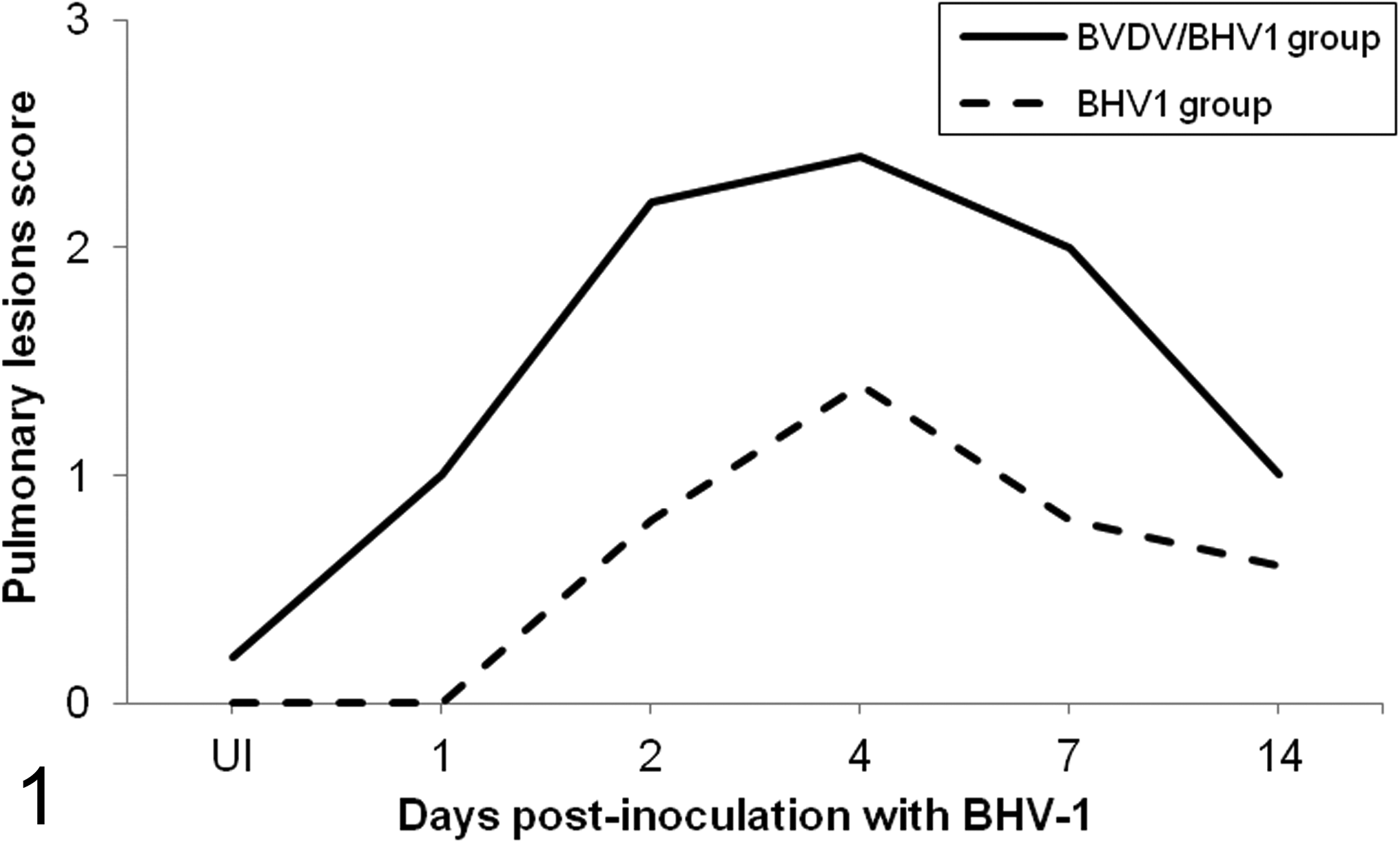
Mean (n = 2 per time point) of the histopathologic lesions scoring in the lung of calves inoculated with herpesvirus 1 subtype 1 (BHV-1.1) vs calves inoculated with bovine viral diarrhea virus (BVDV) and BHV-1.1 at 1, 2, 4, 7, and 14 days postinoculation BHV-1.1 (UI, BHV-1.1 uninfected: negative controls for the BHV-1 group and BVDV infection controls for the BVDV/BHV-1 group). These histopathologic lesions were graded as absent or 0% of pulmonary parenchyma affected (– or 0), mild or 10% to 30% affected (+ or 1), moderate or 30% to 70% affected (++ or 2), or severe or 70% to 100% affected (+++ or 3).
