
Abstract
Mast cells might play an important role as the major effector cells in the immune response against Cryptosporidium parvum. C. parvum is a protozoan parasite that causes cryptosporidiosis in animals and humans worldwide. To investigate the interaction between C. parvum and mast cells during infection, nine 3-day-old male calves were orally challenged with 106 oocysts of C. parvum per calf. The distribution of mast cells in the mucosa of the small intestine was analyzed by toluidine blue staining. The concentrations of histamine and the cytokines interferon-γ, interleukin-4, interleukin-2, and interleukin-12 were measured in the serum, and the histamine levels were also determined from the intestinal contents. The following clinical signs were monitored: nausea, watery diarrhea, dehydration, and weight loss. Oocysts were shed in the feces during the infection period. C. parvum infection induced an increase in mast cell numbers in the mucosa of the small intestine in distinct temporal and spatial patterns. Infection with C. parvum can induce mastocytosis in the entire small intestinal mucosa in immune-competent calves, and the presence of the parasites influences the distribution profile of the mast cells.
Keywords
Cryptosporidium parvum, an epicellular pathogen belonging to the apicomplexa phylum, causes cryptosporidiosis, a food- and water-borne disease that occurs worldwide. 4,7 The organism infects the gastrointestinal tract of both immune-competent and immune-deficient animals and humans. 8,31 In immune-competent individuals, acute infection with C. parvum leads to self-limiting diarrhea. 5 However, in immune-deficient hosts, such as those with acquired immune deficiency syndrome (AIDS), the infection caused by C. parvum is considered to be chronic and life-threatening. 27
Mast cells are multifunctional tissue-dwelling cells capable of secreting a wide variety of mediators. Following activation and degranulation, mast cells release mediators such as histamine, prostanoids and leukotrienes, matrix metalloproteinase-9 (MMP-9), and many chemokines and cytokines, all of which are pivotal to the generation of inflammatory responses. 23 Mast cells directly interact with bacteria and parasites and appear to play a vital role in host defense against pathogens. 18 Some parasitic infections, for example, Nippostrongylus sp, Trichomonas vaginalis, and Giardia lamblia, can induce the accumulation of mast cells, which can protect the host against parasites. 14,24,30
In mice, mastocytosis correlates with the expulsion of Cryptosporidium muris during a primary infection and acts as the defense mechanism upon challenge. 29 Recently, an immune-competent suckling rat model was established. 1,22 In these studies, C. parvum infections were able to induce digestive hypersensitivity (a key pathophysiological factor in functional digestive disorders, such as irritable bowel syndrome), and the intestinal hypersensitivity was closely associated with intestinally activated mast cell accumulation.
To date, there are no reports of the role of mast cells during C. parvum infection in cattle. The present study aimed to investigate the interaction between C. parvum and mast cells in calves during the infection period.
Methods
Parasite and Animals
C. parvum oocysts (Changchun isolate, Jilin Province, China, genotyped as C. parvum-cattle genotype, purified from naturally infected calf feces) were stored in a 2.5% K2Cr2O7 solution for fewer than 3 months before use in the Molecular Parasitology Laboratory of Jilin University, China. The oocysts in the feces were purified by sucrose gradient centrifugation as previously described. 21 The oocysts were bleached and rinsed in phosphate-buffered saline (PBS) before infection.
Nine 3-day-old Holstein-Friesian male calves purchased from farms in the suburb of Changchun were used as the infected group. These animals were orally challenged with 106 oocysts of C. parvum per calf, and blood samples from day 0 were collected after this oocyst challenge. The calves were housed separately in 5-m2 pens that had cement walls and floors with wood shavings and were fed milk thrice daily. In preliminary experiments, the animals began to shed oocysts on day 3 post infection (p.i.); the maximal amount of oocysts was shed on day 6 p.i., and parasite elimination occurred on day 9 p.i. The 9 calves were arbitrarily assigned to 3 groups, A, B, and C (3 calves per group), and they were slaughtered at 3, 6, and 9 days p.i., respectively. At each time point, serum and intestinal samples (the same location for each calf each time) were collected. The control group consisted of 9 calves raised under the same conditions. The serum and intestinal samples of the control group were obtained from the calves in a slaughterhouse. The animals in both the infected and control groups were tested for the presence of other intestinal pathogens by microscopy prior to use of the samples.
Food Intake
The milk intake of group C was recorded during the infection period from 0 to 9 days p.i. Three normal calves, fed under the same conditions in another farm, were used as the control group.
Excretion of Parasite Oocysts
Fecal samples were collected with a plastic bag that was fixed behind the hip of the calves each day after the oocyst challenge. One gram of feces from each sample was homogenized in 1 ml of water and 8 ml of Sheather’s sucrose solution. The number of oocysts in each fecal sample was counted using a cell counter. 33 The parasite burden was evaluated by calculating the number of oocysts per gram (OPG) of feces.
Enzyme-Linked Immunosorbent Assay (ELISA)
ELISA kits (BlueGene Biotech Co., Ltd., Shanghai, China) were used to measure the concentrations of gamma interferon (IFN-γ), interleukin (IL)-4, IL-2, and IL-12 in the serum samples according to the manufacturer’s protocol. The Bovine Histamine ELISA kit (BlueGene Biotech Co., Ltd) was used to measure the concentration of histamine in the serum and intestinal samples according to the manufacturer’s protocol.
