
Abstract
Granular cell tumors (GCTs) are histologically characterized by polygonal neoplastic cells with abundant eosinophilic cytoplasmic granules. In humans, these cells are considered to be derived from Schwann cells, and the cytoplasmic granules are assumed to be autophagosomes or autophagolysosomes. However, the origin and nature of the cytoplasmic granules in canine GCTs have not been well characterized. The present study examined 9 canine lingual GCTs using immunohistochemistry, transmission electron microscopy (TEM), and cell culture and xenotransplantation experiments. In some cases, the tumor cells expressed S100, CD133, and desmin. The cytoplasmic granules were positive for LC3, p62, NBR1, and ubiquitin. TEM revealed autophagosome-like structures in the cytoplasm of the granule-containing cells. The cultured GCT cells were round to spindle shaped and expressed S100, nestin, Melan-A, CD133, LC3, p62, NBR1, and ubiquitin, suggesting that they were of neural crest origin, redifferentiated into melanocytes, and exhibited upregulated autophagy. The xenotransplanted tumors consisted of spindle to polygonal cells. Only a few cells contained cytoplasmic granules, and some had melanin pigments in their cytoplasm. The xenotransplanted cells expressed S100, nestin, Melan-A, and CD133. P62 and ubiquitin were detected, regardless of the presence or absence of cytoplasmic granules, while LC3 and NBR1 were detected only in the neoplastic cells containing cytoplasmic granules. These findings suggest that some xenotransplanted cells redifferentiated into melanocytes and that autophagy was upregulated in the cytoplasmic granule-containing cells. In conclusion, canine lingual GCTs originate from the neural crest and develop cytoplasmic granules via autophagy. In addition, the microenvironment of GCT cells affects their morphology.
Keywords
Granular cell tumors (GCTs) are histologically characterized by proliferating uniform polygonal neoplastic cells that contain abundant eosinophilic granules in their cytoplasm. In humans, GCTs most commonly occur in the tongue, although they can develop in any organ. 13 In dogs, the most common location for GCTs is the oral cavity. 10 GCTs are usually benign, but in rare cases, they can become malignant in both humans and animals. 4,13 In humans, at least 3 of the following criteria need to be met for a diagnosis of malignant GCTs: necrosis, spindle cell proliferation, vesicular nuclei with large nucleoli, increased mitotic activity, a high nuclear-cytoplasmic ratio, and pleomorphism. 11
The histogenesis of GCTs remains unclear. 4,13 Previously, GCTs have been considered to originate from skeletal muscles, neurons, fibroblasts, histiocytes, and myoepithelial cells. 12 Moreover, degenerative changes or trauma-induced proliferation, which occur in a variety of normal and neoplastic cells, were also considered to be involved in the morphogenesis of GCTs. 6,13 Recent reports based on immunohistochemical and electron microscopic studies of humans and animals have suggested that GCTs are derived from neural crest tissue, especially Schwann cells. 4,6,8,11,13,15 Neural crest cells give rise to neurons, glial cells, Schwann cells, pigmented cells such as melanocytes, endocrine cells, muscle cells, craniofacial cartilages, and bones. 3,16,19 Peripheral nerve sheath tumors (PNSTs) also originate from Schwann cells, perineurial cells, or both, 18 although the histopathologic features of GCTs differ from those of PNSTs.
The cytoplasmic granules found in GCT cells have been suggested to be lysosomes, autophagosomes, or autophagolysosomes based on their ultrastructural features. 6,8,15 Autophagy is a process by which cells selectively digest some of their own components, which helps to maintain vital cellular functions during periods of fasting by providing nutrients and removing excessive or damaged organelles, misfolded proteins, and invading microorganisms. Autophagy is rapidly upregulated during periods of starvation, growth factor withdrawal, high bioenergetic demand, developmental transition, oxidative stress, infection, or protein aggregate accumulation. Nutritional status, hormonal factors, temperature, the oxygen concentration, and cell density are involved in the control of autophagy. 5,20 LC3, an autophagy marker, has been detected in human GCT cells, indicating that the cytoplasmic granules seen in these cells are actually autophagosomes or autophagolysosomes. 15
No previous studies have confirmed the exact nature of these cytoplasmic granules in canine GCTs. To elucidate the origin and exact nature of the cytoplasmic granules of GCT cells, 9 canine lingual GCTs were pathologically examined. Immunohistochemistry was performed on all 9 tumors. In addition, each transmission electron microscopy and cell culture and xenotransplantation experiment was performed on only 1 case, respectively.
Materials and Methods
Cases
Nine canine lingual tumors were submitted to the Japan Animal Referral Medical Center or the Department of Veterinary Pathology, Graduate School of Agricultural and Life Sciences, the University of Tokyo, between 2010 and 2013. The case history, lingual location, and behavior of the tumors are summarized in Table 1. No breed, age (10–15 years), or sex (3 males and 6 females) predisposition was detected.
Cases Examined in the Present Study.
aY, years; M, months.
bF, female; M, male; S, spayed.
