
Abstract
In the central nervous system (CNS) of domestic animals, numerous specialized normal structures, unusual cell types, findings of uncertain or no significance, artifacts, and various postmortem alterations can be observed. They may cause confusion for inexperienced pathologists and those not specialized in neuropathology, leading to misinterpretations and wrong diagnoses. Alternatively, changes may mask underlying neuropathological processes. “Specialized structures” comprising the hippocampus and the circumventricular organs, including the vascular organ of the lamina terminalis, subfornical organ, subcommissural organ, pineal gland, median eminence/neurohypophyseal complex, choroid plexus, and area postrema, are displayed. Unusual cell types, including cerebellar external germinal cells, CNS progenitor cells, and Kolmer cells, are presented. In addition, some newly recognized cell types as of yet incompletely understood significance and functionality, such as synantocytes and aldynoglia, are introduced and described. Unusual reactive astrocytes in cats, central chromatolysis, neuronal vacuolation, spheroids, spongiosis, satellitosis, melanosis, neuromelanin, lipofuscin, polyglucosan bodies, and psammoma bodies may represent incidental findings of uncertain or no significance and should not be confused with significant microscopic changes. Auto- and heterolysis as well as handling and histotechnological processing may cause postmortem morphological changes of the CNS, including vacuolization, cerebellar conglutination, dark neurons, Buscaino bodies, freezing, and shrinkage artifacts, all of which have to be differentiated from genuine lesions. Postmortem invasion of micro-organisms should not be confused with intravital infections. Awareness of these different changes and their recognition are a prerequisite for identifying genuine lesions and may help to formulate a professional morphological and etiological diagnosis.
The nervous system represents a complex group of structures, including nervous and connective tissue and vasculature. 6 Different classification schemes have been used to better define its distinct structures. Terminology varies from macroscopy to histology, implying that various structures display a certain degree of variation between different species. In addition, recent studies substantiate the concept of compartmentalization, indicating that similar structures and cell types may differ functionally depending on the cellular microenvironment. 89 This complex organization is further complicated by an organ-specific reaction pattern of various resident cell types and associated structures. However, keeping in mind a few organ-specific components and their histological appearance, together with application of the overall stereotypic cellular response of resident and infiltrating cells, neuropathology should lose most of its diagnostic myth.
Neurological disorders in animals are frequently observed and may be caused by primary alterations of the central nervous system (CNS) or result from secondary involvement in the course of physical, metabolic, degenerative, inflammatory, infectious, or neoplastic processes. Pathologic-anatomic and pathologic-histologic examination is mandatory for diagnosis of neurological diseases in animals. It is essential for any pathologist evaluating brain sections to have a solid knowledge of microscopic neuroanatomy and to recognize the numerous normal specialized structures within the nervous system that should not be confused with pathological changes.33,41 In addition, the pathologist must be able to identify artifacts caused by manipulation or improper handling of nervous tissues that are common within sections of the CNS. These artifacts may mimic genuine lesions or may potentially mask underlying neuropathologic processes. 33
Considering the numerous and various changes during CNS development, aging, and in pathological conditions, it is essential to discriminate clearly between normal structures, incidental findings with uncertain or no significance, pathological changes, and developmentally and age-related changes of significance.2,96,112 Various cell types display increased plasticity. Astrocytes may have a fibrous, protoplasmic, or gemistocytic phenotype or are termed Alzheimer type II cells when exhibiting specific morphological features and immunoreactivity.46,52,113 Various microglial phenotypes, including rod-shaped, amoeboid, and Gitter cells, may be present within CNS lesions.46,99 Similarly, neuronal changes varying from total chromatolysis to necrosis as well as axonal and Wallerian-like changes may be present. 112 The present review focuses on the most commonly encountered neuroanatomic and neurohistologic structures, nonlesional changes, unusual cell types, and postmortem changes that can be observed in the CNS of domestic animals. It aims to demystify neuropathology and helps pathologists to handle the CNS like any other organ.
Normal Structures
Circumventricular Organs
Circumventricular organs (CVOs) are specialized brain structures that are located in the walls of the lateral, third, and fourth ventricles and often protrude into the lumen. 54 They are positioned at the interface between the neuroparenchyma and the cerebrospinal fluid (CSF)–containing ventricles and canals of the brain, mainly at the midsagittal line. The ependyma of the CVOs of the third and fourth ventricles may represent the vestige of a phylogenetically ancient group of protoneurons in the vertebrate brain, which had contact to the CSF. 10 Besides laboratory and domestic mammals, including the albino rat (Rattus norvegicus albinus), common hamster (Cricetus cricetus), domestic rabbit (Oryctolagus cuniculus), cat, dog, goat, 71 and bovine, 73 CVOs have been described in birds (eg, buzzard [Buteo buteo] and domestic pigeon [Columba livia domestica]), 84 as well as in primates (eg, vervet monkey [Cercopithecus pygerythrus]) and human beings. 30 There is no general agreement about the defining features and number of these peculiar brain structures. CVOs may be classified as either ependymal or paraependymal. 10 Ependymal CVOs include the choroid plexus and the subcommissural organ (SCO). Paraependymal CVOs are the subfornical organ (SFO), the median eminence/neurohypophyseal complex, the pineal gland, and the area postrema (AP). In addition, the vascular organ of the lamina terminalis (OVLT) is regarded as a CVO. These 7 structures are depicted in Fig. 1 at a glance.
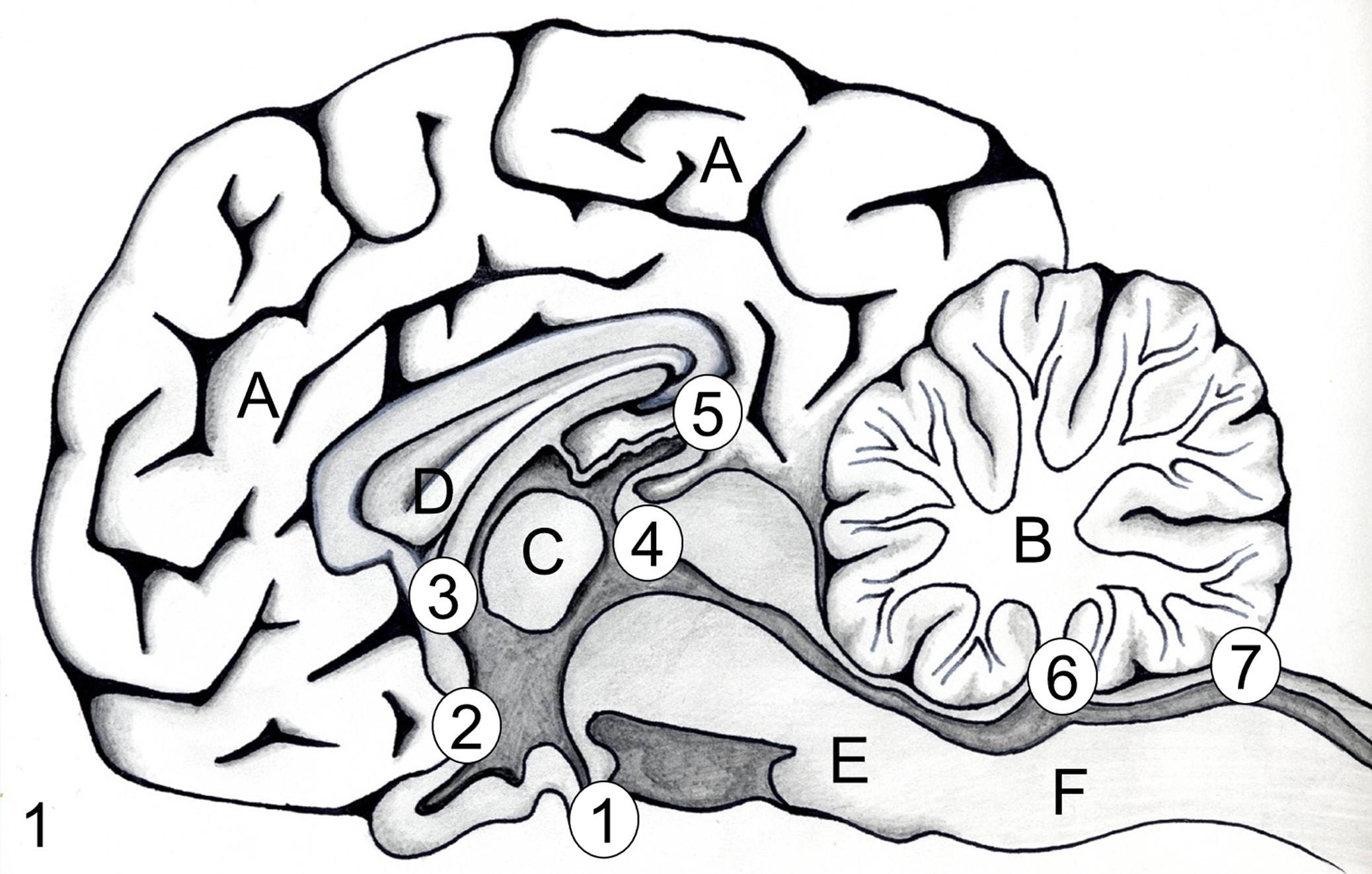
Schematic drawing of the neuroanatomical location of circumventricular organs with specification of their classification and function. Median eminence (ME)/neurohypophyseal complex, secretory, nonneuronal (1); vascular organ of the lamina terminalis, sensory/secretory, neuronal (2); subfornical organ, secretory, neuronal (3); subcommissural organ, secretory, nonneuronal (4); pineal gland, secretory, neuronal (5); choroid plexus of the IV ventricle, sensory, nonneuronal (6); area postrema, secretory, neuronal (7); the choroid plexus of the lateral and III ventricles are not shown. Cerebral cortex (A), cerebellum (B), adhesion interthalamica (C), septum pellucidum (D), pons (E), and medulla oblongata (F).