Immunohistochemical Staining
Samples of the intestinal tract isolated at 3, 6, and 9 days p.i. were fixed immediately in Bouin’s solution for 24 hours and were then dehydrated and embedded in paraffin wax. Histologic 5-μm transverse sections were cut, deparaffinized, and rehydrated. Subsequently, the tissue sections were stained with toluidine blue (TB) or hematoxylin and eosin (HE), as described previously. 3,13 After dehydration and mounting in Canada balsam, the tissue sections were used for counting mast cells and observing pathological changes under the microscope.
Counting Mast Cells
Nine areas (0.16 mm2) were arbitrarily selected in the mucosal samples from each calf, and the mast cell numbers were counted in each area at 400× magnification by 3 independent investigators. The mast cell numbers in each 1-mm2 area were calculated.
Statistical Analysis
The data were expressed as the mean ± standard error of the mean (SEM; 95% confidence interval). One-way or 2-way analyses of variance (ANOVA), Bonferroni post tests, and Student’s t-tests were used to analyze the data. All analyses were performed using GraphPad Prism 5 software. P values <.05 were considered statistically significant.
Results
Clinical Signs
One day after the calves were challenged, nausea was evident. Compared with the food intake of the control group (as shown in Fig. 1 and Fig. 2), the food intake of the infected calves decreased during the prepatent period. Food intake reached its lowest levels at 3 days p.i., when the calves with watery diarrhea began to shed oocysts in their feces. Between days 3 and 6 p.i., the number of oocysts shed in the watery feces increased, dehydration (dry nasal mucosa and sunken eye socket) was greatly increased, and food intake increased slightly and then declined again. Subsequently, the number of excreted oocysts peaked at 5 and 6 days p.i. Thereafter, the condition of the calves improved, accompanied by a decrease in oocyst shedding. At 9 days p.i., the oocysts excretion ceased, and the watery diarrhea ended.
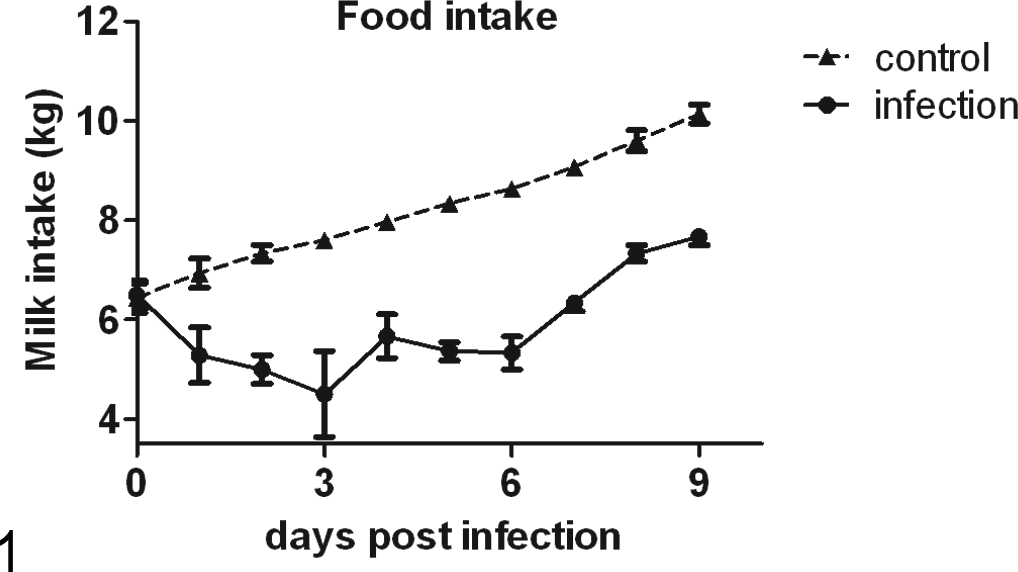
Changes in food intake. Compared with the food intake of the control group, that of the infected calves was significantly lower during the infection period. (P < .01 at 1 day post infection and P < .001 from 2 days post infection to 9 days post infection).