Histopathology and Immunohistochemistry
Tissue samples were obtained by excision biopsy, fixed in 10% phosphate-buffered formalin solution, and embedded in paraffin. In addition, fresh tumor tissue was obtained from case No. 2 and used for the primary culture experiments. Two- to 4-μm-thick deparaffinized sections were stained with hematoxylin and eosin (HE) or periodic acid–Schiff (PAS) stain, and diastase resistance was tested on each section. Immunohistochemistry was performed using the primary antibodies listed in Table 2. Antigen retrieval was performed to detect S100, vimentin, Melan-A, CD133, LC3, p62, NBR1, Iba1, and desmin by autoclaving the deparaffinized sections in citrate buffer (pH 6.0) at 121°C for 10 minutes. No pretreatment was performed in the examinations of nestin, nerve growth factor receptor (NGFR), ubiquitin, and α–smooth muscle actin (SMA) expression. Endogenous peroxidase activity was inactivated by treating the sections with 3% hydrogen peroxide in methanol at room temperature (RT) for 5 minutes. Nonspecific reactions were blocked by incubating the sections in 8% skimmed milk in Tris-buffered saline (TBS) at 37°C for 30 minutes. The sections were then incubated with each primary antibody at 4°C overnight. After washing the sections 3 times with TBS, they were incubated with Dako EnVision+ System horseradish peroxidase (HRP)–labeled polymer anti–mouse or anti–rabbit secondary antibodies (DAKO, Glostrup, Denmark) at RT for 10 minutes, then at 37°C for 40 minutes, and finally at RT for a further 10 minutes. After being washed a further 3 times with TBS, the sections were developed with 0.05% 3,3′-diaminobenzidine and 0.03% hydrogen peroxide in Tris-HCl buffer. Mayer’s hematoxylin was used as a counterstain. The tissue sections used as positive controls are listed in Table 2. The primary antibodies were replaced with TBS to produce the negative control.
Antibodies Used in the Present Study.
ICC, immunocytochemistry; IHC, immunohistochemistry.
Transmission Electron Microscopy
Two 2- to 3-mm cubes of formalin-fixed tumor tissue from case No. 9 were washed with 0.1 M phosphate buffer (PB) before being refixed in 2.5% glutaraldehyde in 0.1 M PB at 4°C for 2 hours and then in 1% osmium tetroxide at 4°C for 2 hours. After being dehydrated in increasing concentrations of ethanol and treated with QY-1 (Nisshin EM Corporation, Tokyo, Japan), the tissues were embedded in Quetol651 resin (Nisshin EM Corporation). Ultrathin sections were stained with uranyl acetate and lead citrate and examined with a Hitachi H-7500 transmission electron microscope (Hitachi High-Technologies Corporation, Tokyo, Japan).
Cell Culture and Xenotransplantation of Cultured Cells Into Immunodeficient Mice
Tumor cells were dissociated from the minced fresh tumor tissue of case No. 2 and seeded in a cell culture flask containing Dulbecco’s modified Eagle’s medium (DMEM; Life Technologies Corporation, Carlsbad, CA) supplemented with 10% fetal bovine serum (FBS; Thermo Fisher Scientific, Waltham, MA) and 1% penicillin-streptomycin (Wako Pure Chemical Industries, Osaka, Japan). The cells were passaged when they reached confluence. Immunocytochemistry was performed using the above-mentioned antibodies at the eighth passage to confirm the characteristics of the cells.
The cultured cells were harvested at the 13th passage before being collected in 3 syringes containing 200 μl phosphate buffered saline (PBS). Then, the 3 cell batches were injected into the lower back subcutaneous tissue of 3 female 5-week-old nude mice (BALB/cAJcl-nu/nu; CLEA Japan, Tokyo, Japan) to examine their transplantability. Each nude mouse was injected with 1.0 × 106 cells. Furthermore, to examine the morphological alterations caused by the type of immunodeficiency, cultured GCT cells at another passage were injected into the lower backs of 3 female 5-week-old SCID mice (FOX CHASE SCID C.B-17/lcr-scid/scidJcl; CLEA Japan). The numbers of cells injected into each SCID mouse were 3.7 × 106, 2.2 × 106, and 1.1 × 106, respectively. After a mass was palpable at the injection site, the tumor volume was calculated every day using the following formula: (length) × (width) × (height) × 3.14 / 6. The end points of the experiment were as follows: (1) when the tumor volume exceeded 10% of the mouse’s body weight, (2) when a mouse lost 20% of its body weight within 2 or 3 days, or (3) when a mouse lost 25% of its body weight within a week. At each end point, the mice were euthanized, and their tumor was resected. The tumor masses were fixed in 10% phosphate-buffered formalin solution. Then, 2- to 4-μm-thick paraffin sections were stained with HE, PAS, or Fontana-Masson silver stain, and immunohistochemistry was performed using the above-mentioned antibodies. All animal experiments were approved by the Committee of Animal Experiments, Graduate School of Agricultural and Life Sciences, the University of Tokyo.