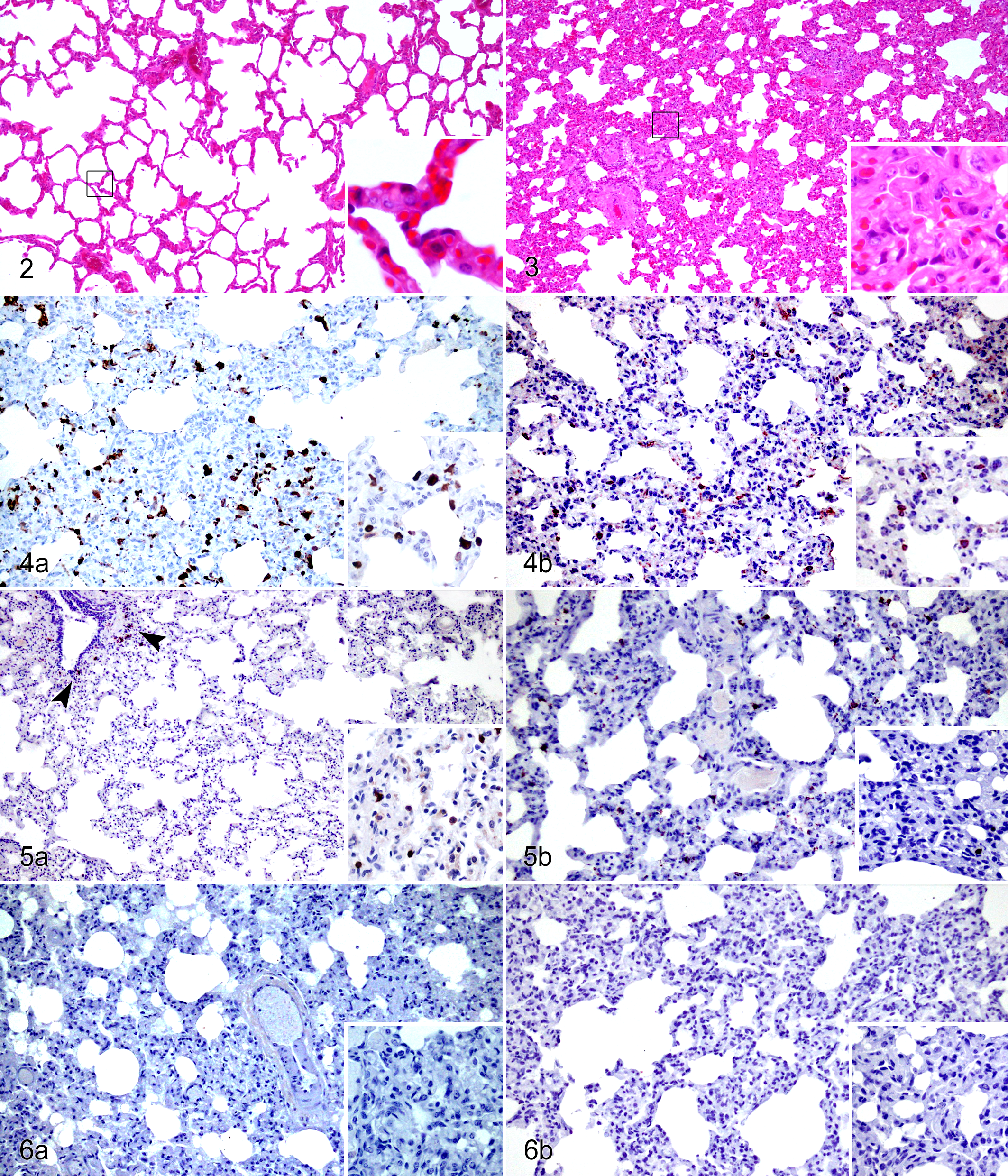
Bovine herpesvirus 1 subtype 1 (BHV-1.1) infection, lung, calf. Minimal alveolar septal thickening was observed at 7 days postinoculation (dpi) (inset). Hematoxylin and eosin (HE).
Mean number (n = 2 per time point) of interstitial macrophages (IMΦs) positive for MAC387, interstitial CD3+ T lymphocytes, and CD79αcy+ B lymphocytes immunolabeled in the lungs of calves coinfected experimentally with bovine viral diarrhea virus (BVDV) and herpesvirus 1 subtype 1 (BHV-1.1) compared with calves inoculated only with BHV-1.1. (UI, BHV-1.1 uninfected: negative controls for the BHV-1 group and BVDV infection controls for the BVDV/BHV-1 group).
In the immunohistochemical examination, mononuclear aggregates present in the pulmonary parenchyma of both animal groups were composed of interstitial macrophages (IMΦs) and T lymphocytes (CD3+) (Fig. 4a,b), which were mostly CD4+, CD8+, and γδ. In addition, a few interspersed CD79αcy+ B cells were also observed. The immunohistochemical techniques confirmed the presence of secretory activity in the pulmonary immune cells, which were immunolabeled against several types of cytokines and presented a different response between both inoculated groups (Fig. 4–6).
