
Abstract
A retrospective study of the pathologic findings in weedy (Phyllopteryx taeniolatus) and leafy (Phycodurus eques) seadragons was performed on specimens submitted to 2 reference laboratories from 1994 to 2012 to determine the range and occurrence of diseases affecting aquarium-held populations. One hundred two and 94 total diagnoses were recorded in weedy and leafy seadragons, respectively. Two of the more common etiologic diagnoses in both species were mycobacteriosis and scuticociliatosis, whereas myxozoanosis was common in weedy seadragons. Metazoan parasite infections were less common etiologic diagnoses. There were no correlations between mycobacteriosis and ciliate protozoan infections in either species. Myxozoanosis was usually found in combination with other diseases and, except for 1 case, was restricted to weedy seadragons. Phaeohyphomycosis, nonmycobacterial bacterial infections, and trauma were also important but less frequent diagnoses. Intestinal coccidiosis was found in weedy but not leafy seadragons. Mineralization of the swim bladder was detected in 26 of 197 leafy seadragons and only 2 of 257 weedy seadragons. Although weedy and leafy seadragons share certain diseases of significance to exhibit populations, there are diseases unique to each species about which the veterinary pathologist, clinician, or diagnostician should be aware.
The Syngnathidae include the weedy seadragon Phyllopteryx taeniolatus (WSD) and leafy seadragon Phycodurus eques (LSD), the seahorses, and the pipefishes. 6 This report summarizes the pathologic findings in 257 WSD and 197 LSD submitted to 2 different reference laboratories from 1994 to early 2012.
Materials and Methods
Seadragons of both species from the archival collections of Northwest ZooPath (NZP) and the Connecticut Veterinary Medical Diagnostic Laboratory (CVMDL) were examined. All seadragons submitted by client institutions using the diagnostic pathology services of the 2 pathology groups from 1994 to 2012 for NZP and from 1999 through 2011 for the CVMDL were included in the study. Unprosected specimens or bony tissues of prosected specimens were decalcified by immersion in 5% formic acid or 0.1% ethylenediaminetetraacetate (EDTA) (Rapid-Cal; Biesecker Bio Chemical, Mt Vernon, WA) or a 0.5 M EDTA solution made in house from component reagents (Fisher Scientific, Pittsburgh, PA) for a duration necessary to allow slicing of the specimen without artifactitious fracture of dermal or skeletal bone, typically 24 to 48 hours. Following the aforementioned specimen preparation, all tissues were processed routinely, embedded in paraffin, sectioned at 4 or 5 μm, mounted on frosted slides, and stained with hematoxylin and eosin (HE). Histologic sections taken from paraffinized tissues that had been decalcified were mounted on glass slides coated with 3-aminopropyltriethoxysilane. Histologic sections from select tissues were stained with Ziehl-Neelson or Fite’s acid-fast stains, Brown & Brenn tissue Gram stain, Gomori or Grocott’s methenamine silver stains, or Fontana-Masson stain. Histologic sections were examined by light microscopy, and lesions were described and reported.
Pathologic findings were given numerical codes based on organ system involved and morphologic and etiologic diagnosis. Clinical histories provided by the submitting institution as well as gross necropsy findings were considered in completing the final anatomic diagnosis. Numerical codes and animal identities were entered into a searchable database (Zairtec Systems, Brecksville, OH). Searches were then performed on organ system, morphologic diagnosis, and, when available, etiologic agent. Up to 4 variables could be searched at one time to look for correlates between disease states. The few discrepancies based on wording of diagnoses were resolved by examining relevant anatomic descriptions from cases and using the most commonly accepted terminology for the case in question.
Results
The 15 etiologic diagnoses affecting more than 1% of submissions are summarized in Table 1. The most important lesions are described as follows.
Summary of 15 Etiologic Diagnoses Affecting Greater Than 1% of Weedy and Leafy Seadragons Submitted to 2 Reference Laboratories From 1994 to 2012.