Results
Histopathology and Immunohistochemistry
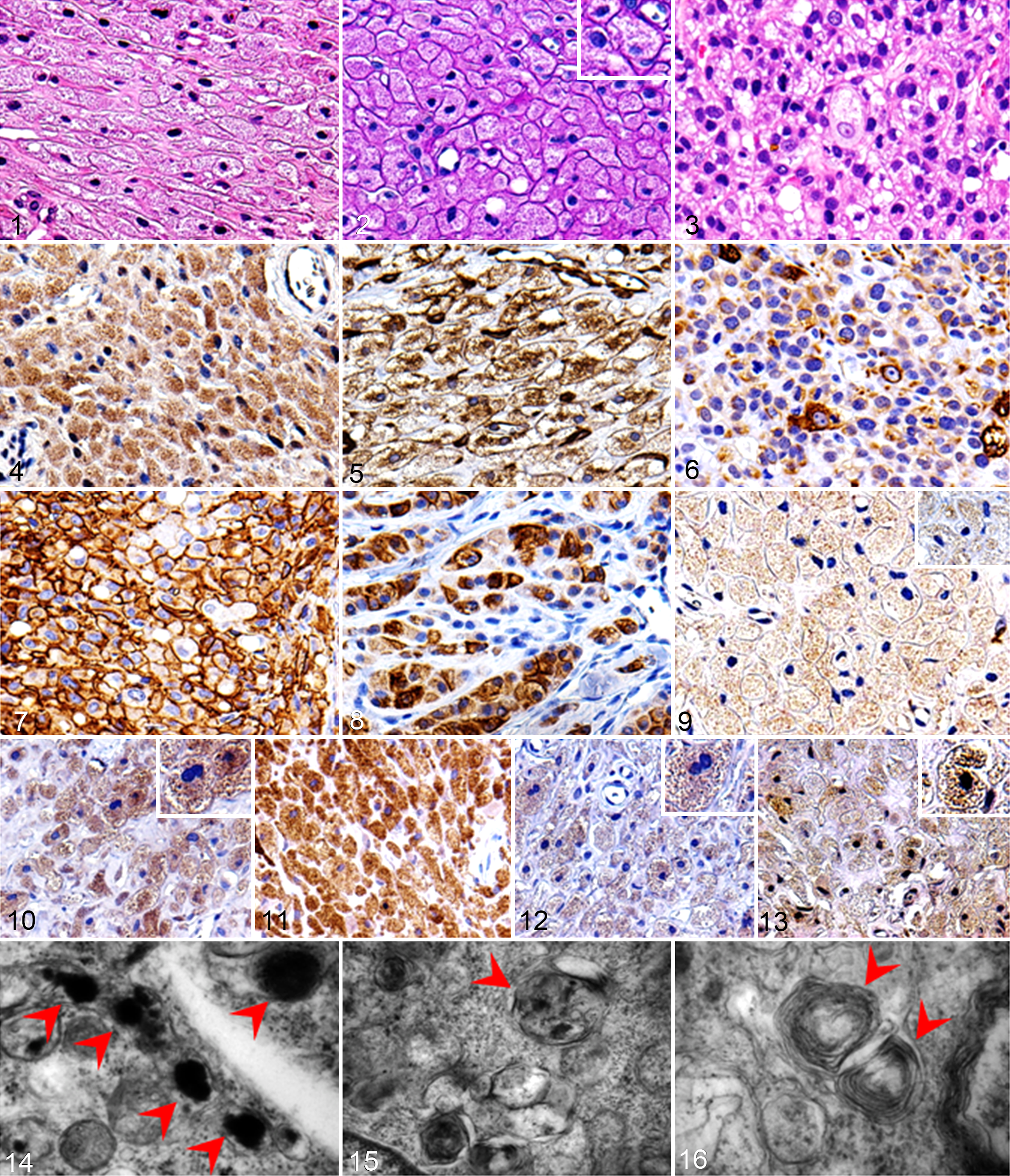
All tumors were histopathologically diagnosed as GCTs, and all except 1 (case No. 9) were benign. The tumor tissues consisted of large round to polygonal cells with abundant eosinophilic cytoplasmic granules (Fig. 1). The granules were PAS positive and diastase resistant in all cases (Fig. 2). The tumor in case No. 9 was considered malignant according to the criteria of human malignant granular cell tumor. It consisted of pleomorphic spindle to polygonal cells. These cells exhibited anisokaryosis, anisocytosis, and increased mitotic activity. A very small number of cells contained abundant eosinophilic cytoplasmic granules (Fig. 3). The neoplastic cells had invaded muscular tissue, and wide-ranging necrotic regions were observed.
Tongue; dog, granular cell tumor (GCT).