Quantitative and Biosynthetic Changes in Pulmonary Immune Cells
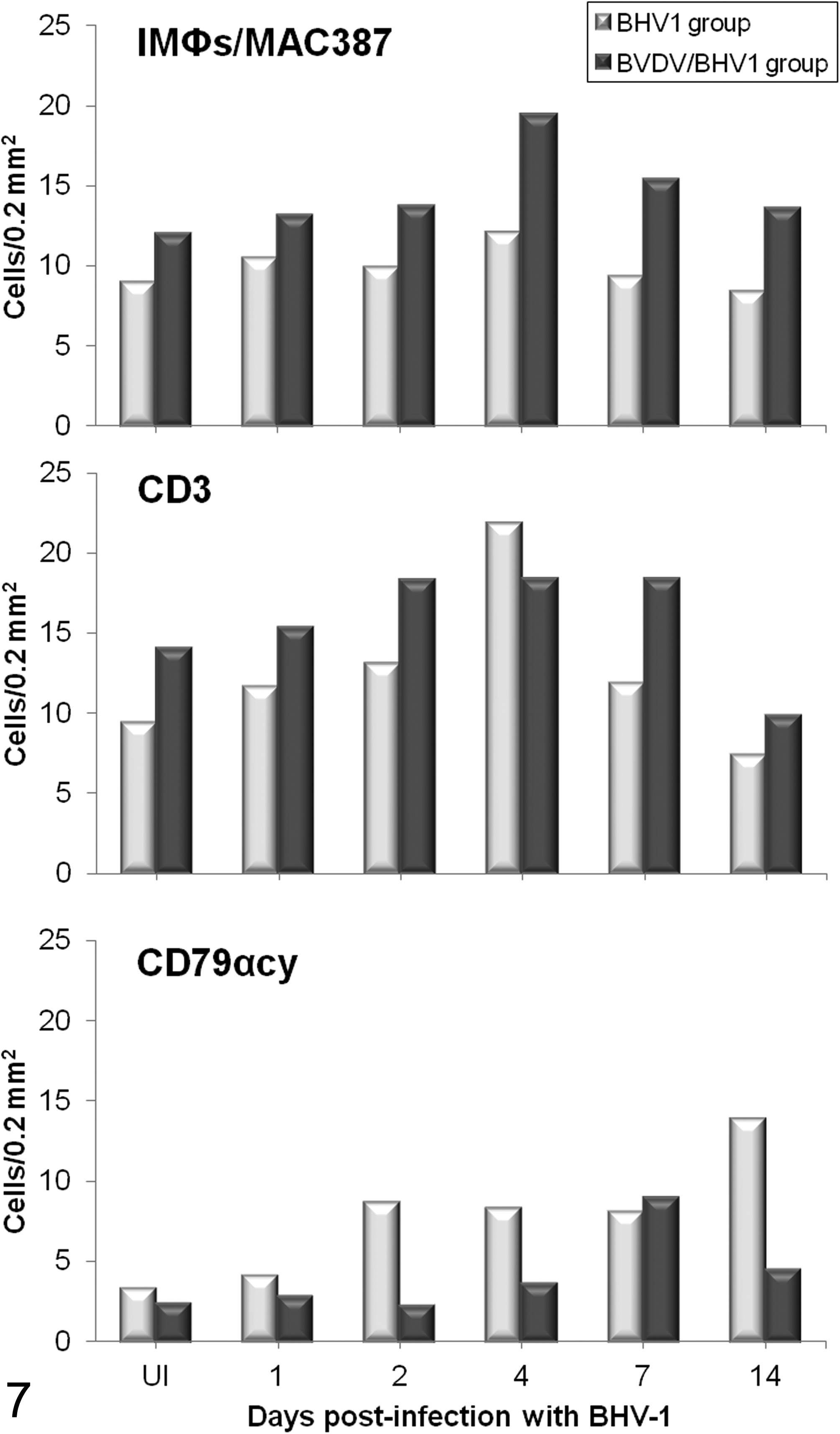
Immunolabeling of IMΦs displayed similar kinetics in single and dual infections, with an observed increase in their number at 4 dpi. However, both inoculated groups presented differences in the magnitude of their responses, demonstrated by the BVDV/BHV-1 group having a higher number of these cells at the 4 dpi peak (Fig. 7).
Interstitial CD3+ T lymphocytes showed similar behavior to BHV-1.1 infection in both inoculated groups, observed as an increase in the number of that particular cell type between 2 and 7 dpi. Nevertheless, the magnitude of this response was higher in the BVDV/BHV-1 group, except for a peak at 4 dpi in the calves infected only with BHV-1 (Fig. 7).
In both inoculated groups, the number of B lymphocytes increased after BHV-1.1 inoculation. This response was earlier and more intense in the BHV-1 group, showing a peak at 14 dpi with values 4 times above the baseline level (from approximately 3.3–13.85 cells/0.2 mm2) (Fig. 7).
Immunolabeling of T-lymphocyte subpopulations revealed that CD4+, CD8+, and γδ+ lymphocytes were increased in number in BVDV-preinfected calves with respect to healthy calves. Following inoculation with BHV-1.1, these cell populations demonstrated a progressive decrease from 4 dpi in coinfected calves, decreasing 2-fold below their baseline level by 14 dpi. In the BHV-1 group, γδ+ lymphocytes showed similar responses to the coinfected group, while a small increase in CD8+ and CD4+ T lymphocytes was observed at 2 and 4 dpi, respectively (Fig. 8). On the other hand, calves infected with BVDV were observed to have a decreased number of FoxP3 regulatory T lymphocytes in comparison with the healthy animals prior to BHV-1 inoculation and remained so until 2 dpi. Subsequent to 2 dpi, the BHV-1 group was observed to have a progressive decrease in FoxP3+ cells, which reached values as low as those observed in the BVDV/BHV-1 group by the end of the experiment (Fig. 8).