CVOs are characterized by several morphological and functional parameters. High vascularization represents a common hallmark of CVOs, characterized by many capillaries reaching near to the ventricular surface, 29 a phenomenon that should not be confused with granulation tissue. Except for the subcommissural organ, the capillaries of all CVOs have a fenestrated endothelium facilitating high permeability and disruption of the blood-brain barrier (BBB) that exists in central capillaries, consisting of both structural and functional components in a complex formed by the cerebrovascular endothelial cells, pericytes, and astroglial cell processes. 60 This particular arrangement allows direct exchange between blood and nervous tissue with bidirectional movement of molecules (eg, hypothalamic polypeptide hormones leaving the brain without disrupting the BBB or substances that do not cross the BBB but may still affect neural functions). 37 The luminal surface of CVOs is covered with morphologically distinct ependymal cells that are devoid of cilia or possess only few on their luminal surface, unlike the surrounding ventricular surface. 93 The ependymal cells are attached to each other by tight junctions restricting the flow of molecules between CSF and the CVO interstitium. 93
CVOs are also grouped according to their anatomical location, cellular composition, or function. The SFO, OVLT, and AP contain neuronal perikarya and have been previously classified as neuronal CVOs, whereas pineal gland, SCO, median eminence/neurohypophyseal complex, and choroid plexus (CP) were regarded as devoid of neurons and termed nonneuronal CVOs. 93 However, classic neurons have been recently described in the pineal gland of man, rabbit, monkey, ferret, and cotton rat. 86 Therefore, it should be reclassified as a neuronal CVO in these species. CVOs may also be distinguished according to their main function, sensitivity to substances, and secretory activity. AP and the SFO are responsive to blood-borne substances, whereas the median eminence/neurohypophyseal complex and pineal gland display predominantly secretory activity. Molecules may directly affect neurons in CVOs that in turn innervate specific nervous centers. 30 Secretory CVOs, like the neurohypophysis, release neurohormones into the bloodstream facilitated by fenestrated capillaries. However, not all CVOs fit into this classification. For example, OVLT seems to be an organ with a double function, as it is sensitive to circulating molecules and simultaneously receives input from the hypothalamus. 93 The SCO has imprecise functions, and many authors exclude it from the CVOs because of the lack of fenestrated endothelium.
Median eminence (ME)/neurohypophyseal complex
The pituitary gland is divided into 3 different parts according to architecture and functions: the adenohypophysis (anterior lobe), the intermediate lobe, and the neurohypophysis (posterior lobe) (Fig. 1). The neurohypophysis belongs to the circumventricular organs as it originates from the floor of the third ventricle. It is divided into a distal part, the neural lobe, and a proximal part, the median eminence, with an external and internal layer. An extension of the third ventricle, the hypophyseal recess, limits the median eminence. The morphology of the neurohypophysis varies considerably according to different species. Histologically, there are axons and neuroglial cells with branching cell processes termed pituicytes (Fig. 2). Pituicytes are regarded as special types of astrocytes collectively also termed aldynoglia by some authors. 49 They appear to be constituted of different populations according to the immunoreactivity for glial fibrillary acidic protein (GFAP; Fig. 3) and S100β protein.49,132 In addition, the nervous lobe shows an intense capillary network characteristic for CVOs. This capillary network receives hypothalamic neurohormones, including vasopressin and oxytocin, via an important nervous tract originating in the magnocellular paraventricular and supraoptic nuclei, which contain large neuroendocrine cells. This tract, the hypothalamo-posthypophyseal tract, courses through the ME to reach the blood vessels of the nervous lobe. 30
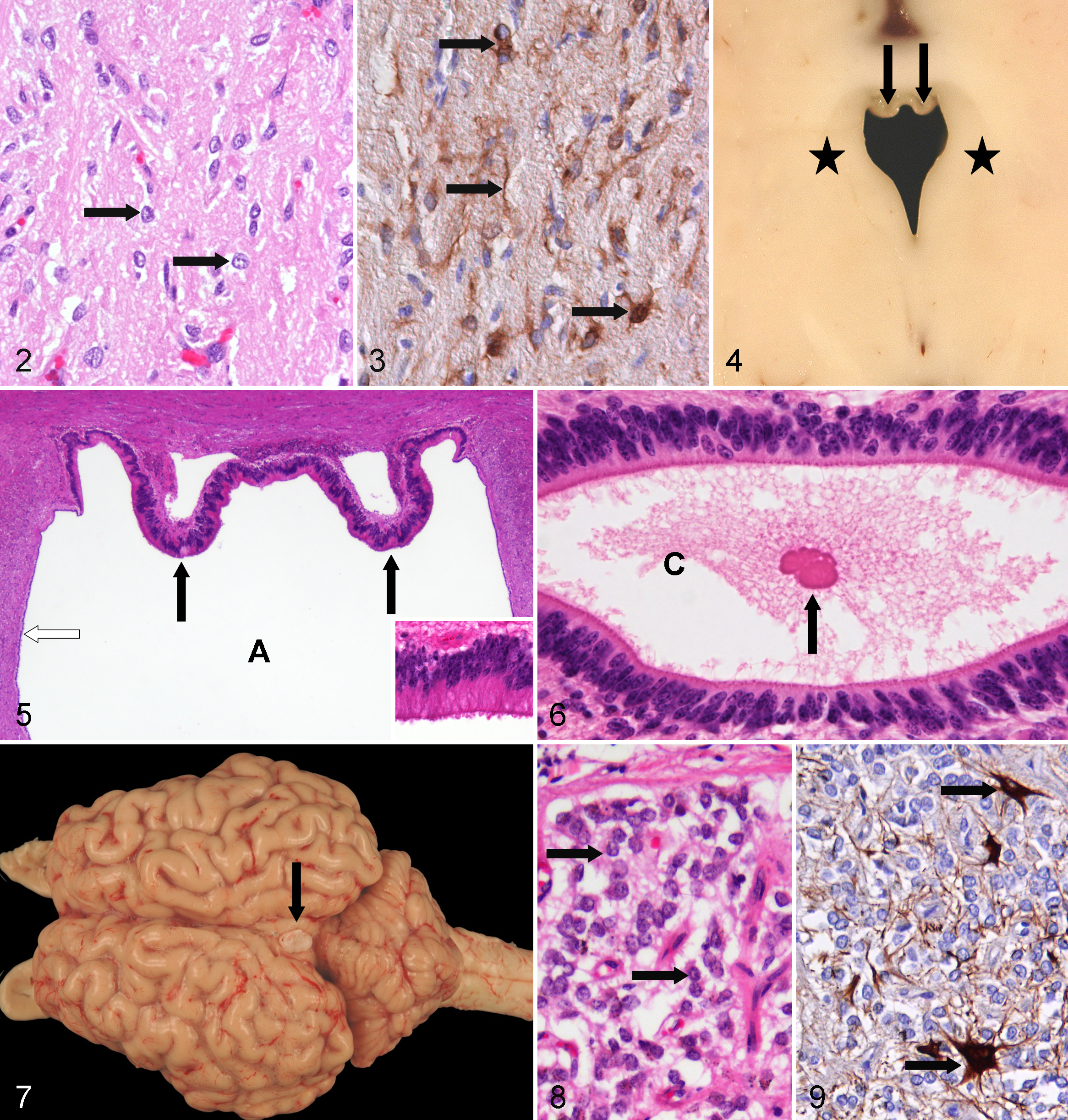
The ME is covered with specialized ependymal cells (ie, tanycytes) and composed of an internal layer containing the above-mentioned tract and an external layer receiving the tuberohypophyseal fibers from the hypothalamus, which transport the neurohormones toward the glandular cells of the adenohypophysis. This hormonal one-way traffic is achieved by a special vascular system: the hypophyseal-portal system. This system is composed of a primary plexus receiving the hypothalamic neurohormones and portal vessels reaching a secondary plexus, which in turn vascularizes the endocrine cells of the adenohypophysis. Herring bodies (corpusculum neurosecretorium) appear as normal hyaline granules in the neurohypophysis. They represent swellings of axonal endings containing clusters of neurosecretory granules and may be misinterpreted as axonal spheroids, viral inclusion bodies, or accumulated cytoplasmic material.
Organum vasculosum of the lamina terminalis (OVLT)
The cavity of the third ventricle is rostrally limited by the lamina terminalis, which extends from the anterior edge of the optic chiasm ventrally to the area of the anterior commissure dorsally. However, its dorsal extension varies considerably according to species. In rodents, it is limited to the suprachiasmatic region, whereas it reaches the anterior commissure in birds. Transversely, the lamina terminalis is limited to a sagittal fold that protrudes into the ventricular cavity above the optic chiasm (Fig. 1). 30
Morphologically, the OVLT consists of a superficial capillary network that lines the deep fold of the lamina terminalis. The deep network is composed of capillary loops originating from the superficial network. The tips of these loops do not reach the ventricular cavity, which is bordered by a fine subependymal vascular network. The typical feature of the OVLT is the relationship with the subarachnoidal spaces of the prechiasmatic cistern through its superficial capillary network and with the ventricular cavity through its deep capillary network. 30 The ependymal cells overlying the OVLT have tight junctions consisting of both zonula occludens and zonula adherens. 93
The blood vessels of the OVLT are devoid of a blood-brain barrier. Thus, osmosensory neurons and nervous fibers gain free access to blood-borne substances and are postulated to contribute to hyperosmolality-induced sympathetic activation. 114 Consequently, OVLT may play a part in controlling the release of vasopressin elaborated in the hypothalamic paraventricular and supraoptic nuclei and therefore act as an osmoregulator. In addition, distinct subpopulations of neurons of the OVLT express angiotensin II receptors mediating the actions of blood-borne angiotensin II. OVLT therefore also plays a role in stimulating water intake. 122 Furthermore, fibers of hypothalamic origin containing luteotropin hormone-releasing hormone (LHRH) reach the OVLT, which might participate in the control of cyclic production of the hypophyseal gonadotropic hormones.
Subfornical organ
The SFO is a small median structure attached to the anterior dorsal surface of the third ventricle at the level of the origin of the right and left columns of the fornix (Fig. 1). 26 It is highly vascularized with capillaries lined either by fenestrated or nonfenestrated endothelium. The surface is covered by flat ependymal cells with few cilia, and neurons are distributed throughout the organ.