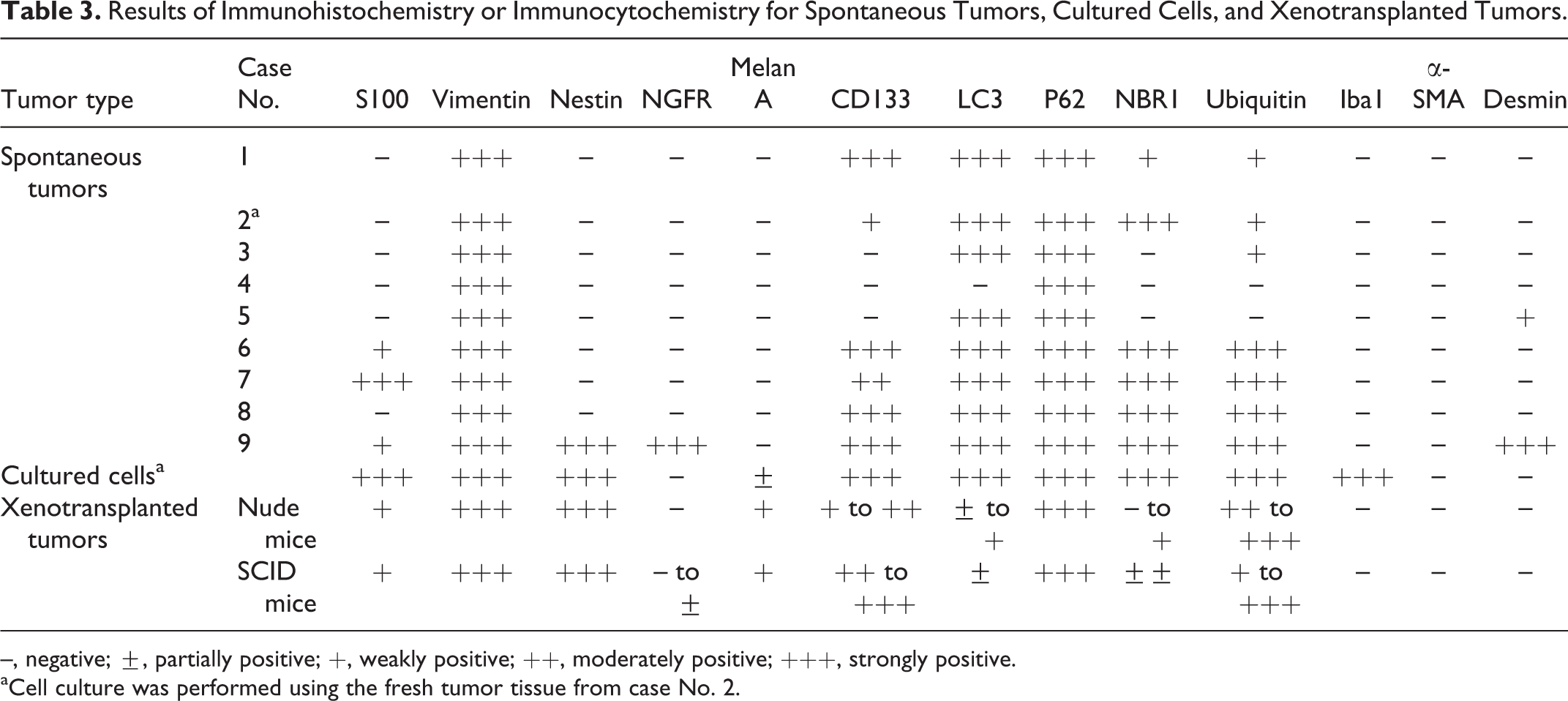
The results of the immunohistochemical examinations are summarized in Table 3. S100 expression was detected in the cytoplasm of the neoplastic cells in case Nos. 6, 7, and 9. Strong S100 expression was detected in case No. 7 (Fig. 4), and weak expression was seen in case Nos. 6 and 9. Vimentin was strongly expressed in the neoplastic cells in all cases (Fig. 5). Nestin was strongly expressed in the neoplastic cells in only case No. 9 (Fig. 6). NGFR was strongly expressed along the cellular membrane of the neoplastic cells in only case No. 9 (Fig. 7). Desmin was detected in the cytoplasm of the neoplastic cells in case Nos. 5 and 9. Case No. 5 exhibited weak desmin expression, whereas case No. 9 exhibited strong desmin expression (Fig. 8). The tests for Melan-A, Iba1, and α-SMA produced negative results in all cases. CD133 expression was detected in the cytoplasmic granules of the neoplastic cells in case Nos. 1, 2, 6 to 8, and 9. Its staining intensity was weak in case No. 2 (Fig. 9), moderate in case No. 7, and strong in case Nos. 1, 6, 8, and 9. Expression of autophagy markers (LC3, p62, NBR1 and ubiquitin) was also observed in the cytoplasmic granules. Strong LC3 expression was detected in 8 cases (all cases except No. 4; Fig. 10). Strong p62 expression was detected in all cases (Fig. 11). NBR1 was detected in case Nos. 1, 2, 6 to 8, and 9 (Fig. 12). It exhibited strong staining intensity in case Nos. 2, 6 to 8, and 9 and weak staining intensity in case No. 1. Ubiquitin expression was detected in case Nos. 1 to 3, 6 to 8, and 9 (Fig. 13). Case Nos. 6 to 9 exhibited strong expression, and case Nos. 1 to 3 exhibited weak expression.
Results of Immunohistochemistry or Immunocytochemistry for Spontaneous Tumors, Cultured Cells, and Xenotransplanted Tumors.
–, negative; ±, partially positive; +, weakly positive; ++, moderately positive; +++, strongly positive.
aCell culture was performed using the fresh tumor tissue from case No. 2.
