
Abstract
The beneficial role that animal shelters play is unquestionable. An estimated 3 to 4 million animals are cared for or placed in homes each year, and most shelters promote public health and support responsible pet ownership. It is, nonetheless, inevitable that shelters are prime examples of anthropogenic biological instability: even well-run shelters often house transient, displaced, and mixed populations of animals. Many of these animals have received minimal to no prior health care, and some have a history of scavenging or predation to survive. Overcrowding and poor shelter conditions further magnify these inherent risks to create individual, intraspecies, and interspecies stress and provide an environment conducive to exposure to numerous potentially collaborative pathogens. All of these factors can contribute to the evolution and emergence of new pathogens or to alterations in virulence of endemic pathogens. While it is not possible to effectively anticipate the timing or the pathogen type in emergence events, their sites of origin are less enigmatic, and pathologists and diagnosticians who work with sheltered animal populations have recognized several such events in the past decade. This article first considers the contribution of the shelter environment to canine and feline disease. This is followed by summaries of recent research on the pathogenesis of common shelter pathogens, as well as research that has led to the discovery of novel or emerging diseases and the methods that are used for their diagnosis and discovery. For the infectious agents that commonly affect sheltered dogs and cats, including canine distemper virus, canine influenza virus, Streptococcus spp, parvoviruses, feline herpesvirus, feline caliciviruses, and feline infectious peritonitis virus, we present familiar as well as newly recognized lesions associated with infection. Preliminary studies on recently discovered viruses like canine circovirus, canine bocavirus, and feline norovirus indicate that these pathogens can cause or contribute to canine and feline disease.
It is likely that Sir Frank McFarlane Burnet, who in 1951 famously predicted the virtual elimination of infectious diseases, never worked with animal shelters. 9 Perhaps pathogens that confront stable host genetics and a stable environment would evolve very slowly, but such situations on this natural earth are increasingly rare, and many intensively housed situations, including the modern animal shelter, are prime examples of biological instability. An estimated 8 to 10 million animals will be housed in animal shelters this year, which is comparable to the total number of dairy cows in the United States (∼9 million) (http://www.ers.usda.gov). Like our managed cowherds, shelters are intensive housing situations where exposure, susceptibility, and transmission of infectious diseases are amplified. According to the American Association of Veterinary Laboratory Diagnosticians (AAVLD), at least 500 state and federally funded pathologists and diagnosticians describe their job in whole or in part as contributing to large animal herd health surveillance in the United States and Canada. For example, food animal–focused diagnostic laboratories alone receive more than $100 million in state and federal funding to perform surveillance for high-impact diseases that would affect the animal agriculture industries (AAVLD, J. Adaska, personal communication, 2012). A complete estimate of the cost of food chain vigilance would also need to include federal (homeland security), state (field personnel and management), farm industry, food industry, and research contributions to this effort. In contrast, very few states support diagnostic investigations or testing of nonprivately owned, small, companion animals like those most often housed in shelters; therefore, there is a paucity of shelter-based infectious disease research. We do know that, predictably, even while animal shelters achieve success at battling overpopulation, problems associated with an unguarded, intensively housed animal population come to light, including pathogen and host transformations that contribute to emergence of disease (Fig. 1).
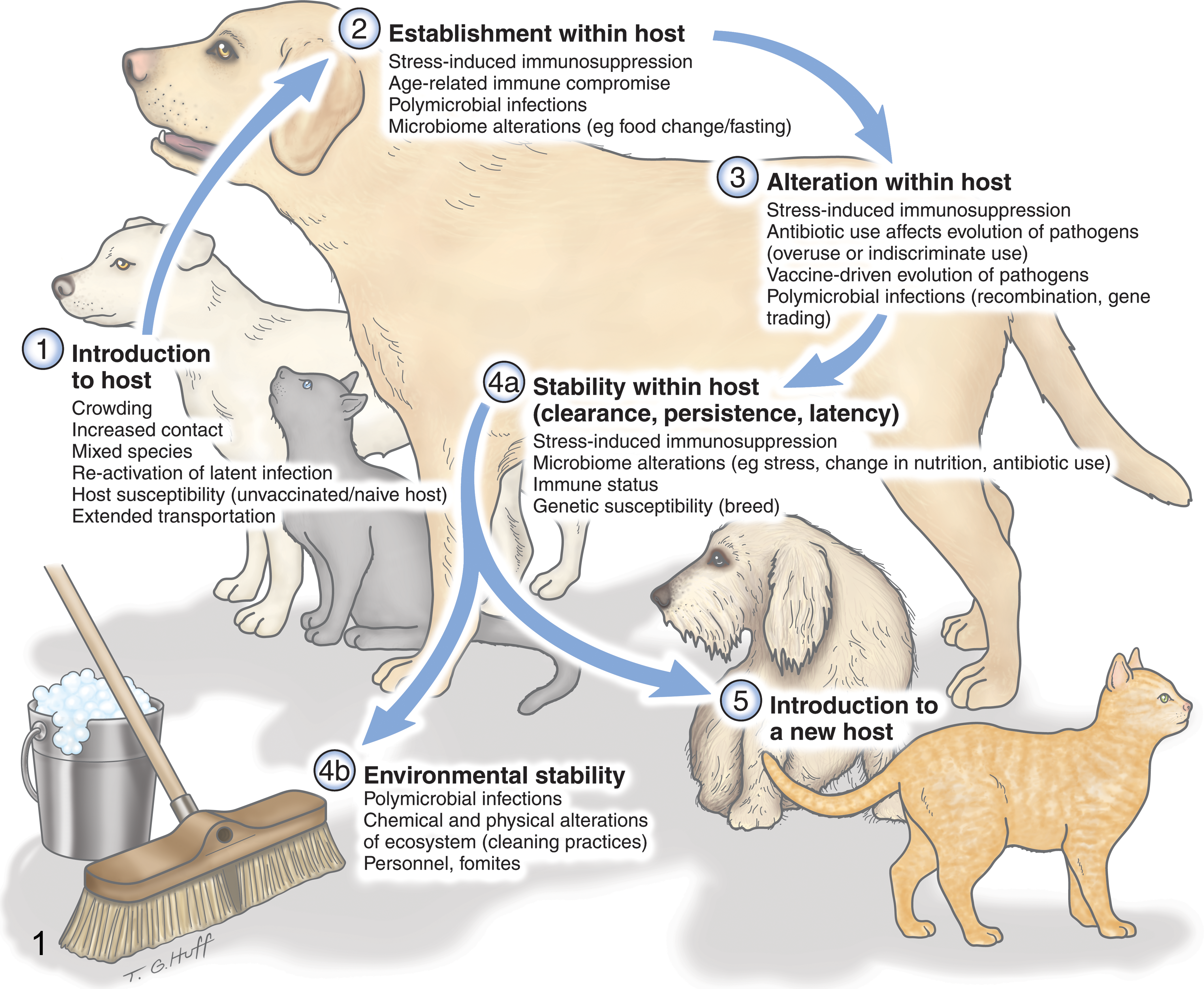
Reasons that an intensively housed environment, like a shelter, can contribute to the emergence of novel pathogens, to novel hosts targeted by a pathogen, or to altered virulence.