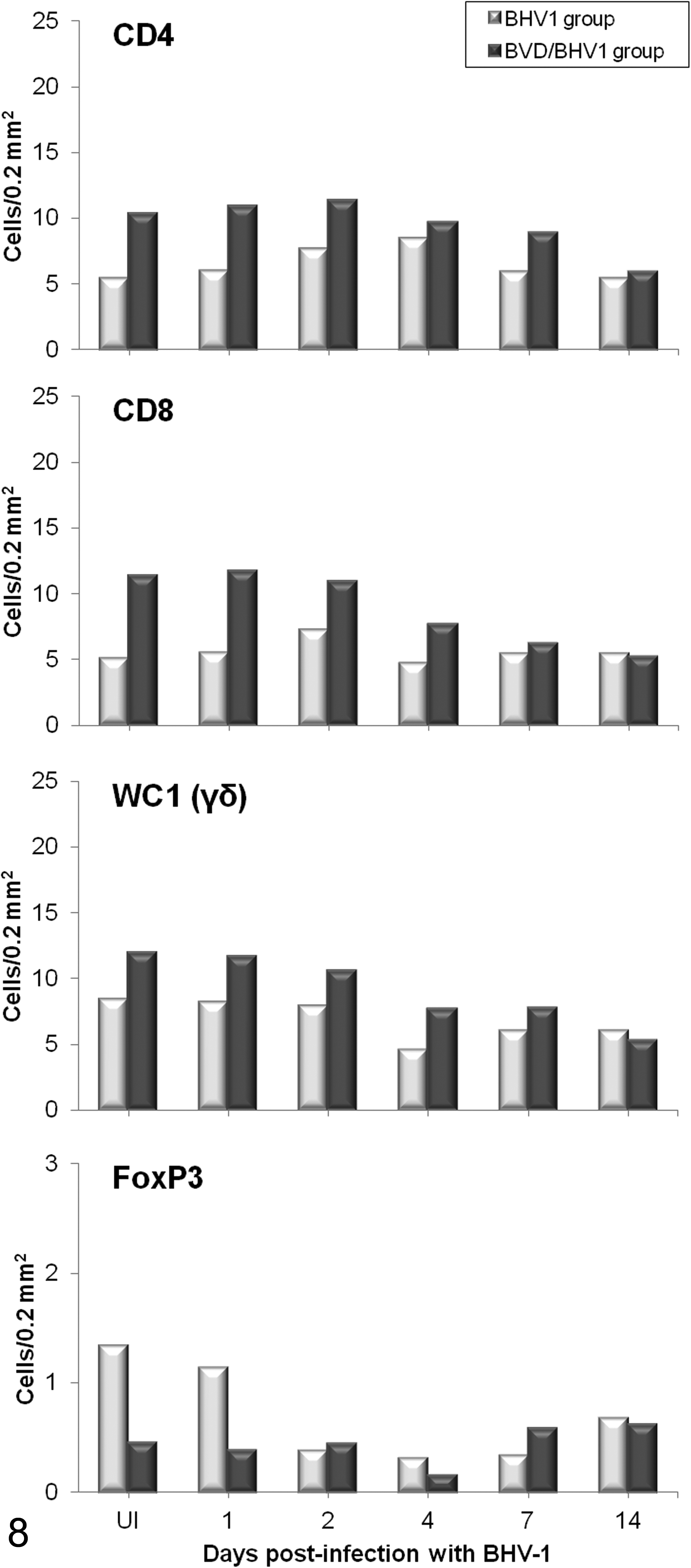
Mean number (n = 2 per time point) of CD4+ T lymphocytes, CD8+ T lymphocytes, γδ+ T lymphocytes, and FoxP3+ regulatory T lymphocytes immunolabeled in the lungs of calves inoculated with herpesvirus 1 subtype 1 (BHV-1.1) vs calves inoculated with bovine viral diarrhea virus (BVDV) and BHV-1.1. (UI, BHV-1.1 uninfected: negative controls for the BHV-1 group and BVDV infection controls for the BVDV/BHV-1 group).
Th1 cytokines, such as IL-12 and IFNγ, which are critical in the cell immune response and play an important role in host defense systems against intracellular microbial agents and viruses, increased after BHV-1.1 inoculation in both inoculated groups, although there were differences in the establishment of this response. In the BHV-1 group, there was an increase at 4 dpi of IMΦs positive for IL-12 and lymphocytes producing IFNγ, whereas the animals previously infected with BVDV displayed a peak of these cytokines at later stages (7 and 14 dpi for IL-12 and 7 dpi for IFNγ) (Fig. 9).
