The use of induced and spontaneous mutant mice and genetically engineered mice (and
combinations thereof) to study cancers and other aging phenotypes to advance improved
functional human life spans will involve studies of aging mice. Genetic background
contributes to pathology phenotypes and to causes of death as well as to longevity.
Increased recognition of expected phenotypes, experimental variables that influence
phenotypes and research outcomes, and experimental design options and rationales can
maximize the utility of genetically engineered mice (GEM) models to translational research
on aging. This review aims to provide resources to enhance the design and practice of
chronic and longevity studies involving GEM. C57BL6, 129, and FVB/N strains are emphasized
because of their widespread use in the generation of knockout, transgenic, and conditional
mutant GEM. Resources are included also for pathology of other inbred strain families,
including A, AKR, BALB/c, C3H, C57L, C58, CBA, DBA, GR, NOD.scid, SAMP, and SJL/J, and
non-inbred mice, including 4WC, AB6F1, Ames dwarf, B6, 129, B6C3F1, BALB/c,129, Het3,
nude, SENCAR, and several Swiss stocks. Experimental strategies for long-term
cross-sectional and longitudinal studies to assess causes of or contributors to death,
disease burden, spectrum of pathology phenotypes, longevity, and functional healthy life
spans (health spans) are compared and discussed.
Genetically modified organisms, especially genetically engineered mice (GEM), are used in the
study of genetic and molecular mechanisms of disease, often with the aim of developing
treatments and interventions to improve health and longevity. A PubMed search (March 11, 2011)
using the keywords transgenic OR knockout AND
mice yielded 137 809 references, with “transgenic” publications beginning
in 1982 and “knockout” publications beginning in 1985, although the term was not widely used
for targeted mutant mice until about 1990. More than half of the citations (72 075) were
published in the past 5 years, 2005–2010. In addition to transgenic and knockout mice,
conditional mutants, and gene traps, mouse research tools include spontaneous mutants, mutants
generated by chemical or physical mutagenesis, inbred strains, recombinant inbred strains,
outbred stocks, collaborative cross-strains, and combinations of these. The International
Knockout Mouse Consortium (IKMC; http://www.knockoutmouse.org/) has
generated embryonic stem (ES) cell lines for more than 16 000 genes since 2008, in its aim to
mutate all protein-coding genes in the mouse by gene trapping and gene targeting in C57BL/6N
ES cells. The IKMC further aims to develop GEM for every protein-coding gene, to make ES cells
and live mice (GEM) publicly accessible through repositories, and to implement broad-based,
systematic phenotyping of these GEM in the International Mouse Phenotyping Consortium (IMPC;
http://www.mousephenotype.org/) by 2020. These coordinated international efforts
are expected to further improve the availability and the utility of GEM research tools to
translational science.1,80,160,192,248 In addition to this international “big team” science initiative, mouse research tools
involving other genetic backgrounds continue to be developed and applied in diverse research
settings. Their rational and optimal implementation requires increased understanding of
expected phenotypes and relevant experimental variables.20,106,211 The Collaborative Cross is another multinational effort to develop mouse genetic tools.
Developed by the “complex trait consortium” to model the complexity of the human genome, this
international effort is developing a mouse reference population with high allelic diversity
generated from a randomized breeding design that outcrossed 8 founder strains, then inbred F2
offspring to obtain new recombinant inbred strains.51,177,222 Pathology data from these recombinant inbred strains are not yet published so are not
included here.
Genetic Background: Expected Pathology Phenotypes and Causes or Contributors to
Death
Since the 1930s, inbred mouse strains have been valued as genetic tools in
translational science.27,28,99,106,132 Differences in life spans and contributors to mortality are expected.174C57BL/6, 129, and FVB/N strains are emphasized here because of their current relevance
and ubiquity in the backgrounds of genetically engineered mice.
C57BL/6 mice are among the most common mice reported in the scientific
literature. C57BL/6N substrains derive from C57BL/6J mice from The Jackson Laboratory (J)
that were sent to National Institutes of Health (N) in 1951.169 Genetic polymorphisms among the substrains have been identified and may be relevant
to some studies.255 C57BL/6J was selected for the initial mouse genome project.239 C57BL/6N mice and ES cells were selected by the IKMC, such that C57BL/6N-based GEM
will be assessed in IMPC phenotyping.55 Various C57BL/6 substrains have been used widely in the development of knockout GEM,
especially as blastocyst recipients of ES cells in targeted mutagenesis.196
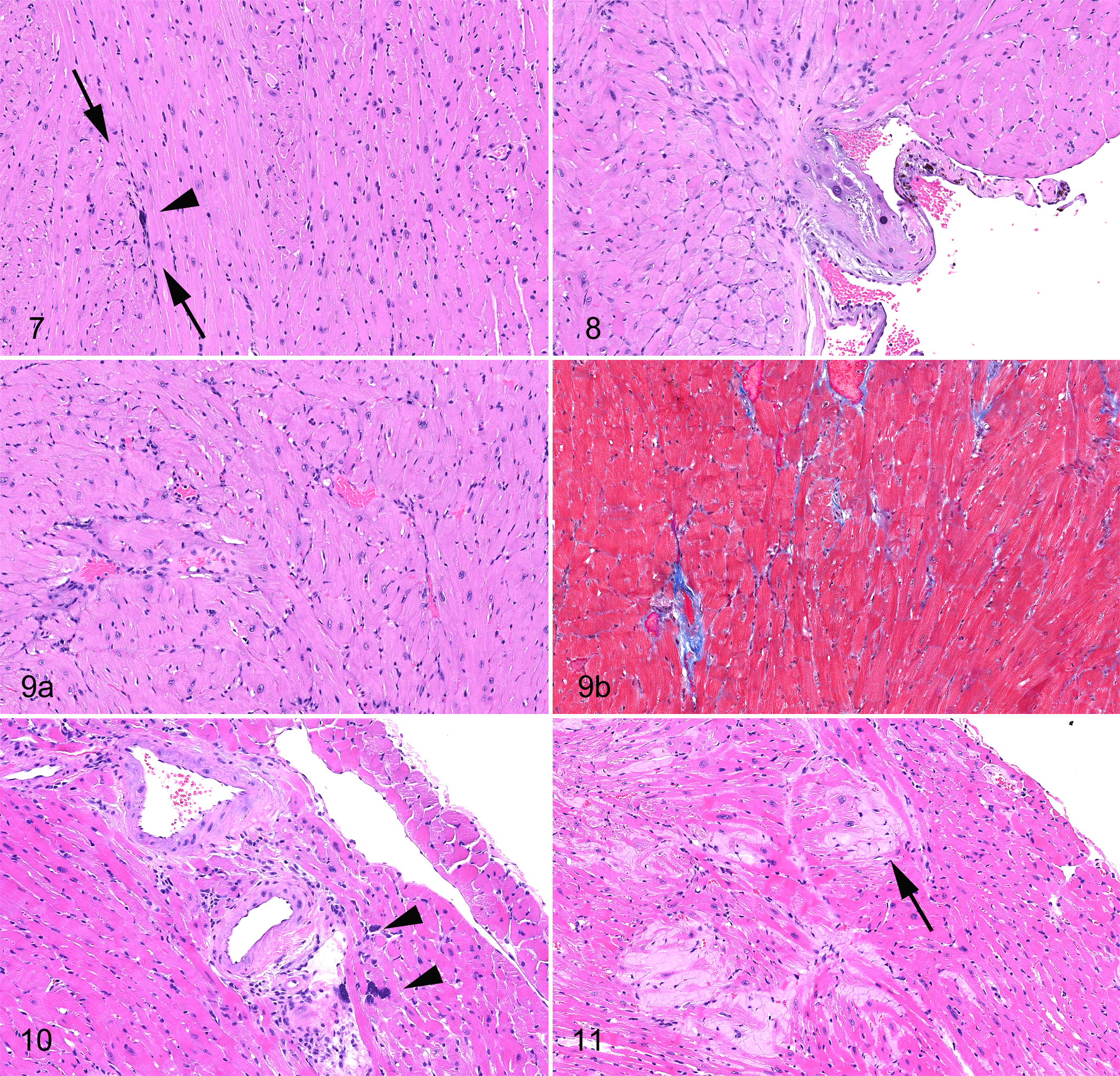
The most common causes and contributors to death in recent studies of aging C57BL/6N or
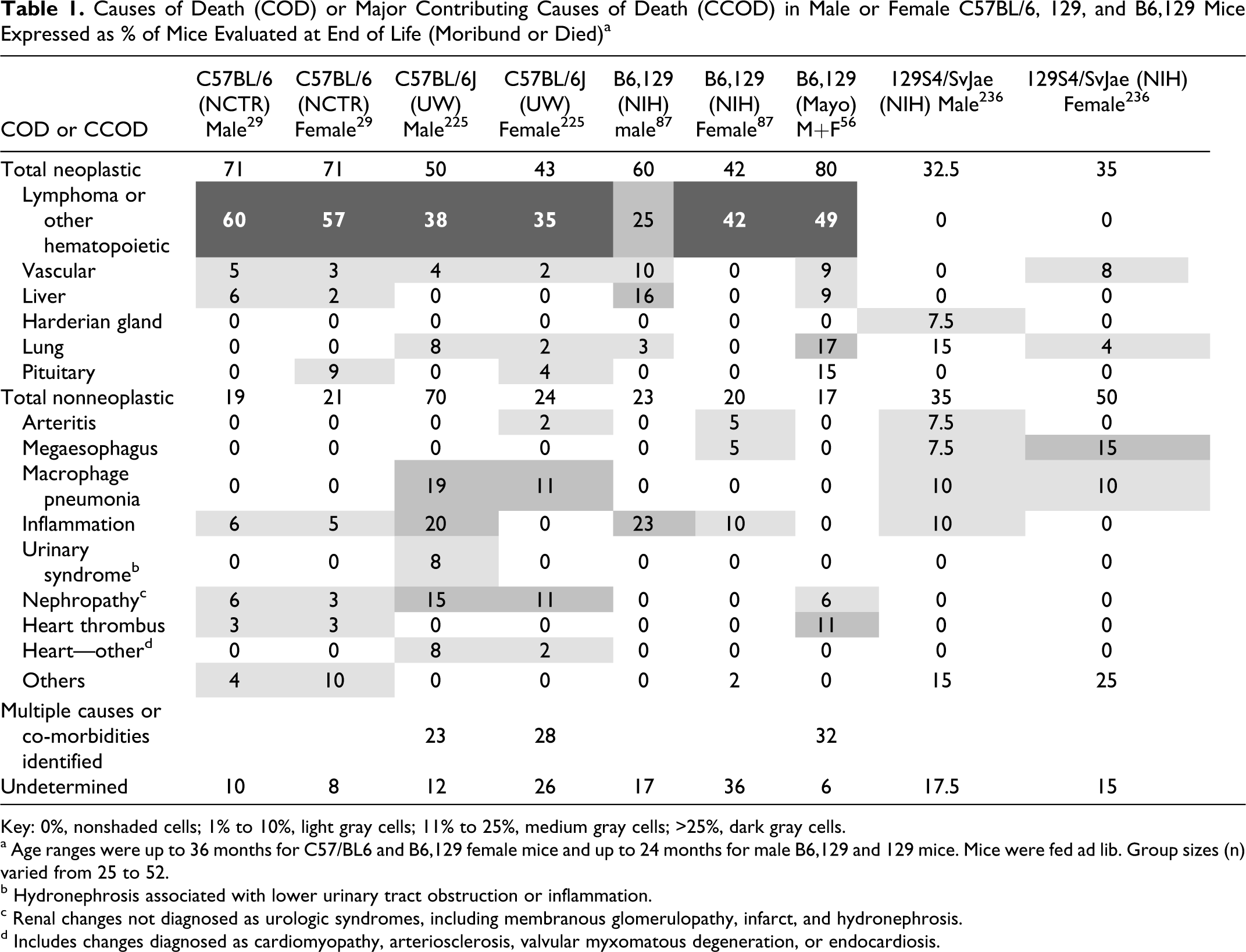
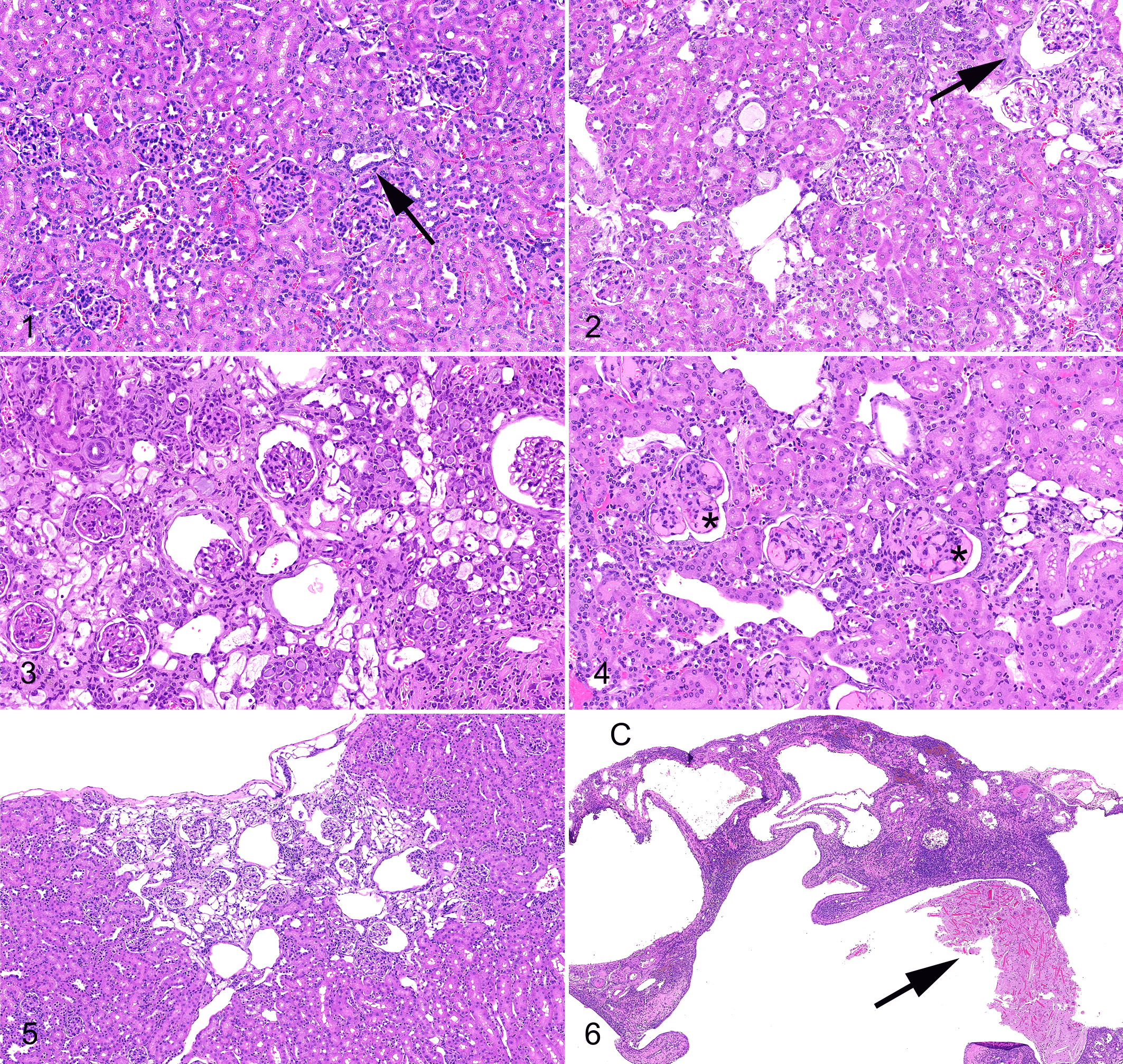
C57BL/6J mice are lymphoma and hematopoietic neoplasms in general (Table 1 ).11,29 Vascular neoplasms, acidophilic macrophage pneumonia, nephropathy (Figures 1–6), inflammatory lesions
(abscesses, etc), and urinary obstructive syndrome in males are potentially significant
contributors. Cardiac changes can be common in some studies of aging C57BL/6 mice but are
not a major cause of death. Figures
1–6 illustrate some of kidney findings, and figures 7-11 illustrate some of heart findings in
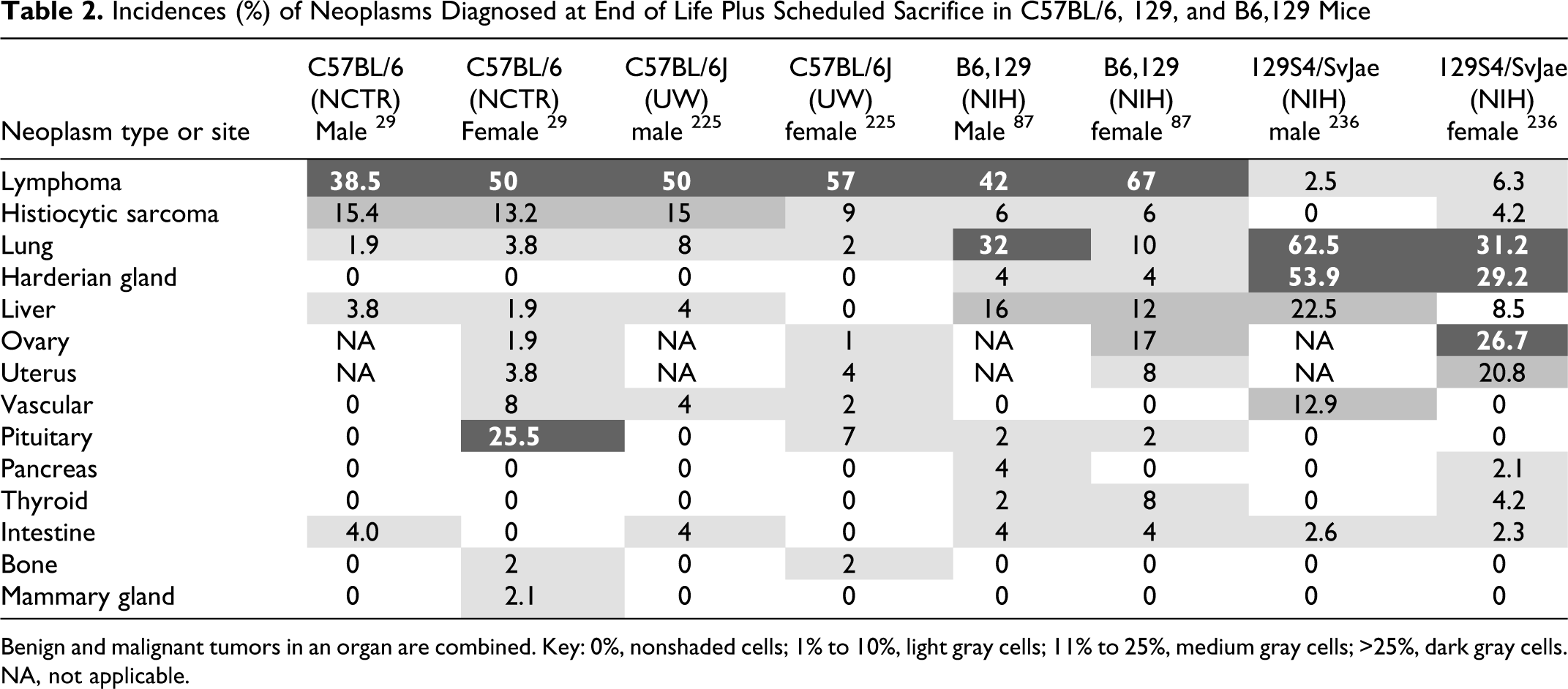
C57BL/6 mice in chronic studies. Neoplasms diagnosed at the end of life and at scheduled
sacrifice (Table 2 ) include
tumors that were not considered to have contributed significantly to death but illustrate
the spectrum of neoplastic findings.
Causes of Death (COD) or Major Contributing Causes of Death (CCOD) in Male or Female
C57BL/6, 129, and B6,129 Mice Expressed as % of Mice Evaluated at End of Life (Moribund
or Died)a
Key: 0%, nonshaded cells; 1% to 10%, light gray cells; 11% to 25%, medium gray cells;
>25%, dark gray cells.
a Age ranges were up to 36 months for C57/BL6 and B6,129 female mice and up
to 24 months for male B6,129 and 129 mice. Mice were fed ad lib. Group sizes (n)
varied from 25 to 52.
b Hydronephrosis associated with lower urinary tract obstruction or
inflammation.
c Renal changes not diagnosed as urologic syndromes, including membranous
glomerulopathy, infarct, and hydronephrosis.
d Includes changes diagnosed as cardiomyopathy, arteriosclerosis, valvular
myxomatous degeneration, or endocardiosis.