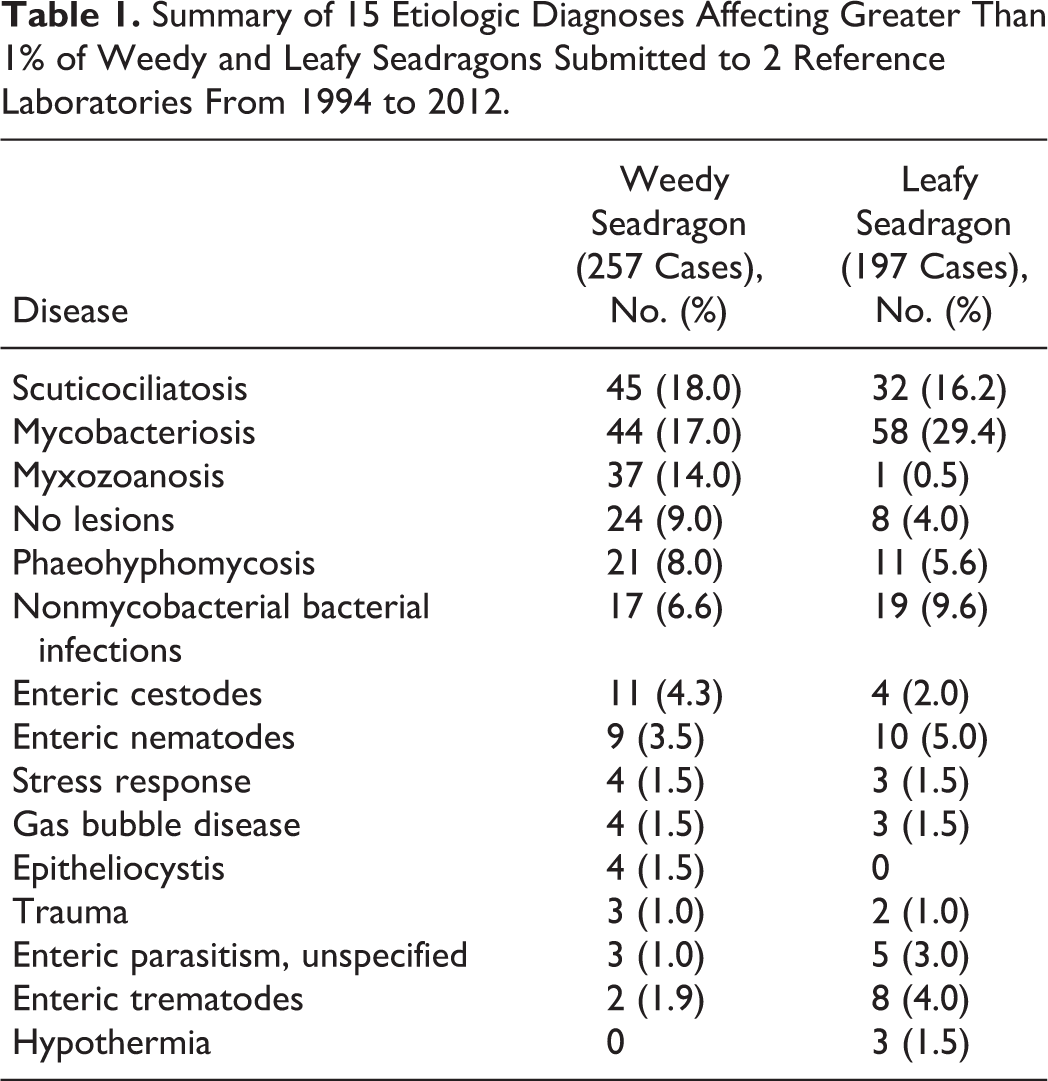
Scuticociliatosis
In most fatal cases of scuticociliatosis, the skin was the primary site of infection, with invasion of underlying tissues (eg, skeletal muscle and connective tissue). Sites of infection were characterized by extensive areas of acute to subacute necrosis with numerous ciliate protozoans, 20 to 30 μm in diameter, some with eosinophilic round cytoplasmic bodies. This finding was associated with epidermal ulceration and dermal edema as well as a mild infiltrate of histiocytes, frequently accompanied by rhabdomyolysis in the subjacent musculature. Scuticociliates were also found within vascular spaces and interstitium of gill lamellae, throughout the serosal surfaces of the coelomic cavity, and within the lumina of lymphatic channels in various tissues, especially the skin and connective tissues. Scuticociliates were also present in the blood vessels of internal organs, where they were associated with tissue necrosis and moderate infiltrates of macrophages and granulocytes (Figs. 1–4).
Leafy seadragon; skin. Visible foci of pink discoloration over head, neck, and trunk caused by ciliated protozoan infection.