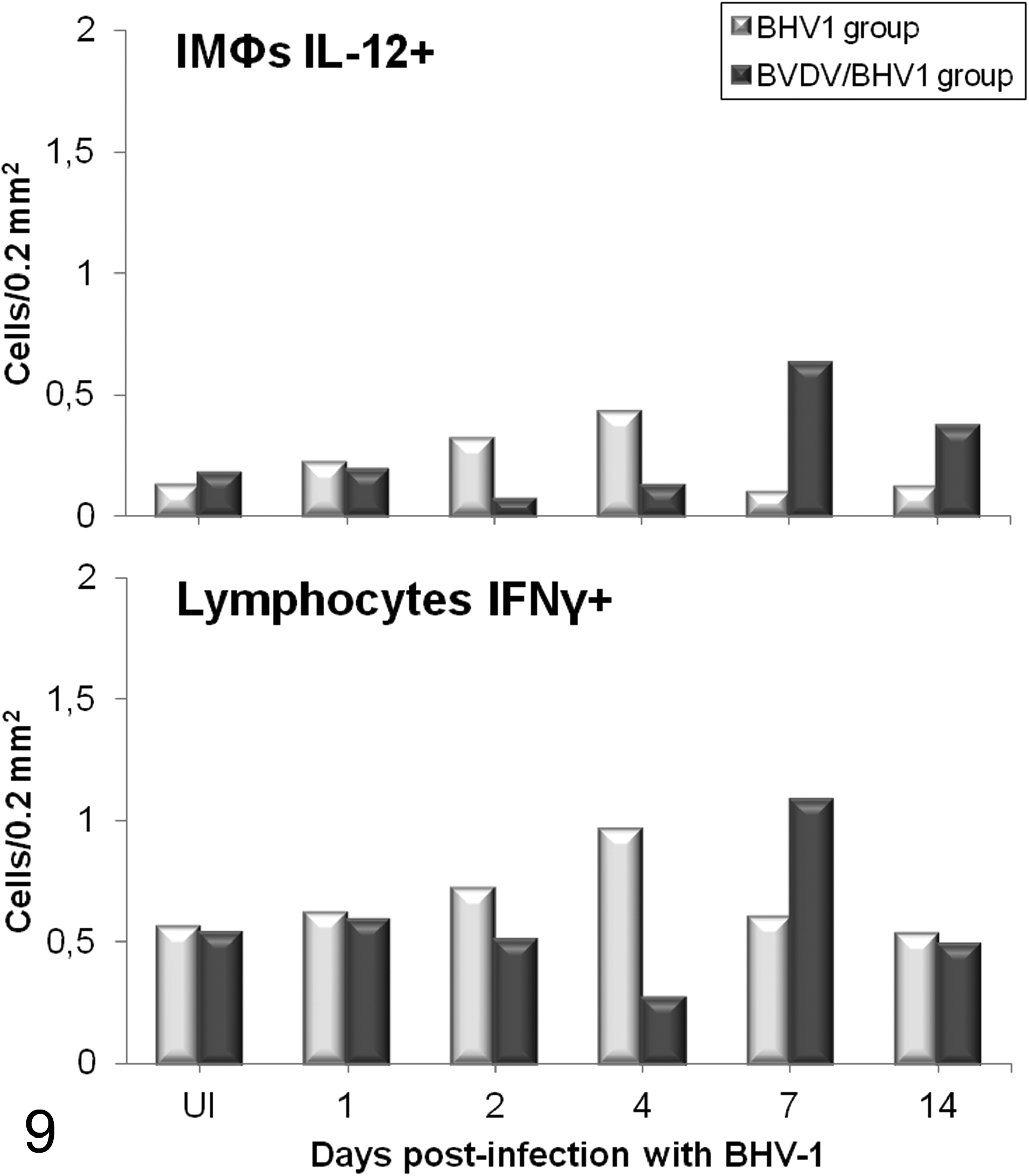
Mean number (n = 2 per time point) of interleukin (IL)–12+ interstitial macrophages (IMΦs) and interferon γ+ (IFNγ+) lymphocytes immunolabeled in the lung of calves coinfected experimentally with bovine viral diarrhea virus (BVDV) and herpesvirus 1 subtype 1 (BHV-1.1) compared with calves inoculated only with BHV-1.1. (UI, BHV-1.1 uninfected: negative controls for the BHV-1 group and BVDV infection controls for the BVDV/BHV-1 group).
Immunohistochemical examination of IL-10 was based on analysis of the expression of this cytokine in different populations of immunocompetent cells, which were distinguished by their morphology. Immunolabeling of IMΦs displayed an intensely positive dark granular and cytoplasmic reaction, whereas the lymphocytes were uniformly immunopositive in all the animals, although there were observable differences in magnitude and kinetics between single and dual infections. Animals preinfected with BVDV demonstrated a lower number of IL-10+ IMΦs and lymphocytes compared with healthy calves. After BHV-1 inoculation, the immunoreactivity of this cytokine in calves affected by dual infections remained low throughout the experiment, while healthy animals infected only with BHV-1 showed a decrease in the number of IL-10+ IMΦs. However, IL-10+ lymphocytes exhibited an increase in the BHV-1 group, reaching the highest levels at 7 dpi and recovering values close to preinoculation levels on the last day; meanwhile, IL-10+ lymphocytes in the BVDV/BHV-1 group showed a trend toward increasing subsequent to infection with the secondary agent (Fig. 10).