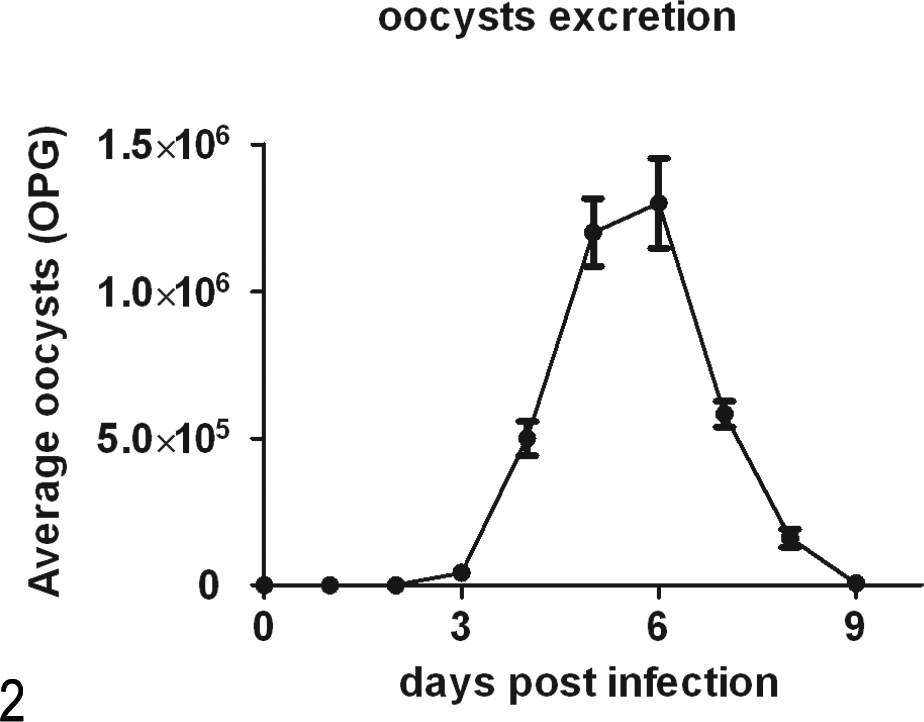
The excretion of parasite oocysts. The parasite burden was evaluated by counting the number of oocysts per gram (OPG) of feces.
Using HE staining (Figs. 3–8), a histopathological evaluation at 6 days p.i. revealed epithelial damage of the small intestine with villous atrophy, crypt hyperplasia, and inflammatory cell infiltration. The intervillous spaces increased at 3 days p.i., reached a peak at 6 days p.i., and then recovered at 9 days p.i. The epithelial damage was less evident at 9 days p.i., and several parasites were found in the ileum. At the peak of infection (6 days p.i.), large numbers of parasites were found in the jejunum, and a large number of immune cells was observed in the lumen of the intestine. However, very few parasites were observed in the jejunum at 3 and 9 days p.i. No parasites were found in the duodenum during the patent period.
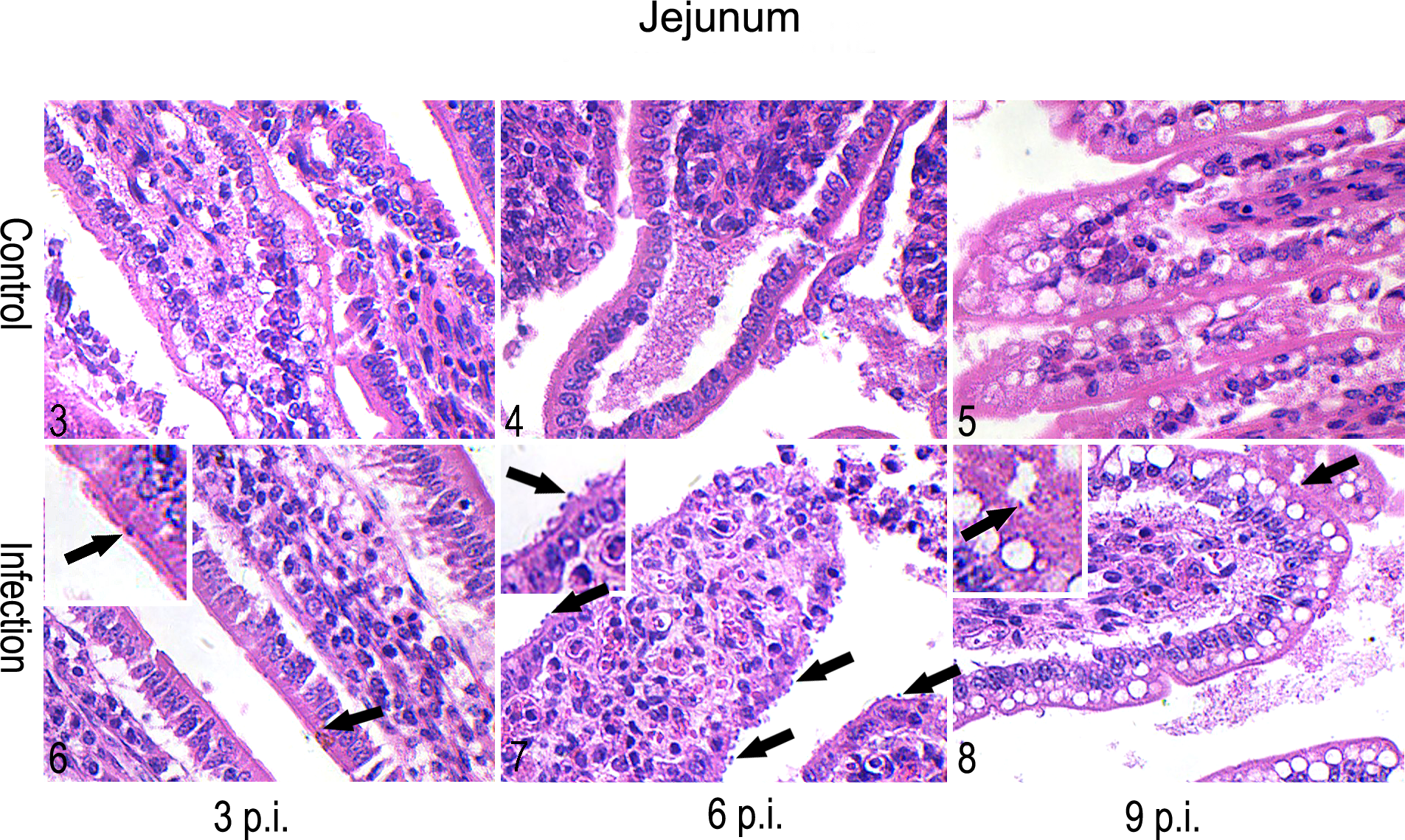
Intestinal mucosa; Holstein-Friesian male calves, C. parvum-infected and control groups. Shown are the comparisons of the intestinal mucosa during C. parvum infection. The representative images shown are from the infected group (6–8) and the control group (3–5) in the jejunum at 3 days p.i. (3 and 6), 6 days p.i. (4 and 7), and 9 days p.i. (5 and 8). The black arrows indicate the parasitophorous vacuoles of C. parvum. Images show the changes of the number of parasitophorous vacuoles and the epithelial damage of the small intestine with villous atrophy and inflammatory cell infiltration in jejunum during infection. The sections were strained with hematoxylin and eosin (HE).