In the past 10 years, emergences of fatal infectious diseases such as respiratory, neurologic, and systemic streptococcal infections (dog and cat); canine influenza; hemorrhagic respiratory Escherichia coli (dog) infection; and virulent systemic feline calicivirus infection have all originated in sheltered populations. 7,21,36,37,70 Novel potential pathogens such as canine circoviruses, bocavirus, kobuvirus, and sapovirus, as well as feline noroviruses, have all been discovered in sheltered animals. 42,50,51,76 In addition, shelters have episodic resurgence of feline panleukopenia, canine parvovirus, and distemper. Stressed animals support higher infection and shedding rates of potential pathogens such as feline enteric coronavirus, canine enteric coronavirus (CECoV), feline herpesvirus, and feline calicivirus. 68 In 1 recent study, a population of animals in a single shelter hosted 12 (feline) or 13 (canine) enteropathogens, many of which were zoonotic. 83 Sheltered animals are faced with increased personal health risks, support infectious disease transmission to the community pet population, and have relatively unguarded access to humans. In a herd health situation such as a shelter, there is merit in establishing an etiologic diagnosis both for disease management and for recognition of potential zoonoses. In this review, we first discuss the mechanisms by which an animal shelter might contribute to pathogen emergence and thereafter present information on common endemic diseases of the shelter, with an emphasis on newly recognized pathologic features or newly recognized diseases. The list of diseases is not comprehensive, and because the reports are limited to natural outbreaks, some of the experimental evidence is based on few animals.
Shelter Factors That Contribute to Pathogen Virulence, Emergence, and/or Host Susceptibility
Intensive Housing
Intensive housing or functioning above shelter capacity is likely to increase direct and indirect (fomite) contact, animal stress, the number and complexity of circulating pathogens, and fluctuations in host population size. All of these factors are of primary importance in determining the forces that drive pathogen evolution. The continuous introduction of new (often juvenile) animals, as well as the rapid turnover and animal transport between shelters, often means new sources of disease and a continuously introduced pool of naive individuals. No modern large animal herd management strategy (even finishing operations where new introductions are common) would tolerate introduction of susceptible animals on a daily basis into their herd in the manner of animal shelters.
The relatively unpredictable crowding and transience of a shelter population can support an explosive expansion of viral or bacterial organisms with periodic host movement or other pathogen population bottlenecks, a situation that could theoretically support the selection of pathogen variants. 34 Several highly virulent isolates of endemic shelter pathogens have been described. For example, sporadic albeit rare outbreaks of virulent systemic caliciviruses presumably arise from an otherwise complex population of lower virulence feline caliciviruses. 19 There have also been several documented clonal outbreaks of Streptococcus zooepidemicus (canine and feline pneumonia) and Streptococcus canis (feline sinusitis, meningitis, and cellulitis). 46,71 In 1 study, a clonal shelter isolate of S. canis that caused fatal cellulitis was sustained in the environment of a single shelter, causing multiple outbreaks that spanned over 2 years despite multiple depopulations and aggressive cleaning protocols. 46
Stress
Stress is a virtually inevitable outcome of keeping animals confined in kennel or cattery environments. Barking dogs may be audible in feline housing areas, and small or inappropriately constructed housing may be suboptimal for typical animal behaviors such as normal postures, playing, elimination, and hiding. Kennels are often very loud; sound levels in animal shelters regularly exceed 100 decibels. 18 Diet changes, separation from an attachment figure, and transport can be stressful to dogs upon introduction to a novel kennel environment. 35 ,82
Stress in sheltered animals has been studied both qualitatively, via observable behavioral changes, and through quantitative physiological metrics, such as catecholamine levels, 4,5 heart rate, 5,94 urinary cortisol/creatinine values, 82 and lymphocyte proliferative assays 4 and fecal corticoid metabolites. 26 Many behavioral and physiological measurements of animals housed in dense group environments indicate the presence of stress. Canine fecal corticoid metabolites have been shown to peak 24 hours after admittance to a kennel, followed by a gradual decline. 26
Numerous studies have linked the presence of stress to the induction of immunodeficiency and resulting vulnerability to various infectious diseases. Studies in multiple species indicate that chronic stress has a deleterious effect on both cell-mediated and humoral immunity. 33 Stress-induced immunosuppression puts cats at risk for newly acquired or reactivated infectious disease. 33 Feline herpesvirus (FHV) infection in particular has been shown to be directly reactivated by stress. 2,28 Contamination of the cattery environment is a primary source of FHV transmission, and the 2 main sources of infection are acutely infected cats and latently infected animals experiencing reactivation. 92 Shelter cats with high stress scores have been shown to be more likely to develop an upper respiratory infection, 91 and respiratory disease in dogs has been linked to shelter-stress–associated immunosuppression. 79
Vaccination
Vaccination plays a central role in shelter-associated disease prevention. As a result, most shelters have some form of a vaccination protocol. While the benefits of vaccination far outweigh any potential adverse effects in an individual or group of animals, the data demonstrating that vaccine administration itself can result in immunosuppression are worth consideration. 32 In dogs, polyvalent vaccines have been shown to significantly suppress absolute lymphocyte count as well as lymphocyte response to mitogen stimulation. 74 Modified-live vaccines are designed to induce a low-grade infection, but for some animals that are stressed, immunosuppressed, or concurrently infected with other microbes, modified-live vaccination can result in clinical disease. There are at least 2 notable and published examples of this occurring in shelters. In 1 experimental study, a group of 5 dogs received the modified-live polyvalent vaccine (canine distemper virus [CDV], adenovirus type 2, parainfluenza virus, and Leptospira-canicola-icterohaemorrhagiae bacterin) 3 days prior to oral challenge with canine parvovirus (CPV). Two of these 5 dogs eventually developed canine distemper viral encephalomyelitis. The study authors concluded that an immunomodulating effect of canine parvovirus infection rendered the vaccinated dogs vulnerable to the attenuated (vaccine) strain of CDV. In a private cattery, Salmonella typhimurium was isolated from the tissues of multiple kittens that died after vaccination with a modified-live virus vaccine containing panleukopenia virus, calicivirus, and herpes virus components. 27
Population scale vaccination programs potentially contribute in other ways to pathogen evolution or virulence for pathogens for which vaccination attenuates clinical disease but not infection/shedding. Ohe et al 62 compared the genogroup of feline calicivirus (FCV) isolates from vaccinated cats with those from unvaccinated cats. Significantly more vaccine “breakdown” strains (presence of new or resistant isolates against which the vaccine is ineffective) were isolated from vaccinated relative to unvaccinated cats. These data suggest the emergence of new/resistant strains against which the vaccine is ineffective. In studies of influenza virus infection in pigs, Gauger et al 29 describe that the pathogenic mechanism for vaccine-associated enhanced respiratory disease is antibody mediated. Whether or not vaccination can influence the clinical outcome in subsequent exposures, or whether vaccine exposure could contribute to future outbreak strains, is unknown.