The functions of the SFO are closely related to those of the vascular organ of the lamina terminalis. The SFO plays an important role in body fluid homeostasis, particularly drinking, cardiovascular regulation, and vasopressin release. 94 Neurons in the SFO organ are activated by central application of nicotine and plasmatic angiotensin II. 81 Most of the fibers originating from these neurons reach the paraventricular and supraoptic hypothalamic nuclei and thus regulate the release of vasopressin. The SFO may induce the sensation of thirst caused by dehydration. 30
Subcommissural organ
The SCO is one of the first brain structures to differentiate in vertebrates.105,109 The SCO is an ancient and conserved structure of the vertebrate brain and, with the exception of a few species (eg, anthropoids and bats), 106 remains fully active throughout life. It is located in the caudodorsal region of the third ventricle, at the entrance of the Sylvian aqueduct (Fig. 1), and is formed in 2 layers. The ependymal cells are highly specialized in the secretion of proteins. The hypendymal cells, situated at the dorsal roof of the entrance of the Sylvian aqueduct, represent the second layer and extend from the rostral to the ventral surface of the posterior commissure and line the roof of this canal as far as to the recessus mesocoelicus. 105 Cell processes of the ependymal and hypendymal cells, containing a secretory material, terminate at the dorsal brain surface adjacent to the subarachnoidal space and on special blood vessels supplying the SCO, which communicate along the perivascular space with the subarachnoidal space. On transverse sections, the organ has different shapes in domestic animals—for example, 2 to 3 folds in dogs, 1 fold in cats and pigs, and 2 folds passing over into a ridge in cattle (Fig. 4). SCO secretory ependymal cells are tall and elongated (Fig. 5). Their apical pole protrudes largely into the ventricular cavity and exhibits only a few kinocilia. 82 They are connected to each other by well-developed zonulae adherentes. There are no neurons in the SCO, but neural innervation of these specialized ependyma has been described. 93
Immunohistochemistry revealed some species variations in the expression of marker molecules. Despite their glial origin, the SCO ependymal cells of cats and rats do not express typical glial markers such as S100 protein or GFAP. However, vimentin, nestin, and cytokeratin expression have been demonstrated. 105 The SCO possesses a dense capillary network without fenestrated walls, unlike other CVOs. The SCO is provided with a double-barrier system, a BBB, and a CSF-SCO barrier.30,105
The specialized ependymal cells of this organ synthesize and secrete high-molecular-mass glycoproteins, including SCO-spondin. The basal secretion of SCO-spondin contributes to the development of the matrix of the posterior commissure. 82 Proteins are also released into the CSF, where they remain soluble 129 or aggregate to form a threadlike supramolecular structure named Reissner’s fiber (RF) 103 that passes through the Sylvian aqueduct and IV ventricle down through the central canal to the filum terminale (Fig. 6). 37 The RF is lacking in adult human beings. 82 This structure is continuously renewed in its cephalic part by addition of secreted material from the SCO. RF material undergoes chemical modifications, disaggregates, and escapes through the dorsal wall of the terminal ventricle to reach local blood vessels. 105
The function of the SCO is unknown. There is evidence that RF facilitates the flow of the cerebrospinal fluid in this narrow aqueduct and is involved in fluid homeostasis. 30 In addition, it has been assumed that the secretory products of the SCO participate in ontogenetic processes in the CNS, such as neuronal differentiation, neuronal aggregation, and axonal pathfinding. 83 Furthermore, RF acts as a pressure mechanoreceptor and cleanser of cerebrospinal fluid 72 because it has the capacity to bind and transport compounds such as dopamine, L-DOPA, and serotonin. 129 There is a close correlation between absence of the SCO and hydrocephalus in rat and mouse strains exhibiting congenital hydrocephalus. 82
Pineal gland
The pineal gland (Fig. 1) develops as a tubular evagination of the diencephalic roof of the neural tube. During phylogeny, the pineal gland develops from a photoreceptive organ into a gland secreting the hormone melatonin (N-acetyl-5-methoxytryptamine) under control of light from the eyes. 104 The pineal gland is an essential organ in the photoneuroendocrine system that contributes substantially to the “inner clock.” In addition, the pineal gland is also of crucial importance for synchronization of gonadal activity and control of reproduction in photoperiodically dependent species. 104 The gland is located at the border between the mesencephalon and the diencephalon of the brain at the posterior wall of the third ventricle just rostrally to the cranial colliculi (Fig. 7). As a part of the epithalamus, it belongs neuroanatomically to the diencephalon. The gland is surrounded by a pial capsule, and in most species, it has a conical or round to oval shape. In rodents, the gland is subdivided into a superficial and a deep part connected by a stalk. 86 The parenchymal cells of most mammalian pineal glands are arranged in cords separated by capillaries. The gland is mainly composed of pinealocytes (Fig. 8) characterized by an ovoid cell shape and moderate cytoplasm with argentophilic cellular processes. 86 Pinealocytes are regarded as paraneurons12,127 and express several retinal antigens 110 and synthesize melatonin and other indolamines that play a crucial role in biorhythm. Besides hormone-producing pinealocytes, interstitial cells, perivascular phagocytes, and peptidergic neuron-like cells can be found. In some species, including man, rabbits, monkeys, ferrets, and cotton rats, typical neurons with Nissl’s substance have been detected. 86 In most species, several but not all of the interstitial cells are immunoreactive for GFAP, resembling fibrillary astrocytes (Fig. 9). 87 In some species, including dogs, 16 horses, 23 and rabbits, 107 cells containing a pigment similar to neuromelanin have been described. In man and ungulates, but not in carnivores, a deposition of granular calcified material composed of calcium and magnesium salts, termed brain sand, acervuli, or corpora arenacea, may be present (Fig. 10) that increases with age and is not associated with a specific disease. 128 In cattle, striated muscle fibers are found within the parenchyma without evidence of their origin or function. 12
The pineal gland is often excluded from the CVOs because its action is more endocrine in nature, and fenestrated endothelia have not been detected in all species. In Mongolian gerbil, Golden hamster, and sheep, the capillaries in the pineal gland are lined by a continuous endothelium. The endothelial cells of the capillaries in the rat pineal gland possess few fenestrations. In the mouse and Djungarian hamster, many fenestrated capillaries are present, 130 similar to other CVOs, but there are also vessels with a morphologically intact blood-brain barrier. 30
Choroid plexus
The CPs, the main producers of CSF, 37 are highly vascularized, leaf-like structures that are attached by thin stalks to the ependymal lining of the brain ventricles and float in the CSF. 28 Four CPs are present, one in each lateral ventricle (CP I and II), one in the third ventricle (dorsal recess; CP III), and one in the fourth ventricle (CP IV; Fig. 1). Each lateral ventricle is connected to the third ventricle through the interventricular foramen. 28 CP IV lies caudal and ventral to the cerebellum on the dorsal side of the brainstem. The CP surface is increased by many protrusions, which appear leaf-like in the lateral ventricles and frond-like in the third and fourth ventricles. The choroid plexus has fenestrated capillaries, but there are tight junctions (zonula occludens) and desmosomes between choroid epithelial cells, thus maintaining a blood-CSF barrier on the apical side. Besides the production of CSF, CP cells also secrete a range of growth factors, neurotrophins, and enzymes into the CSF that eliminate toxic molecules. 102 In addition, CPs are essential for neurite outgrowth and might be a possible source of neural stem cells. 28 CPs thus play a major role in the development, homeostasis, and repair of the central nervous system.31,102
Anatomically, a matrix of loose connective tissue links the vasculature with a monolayer of cuboidal epithelial cells (Fig. 11) sitting on a basement membrane, which are presumed to be of ependymal origin. 1 The epithelial cell surface is increased by numerous microvilli of variable morphology and only few cilia, except during development.