Figure 1. Kidney; aged >24 months, C57BL/6J mouse. Mild (grade 2)
nephropathy. Tubules are multifocally basophilic and dilated with rare intralumenal
hyaline to proteinaceous casts (arrow). Hematoxylin and eosin (HE).
Figure 2. Kidney; aged >24 months, C57BL/6J mouse. Moderate nephropathy
(grade 3). There is renal tubular ectasia with epithelial basophilia or attenuation and
intralumenal proteinaceous casts. Glomeruli are multifocally hypersegmented with mildly
increased mesangial matrix and periglomerular fibrosis (arrow). Hematoxylin and eosin
(HE).
Figure 3. Kidney; aged >24 months, C57BL/6J mouse. Marked (grade 4)
nephropathy. There is severe loss of normal renal tubular epithelium and moderate
periglomerular and interstitial fibrosis. Hematoxylin and eosin (HE).
Figure 6. Kidney; C57BL/6J mouse. Severe hydronephrosis with intrapelvic
eosinophilic crystalline material (arrow) and sloughed cells. The capsule is indicated
(C). Hematoxylin and eosin (HE).
Figure 7. Heart; aged >24 months, C57BL/6J mouse. Mild (grade 2)
cardiomyopathy. The cardiomyocytes are variably sized with occasional karyomegaly and
with compartmentalization of fibers by mild interstitial inflammation (region bracketed
by arrows). Mast cells (arrowhead) are distinctly granulated in mouse tissues.
Hematoxylin and eosin (HE).
Figure 8. Heart; aged >24 months, C57BL/6J mouse. Severe (grade 4)
valvular change with cartilaginous metaplasia and moderately thickened leaflets.
Hematoxylin and eosin (HE).
Figure 9. (A) Heart; aged >24 months, C57BL/6J mouse. Moderate (grade
3) cardiomyopathy. Similar to Figure 7, there is compartmentalization of cardiomyocytes by increased interstitial
fibrosis and inflammation affecting a greater percentage of the tissue. HE. (B) Heart;
aged >24 months, C57BL/6J mouse. Masson’s trichrome stained step section highlighting
increased interstitial fibrosis (blue).
Figure 10. Heart and coronary vessels; aged >24 months, C57BL/6J mouse.
Severe (grade 4) arteriosclerosis with moderate (grade 3) perivascular cardiomyopathy.
Note mast cells (arrowheads). Hematoxylin and eosin (HE).
Figure 11. Heart; aged >24 months, C57BL/6J mouse. Severe amyloidosis.
The interstitium is expanded in coalescing foci by acellular eosinophilic to amphophilic
extracellular material (arrows). Hematoxylin and eosin (HE).
Incidences (%) of Neoplasms Diagnosed at End of Life Plus Scheduled Sacrifice in
C57BL/6, 129, and B6,129 Mice
Benign and malignant tumors in an organ are combined. Key: 0%, nonshaded cells; 1% to
10%, light gray cells; 11% to 25%, medium gray cells; >25%, dark gray cells. NA,
not applicable.
Strain 129 is prevalent in GEM backgrounds because of the early and wide
availability of manipulable and robust parental ES cell lines derived from these mice.16,37,63,188 Targeted homologous recombination technology in these ES cells made possible targeted
manipulations (“knockouts”) of the mouse genome.8,44,113 Genetic advances related to this technology were recognized by the Nobel Committee in
2007, when the Nobel Prize in Physiology or Medicine was awarded to Evans, Capecchi, and Smithies.172 Despite recent selection of C57BL/6N as the source of ES cells for the IKMC project,
129 derived ES cell lines remain among the most available (parental) ES cell lines (see, eg,
ftp://ftp.informatics.jax.org/pub/reports/ES_CellLine.rpt for listing of parental ES cell
lines and origins). Strains (or substrains) of 129 are widely represented in GEM and in
mutant ES cells in repositories worldwide. Many of these can found in the Induced Mutant
Strain Resource (IMSR) database at http://www.findmice.org/IMSRSearchForm.jsp. Genetic and phenotypic differences
are expected among at least sixteen 129 substrains, such that identification and utilization
of the correct 129 substrain may be important for some studies. Correct nomenclature should
indicate 129 substrain of origin.57,66,193,221 The most common neoplastic causes and contributors to death in recent studies of
129S4/SvJae mice in recent studies were lung, liver, and Harderian gland tumors (Tables 1, 2). Acidophilic macrophage pneumonia and nephropathy
have been common findings, as well as important contributors to death in some studies100 (Tables 1, 2).
“B6,129” referring to various combinations of C57BL/6 and 129 strains in the genetic
background, became a common genetic background for knockout mice because chimeras derived
from ES cells of (usually light-colored, agouti) 129 mice injected into blastocysts of
C57BL/6 (black, nonagouti) mice were likely to yield viable and detectable (distinctively
colored) chimeras.113 After multiple generations of backcrossing, outcrossing, and inbreeding to generate
GEM for study, the contributions of C57BL/6 and of 129 in experimental populations vary
widely. Thus, really relevant control mice are difficult to determine or obtain, and the
substrains of origin may even not be known. Genetic analysis can assess strain contributions
in individual mice, to identify the most genetically relevant mice for study aims, or for
backcrossing to achieve congenicity with a desired recipient strain. “B6,129” does not
indicate the relative contributions of either strain. Nomenclature that indicates the number
of backcrossing (N) generations and inbreeding (F) generations, as well as correct names of
the parental strains, improves communication and understanding of the genetic contributions.
B6.129 or B6;129 indicates that the mice are fully backcrossed (>10 N generations) and
congenic with the B6 recipient strain (.) or nearly so (;). Several studies of B6,129 mouse
pathology reveal lesion (phenotype) incidences intermediate between those of the C57BL/6 and
129 strains of origin, as illustrated in Tables 1 and 2, where incidences of hematopoietic, lung, and liver
tumors in B6,129 mice are intermediate compared to the parental strains. B6,129 mice may
develop significant tumor burdens at fairly early time points. One of the authors (J. M.
Ward) found that B6,129 mice at one research facility had incidences of follicular lymphoma
(males, 14.6%; females, 22.5%), lung tumors (males, 9.7%; females, 2.5%), and other tumors
(males, 4.7%; females, 5%) by 18 months, with higher incidences at 2 years of age.87 High tumor incidences in control mice can frustrate interpretation of tumors in
experimental groups.213 This is particularly true in GEM studies that aim to find genotype-relevant
phenotypes (as opposed to studies that aim to assess age-associated morbidities), as GEM
that fail to die or develop obvious phenotypes are aged beyond 1 year in the hopes that they
will develop a significant phenotype. The significance of findings in old GEM, when similar
findings are expected also in the background strains, should be interpreted with caution,
especially with small numbers (n) of experimental and control animals.