Cytokine Concentration in the Serum
The ELISA results (Fig. 9) revealed that the levels of the Th1 cytokines IFN-γ, IL-2, and IL-12 and the Th2 cytokine IL-4 in the sera of the infected group were significantly lower than those in the control group at 3 days p.i. (P < .001). At 6 days p.i., the IFN-γ, IL-4, and IL-12 levels in the infected group were significantly lower than those in the control group. At 9 days p.i., only the IFN-γ and IL-4 levels in the infected group were significantly lower than those in the control group. Almost all of the cytokine concentrations in the infected group were lower than those in the control group, with the exception of IL-12, which was higher in the infected group at 9 days p.i. The Th1/Th2 bias was represented as the ratio of the concentration of a Th1 cytokine (IFN-γ) to the concentration of the Th2 cytokine (IL-4) (Fig. 10). The ratio in the infected group was significantly higher than that in the control group at 6 and 9 days p.i. (P < .01 and P < .05, respectively). In contrast to the gradual decline observed in the control group, the ratio in the infected group showed fluctuations. Little change was observed at 3 days p.i., but the ratio then increased quite rapidly and reached a peak at 6 days p.i. (when the host sheds the maximum number of oocysts), and then the ratio declined in conjunction with parasite clearance at 9 days p.i.
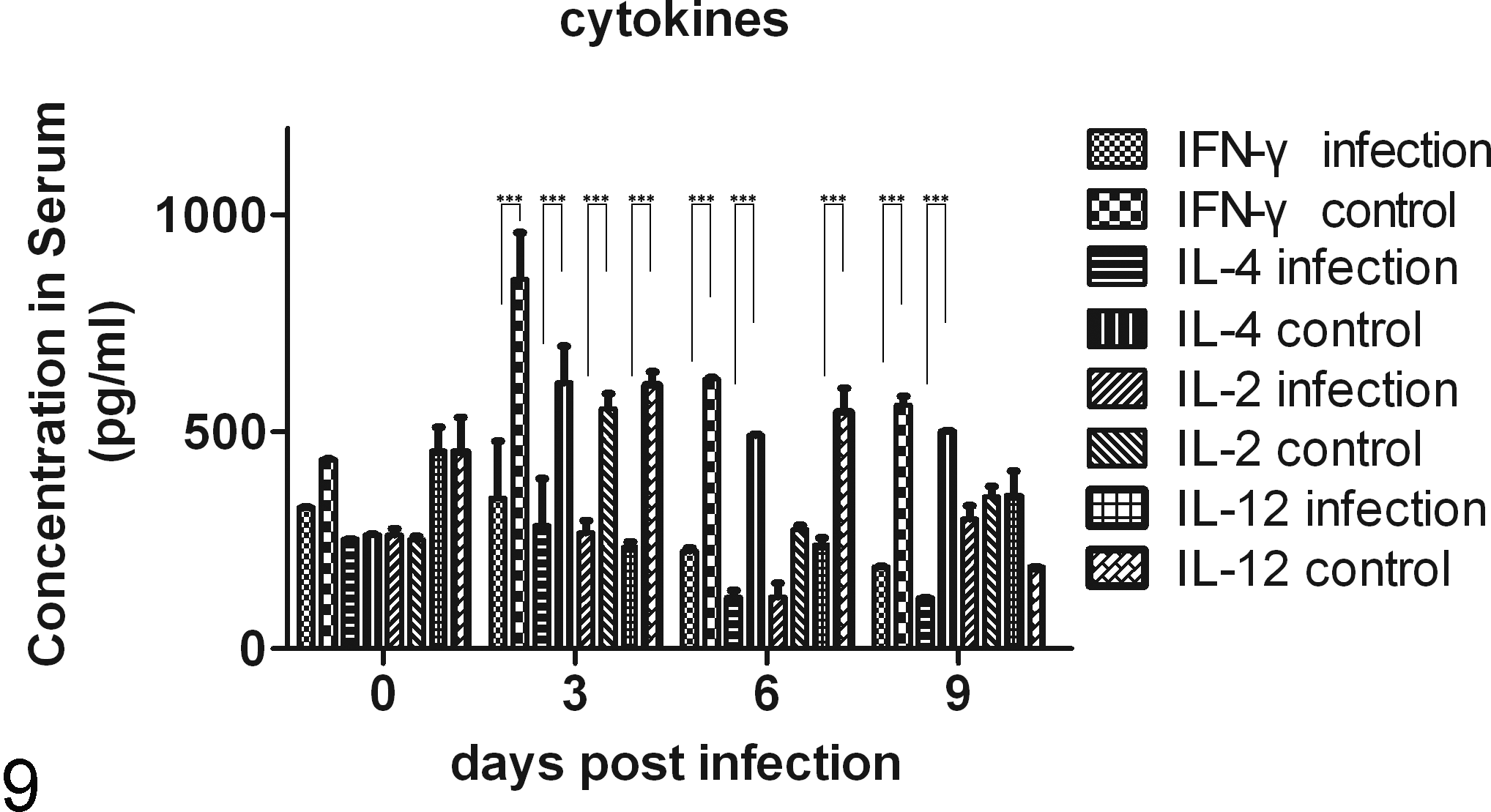
The levels of cytokines in the serum during the infection. The concentrations of IFN-γ, IL-4, IL-2, and IL-12 in the infected group were significantly lower than those in the control group during the patent period. ***P < .001.