Area postrema (AP)
The AP is the most caudally detectable CVO, located at the caudal end of the brainstem at the junction between the medulla at the level of the calamus scriptorius and the spinal cord slightly lateral to the parasympathetic nucleus of the vagus (Fig. 1). The AP consists of 2 right and left masses attached to the inferior angle of the floor of the fourth ventricle linked together along the median line on the obex at the beginning of the central canal (Fig. 12). In rodents and lagomorphs, the AP is often limited to a median mass attached to the obex. In birds, the AP is highly developed as a long strip of capillaries stretching from the obex on the median line to the lateral angles of the fourth ventricle. 30 Owing to a distinct population of small neurons, the AP is considered a true brainstem nucleus. 85 The AP is covered by a monolayer of flat ependymal cells, 12 and due to its rich vascularity, the AP appears spongy. Blood vessels of the AP consist of large-caliber capillaries forming vascular sinusoids that are anastomosed in a dense network with numerous capillary loops (Fig. 13), which may be confused with granulation tissue. The vessels are devoid of the BBB, because of a fenestrated endothelium and the absence of tight junctions, 85 thus enabling blood-borne substances to reach neurons in the AP directly. Unlike normal ependyma with kinocilia, the specialized ependymal cells of the AP are studded with numerous microvilli that often form tufts. There are specialized ependymal cells resembling tanycytes of the ME, which have long cytoplasmic processes impinging the walls of blood vessels in deeper zones of the AP. 85
Together with the neighboring nucleus of the solitary tract and motor dorsal vagal nucleus, the AP forms the dorsomedial medulla or vagal complex, which is an important area of the autonomic system for cardiovascular and respiratory regulation. The AP might also be the origin of the central emetic reflex, which controls the vomiting center located in the reticular formation of the medulla oblongata. 30 In addition, the AP functions as a central chemoreceptive organ. Its anatomical location allows collection of collecting elements from the circulating blood or CSF and of conveying chemical information to nerve elements coursing through the organ. 85
Hippocampus
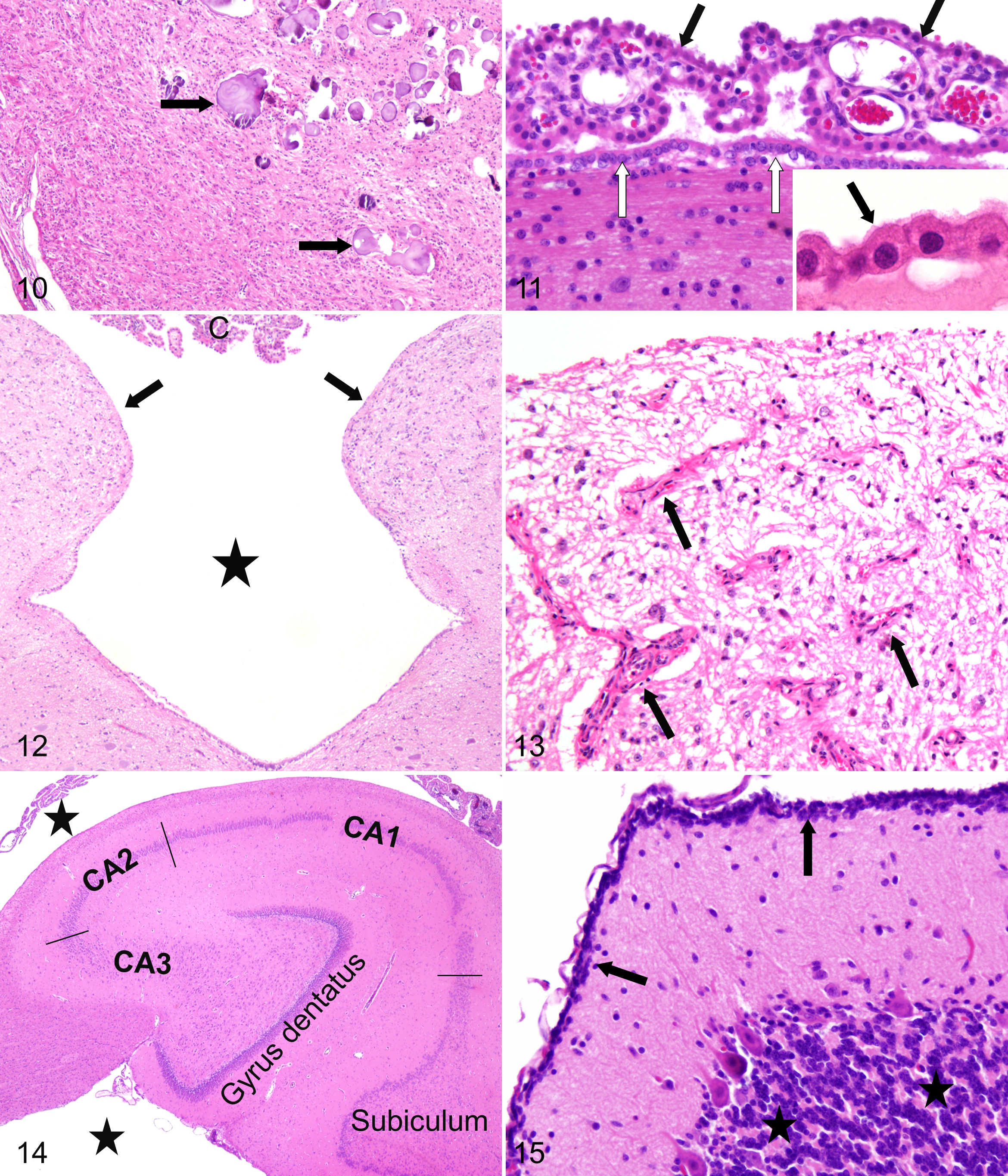
The hippocampal formation is a curved and recurved sheet of the primitive cortex (archipallium), which is located in the medial surface of the temporal lobe, along the floor and medial wall of the temporal horn of the lateral ventricle. The C-shaped formation extends in a curve, starting from the amygdala ventrally in each piriform lobe and progressing caudodorsally and then cranially over the diencephalon. The caudal end of the hippocampal formation tapers under the splenium of the corpus callosum. Dorsal to the caudal thalamus, the hippocampal formation of each cerebral hemisphere meets at the medial plane, and the fornix, the hippocampal commissure, is formed at this region. It is divided into 3 main transverse zones: the dentate gyrus; the hippocampus proper, also termed the hippocampus or cornu ammonis (CA); and the subiculum (Fig. 14). The dentate gyrus and hippocampus proper have a cross-sectional shape resembling 2 interlocking Cs. 131 The subiculum is continuous with one end of the hippocampus proper, and the two together form the shape of the letter S. The subiculum then connects to the entorhinal cortex, forming both the superior and medial aspects of the parahippocampal gyrus. Originally, the hippocampus was subdivided into 4 fields, labeled CA1 to CA4. 76 However, CA4, the deep hilar region, was inconsistently applied to either or both portions of the enclosed pyramidal cell layer and to the cells of the polymorphic layer of the dentate gyrus. However, there is no cytoarchitectonic or connectional reason to distinguish the CA3 pyramidal cells enclosed within the granule cell layer from those that are outside of it, and the use of the confusing term CA4 has been dropped by most authors.3,59 CA1 and subiculum are also termed Sommer’s sector. The main and essential function of the hippocampal formation appears to be essential in the development of short-term declarative memory.
Unusual Cell Types
Cerebellar external germinal cells
During ontogenic development, an actively dividing germinal cell layer remains immediately beyond the pia mater of the cerebellar folia, termed the external germinal layer (Fig. 15), which should not be confused with inflammatory infiltrates. The cells continue to divide and may form a layer up to 12 cells in thickness. Along the inner aspect of this layer, the cells stop dividing and migrate into the cerebellar parenchyma where they differentiate into microneurons, such as basket cells, stellate cells, and granule cells of the inner granular layer of the cerebellum. 135 Therefore, this layer is especially sensitive to X-irradiation, some radiomimetic chemicals, and special viral infections 135 (eg, granule precursor cells located in the cerebellar external granular layer are the major target for Kilham’s rat parvovirus or feline parvovirus replication during the perinatal period).101,111
The external germinal layer has nearly disappeared 20 and 24 days after birth in the mouse and in the rat, respectively. In dogs, external germinal cells will persist as long as 75 days and in the kitten 60 to 84 days after birth, whereas in the calf, they disappear approximately 6 months postnatally. 25
Other CNS progenitor cells (peri- or subventricular cells, subependymal plate, islands of Calleja, hippocampal granule cells)
At different locations, immature round cells (formerly termed spongioblasts), which are remnants of the germinal zone of the primitive neural tube 121 representing neurogenic zones (CNS progenitor cells), can be observed bilaterally and symmetrically in the adult mammalian brain.70,119 In the subgranular zone of the hippocampal dentate gyrus, granule cells are generated, continuously increasing the number of neurons. In the forebrain subventricular zone (SVZ), these cells are also termed peri- or subventricular cells or subependymal plate. They are designated as islands of Calleja when forming distinct island-like clusters (Fig. 16). Clusters of these germinal cells are found at the ventral and lateral aspects of the lateral ventricles and in folds around the mesencephalic duct. On transverse sections of the frontal lobe, occasional clusters of these cells may be found within the white matter. In addition, similar cells are found in the olfactory peduncle and within the olfactory cortex. 121 According to their size, 3 different populations of cells can be recognized: small, intermediate, and large cells (Fig. 17). 9 Many of these immature cells can be immunolabeled with an antibody specific for doublecortin, a microtubule-associated protein expressed by neuronal precursor cells and immature neurons (Figs. 18, 19). 56 Progenitor cells are multipotent for the different neuroectodermal lineages present in the CNS, including the different neuronal and glial subtypes. 115 In addition, CNS progenitors are regarded as the origin for at least some primary brain tumors.98,120
Synantocytes and aldynoglia
Classically, neurons and 3 glial elements, such as astrocytes, oligodendrocytes, and microglia, are recognized in the CNS. NG2 chondroitin sulfate proteoglycan-expressing cells have been proposed to represent a potential fifth element, termed synantocytes. 15 Although these cells are considered oligodendrocyte precursor cells by some, others consider these cells a distinct lineage of CNS cells. These stellate cells display prominent arborization and are related to neurons. Neuronal activity, glutamate, and adenosine triphosphate act on their receptors. It is assumed that synantocytes may be specialized to monitor signals from neurons to glia.