FVB/N mice are albino mice, developed from N:GP (National Institutes of Health
[NIH] general-purpose mouse) Swiss mice. They became popular for the generation of
transgenic mice largely because of their large and injectable pronuclei and their fecundity.7,212 Published data on the pathology of this strain in long-term studies are limited but
indicate a preponderance of spontaneous lung tumors in both sexes. Additional expected
neoplastic findings in females include pituitary gland adenomas, ovarian tumors, lymphomas,
histiocytic sarcomas, Harderian gland adenomas, and pheochromocytomas and in males include
liver tumors, subcutis neural crest tumors, and Harderian gland adenomas.102,143 Skin sensitivity to carcinogens5,97 and ear tag–associated squamous cell carcinoma18 also are reported in this strain. Proliferative mammary lesions and proliferative,
often prolactin-secreting, pituitary lesions may be especially significant when FVB/N is
used as a background strain in models of mammary neoplasia.170,179,234 FVB/N mice are expected to be blind by about the time of weaning, with genetically
determined retinal degeneration due to homozygosity for the rd1 mutation in the
Pde6b gene.50,246 Seizures are reported in some colonies, and seizure-associated brain lesions (neuron
necrosis in the cerebral cortex, hippocampus and thalamus, and astrocyte hypertrophy) or
liver lesions (centrilobular necrosis) also are encountered in these mice.81,153,158 See also Table 3 and
Supplemental Table S1.
Common Pathology Findings in Long-Term Studies (Longer Than 1 Year) in Inbred Strain
Familiesa
aFindings from multiple studies are summarized for each strain family.
Supplemental Table S1 includes the references, substrain information (when reported),
and original terminology.
Other mouse strains and stocks are used for various research purposes.
Increased recognition of their expected pathology phenotypes may increase recognition of
their relevance to additional research areas. Conditional mutants combine transgenic and
knockout or gene trap technologies, often on multiple backgrounds. GEM backgrounds may be
mixed further to generate mice with multiple intended genetic manipulations. Background
genetics are recognized to affect phenotypes associated with many genotypes but frequently
are ignored or not clear in reports and publications.136,194,209 Several references from the past century offer potentially useful perspectives on the
history and expected phenotypes of inbred mice.132,140,163,166,202 Some additional references are especially relevant to understanding the spectrum of
pathology in mice of various strains or stocks.39,40,41,73,146,159,175
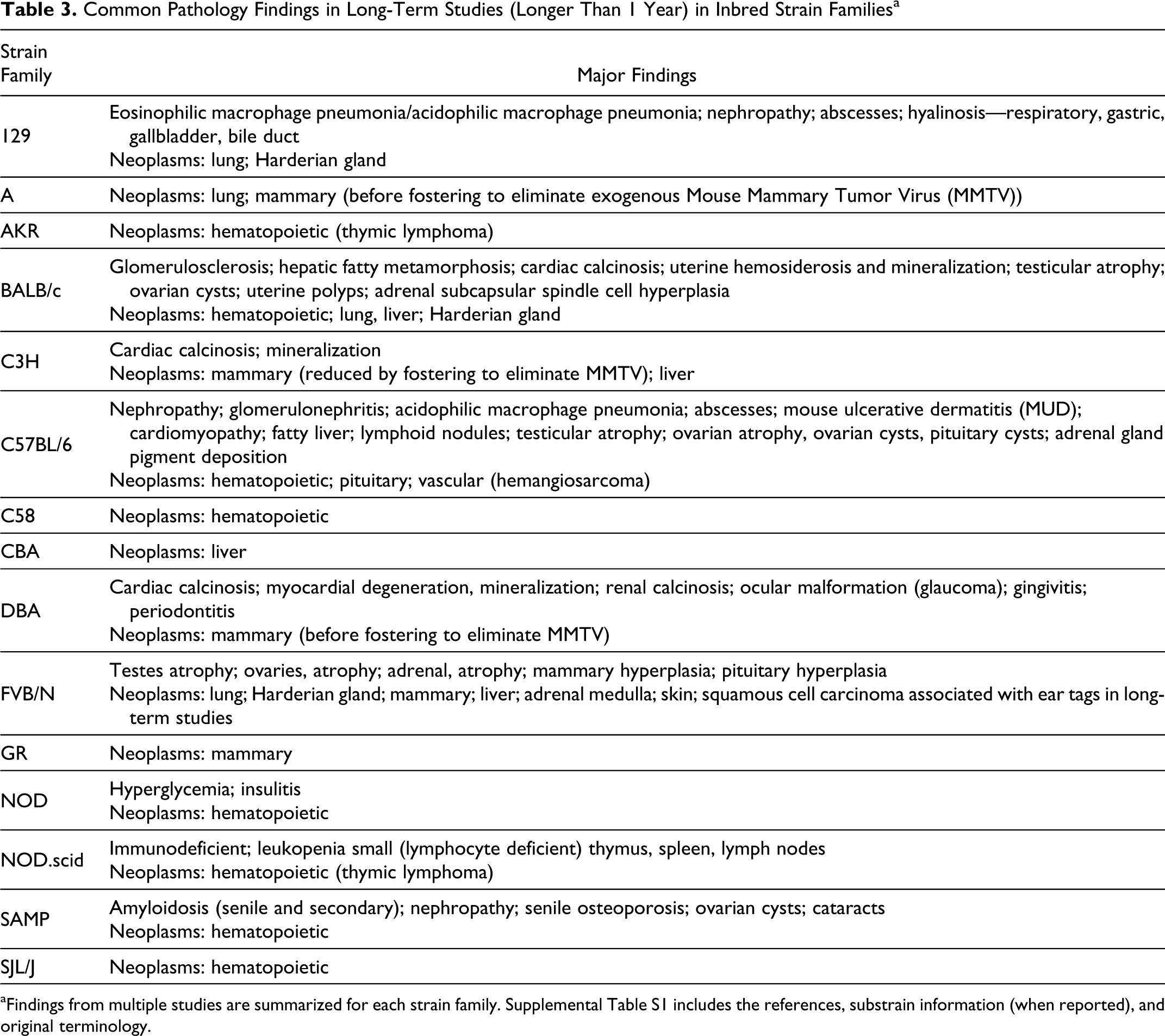
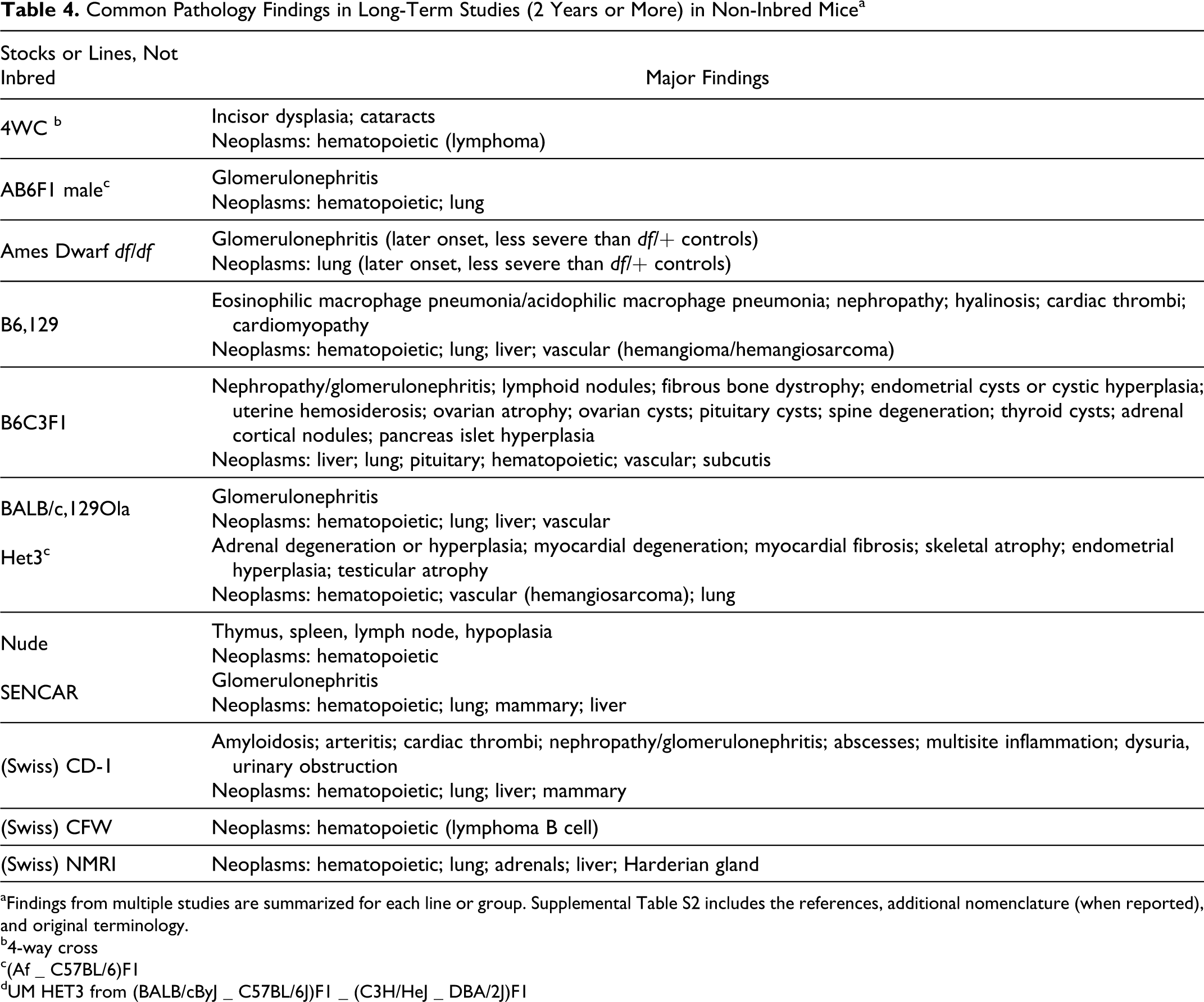
Tables 3 and 4 summarize common pathology
findings from large and long-term (>1 year) studies involving 16 inbred strain families
and several non-inbred stocks. Inbred strain families (Table 3) include 129, A, AKR, BALB/c, C3H, C57BL/6,
C57L, C58, CBA, DBA, FVB/N, GR, NOD.scid, and SJL/J. Non-inbred mice (Table 4 ) include 4WC, AB6F1, Ames dwarf, B6,129,
B6C3F1, BALB/c,129, Het3, nude, Sencar, and several Swiss stocks. Only the most common
findings are listed here, but in summary, they indicate that hematopoietic tumors and lung
and liver tumors should be expected in many of these mice. Nephropathy, amyloidosis, trauma,
dermatitis, abscesses, and cardiac thrombi (or other heart lesions) may shorten or otherwise
affect studies or may be common incidental findings without obvious impact on a study.
Common Pathology Findings in Long-Term Studies (2 Years or More) in Non-Inbred
Micea
aFindings from multiple studies are summarized for each line or group.
Supplemental Table S2 includes the references, additional nomenclature (when
reported), and original terminology.
b4-way cross
c(Af _ C57BL/6)F1
dUM HET3 from (BALB/cByJ _ C57BL/6J)F1 _ (C3H/HeJ _ DBA/2J)F1
Supplemental Tables S1 and S2 include the references for Tables 3 and 4, with substrain information and terminology from
the publications. Users are referred to the original references for study details and for
information on less common lesions that may be relevant or significant in certain substrains
or settings. These tables represent different study designs, different substrains, and
different study conditions. Notable from these tables are similarities between findings
initially reported in inbred strains in the 1930s and findings reported decades later,
although the terminology varies and may have changed over the decades. For example,
neoplasms initially reported as leukemias or reticulum cell sarcomas now are recognized as T
lymphoblastic lymphomas, follicular B cell lymphomas, histiocytic sarcomas, or other less
common types of hematopoietic neoplasms.129,162,237
The Mouse Tumor Biology (MTB; http://tumor.informatics.jax.org) Database and the European Mutant Mouse
Pathology Database (http://www.pathbase.net) are publicly accessible mouse pathology
resources that collect and curate mouse pathology data and images and aim to increase the
value of mice and GEM to the translational research community. The MTB collects neoplasm
data from the primary literature, from other public databases, and by direct submissions
from the scientific community. The MTB database includes spontaneous and induced tumors in
genetically defined mice (inbred, hybrid, mutant, and genetically engineered mice). The data
include standardized tumor names and classifications, pathology reports and images, mouse
strain names, genetic alterations or manipulations (mutations), and literature citations.