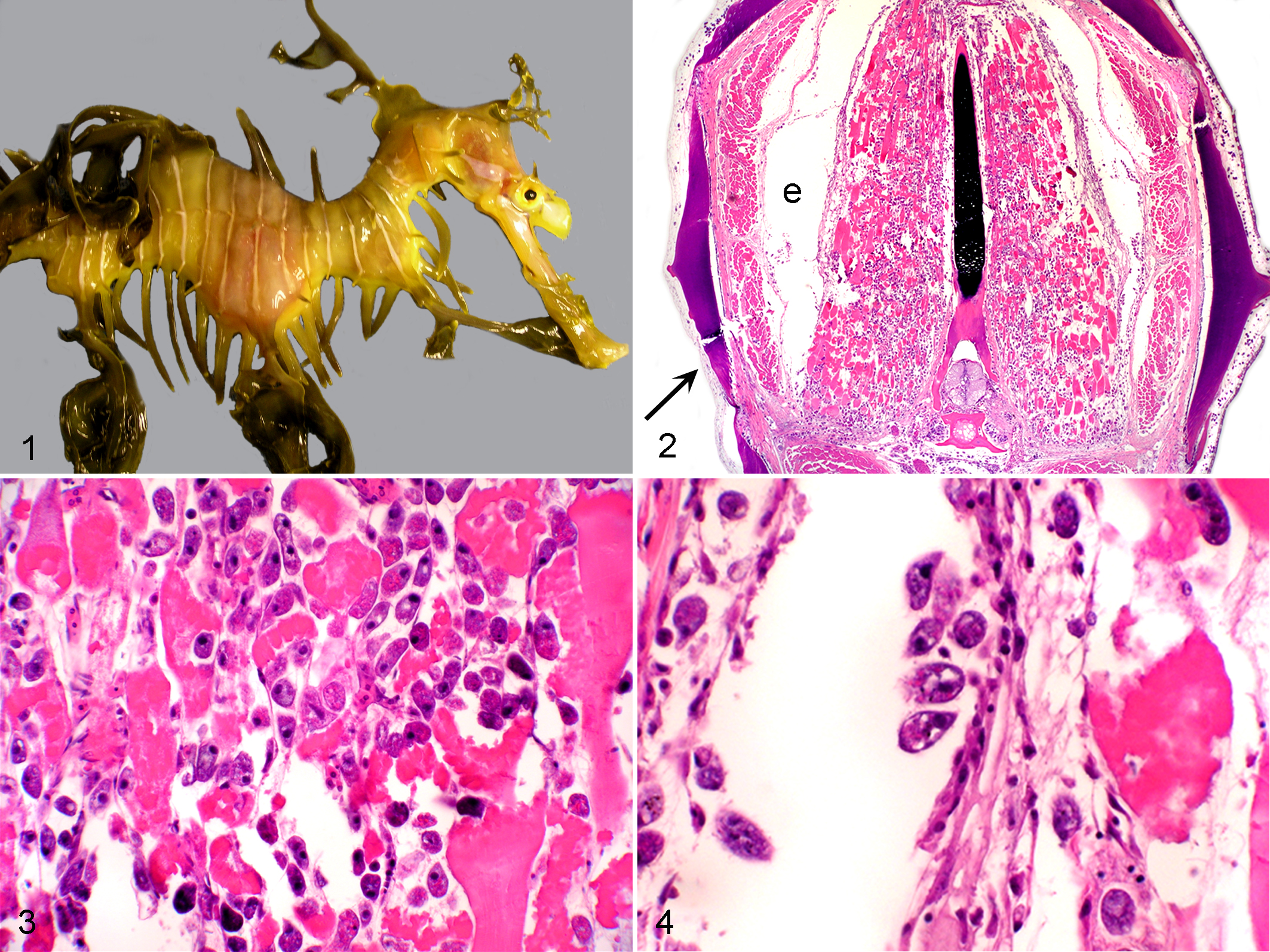
Mycobacteriosis
Lesions were characterized by multiple and occasionally extensive foci of histiocytic inflammation and occasional granuloma formation. Commonly, the clinical history included observation of a chronic skin lesion that appeared both erosive and proliferative, often secondary to injury. This was especially true of injuries to the tip of the snout. The most commonly affected organs in both species were the liver, kidney, heart, gill, and skin, although virtually any organ could be, and was, affected. Organs to which mycobacteria spread contained foci of bacterial embolization and early granulomatous inflammation, a finding that was particularly evident in the gill. Multinucleate giant cells were rarely, if ever, encountered in foci of inflammation, and acid-fast stains (eg, Ziehl-Neelsen and Fite’s) revealed macrophages and granulocytes to be replete with acid-fast bacilli. Rhabdomyolysis in muscle underlying dermal lesions was common, as was severe atrophy of fat, consistent with emaciation (Figs. 5 –8).
Weedy seadragon; swim bladder. The thickened opaque appearance to the bladder wall is secondary to granulomatous inflammation associated with mycobacteriosis.