In addition, it is well recognized that several specialized growth-promoting macroglial cells, including radial glia, tanycytes, olfactory ensheathing cells, pituicytes, Bergmann glia, and Müller glia in the retina, can be encountered. These cells types have been collectively called aldynoglia based on their similarity and specialized function.11,49,75 They share characteristics with astrocytes and oligodendrocytes and resemble Schwann cells and are therefore also called central Schwann-like cells. They express commonly the low-affinity neurotrophin receptor p75.57,95 As an example, in dogs with canine distemper virus–induced white matter lesions, bi- to multipolar p75NTR-expressing cells have been identified with Schwann cell–like characteristics (Figs. 20, 21) prior to the onset of demyelination in parallel to axonal damage characterized by the expression of β-amyloid precursor protein. 57
Radial glial cells originate from neuroepithelial cells and are important for the development of the central nervous system. Except for Bergmann glia in the cerebellum (Figs. 22, 23) and Müller glia in the retina, their radial phenotype is only transient. The long radial processes guide the radial migration of newborn neurons from the ventricular zone to the mantle regions or from the cerebellar external germinal layer into the inner granule cell layer. In addition, radial glial cells characterized by astroglial properties are ubiquitous precursor cells that generate neuronal and glial cells, and they play a pivotal role as key elements in patterning and region-specific differentiation of the CNS. 20
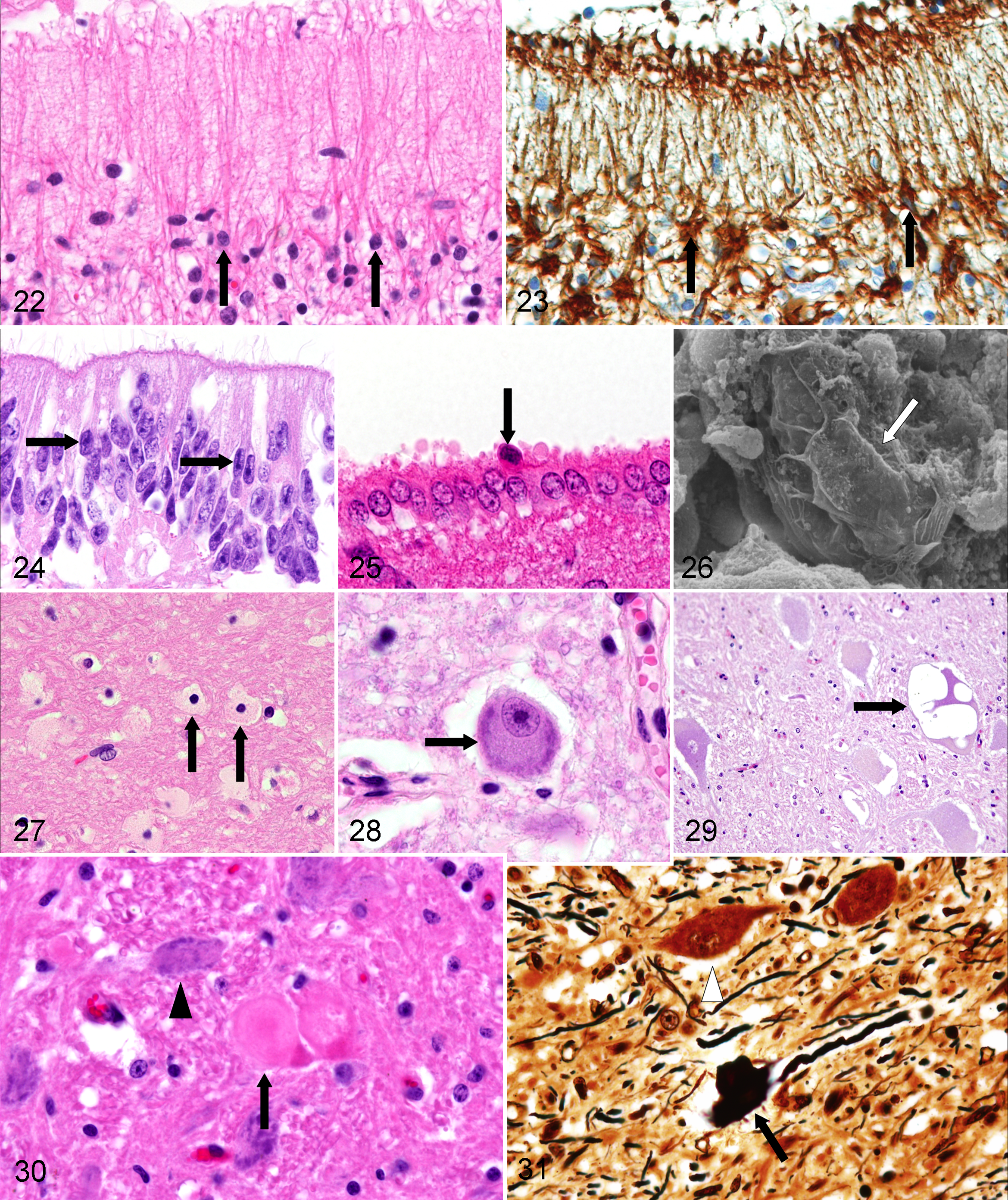
Tanycytes are bipolar specialized ependymal cells located predominantly in the ependymal lining of the floor and ventrolateral walls of the third ventricle. 78 The ventricular surface of tanycytes is in general devoid of kinocilia (Fig. 24). Tanycytes lining the floor of the infundibular recess have long cell processes that project to the vascular surface of the infundibulum (median eminence). In addition, they send processes either to the pial surface or to blood vessels of the mediobasal hypothalamus. The cell processes form bundles and contain high numbers of homogeneously distributed microtubules and, in addition, mitochondria, smooth endoplasmic reticulum, microfilaments, and osmiophilic inclusions. Tanycytes are interconnected with tight junctions, providing a BBB. 10 GFAP has been identified in tanycytes. 134 Derived from light and electron microscopic investigations, tanycytes are supposed to have secretory, absorptive, and transport functions. The latter may be bidirectional (ie, directed to the ventricle and to blood vessels). Tanycytes might be involved in the uptake or delivery of molecules, including infectious agents between the CSF, blood cells, and hypothalamic cells.7,75 They are known to locally modulate the release of gonadotropin-releasing hormone in the median eminence and to display seasonal structural changes in hamsters. 13 Tanycytes may play a role in modulating glutamate concentrations and neurotransmitter supply in the hypothalamic environment. 91
Olfactory ensheathing cells are glial cells that derive from the olfactory placode. The migration of olfactory ensheathing cells is essential for pioneering the olfactory nerve pathway during development by modulating their growth and guidance and for promoting axonal regeneration. They differ from typical glia because they exist in both the peripheral and central nervous system. They share the phenotypes of both astrocytes and Schwann cells. 49
Kolmer cells
On the surface of the choroidal epithelium, some macrophage-like cells are present, called epiplexus or Kolmer cells, first described in amphibians by Kolmer in 1921 and found years later also in mammals. 21 They have a diameter of about 13 to 20 μm, and the nuclei are mostly round, oval, or bilobulated (Fig. 25). The cytoplasm frequently contains tiny vacuoles, 38 and electron microscopy reveals long cellular processes (Fig. 26). Macrophages of monocytic origin infiltrate the stroma of the choroid plexus and enter the ventricle by penetrating through the choroidal epithelial cell (emperipolesis) to reach the epiplexus position. Kolmer cells function as scavenger cells but also play a possible role in immunological responses and iron regulation in the ventricular system or the brain as a whole. 74 Their number can be substantially increased following ventricular infection. 8
Findings of Uncertain or No Significance
Unusual reactive astrocytes in adult cats
An unusual type of hypertrophic astrocyte has been described in adult cats. 43 Histologically, these cells are characterized by an oval to polygonal shape, with a diameter of about 20 μm; short, barely detectable processes; a condensed, hyperchromatic, eccentric nucleus; and abundant, homogeneous, eosinophilic cytoplasm (Fig. 27). Immunophenotyping has revealed an expression of GFAP and S-100 protein, as well as a lack of vimentin. These astrocytes were predominantly located in the white matter of the hindbrain predominantly found in brains with status spongiosus. The occurrence of these cells was less frequently associated with inflammation. In the dentate hilar region, they were found adjacent to degenerated neurons. However, they occur also in brains without other histological changes. The occurrence and function of these cells remain undetermined.
Central chromatolysis
There are some nuclei of the brain, including the olivary nucleus, pontine and supraoptic nuclei, and the lateral cervical nucleus of the spinal cord, in which neurons show central chromatolysis, which is regarded as a normal finding (Fig. 28). 121 In contrast, central chromatolysis in neurons at other localizations as well as peripheral or total chromatolysis represents a significant morphological change that may be present in various CNS conditions, including motor neuron diseases.
Neuronal vacuolation
The presence of cytoplasmic vacuoles in neurons of normal animals has been described in several species. In normal adult cattle, neurons of the red nucleus and the habenular nucleus show frequently cytoplasmic vacuolation as a normal finding.42,48,80 Less commonly, neuronal vacuolation occurs in neurons of the reticular formation or other brainstem nuclei of adult cattle (Figs. 29).48,80 Similarly, neurons of the medulla in sheep display cytoplasmic vacuoles. 121 These incidental neuronal vacuolations in ruminants should not be confused with changes characteristic of spongiform encephalopathies, which also show vacuolation of the neuropil and immunoreactivity for PrPsc. In raccoons (Procyon lotor), neuronal vacuolation has been described in various brainstem nuclei and cerebral neurons without obvious clinical disease. 51
Spheroids
Focal swellings of axons are commonly referred to as spheroids. Particularly in old horses, but even in young individuals, few axonal spheroids, sometimes vacuolated, are often present in the lateral cuneate nucleus of the medulla (Figs. 30, 31). 121 In addition, spheroids are found most frequently in the vestibular nuclei of clinically normal adult cattle. 42 In normal sheep, spheroids are encountered often as an incidental finding in the dorsolateral medulla, particularly the cuneate and gracile nuclei. 55 Axonal spheroids occur also at various sites of the brain and spinal cord of normal dogs, including the cochlear nucleus, 117 gracile nucleus,90,125 cervical spinal cord, 90 and conus medullaris.121,125 Degenerated neurons have to be considered the main differential diagnosis. The occurrence of spheroids in large numbers is indicative for axonopathy or axonal dystrophy.
Spongiosis
Spongiosis or vacuolar degeneration includes vacuolar changes within processes of the neuropil and vesiculation of myelin sheaths. 121 Focal spongiosis has been observed in the white matter of brains originating from adult cattle.62,80 Principally, intramyelinic vacuoles may involve the white matter of the substantia nigra in the rostral mesencephalon (Figs. 32, 33). An association with a clinical syndrome has not been excluded but still lacks confirmation. Differential diagnosis includes artifactual vacuolation; myelin edema; spongiform myelinopathies; spongiform neuronal changes inflicted by viruses, prions, or toxins; metabolic derangements; or lysosomal storage diseases. In single cases, it may be difficult to discriminate between early significant and nonsignificant changes.