Animals entering shelters are usually vaccinated at or shortly after arrival. Exposure to some circulating pathogens typically occurs within a very short time (hours) after arrival, 68 so the shelter management strategy superimposes vaccination with disease exposure. The choice of vaccines, then, is critical. A closely quartered population of animals, immunosuppressed by stress, vaccination, or infection, could facilitate expansion of variant pathogens that in immunocompetent hosts would be unable to survive.
Antibiotics
The use of 1 or more antibiotics is a common therapy in shelter environments. However, antibiotic therapy can potentially alter treated animals’ enteric, skin, and respiratory microbiota. Documented sequelae to antibiotic treatment of animals include enteric bacterial overgrowth (eg, Clostridium difficile), dysregulated defense against viruses, 38 and the emergence of antibiotic-resistant bacterial strains. In a dog kennel, intense prophylactic administration of multiple antimicrobial drugs at parturition induced multidrug resistance of potentially pathogenic bacteria. 57 Bacteria isolated from rectal swabs of cattery cats were more frequently resistant to multiple antibiotics than rectal bacteria isolated from individually owned cats. 59
These results suggest that antimicrobial drug usage in kennels and catteries can result in the emergence of antibiotic-resistant strains of bacteria. Other potentially deleterious consequences of antibiotic therapy should be considered and balanced against the obvious benefit of antibiotic therapy when the etiology is or may include bacteria. For example, a recent study showed that mice with antibiotic-induced alteration of gastrointestinal (GI) bacterial populations had an impairment of virus-specific cell-mediated and humoral immune responses. The indiscriminate/nonspecific use of antimicrobial drugs may have unexpected deleterious consequences.
Polymicrobial Infections
Diagnostics are straightforward when a clinically recognizable disease entity is caused by a single pathogen. With the increased sensitivity of current molecular diagnostic tests and with the increased use of panels that cover multiple potential etiologies, researchers and clinicians are beginning to untangle the more complicated occurrence of disease when it is caused, or more severe, by the presence of multiple pathogens. These types of diseases are alternately called complex or polymicrobial infections.
Several distinct types of polymicrobial infections are recognized. In so-called primary with secondary infections, the presence of 1 microorganism within the host generates a niche for subsequent microorganisms to colonize. Canine infectious respiratory disease (CIRD) complex (“kennel cough”) is a shelter-based example of such a disease, since infection by 1 of several respiratory viruses (canine distemper virus, canine parainfluenza virus, canine herpesvirus, canine respiratory coronavirus [CRCoV]) often precedes 1 or a number of secondary bacterial invaders. 58 The etiology and pathogenesis of CIRD are complex and case specific. Several novel pathogens that contribute or potentially contribute to CIRD complex have been recognized only recently. 71,77
A second type of polymicrobial infection consists of a primary infection with an immunosuppressive pathogen, such as canine distemper virus or parvoviruses. The immunosuppressive virus creates a biological niche for 1 or more opportunistic infections, which would otherwise be asymptomatic or rarely cause disease.
A third type of polymicrobial infection occurs when 2 or more specific pathogens are concurrently required to initiate disease. Whether this type of symbiosis occurs in shelter diseases is currently speculative, but controlled polymicrobial infection studies in other intensive housing situations have been done. Notably, experiments with intensively raised turkeys infected with turkey coronavirus (TCV) and enteropathogenic E. coli (EPEC) provide some guidance. Either one of these microbes alone, even at remarkably high inoculation titers, are considered nonpathogenic in turkeys. However, when birds were concurrently inoculated with low doses of TCV and EPEC, or in succession over a 7-day period, the birds developed severe disease characterized by high mortality, marked growth depression, and enteric lesions. 64 From the viewpoint of pathogenesis, these are difficult pathogen-host interactions to untangle. Many nonpathogenic GI microbes may all occupy essentially the same mucosal niche, including a multitude of viruses, bacteria, protozoa, and metazoan parasites. 52,61,83 Their presence potentially contributes to a complex interplay with one another and could trigger host humoral and cellular immune responses that might exacerbate tissue injury or accelerate morbidity.
Current diagnostic respiratory and diarrhea panels for dogs and cats are very sensitive and can detect several viruses or bacteria simultaneously, but once the results are available, there is often a question about how to interpret the findings, particularly when there are numerous pathogens detected in a single sample. Clinical and diagnostic investigation is complicated when clinical signs of infection with 1 or a combination of pathogens are overlapping and often nonspecific. Traditional pathologic techniques and immunohistochemistry or in situ analyses can help identify both the lesion and presence of the infectious agent within the lesion, giving increased confidence of causality.
Metagenomics and Pathogen Emergence in the Shelter
Compared with the remarkable growth in human pathogen discovery, pathogenic microbes infecting dogs and cats are undersampled and underestimated. For the reasons enumerated above, resident dogs and cats in shelters are fertile ground for pathogen discovery, and pathologists are uniquely suited to identify lesions that are likely to be infectious and unusual in either their presentation, pattern of inflammation, or tissue distribution. These types of enigmatic cases do not occur frequently, but once known pathogens have been effectively ruled out, they can be quite frustrating. As a result, recent nucleic acid amplification techniques are a new and welcome addition to the diagnostic arsenal.