The MTB can be searched by tumor site and by strain or mutant name. The MTB also has
developed a tumor frequency grid at http://tumor.informatics.jax.org/mtbwi/dynamicGrid.do. The grid reflects tumor
frequency based on publications and summarizes published records on spontaneous tumors by
inbred strain family and by organ or system, as reports are published and included in the
Mouse Genome Informatics (MGI) database. For 33 inbred strain families, the grid includes
common and less common tumors and reveals strain predispositions such as Harderian gland
neoplasms in BALB/c and FVB/N, myoepitheliomas in BALB/c, and rhabdomyosarcomas in BALB/c
and A/J. The Pathbase database includes macroscopic images and histopathology
photomicrographs that can be retrieved by searching for specific lesions or by class of
lesion, genetic locus, or other parameters.26,130,206,211,252
Experimental Conditions
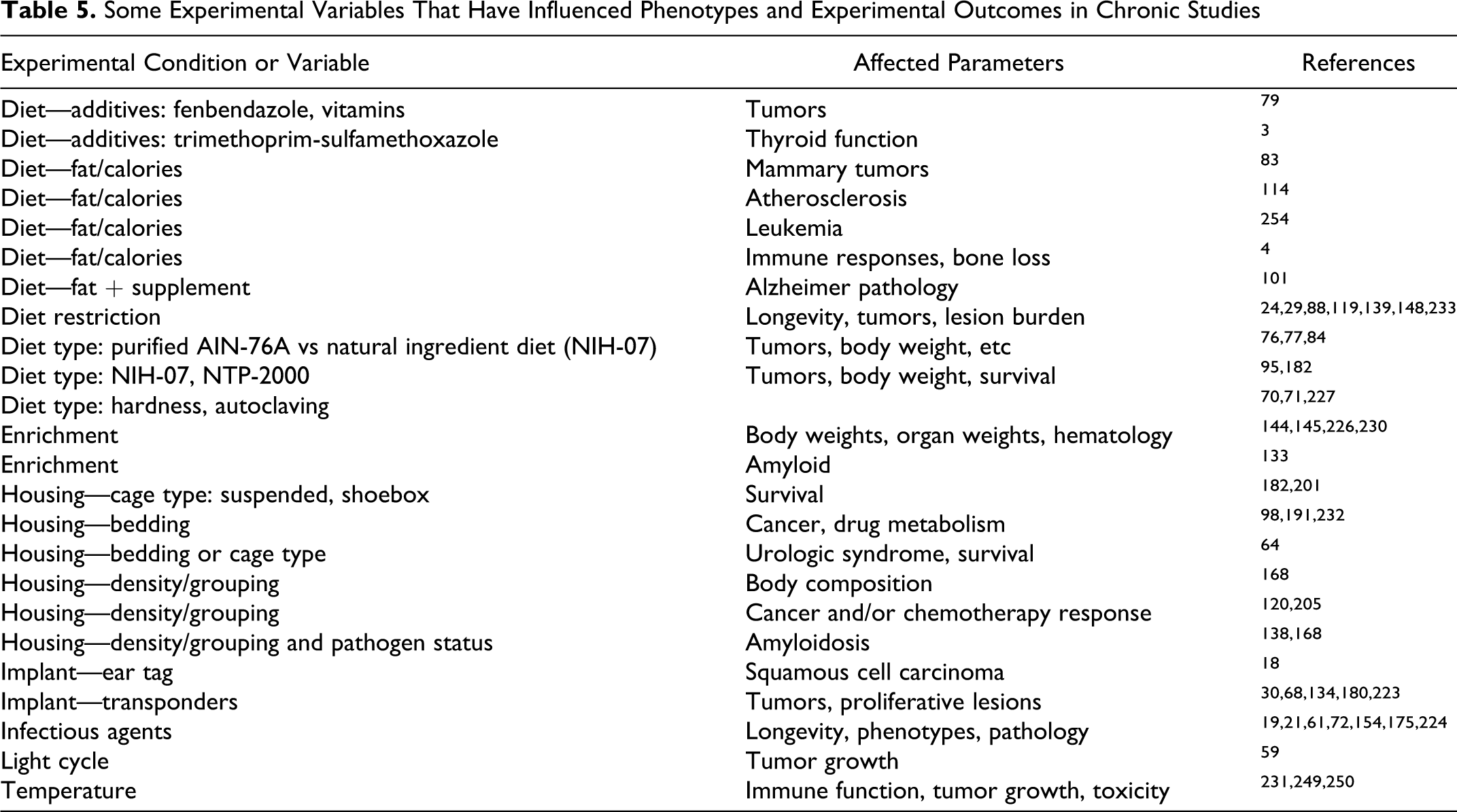
Experimental conditions, especially diet and other environmental factors, have been
recognized to influence mouse phenotypes, survival, and research outcomes.14,61,65,182,186
Diet restriction (DR) or caloric restriction (CR) improves survival and slows
or reduces disease or aging phenotypes in rodent studies. The mechanisms remain a matter of
interest and controversy.149,150,241,242 Longer life spans allow detection of tumors that occur later in life, but diet
restriction that reduces overall tumors may reduce the sensitivity of carcinogenesis bioassays.173 Several “small mice,” such as Ames dwarf or Snell dwarf mice, were noted to be
unusually long lived. These and other small but long-lived animals have led to exploration
of the roles of somatotropic hormones, insulin-like growth factor 1 (IGF-1), and insulin
signaling in life span and development of age-related morbidities.22,23,52,104,214 Inability to consume sufficient calories due to dental or other physical problems, as
well as “voluntary diet restriction” due to diets or additives that are not palatable,
should be considered as potential contributors to small size and increased life span in
relatively undersized mice and is a concern in studies that aim to study or model longevity
by mechanisms other than reduced consumption.69,89,195 Resveratrol and rapamycin are examples of compounds that may extend life span at
least in part by mechanisms that mimic caloric restriction.6,15,25,90 Dietary or caloric excess in current standard caging practices and ad libitum feeding
also raise concerns over the relevance of metabolically morbid, overfed, sedentary, obese
rodents to a normal-weight human population.147 Such concerns also imply that (high-fat or high-calorie) diet challenge may shorten
studies by accelerating morbidities, and perhaps this strategy will simulate some Western
diet–associated morbidities more closely. In addition to issues of dietary restriction or
excess, dietary sources or types of protein and fat, levels of endocrine disruptors
(especially plant source phytoestrogens) or nitrosamines (in animal source protein or fats),
intended additives (treatments), and unintended contaminants are among the dietary variables
that could influence phenotypes and experimental outcomes (Table 5 ).17,96,217,218,240
Some Experimental Variables That Have Influenced Phenotypes and Experimental Outcomes
in Chronic Studies
Commercial laboratory rodent diets are likely to be nutritionally complete and
sufficient, but they vary widely in ingredients and formulation. Processing, handling,
storage, and feeding procedures that ensure preservation of nutrients and freedom from
contamination also vary among distributors and end users. Even when percentages of protein,
fat, carbohydrate, and fiber are similar, the plant, animal, or other sources of these
components can vary substantially. Such variables should be considered in the selection and
standardization of dietary factors to achieve experimental aims.17,43,54,74,96,142,182 Transparency and disclosure are improving with regard to formulation and quality
control for commercial diets and with regard to diets fed to commercially available rodents.
Large vendors of rodent diets now provide much of this information online. Pressure is
increasing to include diet and other relevant husbandry information when reporting animal studies.10,123
Common terms used to describe diets are open formula or
closed formula, which refer to disclosure of the ingredients by the
manufacturer, and natural ingredient or purified, which
refer to the types or sources of ingredients in the diets. Open formula (or open source)
refers to diets for which concentrations of ingredients are publicly available (eg,
http://dvrnet.ors.od.nih.gov/diets_info.asp). Many of today’s commercially
available diets are open formula. Closed formula refers to diets for which the quantitative
ingredient formulation is not publicly available, and composition can vary without public
disclosure.
Natural ingredient diets, sometimes called nonpurified diets, are primarily
cereal based or grain based (including wheat, corn, alfalfa, soy) and also may contain
animal products. NIH-07 and NTP-2000 diets are examples of open-formula, natural ingredient
diets, similar to many common commercial chow diets. From 1980 to 1994, the NIH-07
open-formula, natural ingredient diet was used in National Toxicology Program (NTP) studies
to assess the toxic and carcinogenic potential of chemicals and other agents in rodents. The
NTP-2000 diet has been the primary diet in NTP rodent studies since 1994.182 Soy and alfalfa have been common primary protein sources in natural ingredient diets.
Both are sources of estrogenic activity, which may affect various types of studies.115 Autofluorescence from dietary alfalfa also interferes with some imaging techniques.110
Purified or semipurified diets use exclusively or primarily refined human
food-grade ingredients such as casein, sucrose, cornstarch, and cellulose. The ingredients’
relatively simple chemical compositions facilitate manipulation of individual nutrients for
research purposes. The American Institute of Nutrition (AIN)–purified diet formulations were
developed in the 1970s to establish guidelines for nutritionally adequate purified diets
with inherently less variation than in natural ingredient (cereal-based) diets, to
facilitate interpretation of results among experiments and laboratories. AIN-76 was the
first of these diets. The AIN-76A diet was a modification that included more vitamins.
Subsequent modifications were AIN-93G and AIN-93M, designed for growth and maintenance, respectively.17,135,185
Special diets can be formulated to vary in fat, salt, sugar, vitamins, and
micronutrients or to include antimicrobial agents or test compounds. High-fat diets and
altered preservative levels may require special handling, feeding, and storage to ensure
preservation of nutrients and to delay spoilage or rancidity. Unintended consequences or
phenotypes from antimicrobial agents should be considered.3,79 Altered palatability compared to “control” diets may affect consumption. Pellet
hardness may affect consumption,70,71,216 and a powdered diet may alter tooth wear, leading to incisor overgrowth and malocclusion.111,141
Handling procedures, including autoclaving and storage conditions, also
influence consumption and nutrients or contaminants in rodent diets.70,75,227
Housing density or mice per cage is another important consideration in study
design, with potentially significant impact on life span data (by attrition or early
censoring from a study), on pathology phenotypes (related to wounds and infections), and on
the expense of housing mice. Conspecific (usually male) aggression leads to death or
wounding of cage mates, greater use of females in research, single housing of mice in
long-term studies, and increased expense single housing.33,211,230 Single-housed mice are reported to live longer but weigh more and develop more tumors.92,95
Additional environmental variables that influence mouse phenotypes in long-term
studies are summarized in Table
5.
Experimental Design
Chronic and survival studies involving older GEM will be necessary to assess gene
influences on cancers or other age-related phenotypes, as well as on life span or health
span. Longitudinal or cross-sectional approaches and combinations thereof are used
to assess one or more of the following:
Life span (longevity)
Functional healthy life span (health span)
Efficacy, safety, toxicity, or carcinogenicity of compounds or other interventions
Causes or contributors to death or moribundity
Disease burden or spectrum of pathology
The overall approaches as well as specifics of the implementation, assessment, and
reporting vary with the aims of the studies.56,131,155,167,211,252
Cross-sectional studies assess lesion burden and spectrum of pathology
(phenotypes) at specific time points. These types of studies, especially 2-year rodent
bioassays, are well described in the toxicology literature. The studies are scrutinized by
the Food and Drug Administration (FDA) and other agencies, and best practices are reviewed regularly.117,164,184,251 Methods and data are accessible from the NTP Web site (http://ntp.niehs.nih.gov/) or from
PubMed (http://www.ncbi.nlm.nih.gov/pubmed). The cross-sectional, scheduled sacrifice
study design determines lesion burden and spectrum of pathology at specific time points (eg,
3, 6, 12, 18, 24 months), usually aiming to evaluate drug safety or compound and xenobiotic
carcinogenicity. Animals that die before scheduled sacrifice may be evaluated when feasible,
but examination may be compromised or data lost due to postmortem deterioration of the
tissues. Although such studies frequently use non-inbred Swiss (such as CD-1, ICR, SW) mice
or B6C3F1 mice, and historical control data are available for these mice primarily, the
studies can inform the design, practice, and analysis of studies aiming to understand the
influence of intended gene manipulations on age-associated morbidities (phenotypes) and
their onset and progression in GEM and relevant control mice. The systematic and
comprehensive pathology evaluations are a model for large projects. C57BL/6, 129, and FVB/N
mice are likely to develop tumors and other age-related pathology phenotypes between 12 and
18 months of age, such that a 2-year study may be sufficient to achieve an aim of assessing
if a genotype or intervention reduces neoplasms and age-associated morbidities. When
neoplasms or morbidities in control groups are fewer or occur later, longer aging studies
may be necessary. On the other hand, many GEM studies aim to identify increased or
accelerated neoplasms or other disease phenotypes resulting from a genetic alteration (such
as activation of a putative oncogene or inactivation of a putative tumor suppressor). If the
GEM line does not develop significant phenotypes by 18 months or even by 12 months of age,
there may be little justification to continue the study beyond 12 or 18 months because
neoplasms and age-associated morbidities in the control mice will complicate their
interpretation.
Longitudinal survival studies that assess life span (longevity) also can assess
causes of death (COD) and contributing causes of death (CCOD). In “euthanasia when moribund”
longitudinal studies, definition of moribund state and control of environmental variables,
careful observation of mice, and prompt and expert examination, dissection, and pathology
assessment are key to obtaining useful results.14,56,131,252 Methods to determine COD or, more accurately, CCOD aim to collect diagnostic
specimens, without “shortening” the life span and biasing life span measurements.29,56,127 Because large tumors are identified easily, they often are reported as the COD or
primary CCOD. Regulatory and veterinary concerns to alleviate pain and distress105,107 may dictate early euthanasia and “shorten” life span measurements due to clinically
obvious but preterminal lesions, such as large benign tumors or ulcerative dermatitis.
Suboptimal clinical assessment leading to unexpected deaths, prolonged postmortem intervals,
and deteriorated specimens can compromise determination of COD or CCOD. Requirements of this
study design (euthanasia when moribund) also can preclude procurement of useful clinical
pathology or other test data because animals are not assessed until they are severely
compromised.
A combination strategy of survival and scheduled evaluations (longitudinal and
cross-sectional analysis) to include scheduled physiological evaluations and
clinical pathologycan maximizes health span data from a study that will consume substantial
animal, husbandry, and pathology resources, no matter how it is done.56,131,225,244
Longitudinal survival studies that assess life span (longevity) but do not
assess causes of or contributors to death are used in the National Institute on Aging (NIA)
Interventions Testing Program (ITP; http://www.nia.nih.gov/ResearchInformation/ScientificResources/InterventionsTestingProgram.htm).