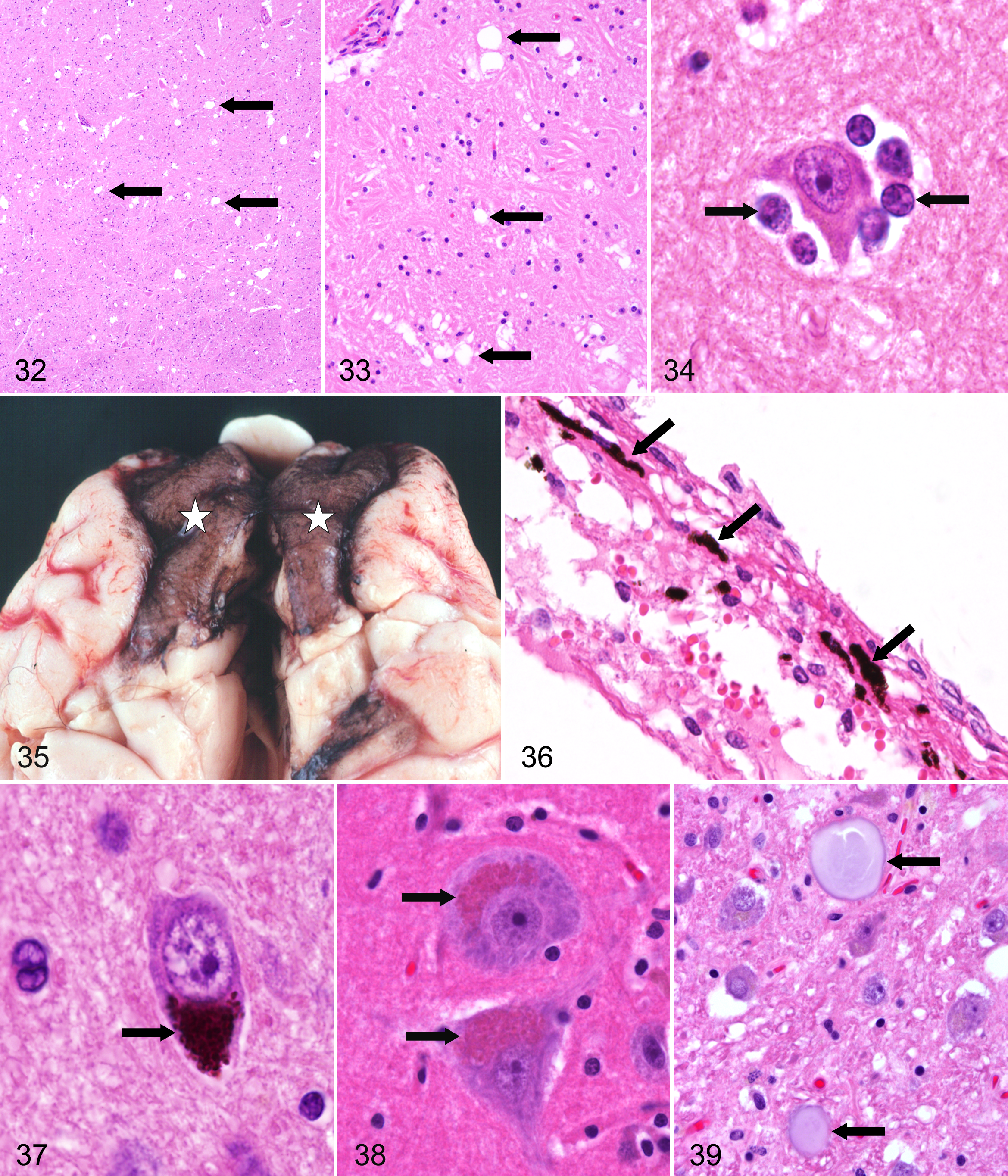
Satellitosis
Perineuronal satellite cells are mostly oligodendrocytes that have an enigmatic affinity to this location 45 and are in contact with the perikaryon of neurons. They are located within the “satellite space,” that is, a shell of 6 μm width around neuronal perikarya (Fig. 34). 92 However, perineuronal cells also may be of astroglial and microglial origin. 121 The number of these perineuronal or satellite oligodendroglial cells varies in different areas of the brain and between species. Usually, they are very common in spinal and trigeminal ganglia and are regarded as a normal finding and should not be confused with lymphocytes. 121 It is assumed that satellite cells contribute to the metabolic function of neuronal homeostasis. However, satellitosis may also occur secondary to neuronal degeneration and may precede neuronophagia, which is characterized by removal of fragments of necrotic neurons by macrophages.
Melanosis
Pigmented melanocytes are regularly found scattered in the leptomeninx of sheep and cattle, particularly in the frontal lobes (Figs. 35, 36) and the spinal cord, the stroma of the choroid plexus, and adjacent to blood vessels within the neuroparenchyma. 32 Particularly in sheep, melanocytes may occur in huge numbers, mimicking neoplastic growth. However, they occur only in the meninges and do not extend into the parenchyma. In other domestic animal species, melanosis of meninges and other CNS structures is rare. 32 Leptomeningeal melanocytes originate from the neural crest elements.
Neuromelanin
Neuromelanin is structurally and biochemically different from melanin that is confined to melanocytes found only in the leptomeninges. Neuromelanin represents an organic polymer of dopamine metabolic products. Compared with lipofuscin, neuromelanin is much darker in hematoxylin and eosin (HE)–stained sections (Fig. 37), is strongly argentophilic, and lacks autofluorescence. 10 Neuromelanic pigment is particularly present in neurons of the substantia nigra. 27 Significant species variability has been observed in the content of neuromelanin in neurons of the substantia nigra, with increasing amounts reported in cats, dogs, primates, and humans, in which it can be even observed macroscopically. 27 Pigment deposition in neurons has to be differentiated from various storage diseases.
Lipofuscin
In neuronal perikarya, an accumulation of golden brown to reddish, fine granular, autofluorescent pigment occurs with aging in all domestic animals. 46 However, not all populations of neurons are similarly affected. Lipofuscin usually does not accumulate in canine Purkinje cells and neurons of the dorsal motor nucleus of the vagal nerve, whereas neurons of the hypoglossal and oculomotor nuclei consistently show pigment accumulation (Fig. 38). 121 Lipofuscin has to be distinguished from neuromelanin and other storage disorder material.
Polyglucosan bodies (corpora amylacea, Lafora bodies)
Polyglucosan bodies (PGBs) are also called corpora amylacea, Lafora or Lafora-like bodies, Bielschowsky’s bodies, or amylopectin bodies, depending on tissue distribution and on clinical preference in human medicine. They share some common histological, histochemical, and ultrastructural characteristics. 44 In domestic animals, Lafora disease occurs in dogs, causing progressive myoclonus. 126 This autosomal recessive genetic disorder in Beagle dogs, Bassets, and Poodles is characterized by accumulation of homogeneous basophilic, periodic-acid Schiff (PAS)–positive inclusions in the perikaryon of neurons, termed Lafora bodies (LBs), a form of PGBs. Similar neurologic disorders have been reported, although less frequently, in cats,50,124 cattle, 116 and cockatiels. 14 In addition, polyglucosan bodies or Lafora-like bodies have been described as incidental cytoplasmic finding in the central nervous system of neurologically normal dogs, 123 cats,67, 68 cattle, 116 and some other species, including a Japanese red fox. 69 PGBs accumulate in the CNS also with aging, and then they are termed corpora amylacea (Fig. 39). 22 They occur commonly with aging (>8 years) 22 but may be observed also in young individuals. 50 They may be preferentially located subpially, perivasculary, and beneath the ependymal lining. Histochemically, corpora amylacea staining is similar to Lafora bodies, indicating a composition of carbohydrates and small amounts of acid mucopolysaccharides. Bielschowsky’s silver stain results in intense coloration. Ultrastructurally, the cytoplasmic accumulations consist of non-membrane-bound, filamentous, interwoven material of 9 nm in diameter associated with fine and coarse granular material. 77 Canine corpora amylacea exhibit positive staining by immunohistochemistry with antibodies against human PGBs 68 and neurofilaments, 47 suggesting a mixed neuronal-glial origin. 77 In dogs with neurologic signs, Lafora-like PGBs located in the hypothalamus and cerebellum are described. They exhibit a lower alcohol-resistant metachromasia than corpora amylacea when stained with toluidine blue and display differences in the expression of GFAP. 77 As differential diagnosis, PGBs have to be discriminated from Buscaino bodies, edema, or storage disorders.
Psammoma bodies
Psammoma bodies are spherical structures, 10 to 50 μm in diameter, consisting of a hyaline matrix with lamellated calcification (Fig. 40). Matrix granules originating from whorled arachnoidal cells serve as the principal calcification nidi. 136 Most commonly, they are located at the spinal rootlets, and their occurrence increases with age. Their significance as a disease indicator is not known. They presumably represent an incidental finding without disease-related significance. However, they are encountered often in meningiomas, particularly in the psammomatous subtype. As differential diagnoses, ossifying pachymeningitis, metastatic vascular calcification, disease-associated dystrophic calcifications, and artifactual dislocation of mineralized tissue fragments have to be considered.
Vascular Filling and Amount of Liquor
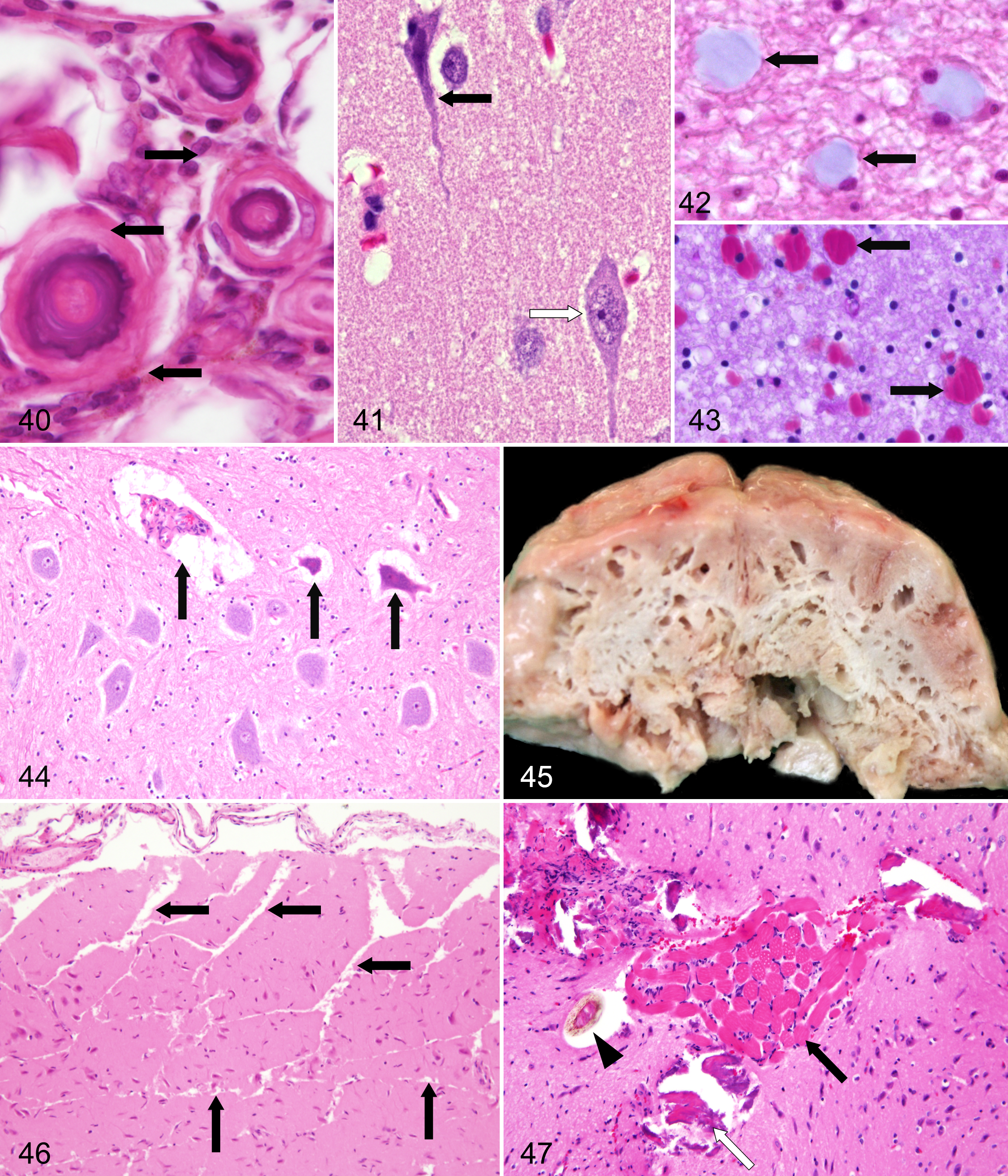
The blood content of blood vessels of the brain and meninges has to be evaluated with particular care. Numerous parameters influence the filling status of blood vessels, including agony, intravital circulatory situation, postmortem positioning of the head, withdrawal of blood, time of separation of the head from the body, and removal of the brain. 32 A similar situation is encountered in the assessment of the amount of liquor. 32 After successful perfusion, CNS vessels lack blood cells, and the perivascular space is no longer visible. Similarly, both aforementioned parameters are useful in evaluating whether perfusion was performed properly.