A staged molecular approach has been used successfully for pathogen detection and discovery in outbreaks of shelter-based disease. 24,53 Discovery typically begins with elimination of known or usual pathogens and is alternately followed by molecular methods such as consensus polymerase chain reaction (PCR), multiplex MassTag PCR platform, or DNA microarray. Feline norovirus, for example, was recently discovered in diarrheic cats based on a consensus PCR strategy using universal primer sets for caliciviruses. 76 A limitation of consensus methods is that they require an estimate or prior knowledge of the suspect pathogen. A metagenomics approach, which involves random nucleic acid amplification followed by high-throughput DNA sequencing (pyrosequencing), can be designed for a less biased amplification. Metagenomics is currently limited to fresh or freshly frozen tissue samples, but several laboratories are exploring ways to extract template from fixed or embedded material. Once a tissue or sample of interest has been identified, some type of particle enrichment is typically done to reduce the otherwise high background of host chromosomal and ribosomal genetic material. Enrichment steps precede amplification and sequencing and vary among laboratories, largely dependent on the character of the tissue sample type and of the targeted pathogen type. Some form of simple filtration is usually done, and in the case of viral discovery, a nuclease treatment is used to digest naked cellular nucleic acids while viral nucleic acids remain protected within the viral capsid. The sequence assembly of overlapping reads is computationally assembled into longer contigs and then subjected to searches against public sequence databases using the Basic Local Alignment Search Tool (BLAST). Sequences of known or closely related viral species are readily identified, but highly divergent sequences may be unrecognizable, so if a novel virus belonged to an uncharacterized viral family, it might be missed by this approach. Viral metagenomics has yielded numerous previously uncharacterized viral genomes from animal samples, including recent discoveries of canine bocavirus, kobuvirus, circovirus, and sapovirus. 42,49 –51
At this point in the discovery pathway, the pathologist has a second important role in determining whether a newly characterized pathogen is causative or a simple contaminant. There are a number of exciting and complementary ways to accomplish this, including comparing prevalence in a set of disease cases (eg, animals in an outbreak population) with healthy controls using PCR, detection of virus-specific antibodies, localization of the pathogen within the affected tissue (in situ hybridization or immunohistochemistry), and/or animal infection studies. In veterinary medicine, deep sequencing is currently used for pathogen discovery, but using this technique for screening of clinical samples is conceivable. As detection methods become even more sensitive, our ability to sort contaminants and orphan pathogens from significant microbes will become increasingly challenging. Ultimately, convincingly and rigorously demonstrating disease association requires confirmation by multiple techniques.
Common Endemic Diseases of the Shelter, With an Emphasis on Newly Recognized Pathologic Features and Newly Recognized Diseases
Feline Herpesvirus 1
In the United States, FHV-1 is the single most important pathogen contributing to feline upper respiratory infection (URI). 10,93 Active shedding of FHV-1 from latently infected cats can be induced by environmental stressors or experimentally by administration of corticosteroid. 28 Virtually all cats infected with FHV become lifelong carriers. Studies have shown that approximately 4% of cats entering a shelter shed FHV, while approximately 50% of cats reactivate a latent infection and begin shedding the virus within a week. 92 Disease recrudescence can result in conjunctivitis, ulcerative keratitis, rhinosinusitis, and/or interstitial pneumonia. Chronic recurrent rhinosinusitis, often considered incurable, may result from bony remodeling of the nasal turbinates as a sequela of acute herpesviral rhinitis. 10 Although FHV-associated mortality is low, clinical signs of rhinosinusitis are discerning criteria in many shelters for euthanasia, so the consequences for affected cats can be profound. 2