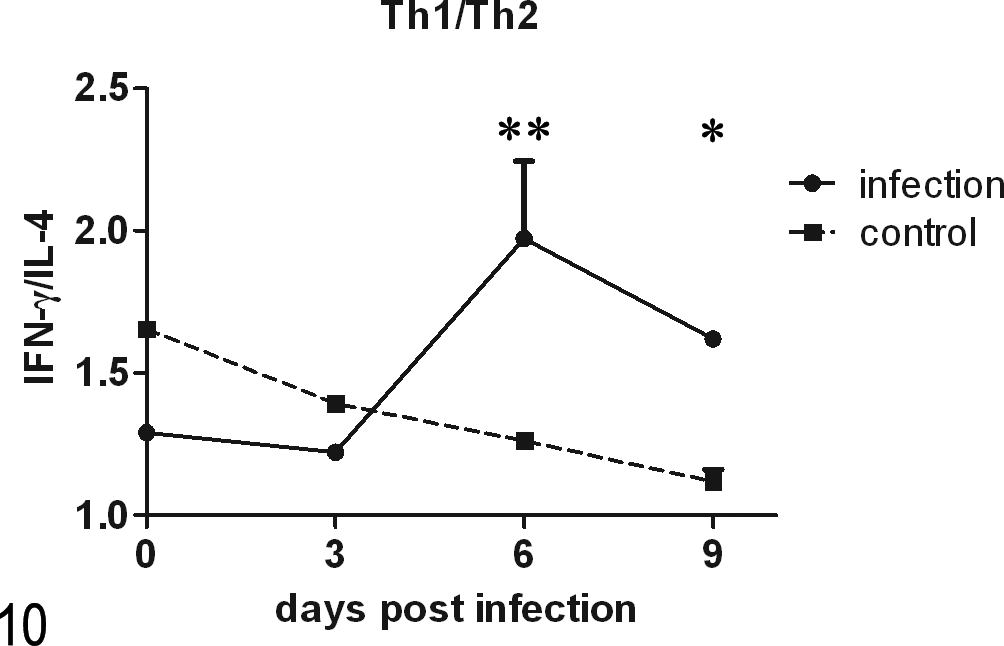
The change in the Th1/Th2 bias. The ratios of the concentrations of IFN-γ and IL-4 reflected the change in the Th1/Th2 bias. *P < .05.
Mast Cells
Figures 11–16 show the transverse sections of the intestines of animals at 6 days p.i. that were stained with TB. Figure 17 indicates that the number of mast cells increased in the mucosa of the small intestine (duodenum, jejunum, and ileum) after challenge with C. parvum oocysts. The mean number of mast cells per square millimeter increased steadily from 3 to 9 days p.i. in the mucosa of the duodenum and jejunum and was not decreased at 9 days p.i. In the ileum, the mast cell numbers in the infected group were significantly greater than those of the control group at 9 days p.i. In other words, the numbers of mast cells were significantly increased in the infected group when the parasites had been eliminated (P < .001). In the jejunum, the mean number of mast cells was significantly greater in the villi of the infected group than in the villi of the control group at 3, 6, and 9 days p.i. (P < .01, P < .001, and P < .001, respectively). The accumulation of mast cells was not significantly different among the duodenum, jejunum, and ileum in the control group (P > .05). The accumulation of mast cells was similar between the jejunum and ileum in the infected group. However, the mean number of mast cells was significantly greater in the duodenum than in the jejunum and ileum at 6 (P < .001, P < .001) and 9 (P < .01 and P < .05, respectively) days p.i.