Avoiding Artifacts
Similar to other organs, the CNS is susceptible to artifacts that might be confused with genuine neuropathological changes. There is a particular propensity for histological artifacts to occur in nervous system tissue samples unless scrupulous attention is paid to all stages of tissue processing (ie, dissection, sampling, fixation, processing, sectioning, and staining).65,66 To avoid artifacts, perfusion fixation is recommended, although this method is not applicable for routine diagnostic pathology. Nevertheless, the CNS should be collected and fixed by free-floating immersion into 10% buffered formalin as soon as possible after death. The formalin should be renewed after a couple of days, especially for brains of large animals. The spinal dura mater should be opened longitudinally to facilitate proper fixation. Special care has to be taken regarding the removal of the CNS out of its bony shell. Gentle handling and avoidance of kinking the spinal cord may are highly recommended to reduce postmortem mechanical damage. Instruments used to open the cranium should never impact the brain, and the dura mater must be carefully removed prior to any attempts to remove the brain. Cranial nerves should be severed with a sharp blade and not pulled from their attachments. 64 In histologically questionable cases of whether a tissue change is genuine or an artifact, one should consider the history and clinical course of the patient’s illness as well as the type and extent of neuropathological findings. Reliable hints for intravital damage include hypertrophy of vascular endothelial cells, infiltration of neutrophilic granulocytes, macrophages within myelin digestion chambers, or (astro)gliosis. The latter could be confirmed by using immunohistochemistry for GFAP and vimentin to detect activated or immature astrocytes as indicators of an intravital process. In addition, immunohistochemical examination may help to assess genuine changes (eg, β-amyloid precursor protein expression in damaged axons may occur as early as 1 hour after intravital injury). 61
Dark neurons
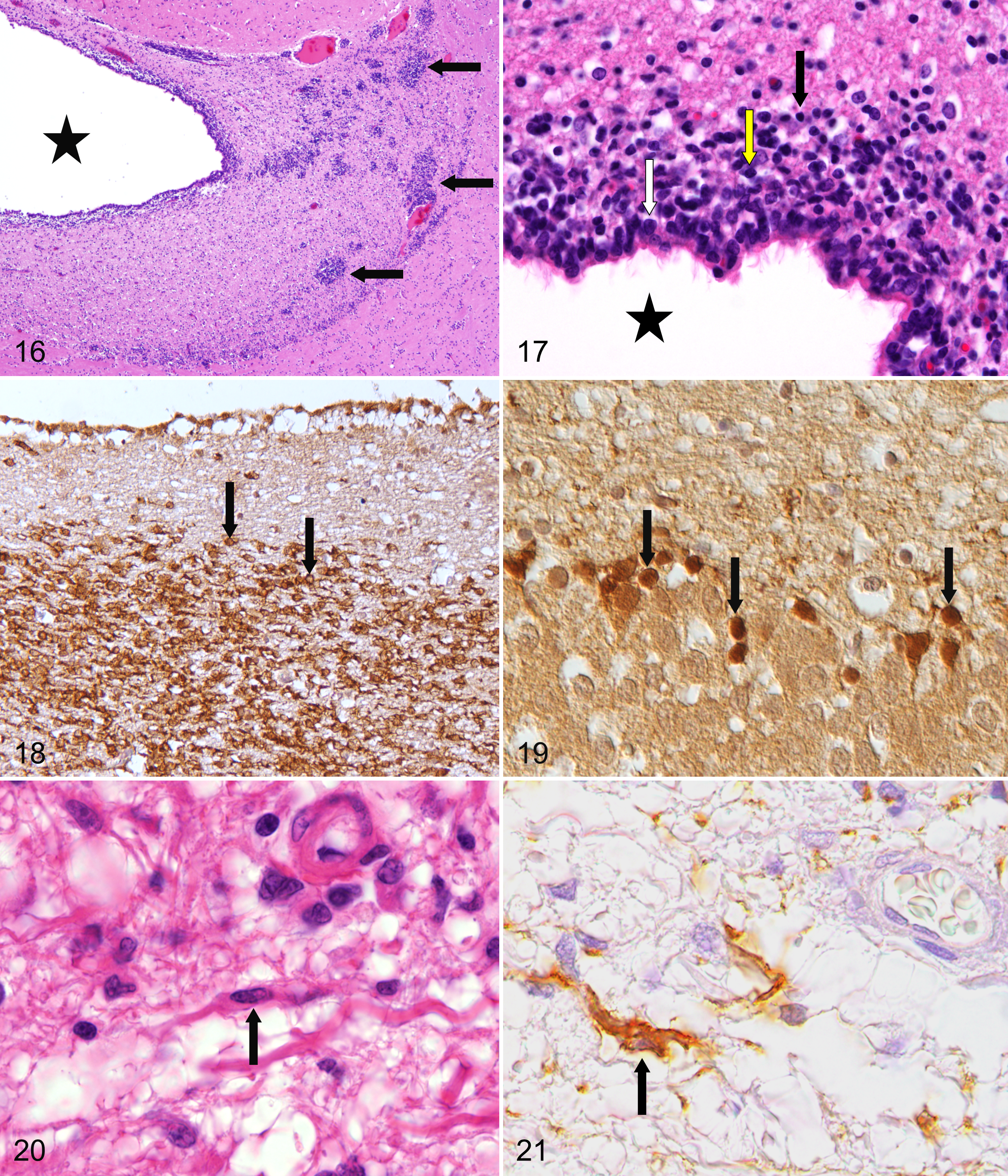
Dark neurons represent a highly interesting and in part controversial topic. In neuropathology, at least 3 types of “dark” neurons—reversible, irreversible, and artifactual—are recognized. They exhibit a similar histological appearance. 45 The artifactual dark neuron may occur scattered throughout the brain and spinal cord and often shows irregular, corkscrew-shaped apical dendrites. The shrunken, dark-stained nucleus may be indistinct within the cell body because it blends into the compacted perikaryal cytoplasm (Fig. 41). At times, slight eosinophilia may be superimposed, 121 giving a dark blue–red tint using HE staining. Affected neurons may be separated from adjacent neuropil, especially within paraffin-embedded tissue, 19 and are often found scattered among histologically intact neurons. It is generally agreed that dark neurons are caused by postmortem trauma or distortion of the unfixed CNS.17,18,32,45,65,66 The “dark” neuron artifact is produced either prior to fixation or even following inadequate perfusion-fixation of the brain as a consequence of regional differences in intensity and speed of fixation, inducing dark neurons in the transitional zones where incompletely fixed neurons are located. 19 The dark neuron change has been readily produced in animals such as pigeons, mice, rats, guinea pigs, squirrels, rabbits, cats, dogs, and monkeys. 66 To avoid dark neurons, perfusion fixation with a delay of several hours prior to removal is recommended.39,40 It is not difficult to recognize a single or a few dark neurons in an otherwise normal-appearing brain, 121 but several reports seem to misinterpret artifactual dark neurons as a genuine lesion. 66 The dark neuron change is clearly related to a contraction process in a perturbed neuron at the time of fixation. 5 However, the biochemical process behind it is not fully defined. Some possible explanations of the neuronal contraction are a postmortem mechanically induced tissue depolarization, glucose deprivation, disruption of peri- and intraneuronal attachments allowing shrinkage during fixation and subsequent dehydration for paraffin embedding, and reduced cytoplasmic osmolarity.5,17,19
However, the occurrence of similarly contracted, darkly stained neurons, occasionally with corkscrew-shaped apical dendrites, in some acute experimental neuropathologic disorders has to be appreciated. A special set of morphological changes characterizes these dark neurons: (1) massive shrinkage, hyperbasophilia, argyrophilia, and an increased electron density; (2) marked compaction of the ultrastructural elements; and (3) aggregation of the nuclear chromatin with a pattern other than that characteristic of apoptosis. 79 These disorders include experimentally induced hypoglycemia, status epilepticus, ischemia, ischemia reperfusion, physical forces such as head injuries, or an electric shock.4,34,35,58,63,97,118 These acute lesions can be differentiated from the dark neuron artifact because distinct neuropathologic changes do not occur in the artifact, including edema in affected regions of the brain reflected by astrocytic swelling, leading to a status spongiosus. 66 In addition, these contracted cells are part of a spectrum of neuronal changes in these acute injuries. Numbers of dark neurons appear to evolve as recognized forms of neuronal necrosis such as acidophilic change. This is in contrast to the monotonous cytological pattern seen in the postmortem mechanically induced artifact. 66 Following compaction, such neurons undergo additional morphological changes, depending on changes within their environment. 36 Independently of the nature of the initiating condition, a proportion of “dark” neurons recover, whereas others die.4,24 However, neuronal death occurs neither by necrosis nor by apoptosis. 36 The debris of these “dark” neurons is removed from an otherwise undamaged environment mainly by astrocytes. Neuronal fragments are transferred to phagocytotic pericytes or discharged into blood vessels. 79 There are special methods available for the identification of acutely degenerating neurons, including autofluorescence in HE-stained slides or after Fluoro Jade B–staining.64,108 In summary, clear differences between neuronal contraction in these acute energy deprivation states and the dark neuron artifact can be ascertained by careful neuropathologic evaluation.