Transmission Electron Microscopy
Transmission electron microscopy (TEM) revealed that the neoplastic cells in case No. 9 contained many vacuoles and lysosome-like electron dense granules in their cytoplasm (Fig. 14). Autophagosome-like structures containing several organelles (Fig. 15) and myelin-like membranous structures (Fig. 16) were also found.
Cell Culture
During the first few passages, round or spindle cells were found in the flask, but round cells predominated during the later passages. The doubling time of the cloned cultured cells was 30.6 hours (Fig. 17). The shape of the cloned cultured cells was spindle to round (Fig. 18). Their nuclei were round to oval and contained several prominent nucleoli, and the nuclear-cytoplasmic ratio was high. These cells exhibited anisokaryosis and anisocytosis. The results of the immunocytochemical examinations of the cultured cells are summarized in Table 3. The cultured cells expressed S100, vimentin, nestin, Melan-A, CD133, LC3, p62, NBR1, ubiquitin, and Iba1. All of these molecules except for Melan-A exhibited strong expression. Melan-A was observed as several dots in the cytoplasm in a few cells. No NGFR, α-SMA, or desmin expression was detected.
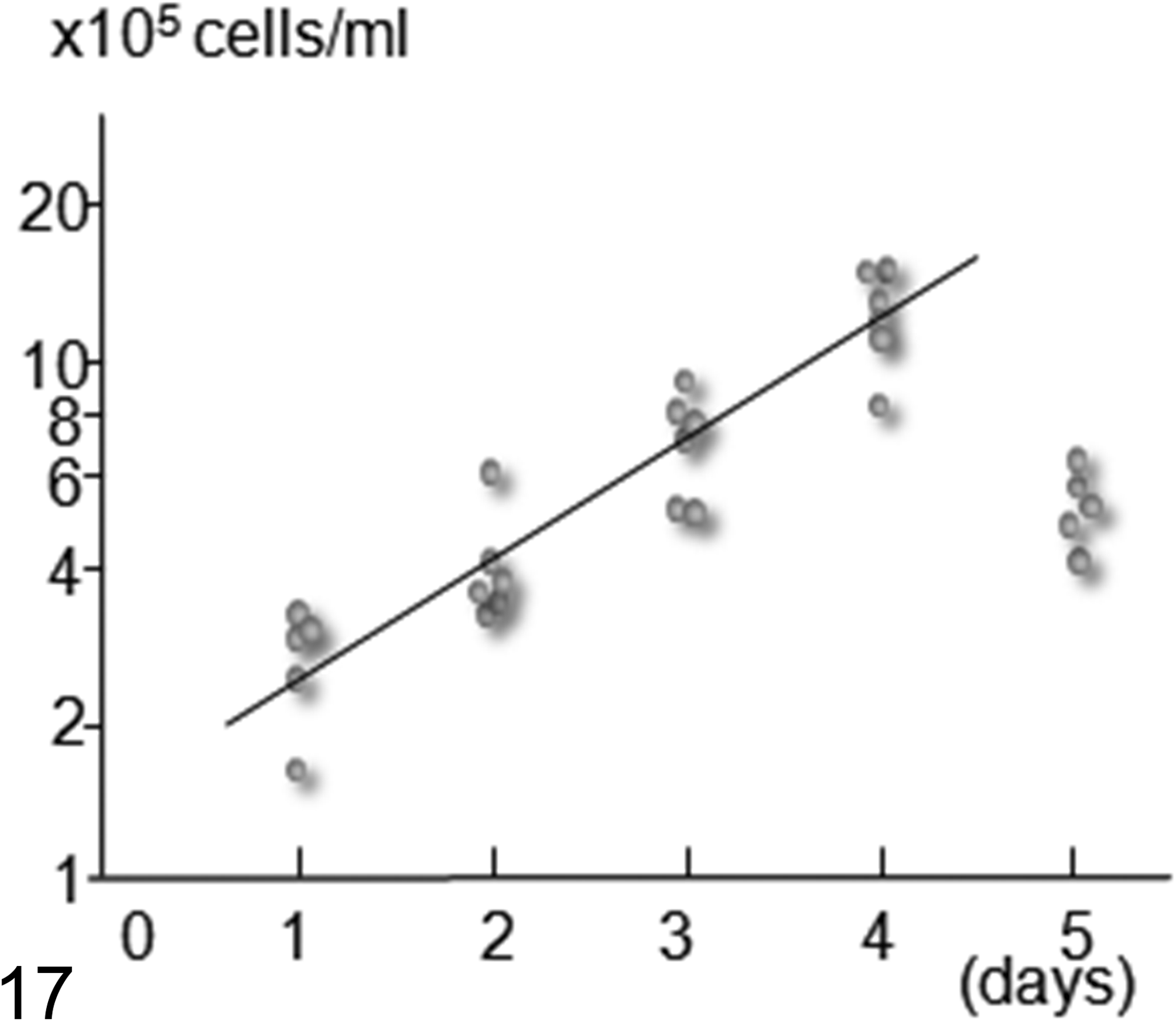
The growth curve of the cloned cultured cells sampled from the fresh tumor tissue of case No. 2. The doubling time was 30.6 hours. The longitudinal axis represents the logarithmic number of cultured cells. The horizontal axis represents the culture period (number of days).