While infections with several pathogens are capable of causing URI in cats, FHV-1 is a major player in recent studies performed in the United States. One study was designed to correlate diagnostic real-time quantitative PCR (qPCR) results for FHV-1, FCV, Mycoplasma felis, Chlamydophila felis, and Bordetella bronchiseptica with histologic lesions in cats with clinical signs of URI. 10 Over the course of a year, 22 cats with URI were examined at necropsy. Twenty were qPCR positive for FHV-1, 7 for M. felis, 5 for FCV, 1 for C. felis, and 0 for B. bronchiseptica. In this study, histologic (inclusion bodies) or antigenic detection of FHV-1 was seen in 18 of 20 cats positive for FHV-1 by qPCR.
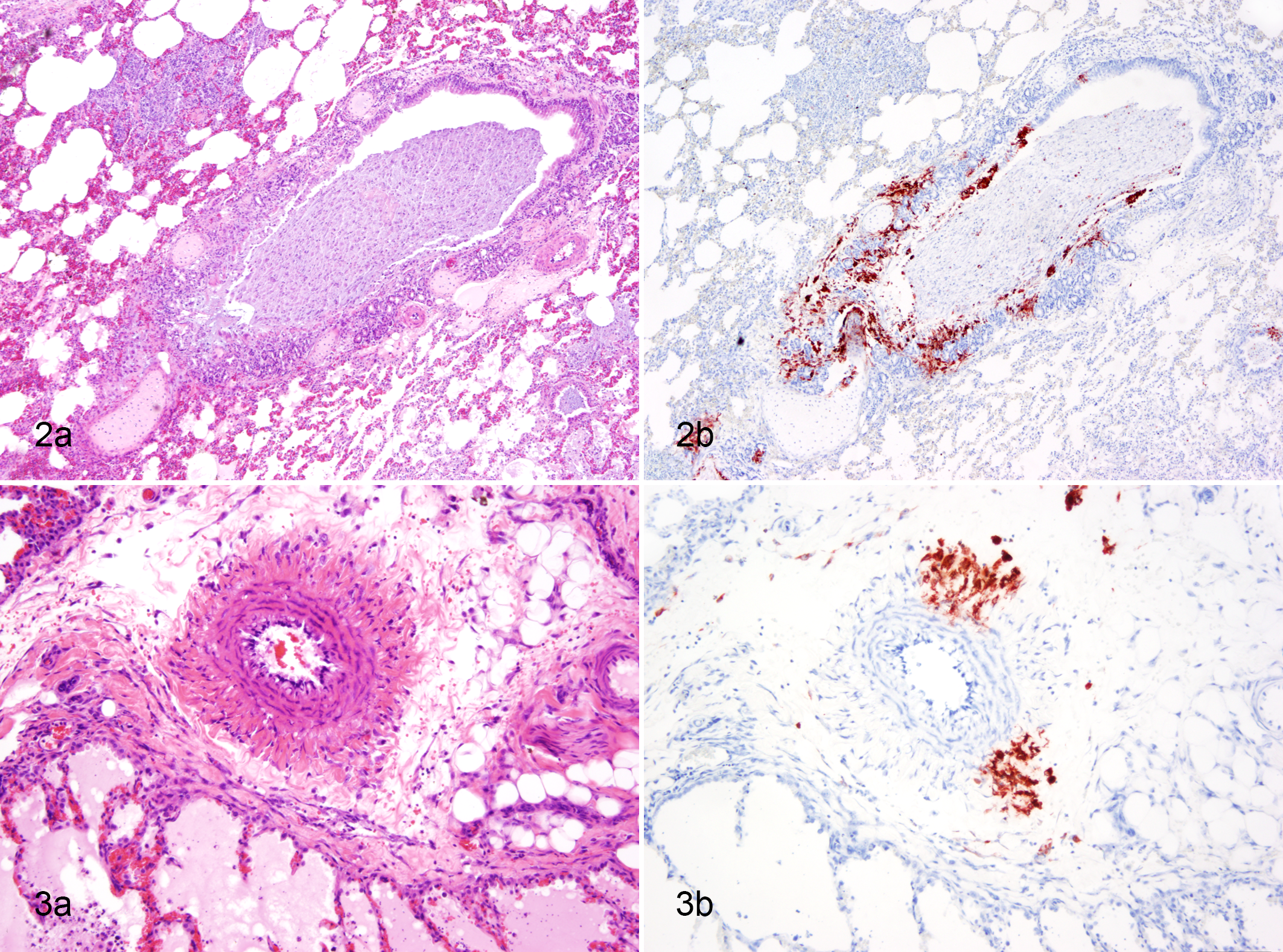
In most shelter FHV infections, virus distribution is limited to the nasal cavity and frontal sinuses. However, other distinct lesions are attributed to infections with FHV. Individual cases of fatal necrotizing tracheitis/bronchitis have been described (Fig. 2A,B). 54 Outbreaks in shelter cats with lesions limited to the trachea and bronchi have been observed on 3 other occasions, one of which resulted in temporary closure of the shelter in Spain (Prada-Puentes, personal communication, 2010). Along with the remarkably bronchocentric lesions, there was a segmental necrotizing arteritis of the large parabronchial pulmonary arteries in the lungs of affected cats (Fig. 3A; P. A. Pesavento, unpublished observation). Along with copious virus in tracheal and bronchial respiratory epithelium, viral antigen was present within cells, presumably macrophages, of the arterial tunica adventitia and media (Fig. 3B). Rarely, FHV-1 has been reported to be a systemic disease, causing vasculitis and hemorrhage in multiple organs (Fig. 7). 36,87
Canine Distemper Virus
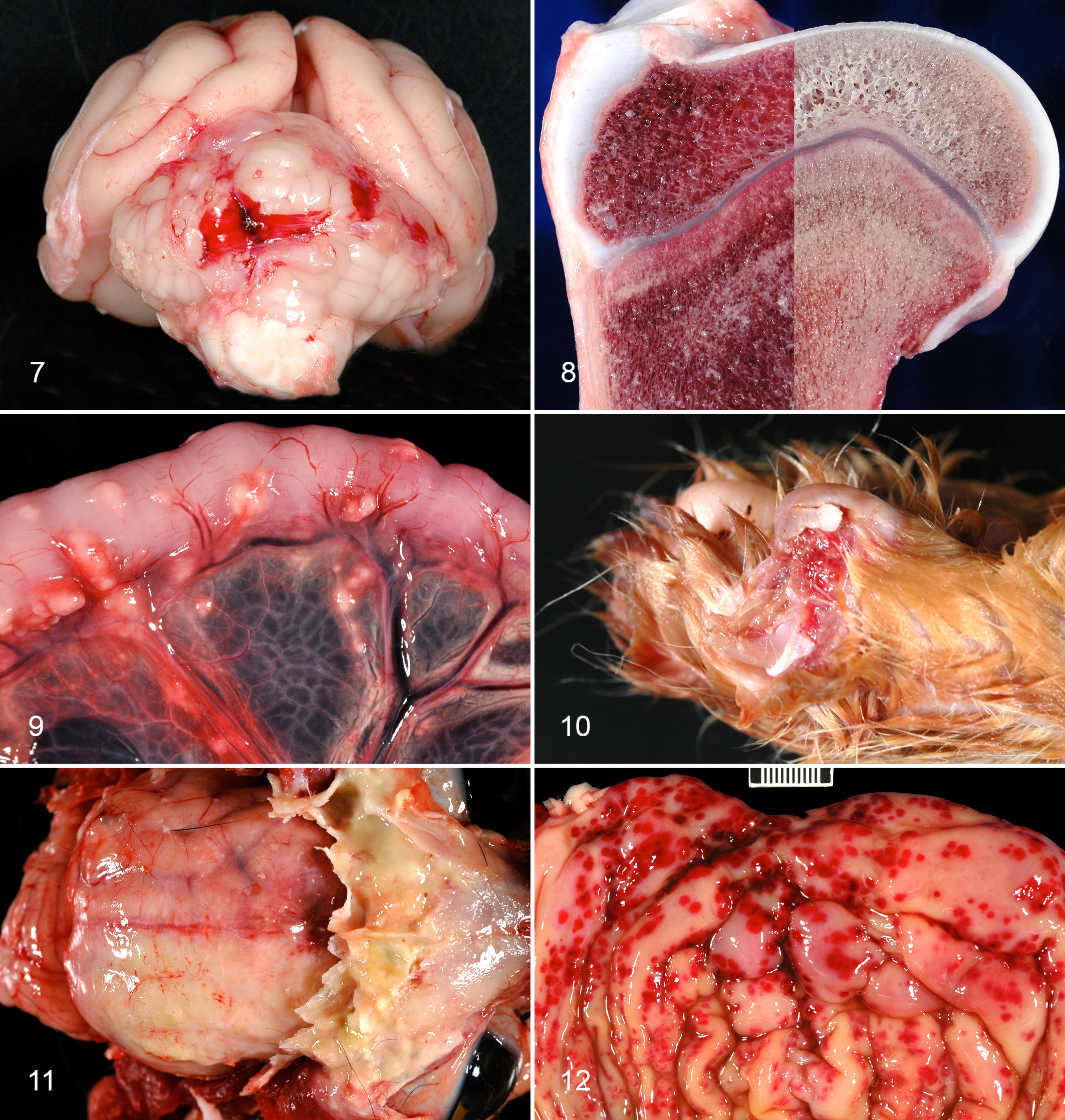
CDV, a morbillivirus in the family Paramyxoviridae, has a long history of kennel-associated disease outbreaks. CDV causes a systemic infection in dogs and wild carnivores, with disease manifestation in multiple organs, including the respiratory tract (rhinitis, tracheobronchitis, interstitial pneumonia), skin (hyperkeratosis of the nose and foot pads), alimentary tract (catarrhal enteritis), central nervous system (necrotizing encephalitis), bones (growth retardation lattices), and eyes (conjunctivitis). CDV-associated growth retardation lattice (Fig. 8) is a metaphyseal sclerosis parallel to the physis resulting from pathologically cross-linked persistent primary spongiosa. This radiographically apparent lesion represents a transient impairment of osteoclastic resorption. In kennel environments, CDV spread is facilitated by a constant supply of infection-susceptible puppies. 32 In surviving puppies, infections have been associated with immunodeficiency, making concurrent or subsequent infections with other viruses such as parvovirus more severe. 32
Recently in Switzerland, a virulent CDV strain with enhanced neuronal tropism, high morbidity and mortality, and rapid spread was identified in numerous wild carnivore species and a domestic dog. 63 The CDV isolate demonstrated a unique molecular signature. Notably, the infected dog had undergone a standard anti-CDV vaccination protocol yet presented with CDV-associated central blindness and circling; infection was confirmed by consistent histologic lesions in brain, positive immunohistochemistry, and reverse transcription (RT)–PCR assays. 63 The authors speculated that the dog likely acquired the CDV infection from contact with wild carnivores, based on similarity in nucleotide and amino acid sequences. Mature dogs that do not receive periodic immunizations may lose their protection and become infected after stress, immunosuppression, or contact with infected and shedding individual animals. 32 In a kennel environment, all of these circumstances may be concurrently in play.