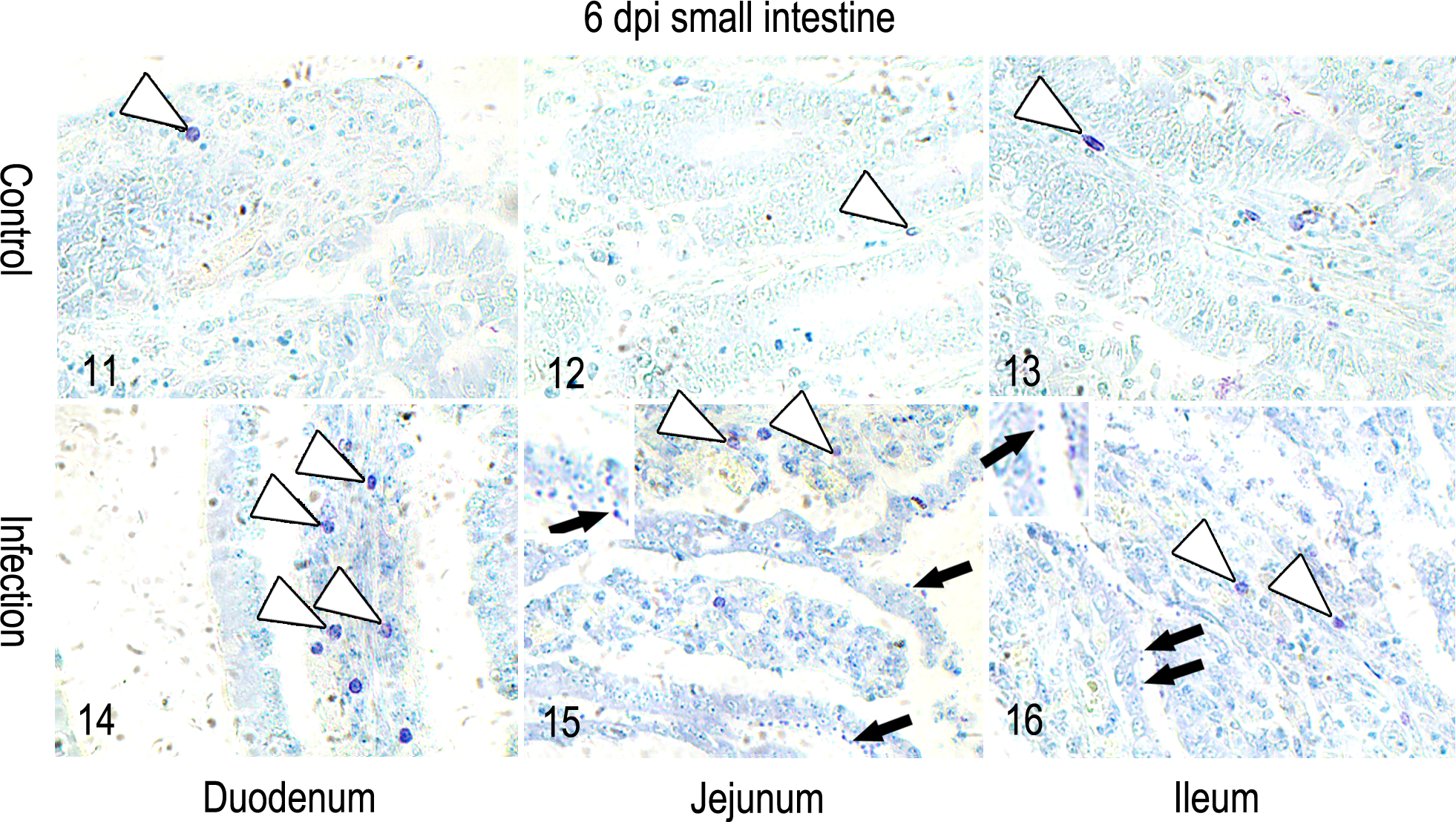
Intestinal mucosa; Holstein-Friesian male calves, C. parvum-infected and control groups. Shown are the comparisons of the accumulation of mast cells in the intestinal mucosa during C. parvum infection. The representative images shown are from 6 days p.i. The white arrows indicate mast cells. The black arrows indicate the parasitophorous vacuoles of C. parvum. Images show the profile of parasitophorous vacuoles and mast cells at 6 days p.i. The sections were strained with toluidine blue (TB).