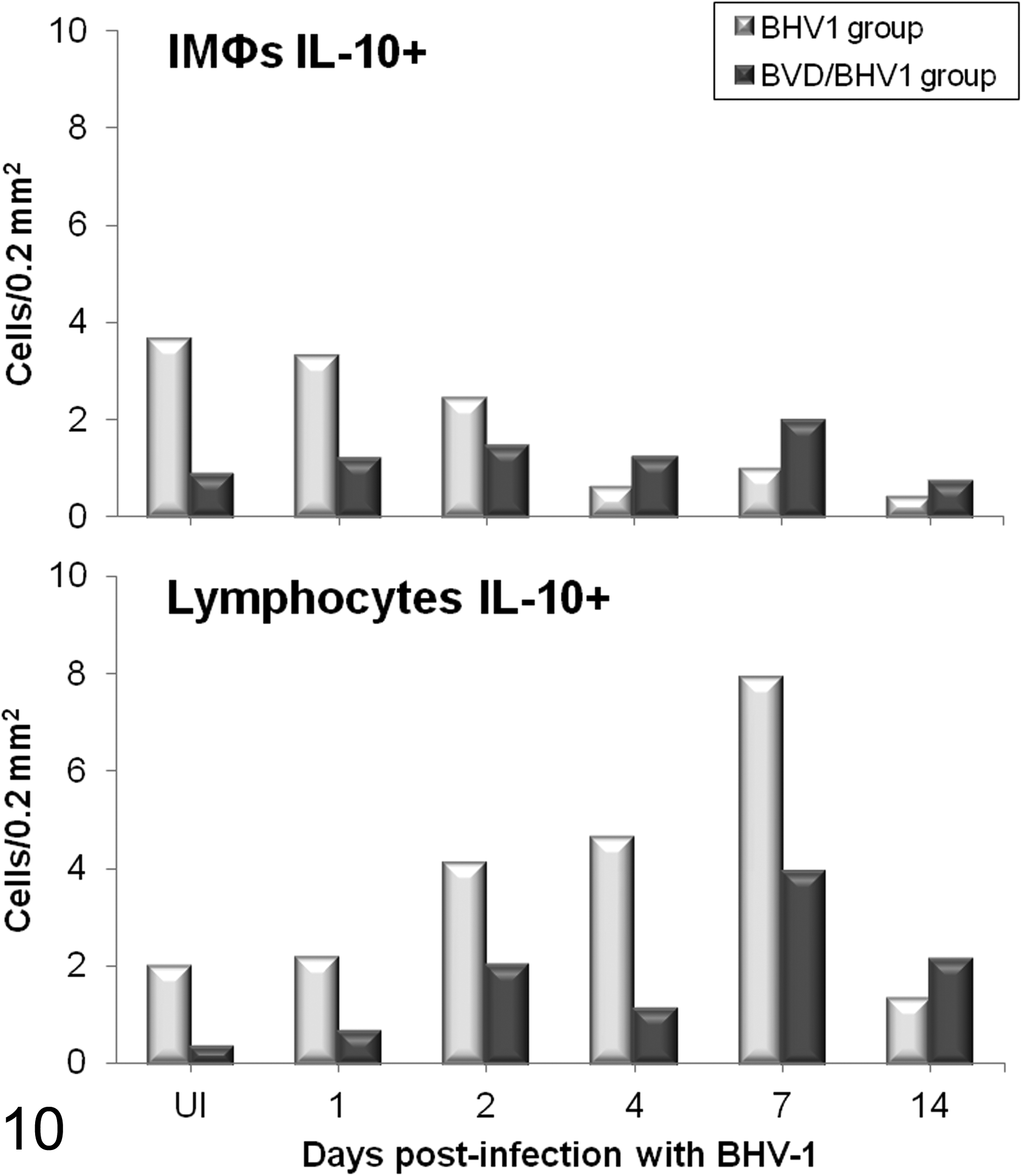
Mean number (n = 2 per time point) of interleukin (IL)–10+ interstitial macrophages (IMΦs) and IL-10+ lymphocytes immunolabeled in the lungs of calves inoculated with herpesvirus 1 subtype 1 (BHV-1.1) vs calves inoculated with bovine viral diarrhea virus (BVDV) and BHV-1.1. (UI, BHV-1.1 uninfected: negative controls for the BHV-1 group and BVDV infection controls for the BVDV/BHV-1 group).
Discussion
The results obtained in this experimental model confirmed and elaborated on the immunological and histopathologic effects of preinfection with BVDV and subsequent coinfection with BHV-1 in calves, which have been previously described in Risalde et al.46 –48 In these publications, it was observed that following the infection with BHV-1, the calves showed changes in pro- and anti-inflammatory cytokine profiles, based on a greater secretion of proinflammatory cytokines (TNFα) and reduced production of anti-inflammatory cytokines (IL-10), leading to an exacerbation of the inflammatory response and to the development of intense clinical respiratory signs and pulmonary lesions. Cytokines involved in the cytotoxic mechanisms responsible for limiting the spread of different pathogens, such as IFNγ, also showed an altered secretion in calves preinfected with BVDV, thus facilitating the spread of BHV-1. Moreover, the presence of BHV-1 favors the persistence of BVDV in target organs and BVDV-related viremia, enhancing this concomitance through a synergic action of their pathogenic mechanisms and a disruption of the maintenance of pulmonary homeostasis by facilitating the establishment of an inflammatory and procoagulant environment, characteristic of the BRDC (Fig. 11).