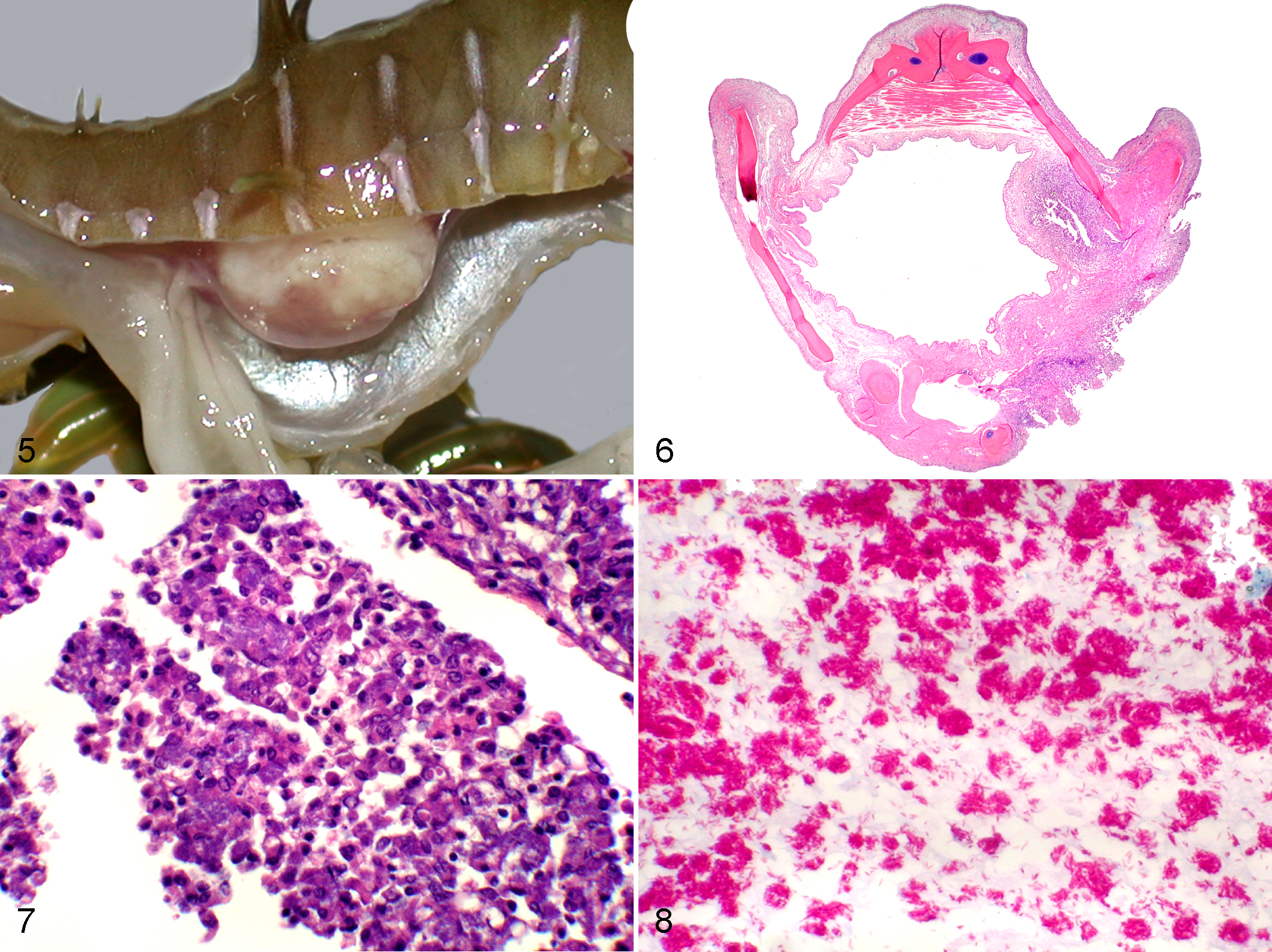
Phaeohyphomycosis
Infections of skin, gill, and internal organs, by melanized fungi, previously have been reported. 19 The lesions consisted of areas of vascular and fibrinoid parenchymal necrosis and granulocytic and histiocytic inflammation with numerous thin-walled, septate fungal hyphae that stain brown in routine HE sections (Fig. 9). The presence of melanin in the walls of these fungal hyphae was demonstrated histochemically using the Fontana-Masson stain.
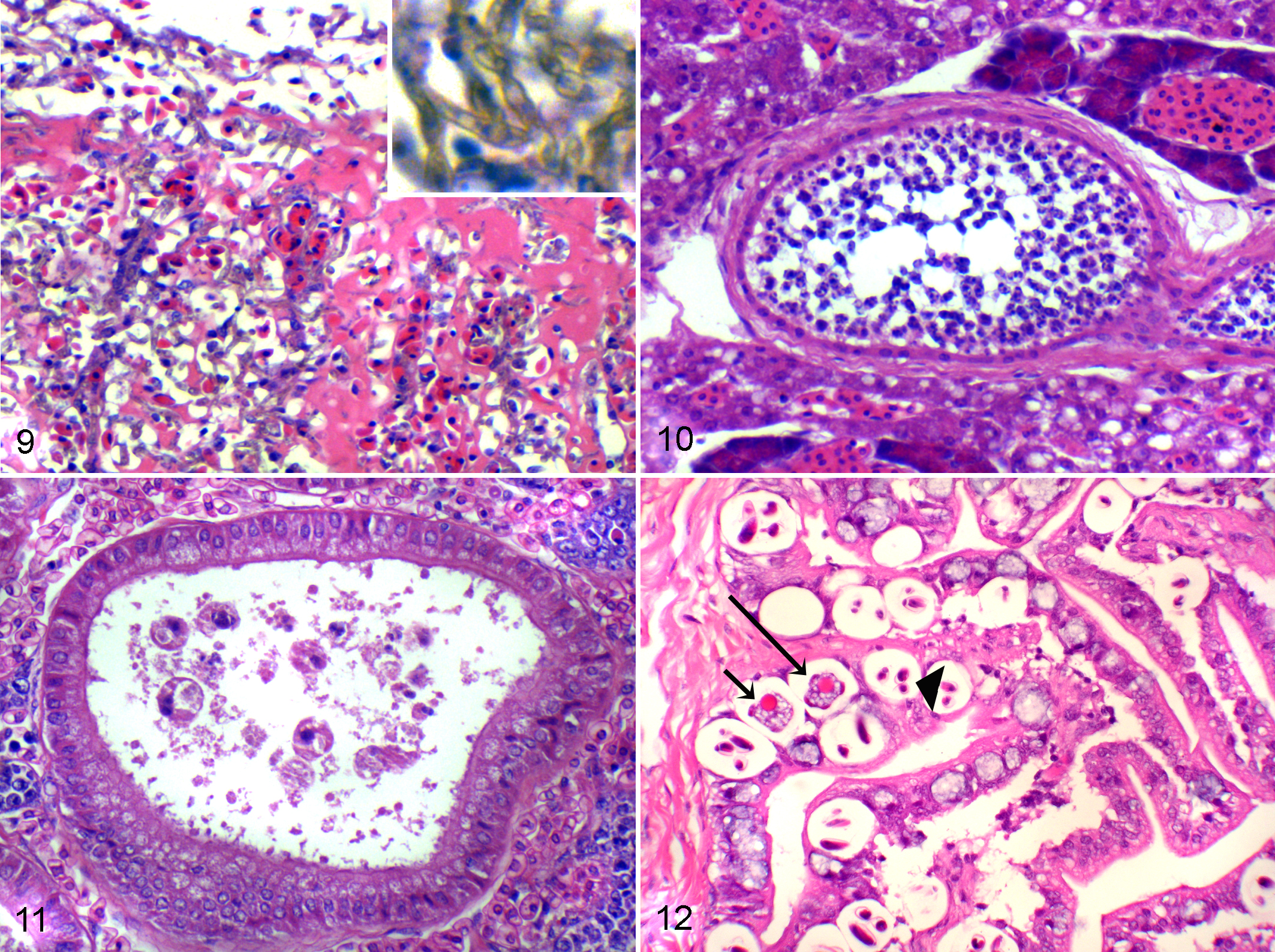
Myxozoanosis
Myxozoanosis has previously been described in weedy seadragons. 10 Renal lesions consisted of both early stages and mature spores of myxozoans visible in proximal and distal tubules, Bowman’s space, and glomerular capillaries. There was accompanying renal tubular dilatation and tubular epithelial cell hypertrophy, and these spores stain periodic acid–Schiff (PAS) positive, with positive staining bodies in the sporoplasm and positive staining of the shell valves. Myxozoan spores were also occasionally seen in bile ducts (Figs. 10, 11).
Weedy seadragon; gill. Congestion, fibrin deposition, and necrosis of filaments and lamellae are associated with intravascular and extravascular brown-pigmented septate fungal hyphae typical of phaeohyphomycosis. Hematoxylin and eosin (HE). Inset: higher magnification of extravascular brown-pigmented fungal elements. HE.
Coccidiosis
Intestinal coccidiosis also has been previously described in weedy seadragons. 27 Developmental stages of the coccidian were present in low to moderate numbers in enterocytes. Stages of the coccidian, which included gamonts and sporulated oocysts, were located within vacuoles in the cytoplasm of enterocytes. Mild, minimal, or no attendant inflammatory response was detected (Fig. 12).
Nonmycobacterial Bacterial Infections
Bacterial infections caused by non–acid-fast bacteria were found in 19 LSD and 17 WSD. The great majority of these were found in cases of dermatitis, often secondary to scuticociliatosis, and cases of septicemia. Where tissue Gram stains (eg, the Brown & Brenn stain) were performed, the bacteria were Gram negative, and the few cultures that were available yielded Vibrio sp.
Gas Bubble Disease
Gas bubble emboli were found in blood vessels in multiple organs, especially gill, facial soft tissues, intestine, and brain. Compression of the surrounding tissue was common.