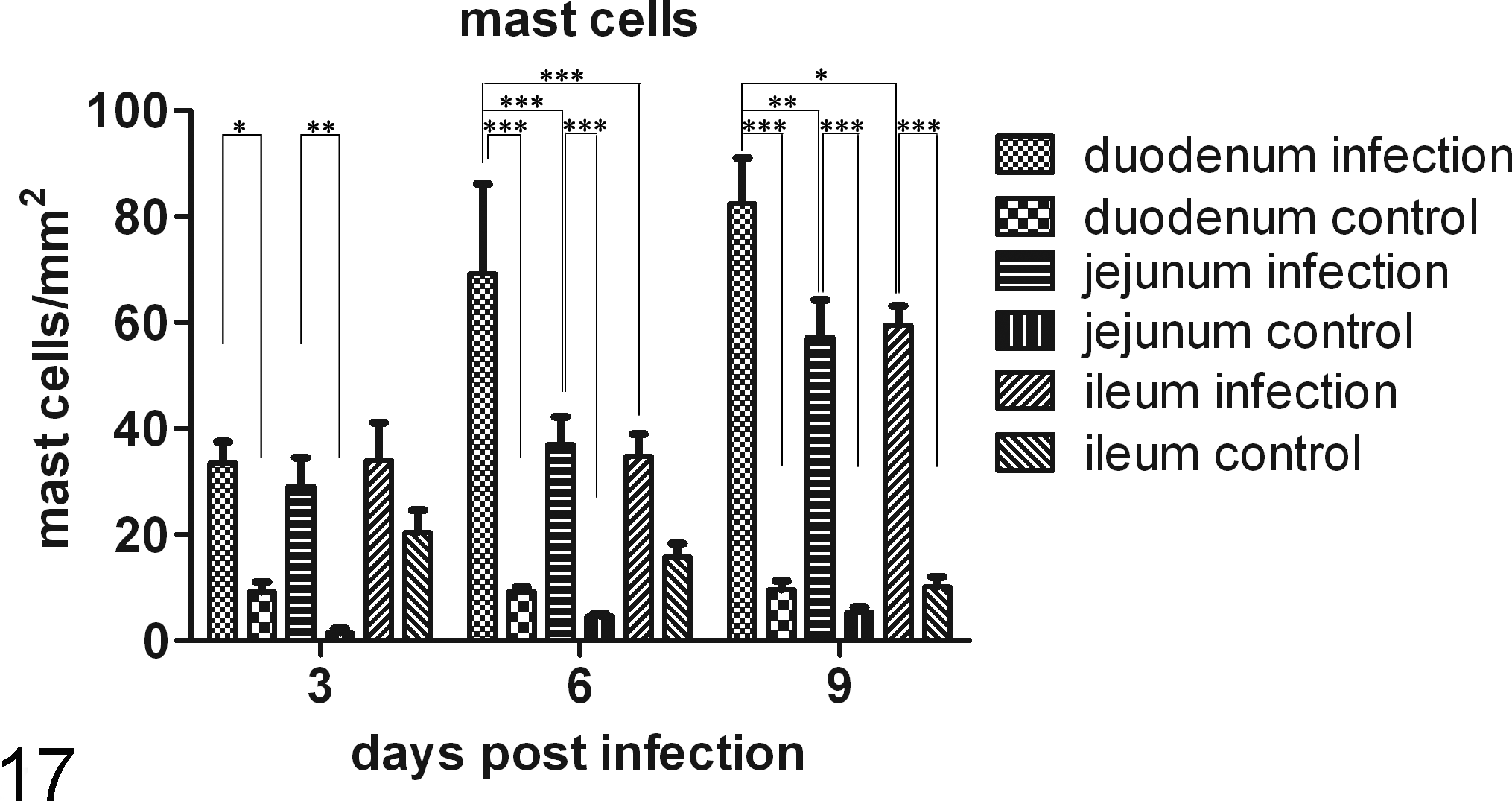
Changes in the number of mast cells during the infection. *P < .05, **P < .01, ***P < .001.
Histamine Concentrations
Figure 18 shows that although there was no statistical significance, the levels of histamine in the serum and intestinal contents in infection groups were higher than the level in the control group (P > .05). The histamine concentrations in jejunum were higher than in duodenum, but the difference showed no statistical significance between the 2 groups (P > .05).
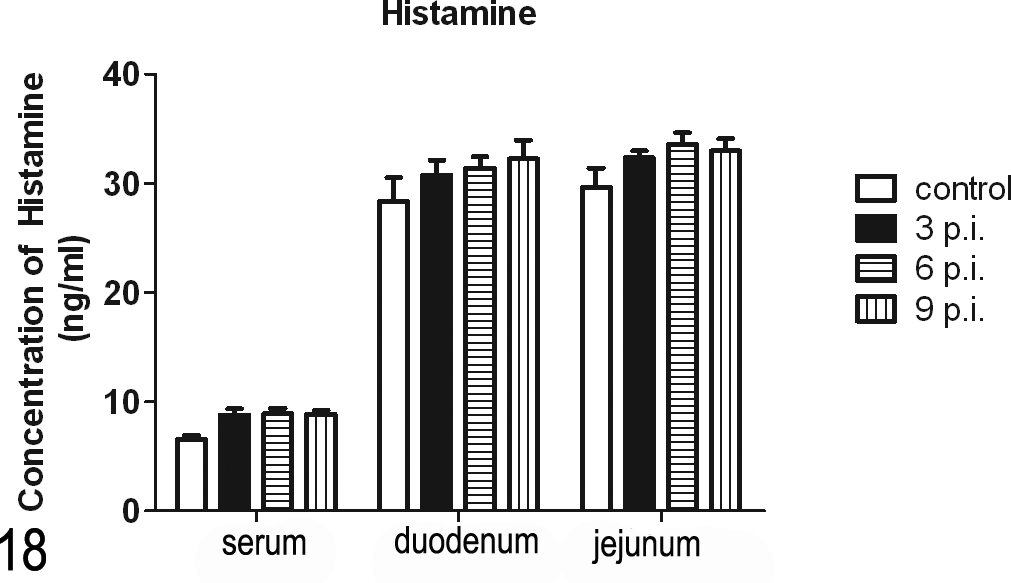
Changes in the histamine concentration during the infection. Although there was no statistical significance, the concentrations in the infection group were higher than in the control group.