HET3 mice used in this program are (F2) offspring of CByB6F1/J and C3D2F1/J (combining
BALB/cBy, C57BL/6, C3H, and DBA/2) strains. Their genetic diversity is intended to “ensure
that causes of age-related mortality will be varied, and thus, that any significant increase
in life span from an intervention will not result merely from a beneficial effect on a
strain-specific disease.”69 Similarly, heterogeneous stocks, such as 4-way cross (4WC) mice, which are (F2)
offspring of (AKR/J × DBA/2J)F1 females and (C57BL/6J × SJL/J)F1 males, and 8-way cross,
also are used in these types of aging studies.151,156 Hematopoietic tumors were the primary cause of death in 4WC mice and found in 55% of
male and 43% of female mice when pathology was evaluated.53
Frequency of tumors (or of other pathology phenotypes) has been defined as the
proportion of mice with tumors in animals submitted for necropsy. The data may be skewed if
some strains are maintained and studied to later ages than others (eg, aging nulliparous
research populations vs younger breeding populations). But once populated with sufficient
data, this approach may help to identify strains, sexes, ages, mutations, or environmental
situations that are likelier to develop or contribute to certain phenotypes and help to
develop testable hypotheses regarding the mechanisms. Compared to large studies that aim
primarily for incidence or prevalence data or responses to an intervention, involving one or
few strains, the MTB database or Pathbase can be mined for frequency data among many strains.26,154,192
Procedural Considerations (Materials and Methods) relevant to pathology in mouse aging
studies. “For scientific, ethical and economic reasons, experiments involving
animals should be appropriately designed, correctly analysed and transparently reported.
This increases the scientific validity of the results, and maximises the knowledge gained
from each experiment.”121 The ARRIVE (Animals in Research: Reporting In Vivo Experiments) guidelines were
published in 2010 in response to identification of deficiencies in the reporting,
experimental design, and statistical analysis of peer-reviewed, published biomedical
research using laboratory animals.121–123,198 They have been endorsed by a number of major publications and funding agencies, and a
similar Guidance for the Description of Animal Research in Scientific
Publications was published recently by the National Academy of Sciences.108 Especially relevant to studies of aging, health span and pathology phenotypes of GEM
are methods for
End-of-life (EOL) determination
Health span assessment
Terminal examinations (necropsy)
Tissue sampling, trimming, and preservation
Nomenclature of mice, genes, and genetic manipulations
Standardized diagnostic criteria and terminology
Grading of lesions
Cause of death (COD, CCOD) assignment
Historical control data (HCD)
Peer review (quality control)
End-of-Life (EOL) Determinations
In much of aging research, life span has been the primary criterion of
interest. It is important to optimize EOL determination for humane and for
scientific reasons. Timely euthanasia of mice based on validated markers of imminent death
allow implementation of end points that alleviate terminal distress, minimally or
insignificantly affect life span data, and permit timely collection of tissue specimens.
Subjective clinical signs used as markers of imminent death (within 24 hours) include (1)
nonresponsiveness to touch, (2) cool body temperature to the touch, (3) slow or labored
respiration, (4) hunched posture with matted fur, and (5) poor body condition score (eg,
per Ullman-Cullere and Foltz228). More objective or quantitative criteria include (1) failure to eat and drink
(determined by food hopper and water bottle weights), (2) loss of body weight,69,131 and (3) body temperature (as determined by telemetry).183 Telemetry transponders offer advantages of rapid definitive animal identification
and temperature measurements, but transponders, microchips, or other implanted devices may
contribute to neoplasia and mortality in susceptible mice.30,68,134,180,223
Health Span Determination
Increasingly, aging studies in mouse models are shifting toward more
characterization of health span and onset of age-related disease.9,124,195,215 At The Jackson Laboratory, a large study is progressing to characterize the aging
of 31 inbred strains.174,211,253 At the University of Washington (UW), studies are under way to define the health
and life span of a long-lived GEM (with targeted mitochondrial overexpression of the
antioxidant enzyme catalase) compared to their wild-type (control) littermates.131 Both projects include physiological testing and pathology. Pathology can identify
(diagnose) many causes of abnormal (positive) findings. For example, abscesses or
inflammatory lesions diagnosed by histopathology are likely to contribute to leukocytosis
identified by hematology or by flow cytometry, cytokine and chemokine responses, observed
and measured splenomegaly (reactive myeloid and/or lymphoid hyperplasia), and possibly
declines in body condition or activity. Otitis diagnosed by histopathology may be relevant
to increased auditory-evoked responses (deafness).157 Otitis interna, arteritis, or infarcts diagnosed by histopathology may be relevant
to clinical signs of vestibular syndromes (rolling, spinning, head tilt).200 Acidophilic macrophage pneumonia, amyloidosis, nephropathy, cardiac thrombi, or
neoplasms diagnosed by histopathology may be primary contributors to failure to thrive or
to perform “normally” in physiologic tests. Both projects combine cross-sectional and
longitudinal approaches, with physiologic testing and pathology at predetermined
cross-sectional time points and end-of-life examinations when mice are moribund. Intensive
and expert standardized, daily monitoring is essential to prevent early censoring for
nonfatal but readily observable lesions (such as rectal prolapse, skin lesions, or masses)
and to prevent loss of pathology data due to unexpected death and postmortem degeneration
(autolysis).
Terminal Evaluations
Systematic, standardized, and comprehensive terminal evaluations can maximize the
usable data from individual animals and the utility and comparability of data within and
between studies.* Prompt
assessment of illness or other concerning disease phenotypes can help to identify problems
and causes early to minimize loss of animals, money, and time and to prevent serious
compromise of the colony, project, or program. A strategic and systematic plan for
pathology should promptly diagnose infections and other complications, assess and validate
positive findings (“phenotypes”) identified by other tests, and assess causes and
contributors to death and to age-associated morbidities.38,192 However, comprehensive pathology protocols that evaluate 40 or more tissues on 10
to 20 slides rapidly become a major expense. Abbreviated protocols that emphasize
confirmed or expected target tissues have been explored as cost-saving alternatives152 and may be statistically justifiable in certain situations. At a minimum,
preservation of perfusion fixed mice or of fixed specimens that cannot be evaluated
promptly may bridge situations where pathology resources are limited and permit later
evaluation to address questions that arise after analysis of other data.
Tissue Sampling, Trimming, and Preservation
To support harmonized and standardized organ processing in regulatory toxicology
studies, European RITA (Registry of Industrial Toxicology Animal-data) and US
NACAD (North American Control Animal Database) working groups developed trimming guides
that describe tissue preparation, sample size, direction of sectioning, and numbers of
sections by organ.13,125,161,190 Orientation of tissue (eg, sagittal, transverse, or coronal) and numbers of
sections per tissue can affect data such as number of tumors identified in a tissue—for
example, 55 tumors in serial sections of 140 thyroids compared to 9 tumors identified in
single random sections of 177 thyroids,62 more heart lesions identified when more sections are examined,219 or more pituitary (or liver) tumors identified when more tissue is examined.181 Collection methods, tissue weights or other measurements, fixatives, tissue
orientation, and processing methods may need to vary according to the aims of a project,
but a systematic approach based on published and reviewed recommendations, with sufficient
detail to reproduce the procedures, will improve the utility of data.32,38,42,67,192,199,207 Regardless of the initial aims of the study, appropriately preserved tissues may be
useful years or even decades later to answer new experimental questions through additional
histology, immunohistochemistry, or newer molecular genetic techniques such as protein
extraction or profiling gene expression.2,126,245 Strategies for identification and curation of specimens should be included in the
planning for long-term studies.
Nomenclature of Mice, Genes, and Genetic Manipulations
At a time when sharing of data and meta-analysis of experimental results is becoming
a dominant mode of scientific investigation, failure to respect formal nomenclatures can
cause confusion and errors and, in some cases, contribute to poor science.209 Correct nomenclature of mice, genes, and genetic manipulations is essential to
accurate reporting of a study. Common errors include incomplete or misleading mouse names,
out-of-date gene names, and incorrect designation of mouse vs human genes and gene
products (proteins). The need for correct nomenclature and practical aspects of using it
are reviewed in this issue and elsewhere.137,209,247
Standardized Diagnostic Criteria and Terminology
Standardized diagnostic criteria and terminology are essential for interpretation of
findings in a study and for comparisons between studies. Differences in
diagnostic criteria and their interpretation contribute to interlaboratory variability in
incidences or rates of tumors (spontaneous and induced) or other lesions. Improved
understanding of mechanisms also can contribute to modifications of criteria or terminology.† Mechanistically, molecularly novel GEM phenotypes may justify novel criteria and
terminology, but expertise in “normal” mouse anatomy and pathology is critical to identify
and interpret novel pathology phenotypes.46,48,109,210 Supplemental Tables S1 and S2 include terminology from the original publications
and illustrate some of the evolution of terminology over the past 8 or more decades.
Large-scale data and project management, as well as accurate and useful
reporting, require attention to terminology. The International Harmonization of
Nomenclature and Diagnostic Criteria for Lesions in Rats and Mice (INHAND) project,
Pathbase, and the National Cancer Institute’s (NCI’s) Mouse Models of Human Cancer
Consortium (MMHCC) are resources for diagnostic criteria and terminology that are highly
relevant to long-term studies in GEM.
The INHAND project is a joint initiative of the Societies of Toxicologic
Pathology from Europe (ESTP), Great Britain (BSTP), Japan (JSTP), and North America (STP)
to develop an internationally accepted nomenclature for proliferative and nonproliferative
lesions in laboratory animals (http://goreni.item.fraunhofer.de/back_inhand.php">http://goreni.item.fraunhofer.de/back_inhand.php).12,187,220,229 The toxicologic pathology community has a long history of expertise in rodent
pathology and interpretation of lesions. The criteria and terminology are primarily
relevant to spontaneous and carcinogen-induced lesions, and they undergo expert scrutiny
and peer review but may not be entirely relevant to GEM pathology.
Pathbase (http://www.pathbase.net) is a database structured around an ontology of
mouse lesions, searchable by pathology terms, developmental stage, anatomy, and gene
attributes (GO terms), developed by bioinformaticians and pathologists specifically to
support GEM phenotyping. Online images are included for some entities. Data and images can
be submitted from the scientific community as well.192,206,208
The NCI MMHCC (http://emice.nci.nih.gov/, http://www.nih.gov/science/models/mouse/resources/hcc.html) has developed
GEM cancer resources, including the Cancer Models Database (CaMOD) (http://cancermodels.nci.nih.gov/), the Cancer Images Database (http://cancerimages.nci.nih.gov/), and consensus papers (combining
veterinary and medical pathology and scientific expertise) on diagnostic criteria and
terminology for neoplasia of the hematopoietic system,129,162 nervous system,86 mammary gland,45,112 prostate,197 lung,171 and intestine,31 as well as on precancers.47 MMHCC terminology is especially useful when mechanisms, morphology, and behavior of
genetically engineered lesions differ from spontaneous and chemically induced lesions in
the toxicology literature.46 Despite differences in project aims and resources, early standardization of
diagnostic criteria and terminology with a widely recognized system will facilitate
comparisons across studies and evaluators and will improve communication and
reporting.
Grading of Lesions
Grading or scoring of lesions is essential to studies that assess severity of
disease conditions or responses to interventions, reviewed in Ward and Thoolen.238 Often grading is a semiquantitative assessment of severity, scored from 0 to 3 or 0
to 4. Requirements of a study, statistical model, or data management system influence
selection of a grading system. The advent of digital microscopy and automation of
measurements and counting can improve quantification35,36,178 or may be useful to validate a grading system. Society of Toxicologic Pathology
Best Practices guidelines for toxicologic histopathology recommend the use of grading
systems that are (1) definable, (2) reproducible, and (3) meaningful.58 The new INHAND liver nomenclature provides an example of grading for nonneoplastic
liver lesions.220 Grading of kidney and/or heart lesions has been used in some studies of aging mice
to assess relative severities of lesions relevant to age-associated morbidities.103,104,131,225
Cause of Death (COD, CCOD) Assignment
Determination of COD or CCOD, as illustrated in Table 1, can be expected to be important to the
aims of aging studies in mice. The National Center for Toxicological Research (NCTR)
developed a system for grading the certainty of cause of death, or cause of morbidity, in
the case of moribund animals as follows:128
If a particular neoplasm (or other pathological condition) can be identified by the
examining pathologist as the most likely reason for the animal’s death or removal,
then it is reported as the probable cause of death. At most, one probable cause of
death may be assigned.
If a probable cause of death is not identified, then a neoplasm (or other lesion)
believed to have contributed to an animal’s death, but not thought to be the singular
cause, may be reported by the pathologist as a contributory cause. More than one
contributory cause of death may be assigned to an animal.
Lesions that are thought to have had a possible influence on an animal’s death or
removal may be listed as equivocal.
If there is no neoplasm or other equivocal cause of death for an animal, then that
animal is reported as having died from an unknown cause.