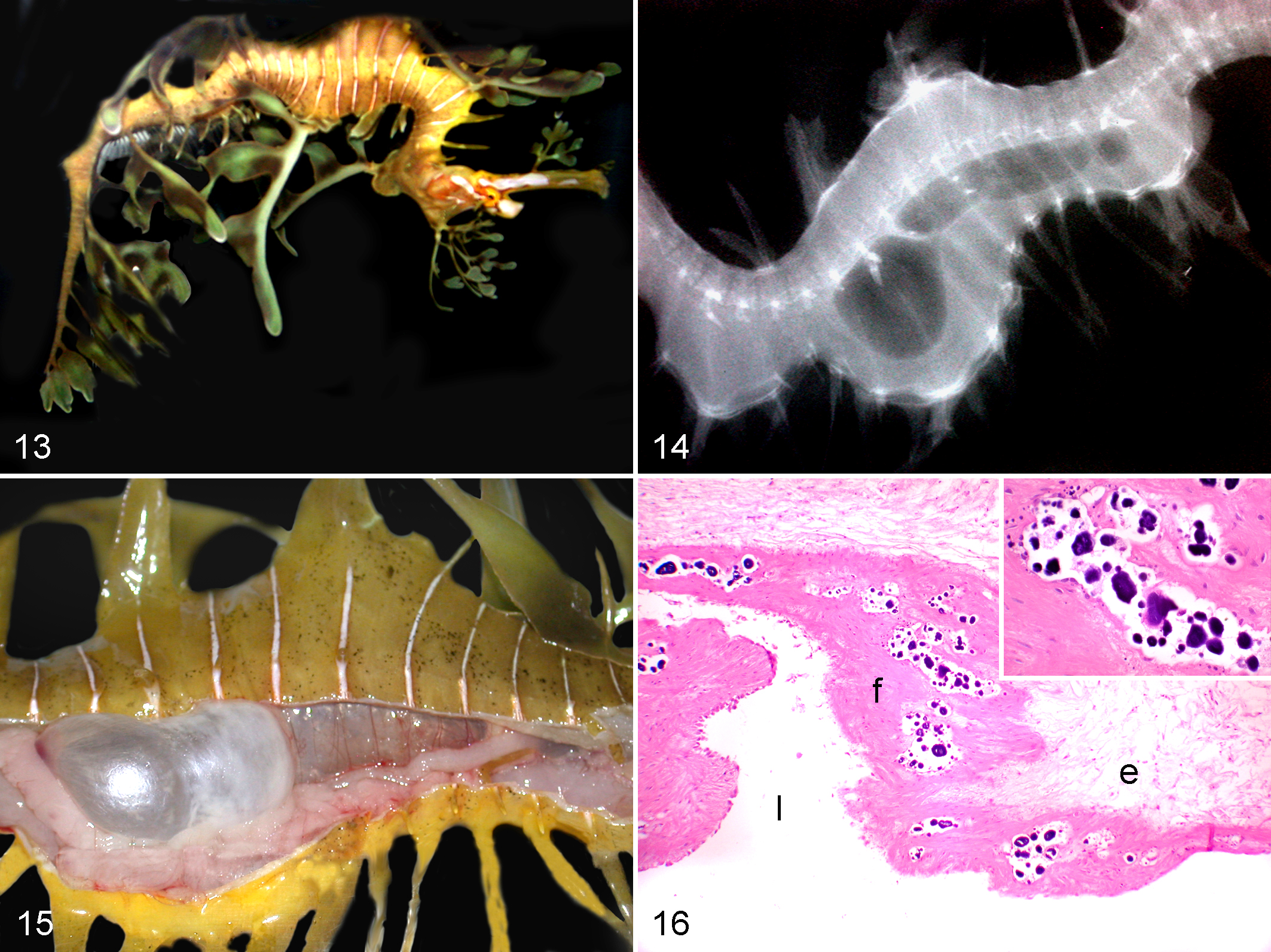
Swim Bladder Mineralization
This lesion consisted of mineralization of the stromal cell layer (subserosal connective tissue) of the swim bladder, with associated edema, causing overall thickening of the wall. Affected bladders were sometimes hyperinflated, and fish often displayed clinical buoyancy abnormalities (Figs. 13–16). There was usually no associated inflammatory infiltrate or evidence of infectious organisms; however, there were sometimes inflammatory lesions in other organs (such as scuticociliatosis or mycobacteriosis), which may or may not have been related to the pathogenesis of the swim bladder mineralization.
Leafy seadragon with positive buoyancy disorder due to swim bladder mineralization. Note that fish is upside down and that a portion of the fish is floating above the water surface.
Uncommon Disorders
WSD had a single case of islet cell neoplasia. LSD had 1 or 2 cases each of intestinal cryptosporidiosis, hepatocellular adenoma, islet cell neoplasia, and trauma.
Miscellaneous Disorders of Undetermined Etiology
In addition, WSD had 1 or 2 case diagnoses of goiter, generalized edema, gill necrosis, nephrolithiasis, renal tubular degeneration, renal hematopoietic depletion, and proliferative enteritis. LSD had an additional 1 or 2 case diagnoses of thyroid follicular hyperplasia, scoliosis, osmotic disturbance, gill necrosis, renal necrosis, nephrolithiasis, renal hematopoietic depletion, pancreatitis, and proliferative enteritis. In these cases, although not visible in the sections examined, enteritis could have been caused by bacterial infection or parasitic infection (eg, nematodes, trematodes, Cryptosporidium, etc) that were not in the plane of section. Thyroid lesions may be secondary diminished seawater iodine in closed-system aquaria versus natural seawater or to lower iodine levels in captive food supply. Pancreatitis in a single LSD was likely subsequent to extension of bacterial infection secondary to enteric trematodes, which were found in low numbers in the examined sections.