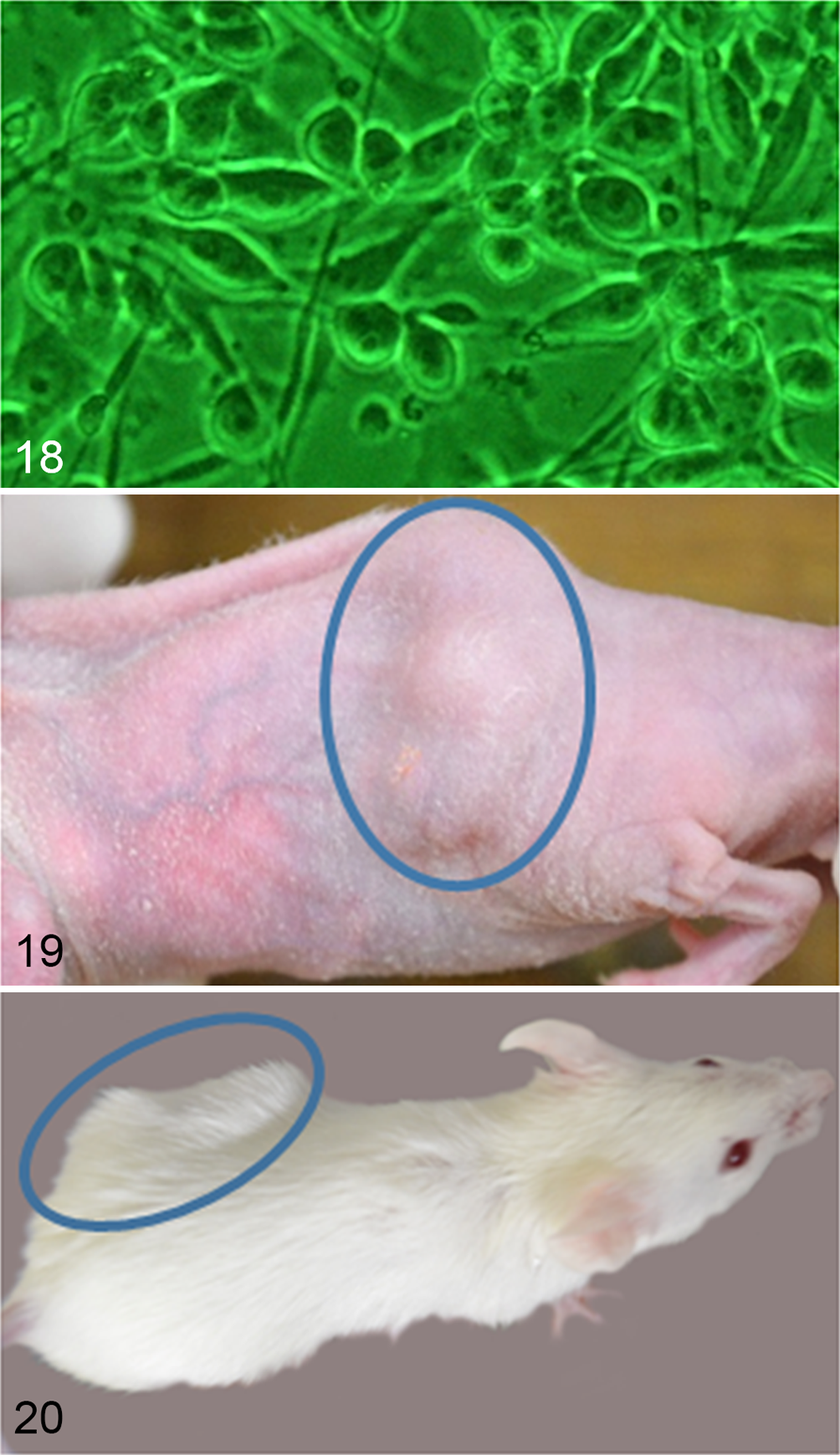
Tongue; dog, granular cell tumor (GCT). Case No. 2. The cloned cultured cells obtained from the fresh tumor tissue of case No. 2 exhibit a spindle-shaped or round morphology. Phase-contrast photomicrograph.