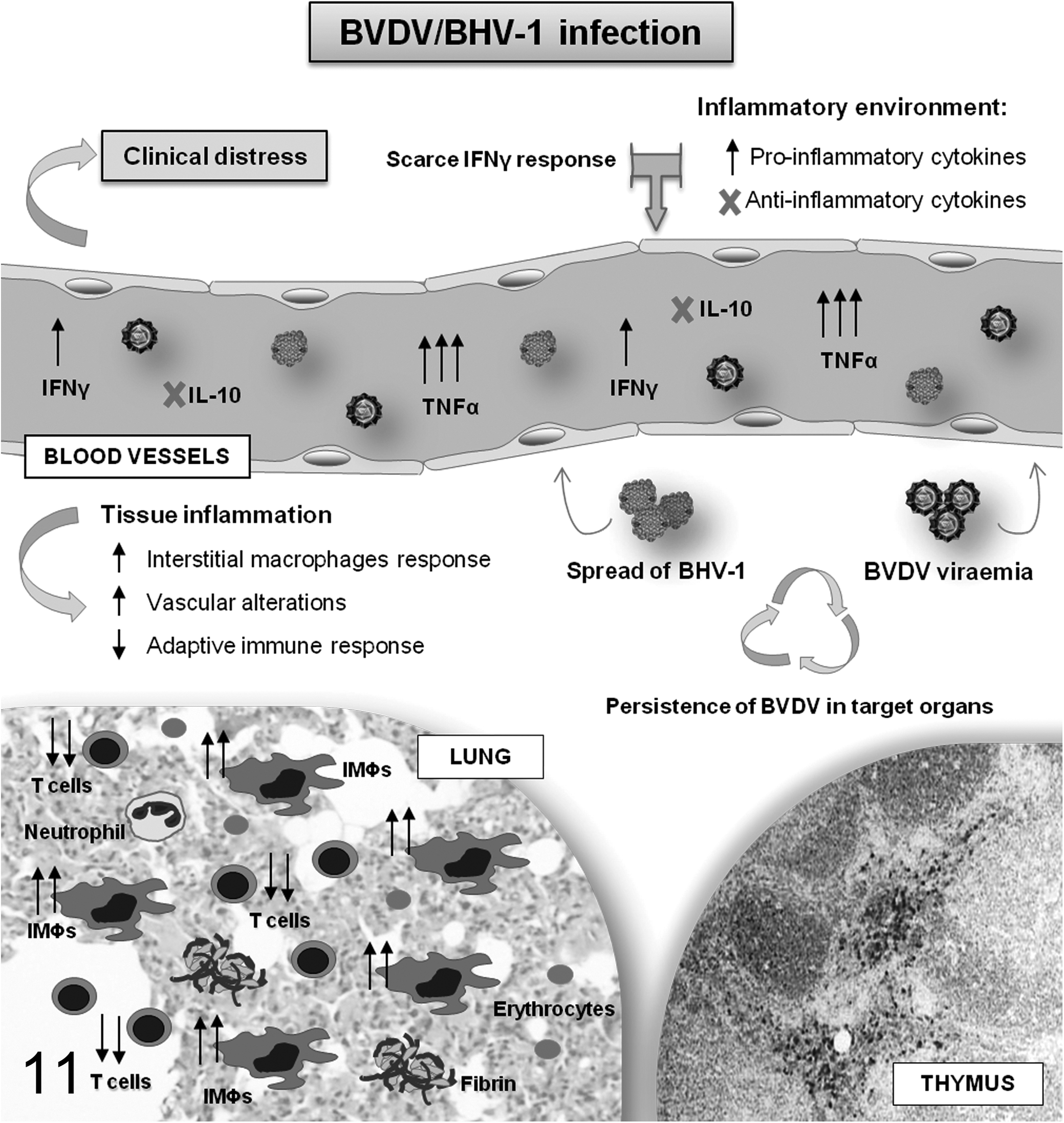
Schematic summary of the main results obtained in the experimental model related to the inflammation and spread of the viruses in calves preinfected intranasally with bovine viral diarrhea virus (BVDV) and challenged at 12 days postinfection (dpi) with bovine herpesvirus 1 subtype 1 (BHV-1.1). IMΦs, interstitial macrophages.
The objective of the present study was to evaluate and characterize the mononuclear cell infiltrates observed in the pulmonary parenchyma of healthy calves and calves with subclinical bovine viral diarrhea (BVD) experimentally inoculated with BHV-1.1, as well as to clarify the role of BVDV on these immunocompetent cells in the local response to the secondary pathogen.
The results demonstrated that following BHV-1 inoculation, both groups of calves presented with a mononuclear cell infiltrate in the pulmonary interstitium comprising mainly IMΦs and T lymphocytes. This mononuclear infiltrate, which was the main component of the alveolar septal thickening consistent with an interstitial pneumonia, increased at 4 dpi and was associated with the most severe respiratory signs in both inoculated groups. The calves preinfected with BVDV presented with respiratory signs such as cough, mucopurulent nasal discharge, dyspnea, and open-mouth breathing, which were consistent with the histopathologic severity of the pulmonary lesions; meanwhile, the calves of the BHV-1 group presented only with a moderate serous nasal discharge. 46
With regard to the immunoreactive cells analyzed in these interstitial aggregates, a marked decrease in the number of Tγδ+ lymphocytes was observed in both inoculated groups from 4 dpi; such cells play a key role in the early stages of inflammation by stimulating the rapid flow of lymphocytes and monocytes to the site of infection. 56
On the other hand, the increase in B-lymphocyte numbers was more marked and occurred earlier in infection in those calves inoculated with BHV-1 alone. The attenuated B-lymphocyte response observed in the BVDV/BHV-1 group may be linked to a severe lymphocyte depletion in BVDV target lymphoid organs, a lesion that has already been described in calves 46 and in other in vivo infections with BVDV, 31,32,37,38,43 in which the depletion has been correlated to the action of apoptotic mechanisms developed during BVD.