Discussion
Scuticociliatosis and vibriosis have been previously reported in weedy and leafy seadragons. 2,6,10 –12,18 Uronema spp are histophagous scuticociliatas, and one study reported Uronema spp to be the most common infectious agent identified in leafy seadragons. 18 Another study found scuticociliatosis to be less common than metazoan parasitism. 12 An aquarium in Japan reported scuticociliate infection in leafy seadragons, causing severe dermatitis and skin slough without invasion of internal organs. Based on the morphology of the organism, however, they believed their organism to be in the genus Philasterides, instead of Uronema. 26 Another aquarium reported DNA evidence supporting Philasterides dicentrarchi as the cause of severe scuticociliate infection of both WSD and LSD. 22 Therefore, we consider all ciliate protozoal infections simply as “scuticociliatosis” here. Interestingly, although another scuticociliate, Brooklynella hostilis, has been found to be a serious opportunistic pathogen in several species of seahorses, none of the scuticociliatosis cases in this study were considered to have the histomorphologic features of Brooklynella sp. 4 Because species identification by histologic examination is tenuous and difficult due to fixation artifact, molecular phylogenetic analyses of sequences polymerase chain reaction (PCR) amplified from scuticociliate infections are needed to fully characterize those ciliates causing lesions in seadragons. An important and distinguishing feature of scuticociliate infection in seadragons is its tissue-invasive quality; ciliates were often identified in the dermis, subdermal connective tissues, muscle, and even coelomic viscera with intravascular scuticociliates commonly found in multiple internal organs and the gill. This pathologic presentation is in contrast to certain other clinically significant ciliates, such as Cryptocaryon irritans, which typically are limited to epithelia of the skin and gill, and although not taxonomically related to these fishes, dolphins have also been found to have intravascular scuticociliates in cases of dermal pathology. 25
Myxozoanosis has been described in seahorses, 27 pipefishes, 15 and specifically WSD, caused by a new species of myxozoan parasite, Sinuolinea phyllopteryx. 10 All but one of the observed cases of myxozoanosis in this study were in WSD. The occasional practice of housing these 2 species together in the same captive environment would seem to encourage cross-species transfer of the organism, although it has been shown for certain species of myxozoans that completion of the life cycle requires an invertebrate alternate host, which may not be readily available in artificial systems. 16 It is interesting to note that although most WSD in captivity are wild-caught, since late 1997, all LSD brought into captivity have been captive-reared offspring from wild-caught males carrying eggs. The eggs are allowed to hatch in captivity, and the adult male is then returned to its capture site. Therefore, although not truly “captive bred,” most recently imported LSD were not exposed to pathogens in nature, only what they might encounter in aquariums. The WSD and LSD may, therefore, not have the same exposure to certain diseases, which might account for some of the differences in disease prevalence between the 2 species. It is also important to note that the myxozoan parasites affecting the liver and kidneys of these specimens are very likely entirely different, and further characterization of the cause of hepatobiliary myxozoanosis would seem prudent.
Disseminated fungal infections were also a common diagnosis in captive seadragons. These have been described in both WSD and LSD and are sometimes caused by infection with 1 of 2 species of Exophiala—for example, Exophiala angulospora and Exophiala aquamarina, a novel species. 5,19 Exophiala spp are common pathogens of many species of fishes but predominantly cool-water species, such as seadragons. 5,8,19,23 Most cases of phaeohyphomycosis were diagnosed by morphology and staining characteristics on light microscopy, although several cases included here were diagnosed by culture and sequencing of PCR-amplified DNA. 19
Similarly, intestinal coccidiosis has been reported to be caused by a novel species of Eimeria in the WSD (Eimeria phyllopterycis). 20,27 Although intestinal coccidiosis was reported in 5 WSD in this study, no cases were found in LSD, which is consistent with the findings in another report. 18 Although other studies have found metazoan parasites to be the most common parasitic infections of syngnathids, 12,18 including seadragons, in this study there were fewer metazoan infections than scuticociliatosis, with 23 individuals having nematodes, trematodes, cestodes, acanthocephalids, or some combination thereof, versus 45 cases of scuticociliatosis in WSD. For LSD, there were a total of 20 individuals with metazoan infections versus 37 protozoal infections. In both species, metazoan infections were usually incidental findings, whereas the scuticociliatosis was almost always the proximate cause of death. Although trematodes were noted in a number of specimens in this study, they were not identified further as monogeneans or digeneans. One study found only digeneans in their population of syngnathids, including seadragons, and digeneans were reported in a study of greater pipefish (Syngnathus acus). 16 However, another did find monogeneans in wild-caught pipefish Syngnathus rostellatus. 1
Neoplasia was identified in only 3 cases: 1 case each of pancreatic islet cell neoplasia in both weedy and leafy seadragons and 1 case of hepatocellular adenoma in a leafy seadragon. A cardiac rhabdomyosarcoma has been reported in a WSD, and it is the only published case of a neoplasm in a seadragon, although neoplasms have been occasionally identified in seahorses. 14,29
There was no apparent correlation between the most common infectious diseases, mainly scuticociliatosis and mycobacteriosis, with zero cases of leafy seadragons having both infections and only 1 WSD having both scuticociliatosis and mycobacteriosis. This is actually less than might be expected based on the expected probability of coinfection determined by random mass sampling in a bivariate population. Several cases of dermal scuticociliatosis in both species had secondary infections of nonmycobacterial bacteria.