Xenotransplantation of Cultured Cells Into Immunodeficient Mice
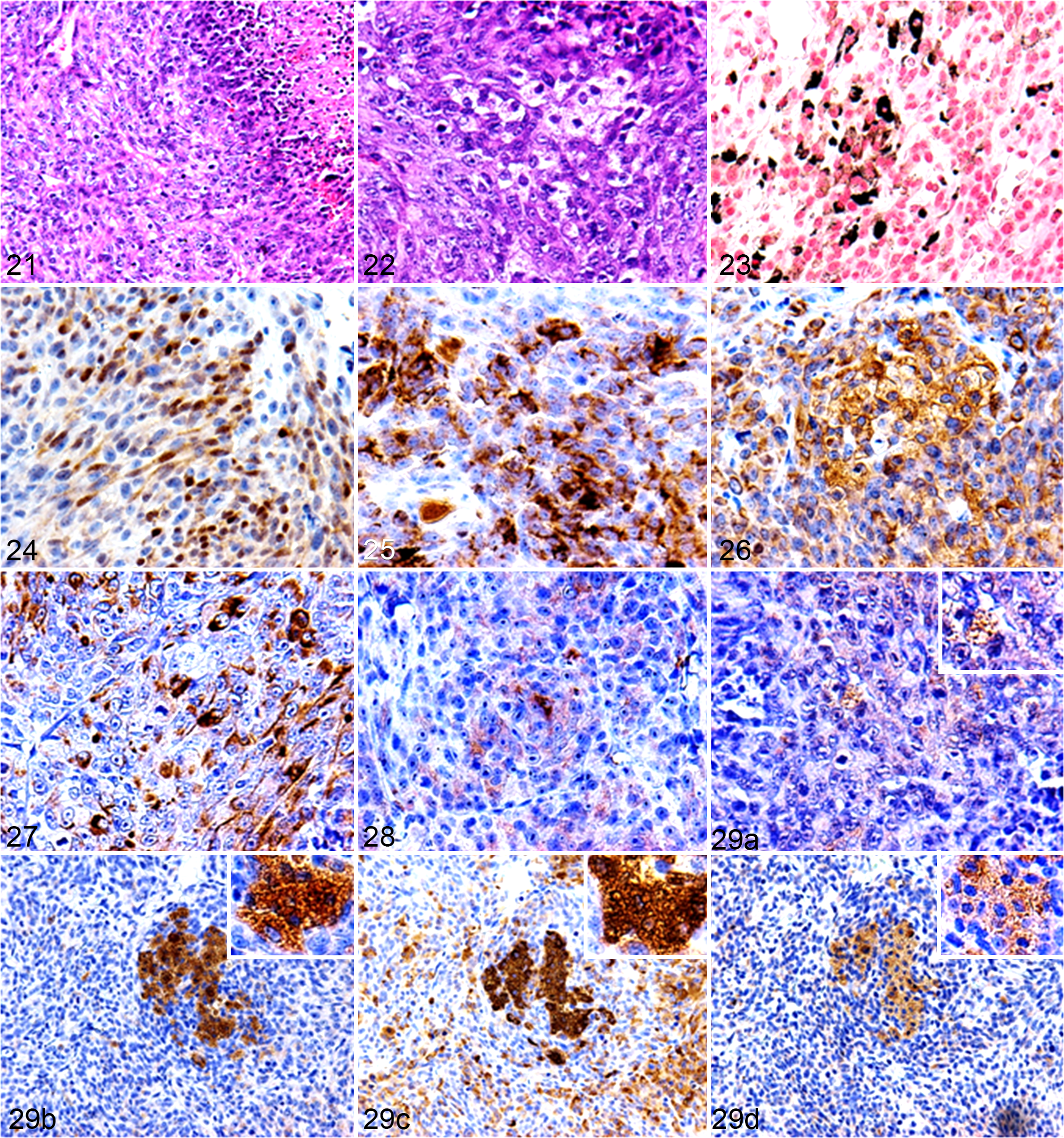
Palpable masses were found in the subcutaneous tissue of the lower backs of 2 of 3 nude mice at 22 and 47 days after inoculation, respectively (Fig. 19), while masses were observed in all SCID mice at 13 to 15 days after inoculation (Fig. 20). The tumor tissues consisted of spindle to polygonal neoplastic cells. These cells exhibited anisocytosis and were arranged in interwoven bundles or whorls, and pseudopalisadings were observed around necrotic lesions (Fig. 21). The nuclei of the neoplastic cells were round to oval and had several large prominent nucleoli. A few cells contained eosinophilic cytoplasmic granules, which were PAS positive and diastase resistant (Fig. 22). In all tumors in nude or SCID mice, Fontana-Masson silver stain revealed that some of the neoplastic cells contained cytoplasmic melanin pigments (Fig. 23).
Subcutis, xenotransplanted tumor; nude mouse. The xenotransplanted tumors consisted of highly pleomorphic cells, and these figures represent the different histologic and immunohistochemical pattern areas of the same tumor arose in a nude mouse.
The results of the immunohistochemical examination of the transplanted tumors are summarized in Table 3. In all tumors, the cytoplasm of the neoplastic cells exhibited S100, vimentin, nestin, Melan-A, CD133, p62, and ubiquitin. S100 and Melan-A were weakly expressed and exhibited localized distributions (Figs. 24 and 25, respectively). Negative to very weak NGFR expression was detected along the membranes of a few tumor cells in SCID mice. Conversely, the expression levels of vimentin and nestin were strong and diffuse (Figs. 26 and 27, respectively). CD133 expression varied from weak to strong (Fig. 28). The cytoplasmic granules of some neoplastic cells were positive for ubiquitin, LC3, p62, and NBR1 (Figs. 29a–d, respectively). LC3 and NBR1 exhibited strong cytoplasmic expression in the granule-containing neoplastic cells, while p62 and ubiquitin expression were detected in the cytoplasm of many neoplastic cells, regardless of the presence or absence of cytoplasmic granules. P62 exhibited strong and diffuse expression, whereas ubiquitin displayed weak to strong expression. Iba1, α-SMA, or desmin expression were not detected.