Discussion
There are 152 species of mammals that have been reported to be susceptible to infection with C. parvum or a C. parvum-like organism. 9 Suckling calves are the main host of C. parvum. 10 The data presented here suggested that infection with C. parvum can induce the accumulation of mast cells in the mucosa of the small intestine in a cattle model.
The clinical signs of the calves challenged with C. parvum oocysts were similar to symptoms described in previous studies. 2,8 Most of the signs and symptoms correlated with the excretion of oocysts, with the exception of the profile of cytokines. Studies of human and animal cryptosporidiosis have indicated that the cytokine IFN-γ is important for resistance to and clearance of the infection. 20 Interferon-γ appears to be a major mediator of host resistance to C. parvum because a lack of IFN-γ is associated with severe, protracted intestinal cryptosporidiosis in humans. 11 The roles of IFN-γ and IL-4 are critical in Cryptosporidium-associated infections because these molecules contribute to the immune response. 17 Mast cells can secrete IL-2, which is a critical Th1 cytokine for Treg proliferation and function. 23 Interleukin-12 was shown to be required for inducing IFN-γ-dependent immunity to C. parvum in mice that lacked T and B cells. 16 However, in the present study, the lower concentration of serum cytokines in the infected group indicated that perhaps the host immune system was suppressed. Furthermore, the ratios between the concentrations of IFN-γ and IL-4 reflected a change in the Th1/Th2 bias. The ratios were skewed toward a Th1 response during the patent period, and the trend peaked when the maximal amount of oocysts was shed; the Th1 response then diminished in association with recovery from the infection. Our data suggest that the C. parvum infection induced a Th1 response, a finding that is in accord with previous studies. 12,26,32 The host gene expression profile may be altered to eradicate the parasite, and the cellular physiological parameters are also likely to be altered to meet the physical and metabolic requirements of the parasite given the success of Cryptosporidium as a mammalian pathogen. 25 The delicate balance between parasitism and the host protective response was shown in infected cultured human epithelial cells, where the expression of more than 200 host genes was altered. 6 The immune system of the calves was still developing, and the infection with C. parvum would therefore affect the response initiated by the immune cells. Perhaps the immune suppression caused by the parasitic infection resulted in the observed decrease in the concentrations of cytokines.
Mast cells are one of the major types of effector cells participating in the immune response to parasitic infections. In this study, mast cells played a major role in immunoregulation. Mast cell-deficient W/Wv adult mice were found to be significantly more susceptible to primary infection with C. parvum than normal adult mice were, 13 and mastocytosis correlated with the defense mechanism of mice challenged with C. parvum. 29 In an immune-competent suckling rat model, the accumulation of mast cells in the mucosa of the small intestine was associated with postinfectious irritable bowel syndrome, which is a common functional gastrointestinal disorder due to jejunal hypersensitivity. 15,28 Mast cells can activate Tregs to suppress inflammation via IL-2. 23 The mastocytosis observed in the current study was significantly different between the duodenum and jejunum where C. parvum was present, indicating that the C. parvum infection induced the accumulation of mast cells in the mucosa of the small intestine and that the distribution profile of mast cells was associated with the location and number of parasites. Because TB can only stain the cytoplasmic granules of mast cells, both the suppression of mastocytosis and the degranulation might be reasons for this phenomenon. Histamine is one of several compounds released during the degranulation of mast cells. 19 To confirm the difference of degranulation in mastocytosis in the duodenum and jejunum, the concentration of histamine was measured by ELISA. The ELISA results suggested that perhaps degranulation was not the main cause for the differences in mast cell numbers observed between the duodenum and jejunum. Furthermore, when the infected and control groups were compared, the concentration of histamine was higher in the infected group than it was in the control group, but the differences were not statistically significant. On one hand, the degranulation of mast cells slightly increased, and on the other hand, C. parvum could suppress the accumulation of mast cells in the small intestine. Perhaps this degranulation and suppression is the reason for the observed mast cell profile. Infection with C. parvum in calves can promote the accumulation of mast cells in the mucosa of the small intestine, and these cells may play a main role in the immune response against C. parvum.
In conclusion, infection with C. parvum induced mastocytosis in the mucosa of the entire small intestine in immune-competent calves, and the presence of the parasites influenced the distribution profile of the mast cells.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the National Key Technology R&D Program of China (No. 2007BAD40B05) and the High Technology Research and Development Program (863) of China (No. 2006AA10A207). We declare that the experiments comply with the current laws of China, where the study was performed.