Similar systems have been implemented in studies of aging mice.29,103,104,131,225
Historical Control Data (HCD)
“The concurrent control group is the most relevant comparator for determining
treatment-related effects in a study,”1,17 but HCD can help to guide study design and determine appropriate n by power
analyses and may offer useful perspective on rare or unusual findings and factors that
contribute to them.213 Pathology practices, including necropsy and trimming procedures and application of
diagnostic criteria and terminology, can affect study data and HCD. HCD are most useful if
these factors are standardized and reported in adequate detail.60,117 In the RITA and NACAD databases60,116,118 and in the NTP database, data are collected by standardized procedures for tissue
sampling and trimming, histopathology is assessed according to internationally harmonized
nomenclature and diagnostic (INHAND) criteria, and validity of data is also confirmed by
peer review. The Mouse Tumor Biology Database (http://tumor.informatics.jax.org/) and Pathbase (http://www.pathbase.net) use standardized mouse anatomy and pathology
ontologies in the Mouse Disease Information System (MoDIS) for data capture, and accept
mouse pathology data and images from the Jackson Aging Center initiative, as well as from
the scientific community.192,211,252
Peer Review, Quality Control
Peer review of pathology data and diagnoses and other quality control
measures are applied in toxicology studies subject to FDA and other regulatory
scrutiny, and peer review is required for incorporation of data into the NTP and RITA databases.118 Peer review has been found to increase study quality with minimal time and cost. It
ensures that consistent criteria are used for diagnoses and increases the reliability of
study data. The Society of Toxicologic Pathologists (STP) has published recommendations
specifically on the peer-review process,165,203,204 and the process is described in the Best Practices Guidelines for Toxicologic Histopathology.34,58,164 Digital and Web-based technologies offer opportunities78,178 for collaborative remote consulting and peer review of GEM pathology in aging and
other phenotyping studies.49,192 Veterinary and medical pathologists working in mouse pathology conduct or
participate in these activities informally by e-mailing images or by multi-institution teleconferences,49 but there is no formal review procedure for mouse pathology except by colleagues
and journal reviewers. Except for pathology publications, journal reviewers often are not
pathologists, and publication of incorrect and misleading pathology diagnoses has been identified.49,109,211
Conclusion
Aging studies are necessarily lengthy, labor intensive, and resource intensive. Housing
costs alone can exceed $1000 for 1 cage of 1 to 5 mice to attain 36 months of age, depending
on institution per diem rates. To ensure accuracy of survival data, investment in expert
daily examination is essential. To maximize data from valuable mice, advance planning for
end-of-life evaluations also is essential. Experimental design that includes
standardization, adequate and systematic pathology evaluation, quality assurance, peer
review, and common sense have much to offer all areas of preclinical translational research,
especially those involving GEM in aging research. A combination of cross-sectional and
longitudinal study design, coupling antemortem physiologic testing with necropsy and
histopathology assessment, offers great opportunity to maximize the translational research
value of mouse research tools. Aiming to optimize design, implementation, analysis, and
reporting of translational studies involving aging mice and GEM, the resources included here
offer historical and contemporary context regarding influences of genetic background and
experimental variables on experimental outcomes relevant to mouse pathobiology.
Supplemental Material
Supplemental Material, VET430696_Supplemental_tables - Pathobiology of
Aging Mice and GEM: Background Strains and Experimental Design
Supplemental Material, VET430696_Supplemental_tables for Pathobiology of Aging
Mice and GEM: Background Strains and Experimental Design by C. F. Brayton, P. M.
Treuting, and J. M. Ward in Veterinary Pathology
Footnotes
The author(s) declared no potential conflicts of interest with respect to the research,
authorship, and/or publication of this article.
The author(s) received no financial support for the research, authorship, and/or
publication of this article.
Supplementary material for this article is available on the Veterinary
Pathology Web site at .
Abbreviations
*
References 42, 131, 176, 192, 211, 225, 235.
References 34, 82, 85, 91, 93, 94, 116, 189, 243.
References
1.
AbbottA. Mouse project to find each gene’s role: International
Mouse Phenotyping Consortium launches with a massive funding commitment.
Nature.
2010;465(410).
2.
Al-MullaF, ed. Formalin-Fixed Paraffin-Embedded Tissues:
Methods and Protocols. Totawa, NJ:
Humana Press; 2011.
3.
AltholtzLYLa PerleKMQuimbyFW. Dose-dependant hypothyroidism in mice induced by
commercial trimethoprim-sulfamethoxazole rodent feed. Comp
Med.
2006;56(5):395–401.
4.
AmarSZhouQShaik-DasthagirisahebY. Diet-induced obesity in mice causes changes in immune
responses and bone loss manifested by bacterial challenge. Proc
Natl Acad Sci.
2007;104(51):20466–20471.
5.
AnikinIVKozlovAPPopovAV. Susceptibility of wild and knockout p53 FVB/N line mice to
benz(a)pyrene-induced subcutaneous sarcoma. Vopr Onkol.
2002;48(6):700–702.
6.
AnisimovVNZabezhinskiMAPopovichIG. Rapamycin extends maximal lifespan in cancer-prone
mice. Am J Pathol.
2010;176(5):2092–2097.
7.
AuerbachABNorinskyRHoW. Strain-dependent differences in the efficiency of
transgenic mouse production. Transgenic Res.
2003;12(1):59.
8.
AuerbachWDunmoreJHFairchild-HuntressV. Establishment and chimera analysis of 129/SvEv- and
C57BL/6-derived mouse embryonic stem cell lines.
Biotechniques.
2000;29(5):1024–1028,
1030, 1032.
9.
AustadS. Recent advances in vertebrate aging research
2009. Aging Cell.
2010;9(3):297–303.
10.
AyalaJESamuelVTMortonGJ. Standard operating procedures for describing and
performing metabolic tests of glucose homeostasis in mice. Dis
Model Mech.
2010;3(9–10):525–534.
11.
BabbittJTKharaziAITaylorJM. Hematopoietic neoplasia in C57BL/6 mice exposed to
split-dose ionizing radiation and circularly polarized 60 Hz magnetic
fields. Carcinogenesis.
2000;21(7):1379–1389.
12.
BachUHaileyJRHillGD. Proceedings of the 2009 National Toxicology Program
Satellite Symposium: Tumor pathology and INHAND (International harmonization of
Nomenclature and Diagnostic Criteria for lesions in rats and mice)
nomenclature. Toxicol Pathol.
2010;38(1):9–36.
13.
BahnemannRJacobsMKarbeE. RITA—registry of Industrial Toxicology Animal-data—guides
for organ sampling and trimming procedures in rats. Exp Toxicol
Pathol.
1995;47(4):247–266.
14.
BargerJLWalfordRLWeindruchR. The retardation of aging by caloric restriction: its
significance in the transgenic era. Exp Gerontol.
2003;38(11–12):1343–1351.
15.
BargerJLKayoTVannJM. A low dose of dietary resveratrol partially mimics caloric
restriction and retards aging parameters in mice. PLoS
One.
2008;3(6):e2264.
16.
BaribaultHKemlerR. Embryonic stem cell culture and gene targeting in
transgenic mice. Mol Biol Med.
1989;6(6):481–492.
17.
BarnardDELewisSMTeterBB. Open- and closed-formula laboratory animal diets and their
importance to research. J Am Assoc Lab Anim Sci.
2009;48(6):709–713.
18.
BaronBWLanganGHuoD. Squamous cell carcinomas of the skin at ear tag sites in
aged FVB/N mice. Comp Med.
2005;55(3):231–235.
19.
BartholdSW. Opportunistic infections in research rodents: the
challenges are great and the hour is late. ILAR J.
1998;39(4):316–321.
20.
BartholdSW. Genetically altered mice: phenotypes, no phenotypes, and
faux phenotypes. Genetica.
2004;122(1):75–88.
21.
BartholdSW. Microbes and the evolution of scientific fancy
mice. ILAR J.
2008;49(3):265–271.
22.
BartkeABrown-BorgH. Life extension in the dwarf mouse. In:
GeraldPS, ed. Current Topics in Developmental Biology. Vol
63. New York, NY: Academic
Press;
2004:189–225.
23.
BartkeAMasternakMMAl-RegaieyKA. Effects of dietary restriction on the expression of
insulin-signaling-related genes in long-lived mutant mice.
Interdiscip Top Gerontol.
2007;35:69–82.
24.
BartkeABonkowskiMMasternakM. THow diet interacts with longevity genes.
Hormones (Athens).
2008;7(1):17–23.
25.
BaurJAPearsonKJPriceNL. Resveratrol improves health and survival of mice on a
high-calorie diet. Nature.
2006;444(7117):337–342.
26.
BegleyDAKrupkeDMNeuhauserSB. The Mouse Tumor Biology database (MTB): a central
electronic resource for locating and integrating mouse tumor pathology
data. Vet Pathol. 2011Jan31. [Epub ahead of print]
27.
BittnerJJ. The breeding behavior and tumor incidence of an inbred
albino strain of mice. Am J Cancer.
1935;25:113–121.
28.
BittnerJJ. The breeding behavior and tumor incidence of a black
agouti stock of mice. Am J Cancer.
1935;25:614–620.
29.
BlackwellBNBucciTJHartRW. Longevity, body weight, and neoplasia in ad libitum–fed
and diet-restricted C57BL6 mice fed NIH-31 open formula diet.
Toxicol Pathol.
1995;23(5):570–582.
30.
BlanchardKTBarthelCFrenchJE. Transponder-induced sarcoma in the heterozygous p53+/–
mouse. Toxicol Pathol.
1999;27(5):519–527.
31.
BoivinGPWashingtonKYangK. Pathology of mouse models of intestinal cancer: consensus
report and recommendations. Gastroenterology.
2003;124(3):762.
32.
BolonBBraytonCCantorGH. Editorial: best pathology practices in research using
genetically engineered mice. Vet Pathol.
2008;45(6):939–940.
33.
BolonBBartholdSWBoydKL. Gender agenda: sex bias can be justified in animal
research. Nature.
2010;466(7302):28.
34.
BoormanGAHasemanJKWatersMD. Quality review procedures necessary for rodent pathology
databases and toxicogenomic studies: the National Toxicology Program
experience. Toxicol Pathol.
2002;30(1):88–92.
35.
BoyceJTBoyceRWGundersenHJ. Choice of morphometric methods and consequences in the
regulatory environment. Toxicol Pathol.
2010;38(7):1128–1133.
36.
BoyceRWDorph-PetersenKALyckL. Design-based stereology: introduction to basic concepts
and practical approaches for estimation of cell number. Toxicol
Pathol.
2010;38(7):1011–1025.
37.
BradleyAEvansMKaufmanMH. Formation of germ-line chimaeras from embryo-derived
teratocarcinoma cell lines. Nature.
1984;309(5965):255–256.
38.
BraytonCJusticeMMontgomeryCA. Evaluating mutant mice: anatomic
pathology. Vet Pathol.
2001;38(1):1–19.
39.
BraytonC. Spontaneous diseases in commonly used mouse
strains. In: FoxJGBartholdSWDavissonM, eds. The Mouse in Biomedical Research: Diseases.
Vol II. 2nd ed. Burlington,
MA: Elsevier;
2006:623–717.
40.
BronsonRT. Rate of occurrence of lesions in 20 inbred and hybrid
genotypes of rats and mice sacrificed at 6 month intervals during the last years of
life. In: HarrisonDE, ed. Genetic Effects on Aging II.
Caldwell, NJ: Telford
Press;
1990:279–358.
41.
BronsonRTLipmanRD. Reduction in rate of occurrence of age related lesions in
dietary restricted laboratory mice. Growth Dev Aging.
1991;55(3):169–184.
42.
BronsonRT. Pathologic phenotyping of mutant mice.
Methods Mol Biol.
2009;530:435–461.
43.
BrownNMSetchellKD. Animal models impacted by phytoestrogens in commercial
chow: implications for pathways influenced by hormones. Lab
Invest.
2001;81(5):735–747.
44.
CapecchiMR. Gene targeting in mice: functional analysis of the
mammalian genome for the twenty-first century. Nat Rev
Genet.
2005;6(6):507–512.
45.
CardiffRDAnverMRGustersonBA. The mammary pathology of genetically engineered mice: the
consensus report and recommendations from the Annapolis meeting.
Oncogene.
2000;19(8):968–988.
46.
CardiffRDRosnerAHogarthMA. Validation: the new challenge for
pathology. Toxicol Pathol.
2004;32(suppl
1):31–39.
47.
CardiffRDAnverMRBoivinGP. Precancer in mice: animal models used to understand,
prevent, and treat human precancers. Toxicol Pathol.
2006;34(6):699–707.
48.
CardiffRDWardJM, Barthold SW. ‘One medicine—one pathology’: are veterinary
and human pathology prepared?Lab Invest.
2008;88(1):18–26.
49.
CardiffRD. How to phenotype a mouse. Dis
Model Mech.
2009;2(7–8):317–321.
50.
ChangBHawesNLHurdRE. Retinal degeneration mutants in the mouse.
Vision Res.
2002;42(4):517–525.
51.
CheslerEJMillerDRBranstetterLR. The Collaborative Cross at Oak Ridge National Laboratory:
developing a powerful resource for systems genetics. Mamm
Genome.
2008;19(6):382–389.
52.
ChibaTTsuchiyaTKomatsuT. Development of a bioassay to screen for chemicals
mimicking the anti-aging effects of calorie restriction. Biochem
Biophys Res Commun.
2010;401(2):213–218.
53.
ChrispCETurkePLucianoA. Lifespan and lesions in genetically heterogeneous
(four-way cross) mice: a new model for aging research. Vet
Pathol.
1996;33(6):735–743.
54.
ClarkeHECoatesMEEvaJK. Dietary standards for laboratory animals: report of the
Laboratory Animals Centre Diets Advisory Committee. Lab
Anim.
1977;11(1):1–28.
55.
CollinsFSRossantJWurstW. A mouse for all reasons.
Cell.
2007;128(1):9–13.
56.
ConoverCABaleLKMaderJR. Longevity and age-related pathology of mice deficient in
pregnancy-associated plasma protein-A. J Gerontol A Biol Sci Med
Sci.
2010;65(6):590–599.
57.
CookMNBolivarVJMcFadyenMP. Behavioral differences among 129 substrains: implications
for knockout and transgenic mice. Behav Neurosci.