Canine Influenza
In 2004, an H3N8 canine influenza virus (CIV) closely related to equine influenza virus was isolated from racing Greyhound dogs with severe respiratory disease. 21 CIV was subsequently associated with multiple outbreaks in shelter dogs and circulated in shelters and racing Greyhounds throughout the United States. 14,21,25,65 These findings came as a surprise to many, since at the time, dogs were thought to be refractory to infection with influenza viruses. 30 CIV infection in dogs is rarely fatal unless complicated by concurrent bacterial pneumonia, 14 and it is notable that the most important differential diagnosis for hemorrhagic pneumonia in dogs is S. zooepidemicus (see below). To complicate matters, all dogs diagnosed with hemorrhagic pneumonia and CIV in 1 outbreak involving racing Greyhounds had concurrent infections with S. zooepidemicus. 98
The interspecies transmission of avian influenza virus H3N2 to dogs was first reported in South Korea during 2007. 88 Infection of dogs with H3N2 CIV resulted in a severe respiratory syndrome characterized by high fever, coughing, severe bronchointerstitial pneumonia, and necrosuppurative tracheobronchitis. 41,88 An outbreak of H3N2 canine influenza virus in cats has recently been reported in an animal shelter in South Korea. 40
Recent evidence indicates H3N8 CIV infection in dogs, including shelter dogs, is uncommon. In a survey of 1268 shelter and pet dogs, H3N8 CIV seroprevalence peaked in 2007 at 62%, then decreased in 2008 (38%) and 2009 (15%). 1 Seroprevalence was associated with geographic region and canine population density (ie, shelter or boarding facilities). An H3N8 CIV seroprevalence rate of 0% was identified in unvaccinated dogs (n = 93) participating in a Flyball tournament in Pennsylvania, 95 and a survey of 225 dogs in Ontario, Canada, identified a single Greyhound that was seropositive. Since this dog was originally from Florida, the authors concluded that CIV was not locally present. 47 In a recent survey, no CIV-seropositive dogs were identified in New Zealand (n = 251) 45 or Italy (n = 637). 75 Therefore, recent seroprevalence evidence suggests that H3N8 CIV seroprevalence is uncommon, while H3N2 CIV infections in dogs and cats may be emerging diseases. A more detailed account of canine influenza infection is described elsewhere in this issue (Priestnall et al 80 ).
Feline Calicivirus
At any given time in a shelter environment, a wide range of cats (3%–100%) shed FCV. 20,81 Infected and shedding cats are often asymptomatic or demonstrate mild upper respiratory signs with or without lingual ulcers. 81 FCV infection in shelter cats is so common that vaccination with a modified-live (attenuated) vaccine is widely practiced, and diagnosis of true FCV-associated disease is problematic. Widespread vaccination generally prevents clinical expression of FCV infection but does not prevent infection or shedding. 81
Aside from its contribution to the morbidity associated with feline URI, of particular concern in the shelter are rare but more pathogenic forms of caliciviruses, which have a remarkable ability to emerge from an otherwise docile background. 20,37,84,85 While many factors, including host age and dose, can contribute to disease outcome, the fact that viral factors are important has been demonstrated by experimental infections using purified viral isolates that predictably cause mild and systemic disease. Twelve highly fatal (hypervirulent) outbreaks of FCV-associated disease have been confirmed by the University of California, Davis shelter animal medicine service since hypervirulent isolates were first recognized in 2000. 66 The viral isolates are genetically distinct and arose independently. Other reports from the United States and Europe concur that hypervirulent FCV occurs rarely and sporadically. 20,37,84
Infection with hypervirulent FCV can result in 1 or more of a diverse spectrum of clinical manifestations, including lameness, diarrhea, pneumonia, edema, and hemorrhage. Confirmation of causation is arduous; since FCV is ubiquitous and there is no known genetic signature that distinguishes hypervirulent isolates, it requires some combination of virus isolation from multiple affected tissues, along with histopathology and immunohistochemistry that link the presence of the virus with the systemic disease. 72 In cats clinically affected with low-virulence field strains of FCV, the virus is generally limited to oral mucosa or respiratory epithelial cells. In cats naturally infected with a hypervirulent FCV, viral antigen can be demonstrated systemically within endothelial cells as well as epithelial cells in multiple tissue types. Systemic forms are acute and can include 1 or a combination of lesions, including ulcerative dermatitis, arthritis, vasculitis, hepatitis, pancreatitis, and/or pneumonia. Mortality rates in virulent FCV epizootics are often high, ranging from 33% to 60%. FCV-vaccinated adult cats are often affected, along with kittens. 72 The largest documented outbreak affected 54 cats over a 20-day period, but most outbreaks are more limited in the number of affected animals (2–12 cats) and in duration. 37
Coyne et al 19 have recently published a large-scale analysis of the genetic diversity of FCV. This study showed that in both the client-owned cat population and shelters with good biosecurity, multiple strains of FCV continuously circulate, and FCV is in perpetual circulation in cattery environments.