Buscaino bodies (mucocytes, metachromatic bodies, Grynfeltt bodies)
Buscaino bodies or “mucocytes” represent glassy, pale, gray-blue, metachromatic, often PAS-positive, rounded, oval, or lobulated, mucin-like material of approximately 100 μm in diameter that are usually dispersed in the neuropil but preferentially located in the white matter (Figs. 42, 43).33,39,45,46 The exact nature and development of these structures are not known, but they are thought to represent alterations of myelin fixation with solubilization and subsequent precipitation, as well as the handling of nervous tissues too soon after perfusion (ie, before the chemical process of fixation has been completed).33,39 Buscaino bodies have to be differentiated from corpora amylacea by neuroanatomical distribution and at least partial birefringence under polarized light, edema, or material accumulated in storage disorders.
Shrinkage artifacts
Retraction spaces are commonly observed around neurons, glial cells, and blood vessels and are caused by shrinkage (Fig. 44).33,39 These artifactual changes are often due to insufficient dehydration and inadequate impregnation with paraffin wax upon embedding. 45 Perivascular retraction spaces have to be differentiated from perivascular or astrocytic edema. Pericellular retraction spaces with cellular dropouts may mimic spongiosis.
Vacuolization
Saprophytic gas-producing bacteria give rise to postmortem grossly visible cyst formations (“Swiss cheese”) (Fig. 45). 32 This postmortem change has to be differentiated from gas bubbles in various tissues, including brain of marine mammals entangled and drowned at depth in gillnets. 88
Neuropil vacuolization is a disturbing but reproducible histotechnological artifact. 133 Besides freezing and autolysis, high concentrations of alcohol or prolonged exposure (eg, 48 hours) to alcohol of 70% may cause artifactual vacuolization of the neuropil, particularly of white matter.41,133 This artifactual vacuolization has to be differentiated from microcavitation associated with disease (eg, certain toxic disorders and altered metabolic states in white matter 53 and in transmissible gray matter encephalopathies) because light microscopically, the vacuoles appear similar. 100
Freezing artifacts, soft tissue fragments, mineralized debris, and leptomeningeal emphysema
Clefts and linear fissures within the nervous tissue after freezing represent an artifactual distortion resulting from formation of ice crystals (Fig. 46). Although pathologists should be aware of the effects of freezing, new technical staff or submitting veterinarians may not always understand the difference between preserving a carcass in the refrigerator and placing it in the freezer. 64
Soft tissue and bone fragments may be dislocated into the central nervous tissue (Fig. 47) when using electric or manual saws during the opening of the cranium to remove the brain. 39 In particular mineralized debris has to be differentiated from psammoma bodies.
Removal of the spinal cord has to be performed carefully with minimal kinking before fixation because otherwise, a displacement of the neuropil may simulate the appearance of tumors or developmental abnormalities. 39 Forcible removal of the skull may cause distention and artifactual emphysema of the leptomeninx (Figs. 48, 49) as well as separation of brain structures (eg, molecular layer from Purkinje cell and granule cell layer of the cerebellum), particularly in large animals (Fig. 50).
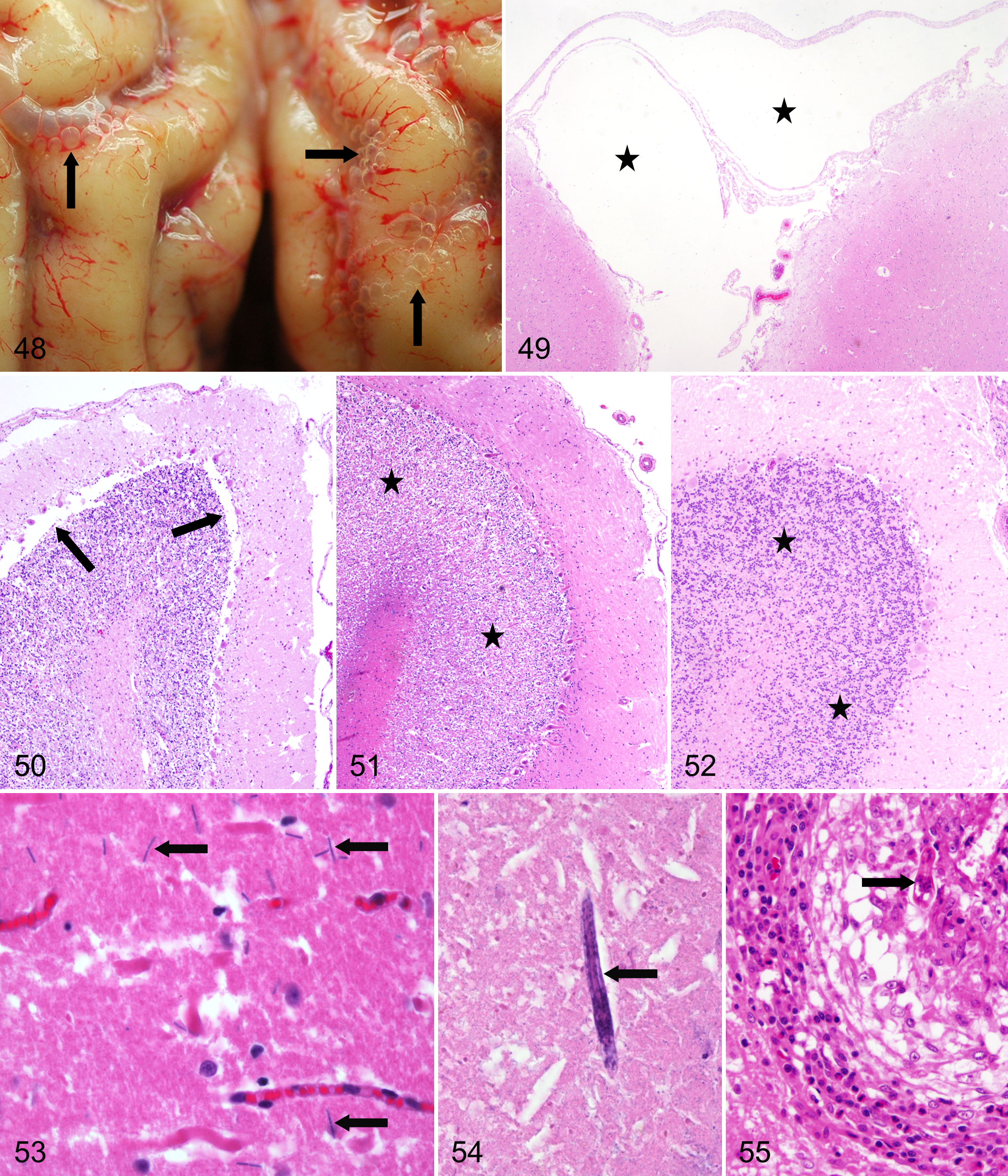
Postmortem Changes of Cells and Neuroparenchyma
Cerebellar conglutination
The cerebellar granular cell layer of cattle and other bovidae is particularly susceptible to postmortem lysis, whereas the Purkinje cells remain relatively well preserved. This postmortem change is also known as cerebellar conglutination (Figs. 51, 52). In autolytic CNS tissue, glial cells appear small and hyperchromatic, resulting in a seemingly much higher number of oligodendrocytes than normal. 121
Hemolysis
Hemolysis allows diffusion of hemoglobin through permeable blood vessels into adjacent neuroparenchyma, resulting in a spotted red appearance. As differential diagnosis, intravital hemorrhages have to be considered. Additional handling-related displacements of gray and white matter may result in a “toothpaste” appearance of the spinal cord. 32
Bacteria, fungi, and nematodes
Advanced postmortem decomposition allows bacteria, fungi, and nematodes to invade the CNS. Bacteria (Fig. 53) and fungi usually reach the CNS via blood vessels and have to be differentiated from intravital embolic-metastatic changes, which are accompanied by vital reactions (eg, vascular endothelial hypertrophy, infiltration of inflammatory cells, and activation of glial cells). Earth nematode invasion of the CNS occurs in buried animals and must be distinguished, especially in forensic cases, from intravital infestation of migrating larvae (eg, Toxocara sp or Baylisascaris procyonis) by the lack of cellular inflammatory response (Figs. 54, 55). They may enter the cranial cavity via natural openings, such as the canales nervi optici or the foramen occipitale magnum, or via acquired fracture clefts of the skull.
Concluding Remarks
We hope that the presented information, together with the provided images, will assist pathologists in becoming more familiar with normal architecture, nonlesions, unusual cell types, artifacts, and postmortem changes in the CNS. Working with the nervous system requires in-depth knowledge of neuroanatomic and neurohistologic architecture as well as some skills about technical procedures for collecting the CNS tissue and dissecting the area of interest without causing too many artifacts to ensure that high-quality material for interpretation is obtained. Overall, the myth surrounding findings and interpretation of CNS tissue-specific reaction patterns will be best resolved not by avoiding CNS tissue analysis but instead by continuously tackling and investigating cases of nervous disorders as for any other organ system. This will ensure familiarization with neuromorphological and neuropathological findings and becoming confident with neuropathological descriptions, interpretations, and formulating diagnoses. It is recommended that less experienced pathologists should use standard text books of neuroanatomy and neurohistology for orientation, together with a good review article or textbook about CNS disease for data interpretation. Sometimes this should be done before, during, and after reading slides. Even experienced neuropathologists have to rely on such support. This should not be interpreted as a weakness; especially in the CNS, such aids are required to enable a professional interpretation and diagnosis.
Footnotes
Acknowledgement
The drawing skills of Mrs Kerstin Schöne for the generation of Fig. 1 are gratefully acknowledged. We thank Dr M. von Keyserlingk, State Veterinary Institute, Hannover, Germany, for providing the ![]() .
.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.