2002;116(4):600–611.
58.
CrissmanJWGoodmanDGHildebrandtPK. Best practices guideline: toxicologic
histopathology. Toxicol Pathol.
2004;32(1):126–131.
59.
DauchyRTDupepeLMOomsTG. Eliminating animal facility light-at-night contamination
and its effect on circadian regulation of rodent physiology, tumor growth, and
metabolism: a challenge in the relocation of a cancer research
laboratory. J Am Assoc Lab Anim Sci.
2011;50(3):326–336.
60.
DeschlUKittelBRittinghausenS. The value of historical control data: scientific
advantages for pathologists, industry and agencies. Toxicol
Pathol.
2002;30(1):80–87.
61.
DubosRSavageDSchaedlerR. Biological Freudianism: lasting effects of early
environmental influences. Int J Epidemiol.
2005;34(1):5–12.
62.
EustisSLHaileyJRBoormanGA. The utility of multiple-section sampling in the
histopathological evaluation of the kidney for carcinogenicity studies.
Toxicol Pathol.
1994;22(5):457–472.
63.
EvansMJKaufmanMH. Establishment in culture of pluripotential cells from
mouse embryos. Nature.
1981;292(5819):154–156.
64.
EverittJIRossPWDavisTW. Urologic syndrome associated with wire caging in AKR
mice. Lab Anim Sci.
1988;38(5):609–611.
65.
EverittJIFosterPM. Laboratory animal science issues in the design and conduct
of studies with endocrine-active compounds. ILAR J.
2004;45(4):417–424.
66.
FestingMFSimpsonEMDavissonMT. Revised nomenclature for strain 129 mice.
Mamm Genome.
1999;10(8):836.
67.
FietteLSlaouiM. Necropsy and sampling procedures in
rodents. Methods Mol Biol.
2011;691:39–67.
68.
FloydEMannPLongG. The Trp53 hemizygous mouse in pharmaceutical development:
points to consider for pathologists. Toxicol Pathol.
2002;30(1):147–156.
69.
FlurkeyKAstleCMHarrisonDE. Life extension by diet restriction and N-acetyl-L-cysteine
in genetically heterogeneous mice. J Gerontol A Biol Sci Med
Sci.
2010;65(12):1275–1284.
70.
FordDJ. Effect of autoclaving and physical structure of diets on
their utilization by mice. Lab Anim.
1977;11(4):235–239.
71.
FordDJ. Influence of diet pellet hardness and particle size on
food utilization by mice, rats and hamsters. Lab Anim.
1977;11(4):241–246.
72.
FranklinCL. Microbial considerations in genetically engineered mouse
research. ILAR J.
2006;47(2):141–155.
73.
FrithCHWardJM. AColor Atlas of Neoplastic and Non Neoplastic Lesions in Aging Mice.
New York, NY: Elsevier;
1988.
74.
FuXBoothSLSmithDE. Vitamin K contents of rodent diets: a
review. J Am Assoc Lab Anim Sci.
2007;46(5):8–12.
75.
FullertonFRGreenmanDLKendallDC. Effects of storage conditions on nutritional qualities of
semipurified (AIN-76) and natural ingredient (NIH-07) diets. J
Nutr.
1982;112(3):567–573.
76.
FullertonFRGreenmanDLMcCartyCC. Increased incidence of spontaneous and
2-acetylaminofluorene-induced liver and bladder tumors in B6C3F1 mice fed AIN-76A diet
versus NIH-07 diet. Fundam Appl Toxicol.
1991;16(1):51–60.
77.
FullertonFRGreenmanDLBucciTJ. Effects of diet type on incidence of spontaneous and
2-acetylaminofluorene-induced liver and bladder tumors in BALB/c mice fed AIN-76A diet
versus NIH-07 diet. Fundam Appl Toxicol.
1992;18(2):193–199.
78.
GabrilMYYousefGM. Informatics for practicing anatomical pathologists:
marking a new era in pathology practice. Mod Pathol.
2010;23(3):349–358.
79.
GaoPDangCVWatsonJ. Unexpected antitumorigenic effect of fenbendazole when
combined with supplementary vitamins. J Am Assoc Lab Anim
Sci.
2008;47(6):37–40.
GoelzMFMahlerJHarryJ. Neuropathologic findings associated with seizures in FVB
mice. Lab Anim Sci.
1998;48(1):34–37.
82.
GopinathC. Spontaneous tumour rates: their use to support rodent
bioassays. Toxicol Pathol.
1994;22(2):160–164.
83.
GordonRRHunterKWLa MerrillM. Genotype X diet interactions in mice predisposed to
mammary cancer, II: tumors and metastasis. Mamm Genome.
2008;19(3):179–189.
84.
GreenmanDLFullertonFR. Comparison of histological responses of BALB/c and B6C3F1
female mice to estradiol when fed purified or natural-ingredient diets.
J Toxicol Environ Health.
1986;19(4):531–540.
85.
GreimHGelbkeHPReuterU. Evaluation of historical control data in carcinogenicity
studies. Hum Exp Toxicol.
2003;22(10):541–549.
86.
GutmannDHBakerSJGiovanniniM. Mouse models of human cancer consortium symposium on
nervous system tumors. Cancer Res.
2003;63(11):3001–3004.
87.
HainesDCChattopadhyaySWardJM. Pathology of aging B6;129 mice.
Toxicol Pathol.
2001;29(6):653–661.
HarrisonDERoderickTH. Selection for maximum longevity in mice.
Exp Gerontol.
1997;32(1–2):65–78.
90.
HarrisonDEStrongRSharpZD. Rapamycin fed late in life extends lifespan in genetically
heterogeneous mice. Nature.
2009;460(7253):392–395.
91.
HasemanJKHuffJBoormanGA. Use of historical control data in carcinogenicity studies
in rodents. Toxicol Pathol.
1984;12(2):126–135.
92.
HasemanJKBourbinaJEustisSL. Effect of individual housing and other experimental design
factors on tumor incidence in B6C3F1 mice. Fundam Appl
Toxicol.
1994;23(1):44–52.
93.
HasemanJKHaileyJRMorrisRW. Spontaneous neoplasm incidences in Fischer 344 rats and
B6C3F1 mice in two-year carcinogenicity studies: a National Toxicology Program
update. Toxicol Pathol.
1998;26(3):428–441.
94.
HasemanJK. Using the NTP database to assess the value of rodent
carcinogenicity studies for determining human cancer risk. Drug
Metab Rev.
2000;32(2):169–186.
95.
HasemanJKNeyENyskaA. Effect of diet and animal care/housing protocols on body
weight, survival, tumor incidences, and nephropathy severity of F344 rats in chronic
studies. Toxicol Pathol.
2003;31(6):674–681.
96.
HeindelJJvom SaalFS. Meeting report: batch-to-batch variability in estrogenic
activity in commercial animal diets—importance and approaches for laboratory animal
research. Environ Health Perspect.
2008;116(3):389–393.
97.
HenningsHGlickABLowryDT. FVB/N mice: an inbred strain sensitive to the chemical
induction of squamous cell carcinomas in the skin.
Carcinogenesis.
1993;14(11):2353–2358.
98.
HestonWE. Testing for possible effects of cedar wood shavings and
diet on occurrence of mammary gland tumors and hepatomas in C3H-A-vy and C3H-Avy-fB
mice. J Natl Cancer Inst.
1975;54(4):1011–1014.
99.
HoagWG. Spontaneous cancer in mice. Ann N
Y Acad Sci.
1963;108:805–831.
100.
HoenerhoffMJStarostMFWardJM. Eosinophilic crystalline pneumonia as a major cause of
death in 129S4/SvJae mice. Vet Pathol.
2006;43(5):682–688.
101.
HooijmansCRVan der ZeeCEDederenPJ. DHA and cholesterol containing diets influence
Alzheimer-like pathology, cognition and cerebral vasculature in APPswe/PS1dE9
mice. Neurobiol Dis.
2009;33(3):482–498.
102.
HuangPDudaDGJainRK. Histopathologic findings and establishment of novel tumor
lines from spontaneous tumors in FVB/N mice. Comp Med.
2008;58(3):253–263.
103.
IkenoYBronsonRTHubbardGB. Delayed occurrence of fatal neoplastic diseases in ames
dwarf mice: correlation to extended longevity. J Gerontol A Biol
Sci Med Sci.
2003;58(4):291–296.
104.
IkenoYHubbardGBLeeS. Reduced incidence and delayed occurrence of fatal
neoplastic diseases in growth hormone receptor/binding protein knockout
mice. J Gerontol A Biol Sci Med Sci.
2009;64(5):522–529.
105.
Institute for Laboratory Animal Research–National Research
Council. Guide for the Care and Use of Laboratory Animals.
8th ed. Washington, DC:
National Academies Press;
2010.
106.
Institute for Laboratory Animal Research, Committee on
Mammalian Models for Research on Aging. Mammalian Models for Research
on Aging. Washington, DC:
National Academies Press;
1981.
107.
Institute for Laboratory Animal Research.
1996. Guide for the Care and Use of Laboratory Animals.
7th ed. Washington, DC:
National Academies Press;
1996.
108.
Institute for Laboratory Animal Research.
Guidance for the Description of Animal Research in Scientific
Publications. Washington, DC:
National Academies Press;
2011.
InoueYIzawaKKiryuS. Diet and abdominal autofluorescence detected by in vivo
fluorescence imaging of living mice. Mol Imaging.
2008;7(1):21–27.
111.
ItoGMitaniSKimJH. Effect of soft diets on craniofacial growth in
mice. Anat Anz.
1988;165(2–3):151–166.
112.
JeffreyEGRobertCLotharH. Validation of transgenic mammary cancer models: goals of
the NCI mouse models of Human Cancer Consortium and the mammary cancer
CD-ROM. Transgenic Res.
2002;11(6):635–636.
113.
JiangLI, Nadeau JH. 129/Sv mice—a model system for studying germ
cell biology and testicular cancer. Mamm Genome.
2001;12(2):89–94.
114.
JovenJRullAFerreN. The results in rodent models of atherosclerosis are not
interchangeable: the influence of diet and strain.
Atherosclerosis.
2007;195(2):e85–e92.
115.
KatoHIwataTKatsuY. Evaluation of estrogenic activity in diets for
experimental animals using in vitro assay. J Agric Food
Chem.
2004;52(5):1410–1414.
116.
KeenanCHughes-EarleACaseM. The North American Control Animal database: a resource
based on standardized nomenclature and diagnostic criteria.
Toxicol Pathol.
2002;30(1):75–79.
117.
KeenanCElmoreSFrancke-CarrollS. Best practices for use of historical control data of
proliferative rodent lesions. Toxicol Pathol.
2009;37(5):679–693.
118.
KeenanCElmoreSFrancke-CarrollS. Potential for a global historical control database for
proliferative rodent lesions. Toxicol Pathol.
2009;37(5):677–678.
119.
KeenanKPBallamGCSoperKA. Diet, caloric restriction, and the rodent
bioassay. Toxicol Sci.
1999;52(2)(suppl):24–34.
120.
KerrLRAndrewsHNStrangeKS. Temporal factors alter effects of social housing
conditions on responses to chemotherapy and hormone levels in a Shionogi mammary tumor
model. Psychosom Med.
2006;68(6):966–975.
121.
KilkennyCParsonsNKadyszewskiE. Survey of the quality of experimental design, statistical
analysis and reporting of research using animals. PLoS
One.
2009;4(11):e7824.
122.
KilkennyCAltmanDG. Improving bioscience research reporting: ARRIVE-ing at a
solution. Lab Anim.
2010;44:377–378.
123.
KilkennyCBrowneWJCuthillIC. Improving bioscience research reporting: the ARRIVE
guidelines for reporting animal research. PLoS Biol.
2010;8(6):e1000412.
124.
KirklandJLPetersonC. Healthspan, translation, and new outcomes for animal
studies of aging. J Gerontol A Biol Sci Med Sci.
2009;64(2):209–212.
125.
KittelBRuehl-FehlertCMorawietzG. Revised guides for organ sampling and trimming in rats and
mice—part 2. A joint publication of the RITA and NACAD groups.
Exp Toxicol Pathol.
2004;55(6):413–431.
Online details athttp://reni.item.fraunhofer.de/reni/trimming/index.php
126.
KlopfleischRWeissATGruberAD. Excavation of a buried treasure—DNA, mRNA, miRNA and
protein analysis in formalin fixed, paraffin embedded tissues.
Histol Histopathol.
2011;26(6):797–810.
127.
KodellRLFarmerJHGreenmanDLFrithCH. Estimation of distributions of time to appearance of tumor
and time to death from tumor after appearance in mice fed
2-acetylaminofluorene. J Environ Pathol Toxicol.
1980;3(3, spec
no):89–102.
128.
KodellRLBlackwellBNBucciTJ. Cause-of-death assignment at the National Center for
Toxicological Research. Toxicol Pathol.
1995;23(3):241–247.
129.
KoganSCWardJMAnverMR. Bethesda proposals for classification of nonlymphoid
hematopoietic neoplasms in mice. Blood.
2002;100(1):238–245.
130.
KrupkeDMBegleyDASundbergJP. The Mouse Tumor Biology database.
Nat Rev Cancer.
2008;8(6):459–465.
LazarovORobinsonJTangYP. Environmental enrichment reduces Abeta levels and amyloid
deposition in transgenic mice. Cell.
2005;120(5):701–713.
134.
Le CalvezSPerron-LepageMFBurnettR. Subcutaneous microchip-associated tumours in B6C3F1 mice:
a retrospective study to attempt to determine their histogenesis.
Exp Toxicol Pathol.
2006;57(4):255–265.