Feline and Canine Parvoviruses
Beginning in the late 1970s, the newly emergent canine parvovirus strain CPV2 caused multiple fatal epizootics of hemorrhagic gastroenteritis in kennels and shelters worldwide. Evidence links the emergent CPV-2 with feline panleukopenia virus (FPV), 13 although viral emergence from a closely related host cannot be absolutely excluded. This remarkable feline-canine species jump was apparently accomplished with only a 6– or 7–amino acid substitution between FPV and CPV-2, mostly in the transferrin receptor binding domain of the capsid VP2 protein. 86 Since that time, a progressive series of antigenic variants based on single amino acid substitutions in VP2 (most recently CPV-2a/426Glu) have emerged and are now distributed worldwide. In dogs, etiologic diagnosis through fecal enzyme-linked immunosorbent assay (ELISA) is relatively straightforward, although false-negative results are relatively common (lack of viral shedding at the time of sampling, viral antigenic drift). The current vaccine series is generally effective regardless of the CPV strain type, but some unique biological behaviors of CPV are troubling with regard to its current and potential threat in shelters. CPV has been shown to evolve very rapidly in comparison to FPV 3,23 ; while it is controversial whether CPV2c is more virulent in dogs, 12,22 it does have an extended host range. Unlike its predecessor CPV2, CPV2c can infect and cause disease in cats. Given the remarkably high concentration of CPV shed in feces (109 PFU/g), 56 the extreme environmental stability of parvovirus, and the diverse range of carnivore host (skunks, raccoons, foxes), CPV is likely to persist as a perennial problem in shelter environments.
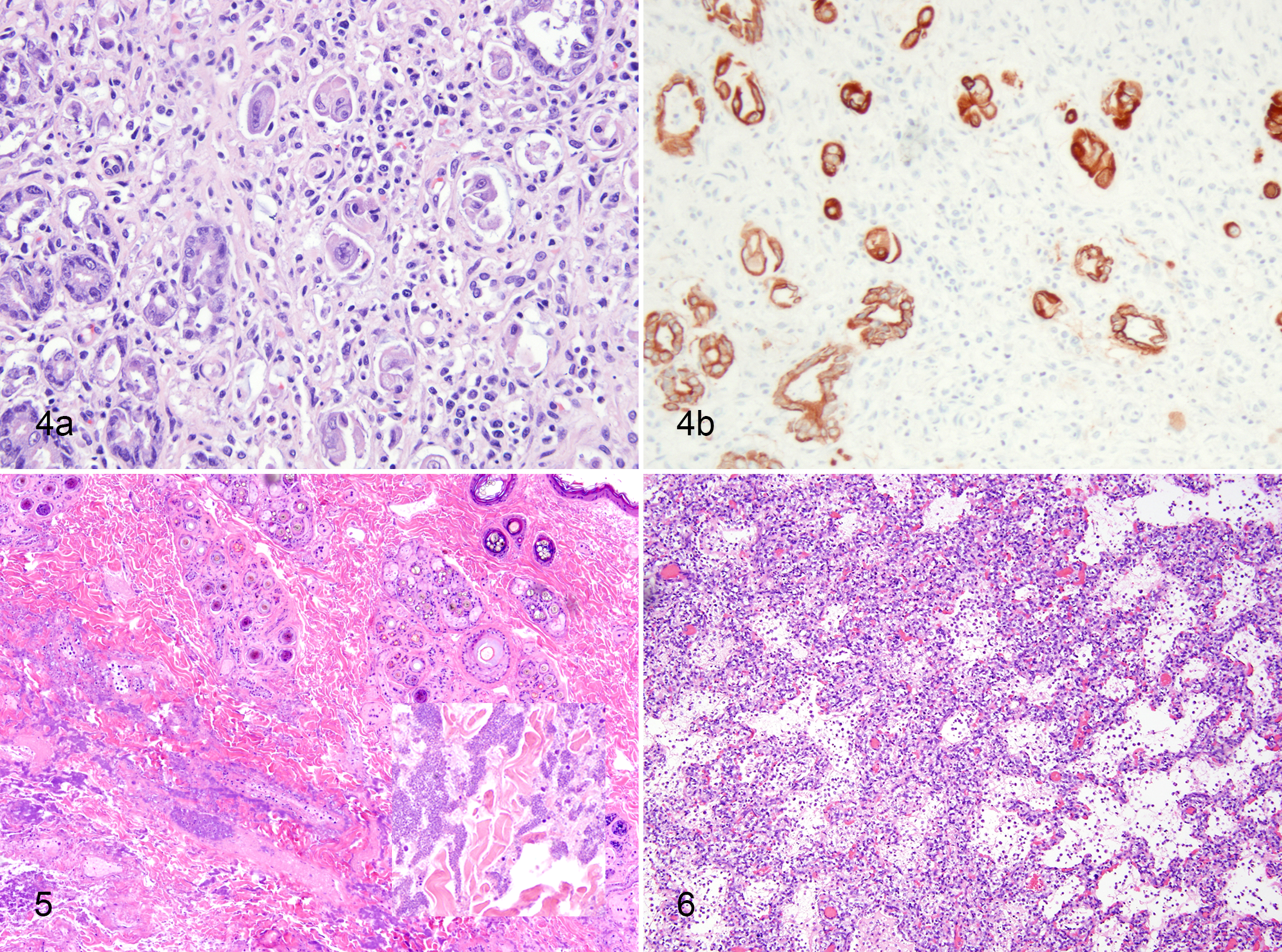
Regardless of any alterations in biological behavior, the gross and histologic presentation and pathogenesis of canine or feline parvoviruses in dogs and cats are consistent. Both viruses cause acute disease, with viral infection and replication in a broad spectrum of mitotically active cells. In neonatal animals, tissue tropism includes the myocardium, although we have seen at least 3 cases of parvovirus-associated myocarditis in older puppies (ages 3–5 months) over the past few years. In most cases in puppies and older dogs, the virus replicates in lymphoid tissues and rapidly replicating crypt cells of the ileum and small intestine. Potentially fatal hemorrhagic enteritis is the main gross lesion observed. In canids, this is easily recognizable on gross examination as segmental hemorrhagic enteritis. We have found that fatal cases of feline panleukopenia can be much more subtle, with diffuse dilation of the small intestines, but sometimes no grossly observable hemorrhage. Both cats and dogs, however, have very similar histologic lesions. During acute infection, there is crypt-oriented epithelial necrosis. Because the lesion is segmental, multiple sections of the affected small intestine should be taken, as epithelium quickly sloughs into the crypt spaces (crypt “abscesses”). While crypt material is typically positive by immunohistochemistry, it is unnerving to make a diagnosis based solely on cellular debris. In late or healing stages of infection, the regenerating epithelium can be striking with hyperplastic glands nested in an inflamed and expanded lamina propria (Fig. 4a). At this stage, neither the bizarre epithelial cells (Fig. 4b) nor adjacent glands will typically contain virus, but individual macrophages within the gut-associated lymphoid tissue are sometimes positive by immunohistochemistry.