Discussion
The morphologies of the 8 benign GCTs examined in the present study were consistent with those of the GCTs described in previous reports. 4,6,10 –13,15 Only the tumor in case No. 9 was diagnosed as a malignant GCT. Cytoplasmic granules were found in both the benign and malignant GCTs. The granules were PAS positive and diastase resistant in all cases. These findings suggest that the cytoplasmic granules contained particular glycoproteins regardless of the malignancy of the tumor. S100 and NGFR were detected in the neoplastic cells of case Nos. 6, 7, and 9; desmin expression was observed in only case No. 9; and CD133 was found in case Nos. 1, 2, 6 to 8, and 9. The neoplastic cells in case No. 9 expressed S100, nestin, NGFR, CD133, and desmin. These results suggest that the tumors originated from neural crest cells, such as Schwann cells or striated muscle cells, and some contained possible cancer stem cells because nestin, NGFR, and CD133 are stem cell markers. 2,3,14,17 The ability of neural crest cells to differentiate into a diverse range of cell types explains the expression of Schwannian and striated muscle markers in the neoplastic cells. The cytoplasmic granules in the neoplastic cells were positive for LC3, p62, NBR1, and ubiquitin. LC3 is specifically localized on the membranes of autophagosomes and is a useful marker of autophagy. 15 P62 and NBR1 bind to ubiquitinated proteins, and p62 also acts as an adaptor protein for both ubiquitin and LC3. All of these molecules are required for autophagosome production. 5,9 Moreover, TEM revealed autophagosome-like membranous structures in the cytoplasm of the neoplastic cells. The immunohistochemical and ultrastructural findings of the present study suggest that the cytoplasmic granules found in canine lingual GCT cells are autophagosomes or autophagolysosomes. CD133, a cancer stem cell marker, was detected on the membranes of the cytoplasmic granules in the in vivo tumor cells, cultured cells, and xenotransplanted tumor cells in the present study. Chen et al 1 demonstrated that CD133 plays a role in cell survival by regulating autophagy and glucose uptake in hepatoma cells and that it might also be necessary for cancer stem cell survival in tumor microenvironments. Therefore, canine GCTs might contain cancer stem cells. In addition, the cytoplasmic granules might form during the adaptation of the neoplastic cells to their microenvironments.
The cultured GCT cells lost their characteristic cytoplasmic granules and came to exhibit malignant morphological features. They were immunopositive for neural markers, autophagy markers, and stem cell markers. Melan-A expression was also detected in a few of the cultured cells. These findings suggest that the neoplastic cells dedifferentiated and acquired the characteristics of neural crest stem cells during the culture period and that some cells differentiated into Schwann cells or melanocytes. Furthermore, autophagy was upregulated during the proliferation of the cells in their new microenvironment. Widera et al 19 demonstrated that the multipotency of Schwann cells can be restored by culturing them. The latter finding and the present results support the hypothesis that GCTs originate from neural crest cells.
The xenotransplanted tumors consisted of polygonal to spindle neoplastic cells arranged in interwoven bundles, and pseudopalisadings were observed around necrotic lesions. Some of the neoplastic cells contained fine eosinophilic granules or dark brown melanin pigments in their cytoplasm. In addition, they were positive for autophagy markers or Melan-A. In contrast, the neoplastic cells of the in vivo tumors were positive for autophagy markers and negative for Melan-A. These findings suggest that the neoplastic cells of the in vivo tumors dedifferentiated during the culture period and that some of them redifferentiated into melanocytes after their xenotransplantation. Dupin et al 3 demonstrated that multipotency can be restored to embryonic pigmented cells and Schwann cells by subjecting them to new environmental conditions and that cells derived from the neural crest are phenotypically unstable and possess a broad differentiation plasticity. Previous reports 3,19 support our hypothesis that GCT cells originate from the neural crest, dedifferentiate during culturing, and redifferentiate into melanocytes during xenotransplantation. P62 and ubiquitin were distributed diffusely throughout the xenotransplanted tumors, regardless of the presence or absence of cytoplasmic granules. On the other hand, LC3 and NBR1 were detected only in the cytoplasmic granule-containing neoplastic cells. These findings suggest that many of the xenotransplanted neoplastic cells contained ubiquitinated proteins bound to adopter proteins, but autophagy was upregulated only in the few neoplastic cells that contained cytoplasmic granules. Therefore, the cytoplasmic granules in canine lingual GCTs are formed through autophagy, and the morphology of neoplastic GCT cells is affected by their microenvironment. Recently, it has been suggested that autophagy might play not only a suppressive role in the initiation stages of cancer but also a maintaining role in tumor growth in the later stages of tumorigenesis. 7 According to this hypothesis, autophagy might play a maintaining role in the neoplastic cells of benign GCTs or cultured GCT cells, as well as a suppressive or maintaining role in the neoplastic cells of xenotransplanted tumors.
In conclusion, the present findings suggest that canine lingual GCTs originate from the neural crest and that cytoplasmic granules are formed through autophagy. In addition, the morphology of neoplastic GCT cells is affected by their microenvironment.
Footnotes
Acknowledgment
We thank Sanritsu Zelkova Veterinary Laboratory and Veterinary Medical Center, the University of Tokyo, for their collaboration with the sample collection.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.