A difference between groups was observed in the behavior of CD8+, CD4+, and Foxp3+ regulatory T cells, as well as in the different cytokines studied in response to the secondary infectious agent. After BHV-1 inoculation and according to the systemic findings, 35 healthy calves demonstrated a slight and early increase of CD8+ T lymphocytes in the lungs. These cytotoxic cells appear to have a key role in preventing the spread of this pathogen. 36,56 There was also an increase in the number of CD4+ T lymphocytes at 4 dpi, coinciding with the detection of BHV-1 and with marked responses of IL-12 and IFNγ, cytokines produced by Th1 cells and known to have a major antiviral function and the ability to induce the lysis of BHV-1–infected cells. 2,5,51 In addition, Foxp3+ T regulatory cells as well as IL-10 play an important role in the regulation of inflammatory responses, primarily limiting tissue damage but in some cases also promoting persistence of pathogens. 4 –6 Differences in their cell expression was observed between healthy animals and calves inoculated with BVDV, insomuch as the calves preinfected with BVDV demonstrated a lower expression of Foxp3+ T regulatory cells with respect to healthy animals, a fact that could account for the earlier inflammatory changes observed in this group subsequent to BHV-1 inoculation. However, after the challenge with the virus, calves of the BHV-1 group also showed a loss of expression of Foxp3, which was likely due to the inhibition in the production of Foxp3+ T regulatory cells from naive CD4+ cells and the conversion of T regulatory cells into Foxp3– T effector cells as influenced by IL-1, IL-12, and IFNγ during the inflammatory processes in peripheral tissues. 13 These Foxp3– cells may correspond to a T regulatory 1 cell subpopulation, which can be generated and mediate their suppressive activity through IL-10 activity. 50,57 Thus, IL-10 and IFN-γ production by lymphocytes seems to be related to antigen stimulation in the calves infected only with BHV-1, contributing to its control both in limiting the inflammatory response and in clearing the virus, as has been observed in other infectious diseases. 11,12,16
The presence of BVDV was detected by polymerase chain reaction (PCR) in the lung of the BVDV/BHV-1 group from 0 to 7 dpi (12–19 dpi BVDV) and the BHV-1 from 2 to 14 dpi; likewise, in calves of the BHV-1 group, BHV-1 was detected from 4 to 7 dpi with concurrent confirmation that the animals were BVDV negative. 46,47
Taken in conjunction, these results suggest that calves in the BHV-1 group develop an adequate adaptive immune response to BHV-1 at the pulmonary level, contributing to the elimination of the virus at 7 dpi. However, BVDV preinfected calves, while displaying greater CD8+ and CD4+ T-lymphocyte numbers prior to BHV-1 inoculation, failed to respond to the presence of this pathogen in the lung (from 2 dpi); indeed, a lymphocyte depletion was observed in these animals from 4 dpi. This inhibition in the proliferative response of CD8+ and CD4+ T lymphocytes, which has also been reported in infections with NCP strains of BVDV, 26,34 coupled with alterations in IFNγ production and a slight expression of IL-10, 30,45,48 might impair the development of an adaptive immune response to the secondary viral agent in the lung, thereby producing significant inflammatory lesions and favoring the persistence of BHV-1 in the lungs, where it was detected until the end of the study (14 dpi).
In conclusion, animals preinoculated with BVDV, despite having a transient infection, exhibited an alteration in the response of Th1 cytokines that play a key role in the immune response, as well as having an impaired local cell-mediated response to the secondary respiratory pathogen, marked essentially by an inhibition in the response of CD8+ and CD4+ lymphocytes. These observations, along with the fact that the alteration in the IL-10 response could mediate T regulatory cells function in the lung, suggest that further studies are required to increase our understanding of the role of this cell population in the pathogenesis of BVD and their possible use for immunotherapy.
Footnotes
Acknowledgements
We thank the Institute für Virologie, TIHO (Hannover, Germany) and Laboratorios HIPRA SA (Girona, Spain) for providing the BVDV and BHV-1.1, respectively, and the farm “Las Rozuelas del Valle” (Torrecampo, Spain) for providing the animals. We thank specially to G. Muñoz for her technical assistance.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the Junta de Andalucía-FEDER (P09-AGR-4671). M. A. Risalde and V. Molina were supported by a predoctoral grant from the Spanish Ministry of Education and Science. P. J. Sánchez-Cordón holds a contract from the “Ramón y Cajal programme” of Spanish Ministry of Education and Science. F. Romero-Palomo was supported by a predoctoral grant from the Andalusian Regional Government-ERDF associated with project P09-AGR-4671.