Feline Infectious Peritonitis
Despite substantial long-term research efforts, there is currently no effective diagnostic protocol that can reliably discriminate the relatively avirulent feline enteric coronavirus (FECV) from the pathogenic feline infectious peritonitis (FIP) virus. However, ongoing efforts in multiple laboratories have identified, cataloged, and defined multiple genetic determinants that distinguish FECV from FIP isolates of feline coronavirus (FCoV). 8,16,17,67,90 Any one of several mutations is thought to alter viral tropism to specific subsets of monocytes and macrophages, facilitating systemic colonization and the emergence of morbidity.
For FCoV infection of cats, the complex interplay of virus and host strongly suggests that both viral and host factors contribute to the emergence of disease. 60 FIP remains a disease primarily of young cats with significant breed predilections, with increased prevalence in British Shorthair, Devon Rex, Birman, Burmese, and Abyssinian cats. 73,97 Some breeders report that 100% of the kittens die within 1.5 years (P. A. Pesavento, unpublished observation). Regardless of whether the affected animal is of mixed or pure breed, the gross and histologic findings are unchanged from what Wolfe and Griesemer 96 described in 1966: “severe peritonitis characterized by excessive abdominal fluid and the deposition of granular gray-white exudate on the abdominal viscera. Sharply defined serosal elevations, 0.5 to 2.0 mm in diameter, on the omentum, mesentery, and viscera were caused by plaques of exudate and severe subjacent inflammation” (Figure 9). The reported prevalence of FIP is consistently higher in animals from a shelter environment. Whether this is an indication of a higher exposure to the enteric form of FCoV, host compromise, or the presence of more virulent isolates is unknown. A more thorough discussion of the pathogenesis and histologic characteristics of feline coronaviruses occurs in the article by Kipar et al 44 in this issue.
β-Hemolytic Streptococci
Many in the medical community believe that emerging viruses pose a greater threat than bacteria because of the perception that immediate and effective antibiotic therapy for most bacterial diseases is currently available. In the modern world, this viewpoint is likely to prove naive, since the current impact of Yersinia pestis, clostridia, mycobacteria, and resistant forms of Streptococcus (MRSA) and Staphylococcus spp underscore the remarkable evolutionary plasticity, tenacity, and antibiotic resistance of bacterial pathogens. In shelter environments, the β-hemolytic S. canis and S. zooepidemicus are the most common streptococcal pathogens cultured in small animals, and they have been associated with disease outbreaks with high morbidity and mortality in both cats and dogs.
S. canis is isolated from the nasal cavity of up to 10% of cats with chronic URIs. Disease associated with this pathogen can include polyarthritis, urogenital infections, and neonatal septicemia. 48 Most previous reports reflect transient outbreaks of disease affecting young animals in closed colonies. 31,39 However, within the past 4 years, there have been multiple reports of shelter outbreaks affecting hundreds of intensively housed cats. Infection has been frequently associated with either necrotizing fasciitis (toxic shock syndrome) (Figs. 5, 10) or sinusitis and meningitis (Fig. 11). 69,89 There are several disturbing features of these outbreaks. First, while the cultured bacteria were susceptible to multiple antibiotics in vitro, in vivo treatment was not always successful. Second, despite extensive environmental cleaning (including cage removal and replacement), the same clone of bacteria persisted in the environment of at least 1 shelter for more than 18 months, with new and fatal cases occurring within 2 days of returning cats to the shelter. Third, cats of all ages and of all vaccination status were affected. Fourth, S. canis was the sole pathogen identified in most cases; other pathogens were variably but inconsistently present. 46,69 We have very little understanding of the differences between ubiquitous and virulent strains, the importance of clonality, or the triggers for outbreaks. The remarkable capacity for episodic virulence and the adaptability of the streptococci are evident in many species.
S. equi subsp zooepidemicus can be found infrequently in the oral cavity of asymptomatic dogs, but its presence is significantly associated with CIRD. 15 Until recently, S. zooepidemicus was not widely recognized as a primary canine pathogen, but outbreaks are increasingly frequent or recognized with increasing frequency. 11,43,71,77,78 All reports describe acute, fatal hemorrhagic pneumonia that in 1 case was observed in more than 1000 mixed-breed dogs in a single shelter. 71 Gross lesions included hemothorax and acute, fibrinosuppurative pneumonia with intralesional Gram-positive cocci (Fig. 6). While diagnostic tests in individual cases revealed inconsistent but potentially contributory co-pathogens, the evidence from clinical, pathological, and molecular clonality analyses indicates that S. zooepidemicus was strongly associated with acutely fatal respiratory infection in dogs. There are individual reports of S. zooepidemicus causing rhinitis 7,55 and pneumonia 6 in cats. The pathogenesis of the streptococcal infections is discussed in depth elsewhere in this issue (Priestnall et al 80 ).
Canine Circovirus
Among recent discoveries of viruses in sheltered animals, steps in demonstrating associated disease are most persuasive in the case of canine circovirus. 42,49 Canine circovirus has been present in a growing number of cases of dogs with vasculitis, histiocytic inflammation, or a combination thereof (Fig. 12). 49 In independent studies, 3% to 6% of normal dogs and up to 11% of dogs with thrombocytopenia were PCR positive for canine circovirus in peripheral blood. 42,49 In pilot studies to establish whether canine circovirus contributes to canine disease, in situ hybridization was used to establish the presence of the virus in tissues of dogs with vasculitis and/or histiocytic inflammation. 49 As has been shown for porcine circovirus 2, virus is most abundantly present in primary and secondary lymphoid tissues in affected dogs. 49 Studies that are ongoing in a number of laboratories are designed to examine the prevalence and pathogenesis of canine circovirus infection.
Conclusion
While it is not possible to anticipate the time of pathogen emergence or the next species-jumping event, their place of origin is, in our experience, nearly predictable. Shelter management strategies and emergent factors such as intensive housing, stress, immunosuppression, modified live vaccination, and antimicrobial administration culminate in a biological “perfect storm,” contributing to the emergence and possible persistence of virulent infectious pathogens. It follows that we see common pathogens causing more severe disease and newly recognized emerging pathogens becoming manifest in the shelter environment. Early recognition of emerging pathogens has the immediate benefit of improving disease control and identifying potential zoonoses. There is also great value in characterizing circulating pathogens, since crucial reference genomes will open avenues of research into the population dynamic of pathogens and the evolution of virulence.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.