Myxozoanosis was almost always encountered with some other disease process, the more common of which were scuticociliatosis (8 cases), branchitis (7 cases), cestodiasis (7 cases), and systemic fungal infection (5 cases).
It is also interesting to look at the absence of certain diseases in the population of seadragons in this study. Hepatitis and pancreatitis have been reported in tiger-tailed seahorses (Hippocampus comes) associated with blister-like lesions of the skin and inclusion bodies in the pancreatic lesions resembling birnavirus inclusion bodies. 24 However, no such cases were seen in seadragons in the present study; only 1 case of pancreatitis was observed in a LSD, and viral inclusions were not observed. The only indications of viral disease were 2 cases of branchitis resembling viral infection. Similarly, although Glugea heraldi, the microsporidian known to cause so-called blister disease in seahorses, is one of the better described pathogens of seahorses, 3,28 no cases of Glugea sp or other microsporidian infection were observed in the seadragons in this study. Similarly, epitheliocystis was observed in 4 weedy seadragons but not in any leafy seadragons. Epitheliocystis has previously been reported in the leafy seadragon, 13 with molecular evidence associating Chlamydiales bacteria with the infection. 17
By far the most serious infectious diseases affecting WSD and LSD are scuticociliatosis, mycobacteriosis, and infection by melanized fungi (eg, Exophiala spp). Strategies for improving captive longevity of this species will need to concentrate on prevention and control of these organisms. The role of live foods for carrying bacterial, mycobacterial, and myxozoan pathogens should be investigated. Similarly, the effects that different aquarium temperatures have on pathogen prevalence (eg, lower temperatures may favor myxozoanosis, whereas higher temperatures may favor mycobacteriosis) should also be studied. Gas bubble disease, although common in seahorses, was not commonly identified in seadragons in this study. However, some cases do occur, possibly subsequent to super-saturation events in the water causing absorption through the gills.
Mineralization of the swim bladder is an interesting and not well-understood problem primarily of the leafy seadragon. Whether this is a caused by diet, water chemistry, or other as yet unknown causes is open to speculation. Aquarists often supplement closed seawater systems with calcium hydroxide to maintain adequate dissolved calcium levels for coral growth, as well as “gut-loading” feeder mysid shrimp with food supplemented with calcium. What role these supplementation regimes, if any, play in the formation of the mineralization of the swim bladder would seem to be worthy of study. Renal calcinosis has been described in salmonids fed synthetic diets deficient in magnesium, 7 and the role of dietary magnesium also probably warrants investigation. Metastatic mineralization of tissues has been reported in salmonids subjected to prolonged, elevated carbon dioxide levels in the water, although tissues other than the swim bladder were affected. 9 Considering that seadragons usually must be kept in relatively large, well-filtered aquarium systems with relatively low biomass, it would seem unlikely that these seadragons were subjected to the kind of chronic, elevated carbon dioxide levels necessary to produce metastatic mineralization, which is more typical of a hatchery setting where aquarium biomass levels are high.
Swim bladder hypoplasia is an occasional clinical diagnosis, where the absolute size of the swim bladder in a juvenile does not develop to the normal proportions of an adult during growth in captivity. Seadragons with swim bladders that fail to develop properly in this fashion cannot maintain neutral buoyancy. This has been observed not only in institutions contributing to this study but also in other institutions as well (P. Mohan, personal communication, 2012). In one institution, this disorder was associated with calcium hydroxide supplementation of the tank water; in another, there was no such supplementation, but weekly water changes were performed, and the animals were kept in relatively shallow tanks with little opportunity for vertical migration (D. Richardson, personal communication, 2012; P. Mohan, personal communication, 2012). This problem has not been seen in almost 10 years, and until additional cases are observed, and both clinical and husbandry correlates are well described, the pathogenesis of swim-bladder hypoplasia will remain unknown. Interestingly, the swim bladders in these cases were histologically normal, with no microscopic lesions to account for the failure of the organ to grow to normal size.
Continued application of rigorous pathologic surveillance for disease combined with clinical studies of these problems should help diminish the morbidity and mortality of these delicate and fascinating creatures in captivity.
Footnotes
Acknowledgements
We thank Histology Consulting Service and the Histology Laboratory of the CVMDL for histotechnological services, Elizabeth Post and Sue Hutton of Northwest ZooPath for data retrieval, and Christie Buie of Northwest ZooPath for image layout. We also thank Daryl Richardson, Paula Carlson, Charles Innis, VMD, Dipl. ABVP (Reptile and Amphibian Practice), Akinyi Nyaoke, BVM, MSc, PhD, James Raymond, DVM, Dipl. ACVP, Inga Sidor, DVM, Dipl. ACVP, Karen Wolf, DVM, Dipl. ACZM, and veterinary anatomic pathology residents of the Department of Pathobiology and Veterinary Science, University of Connecticut for help, advice, and support.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.