135.
LichtmanAHClintonSKIiyamaK. Hyperlipidemia and atherosclerotic lesion development in
LDL receptor-deficient mice fed defined semipurified diets with and without
cholate. Arterioscler Thromb Vasc Biol.
1999;19(8):1938–1944.
136.
LinderCC. The influence of genetic background on spontaneous and
genetically engineered mouse models of complex diseases. Lab
Anim (NY).
2001;30(5):34–39.
137.
LinderCC. Mouse nomenclature and maintenance of genetically
engineered mice. Comp Med.
2003;53(2):119–125.
138.
LipmanRDGaillardETHarrisonDE. Husbandry factors and the prevalence of age-related
amyloidosis in mice. Lab Anim Sci.
993;43(5):439–444.
139.
LipmanRDSmithDEBlumbergJB. Effects of caloric restriction or augmentation in adult
rats: longevity and lesion biomarkers of aging. Aging
(Milano).
1998;10(6):463–470.
140.
LittleCC. The genetics of spontaneous tumor
formation. In: SnellGDBittnerJJHestonWE, eds. The Biology of the Laboratory Mouse.
Philadelphia, PA:
Blakiston;
1941:248–278.
141.
LoscoPE. Dental dysplasia in rats and mice.
Toxicol Pathol.
1995;23(6):677–688.
142.
LuijtenMThomsenARvan den BergJA. Effects of soy-derived isoflavones and a high-fat diet on
spontaneous mammary tumor development in Tg.NK (MMTV/c-neu) mice.
Nutr Cancer.
2004;50(1):46–54.
143.
MahlerJFStokesWMannPC. Spontaneous lesions in aging FVB/N mice.
Toxicol Pathol.
1996;24(6):710–716.
144.
MarashiVBarnekowAOssendorfE. Effects of different forms of environmental enrichment on
behavioral, endocrinological, and immunological parameters in male mice.
Horm Behav.
2003;43(2):281–292.
145.
MarashiVBarnekowASachserN. Effects of environmental enrichment on males of a docile
inbred strain of mice. Physiol Behav.
2004;82(5):765.
146.
MaronpotRRBoormanGAGaulBW, eds. Pathology of the Mouse: Reference and Atlas.
Vienna, IL: Cache River
Press; 1999.
147.
MartinBJiS, Maudsley S, et al. “Control” laboratory rodents are
metabolically morbid: why it matters. Proc Natl Acad Sci U S
A.
2010;107(14):6127–6133.
148.
MasoroEJ. Use of rodents as models for the study of “normal aging”:
conceptual and practical issues. Neurobiol Aging.
1991;12(6):639–643.
149.
MasoroEJ. Overview of caloric restriction and
ageing. Mech Ageing Dev.
2005;126(9):913–922.
150.
MasoroEJ. Caloric restriction-induced life extension of rats and
mice: a critique of proposed mechanisms. Biochim Biophys
Acta.
2009;1790(10):1040–1048.
151.
McClearnGE. Heterogeneous stocks and selective breeding in aging
research. ILAR J.
2011;52(1):16–23.
152.
McConnellEE.Pathology requirements for two-year rodent studies, II: alternative
approaches. Toxicol Pathol.
1983;11(1):65–76.
153.
McLinJPStewardO. Comparison of seizure phenotype and neurodegeneration
induced by systemic kainic acid in inbred, outbred, and hybrid mouse
strains. Eur J Neurosci.
2006;24(8):2191–2202.
154.
MikaelianIIchikiTWardJM. Diversity of spontaneous neoplasms in commonly used inbred
strains and stocks of laboratory mice. In: HedrichHJ, ed. The Laboratory Mouse.
London: Elsevier Academic
Press;
2004:345–354.
155.
MillerRA. Principles of animal use in gerontological
research. In: ConnPM, ed. Handbook of Models for Human Aging.
Amsterdam: Elsevier Academic
Press;
2006:21–31.
156.
MillerRAHarrisonDEAstleCM. An aging interventions testing program: study design and
interim report. Aging Cell.
2007;6(4):565–575.
157.
MitchellCRKemptonJBScott-TylerB. Otitis media incidence and impact on the auditory brain
stem response in lipopolysaccharide-nonresponsive C3H/HeJ mice.
Otolaryngol Head Neck Surg.
1997;117(5):459–464.
158.
MohajeriMHMadaniRSainiK. The impact of genetic background on neurodegeneration and
behavior in seizured mice. Genes Brain Behav.
2004;3(4):228–239.
159.
MohrUDungworthDLWardJM, eds. Pathobiology of the Aging Mouse.
Washington, DC: ILSI
Press; 1996.
160.
MooreM; IMPC Steering Committee. The International Mouse Phenotyping
Consortium: Initial Business Plan. Bethesda, MD:
National Institutes of Health;
2010.
161.
MorawietzGRuehl-FehlertCKittelB. Revised guides for organ sampling and trimming in rats and
mice—part 3. A joint publication of the RITA and NACAD groups.
Exp Toxicol Pathol.
2004;55(6):433–449.
Online information athttp://reni.item.fraunhofer.de/reni/trimming/index.php
162.
MorseHC IIIAnverMRFredricksonTN. Bethesda proposals for classification of lymphoid
neoplasms in mice. Blood.
2002;100(1):246–258.
163.
MorseHCI. Origins of Inbred Mice. New York,
NY: Academic Press;
1978.
164.
MortonDKempRKFrancke-CarrollS. Best practices for reporting pathology interpretations
within GLP toxicology studies. Toxicol Pathol.
2006;34(6):806–809.
165.
MortonDSellersRSBarale-ThomasE. Recommendations for pathology peer review.
Toxicol Pathol.
2010;38(7):1118–1127.
166.
MurphyED. Characteristic tumors. In: GreenE, ed. Biology of the Laboratory Mouse; by the Staff of the Jackson
Laboratory. 2nd ed. New York,
NY: Dover;
1966.
167.
NadonNLStrongRMillerRA. Design of aging intervention studies: the NIA
interventions testing program. Age (Dordr).
2008;30(4):187–199.
168.
NagyTRKrzywanskiDLiJ. Effect of group vs. single housing on phenotypic variance
in C57BL/6J mice. Obes Res.
2002;10(5):412–415.
NietoAIShyamalaGGalvezJJ. Persistent mammary hyperplasia in FVB/N
mice. Comp Med.
2003;53(4):433–438.
171.
NikitinAYAlcarazAAnverMR. Classification of proliferative pulmonary lesions of the
mouse: recommendations of the mouse models of human cancers consortium.
Cancer Res.
2004;64(7):2307–2316.
National Toxicology Program. Effect
of dietary restriction on toxicology and carcinogenesis studies in F344/N rats and
B6C3F1 mice. Natl Toxicol Program Tech Rep Ser.
1997;460:1–414.
174.
PaigenBPetersLLYuanR. Mice as a mammalian model for research on the genetics of
aging. ILAR J.
2011;52(1):4–15.
175.
PercyDHBartholdSW. Pathology of Laboratory Rodents and Rabbits.
3rd ed. New York, NY:
Wiley-Blackwell; 2007.
176.
Pettan-BrewerCTreutingPM. Practical pathology of aging mice.
Pathobiol Aging Age Relat Dis.
2011;1:7202.
177.
PhilipVMSokoloffGAckert-BicknellCL. Genetic analysis in the Collaborative Cross breeding
population. Genome Res.
2011;21(8):1223–1238.
178.
PottsSJ. Digital pathology in drug discovery and development:
multisite integration. Drug Discov Today.
2009;14(19–20):935–941.
179.
RadaelliEArnoldAPapanikolaouA. Mammary tumor phenotypes in wild-type aging female FVB/N
mice with pituitary prolactinomas. Vet Pathol.
2009;46(4):736–745.
180.
RaoGNEdmondsonJ. Tissue reaction to an implantable identification device in
mice. Toxicol Pathol.
1990;18(3):412–416.
181.
RaoGNHasemanJKGrumbeinS. Growth, body weight, survival, and tumor trends in
(C57BL/6 × C3H/HeN) F1 (B6C3F1) mice during a nine-year period.
Toxicol Pathol. 1990;18(1, pt
1):71–77.
182.
RaoGNCrockettPW. Effect of diet and housing on growth, body weight,
survival and tumor incidences of B6C3F1 mice in chronic studies.
Toxicol Pathol.
2003;31(2):243–250.
183.
RayMAJohnstonNAVerhulstS. Identification of markers for imminent death in mice used
in longevity and aging research. J Am Assoc Lab Anim
Sci.
2010;49(3):282–288.
184.
ReaganWJIrizarry-RoviraAPoitout-BelissentF. Best practices for evaluation of bone marrow in
nonclinical toxicity studies. Vet Clin Pathol.
2011;40(2):119–134.
185.
ReevesPGNielsenFHFaheyGC Jr. AIN-93 purified diets for laboratory rodents: final report
of the American Institute of Nutrition ad hoc writing committee on the reformulation of
the AIN-76A rodent diet. J Nutr.
1993;123(11):1939–1951.
186.
RelieneRSchiestlRH. Differences in animal housing facilities and diet may
affect study outcomes: a plea for inclusion of such information in
publications. DNA Repair (Amst).
2006;5(6):651–653.
187.
RenneRBrixAHarkemaJ. Proliferative and nonproliferative lesions of the rat and
mouse respiratory tract. Toxicol Pathol.
2009;37(7)(suppl):5S–73S.
188.
RobertsonEJ. Using embryonic stem cells to introduce mutations into the
mouse germ line. Biol Reprod.
1991;44(2):238–245.
189.
RoeFJ. Historical histopathological control data for laboratory
rodents: valuable treasure or worthless trash?Lab Anim.
1994;28(2):148–154.
190.
Ruehl-FehlertCKittelBMorawietzG. Revised guides for organ sampling and trimming in rats and
mice—part 1. Exp Toxicol Pathol.
2003;55(2–3):91–106.
Online information athttp://reni.item.fraunhofer.de/reni/trimming/index.php
191.
SabineJR. Exposure to an environment containing the aromatic red
cedar, Juniperus virginiana: procarcinogenic, enzyme-inducing and
insecticidal effects. Toxicology.
1975;5(2):221–235.
192.
SchofieldPNDubusPKleinL. Pathology of the laboratory mouse: an International
Workshop on Challenges for High Throughput Phenotyping. Toxicol
Pathol.
2011;39(3):559–562.
SellersR. The gene or not the gene—that is the question:
understanding the genetically engineered mouse phenotype. Vet
Pathol. 2011Oct4. [Epub ahead of print]
195.
SelmanCWithersDJ. Mammalian models of extended healthy
lifespan. Philos Trans R Soc Lond B Biol Sci.
2011;366(1561):99–107.
196.
SeongESaundersTLStewartCL. To knockout in 129 or in C57BL/6: that is the
question. Trends Genet.
2004;20(2):59.
197.
ShappellSBThomasGVRobertsRL. Prostate pathology of genetically engineered mice:
definitions and classification. The consensus report from the Bar Harbor meeting of the
Mouse Models of Human Cancer Consortium Prostate Pathology Committee.
Cancer Res.
2004;64(6):2270–2305.
198.
SiegelV. Reproducibility in research. Dis
Models Mech.
2011;4(3):279–280.
199.
SlaouiMFietteL. Histopathology procedures: from tissue sampling to
histopathological evaluation. Methods Mol Biol.
2011;691:69–82.
200.
SouthardT, Brayton CF. spontaneous unilateral brainstem infarction in
Swiss mice. Vet Pathol.
2011;48(3):726–729.
201.
StarkDM. Wire-bottom versus solid-bottom rodent caging issues
important to scientists and laboratory animal science specialists.
Contemp Top Lab Anim Sci.
2001;40(6):11–14.
202.
StorerJB. Longevity and gross pathology at death in 22 inbred mouse
strains. J Gerontol.
1966;21(3):404–409.
203.
Society of Toxicologic Pathologists.
Peer review in toxicologic pathology: some
recommendations. Toxicol Pathol.
1991;19(3):290–292.
204.
Society of Toxicologic Pathologists.
Documentation of pathology peer review: position of the Society of
Toxicologic Pathologists. Toxicol Pathol.
1997;25(6):655.
205.
StrangeKSKerrLRAndrewsHN. Psychosocial stressors and mammary tumor growth: an animal
model. Neurotoxicol Teratol.
2000;22(1):89–102.
206.
SundbergBASchofieldPNGruenbergerM. A data-capture tool for mouse pathology
phenotyping. Vet Pathol.
2009;46(6):1230–1240.
SundbergJPSundbergBASchofieldP. Integrating mouse anatomy and pathology ontologies into a
phenotyping database: tools for data capture and training. Mamm
Genome.
2008;19(6):413–419.
209.
SundbergJPSchofieldPN. Commentary: mouse genetic nomenclature. Standardization of
strain, gene, and protein symbols. Vet Pathol.
2010;47(6):1100–1104.
210.
SundbergJPWardJMHogeneschH. Training pathologists in mouse pathology.
Vet Pathol. 2010 Sep3. [Epub ahead of print]
211.
SundbergJPBerndtASundbergBA. The mouse as a model for understanding chronic diseases of
aging: the histopathologic basis of aging in inbred mice.
Pathobiol Aging Age Relat Dis.
2011;1:7179.
212.
TaketoMSchroederACMobraatenLE. FVB/N: an inbred mouse strain preferable for transgenic
analyses. Proc Natl Acad Sci U S A.
1991;88(6):2065–2069.
213.
TaroneREChuKCWardJM. Variability in the rates of some common naturally
occurring tumors in Fischer 344 rats and (C57BL/6N × C3H/HeN)F1 (B6C3F1)
mice. J Natl Cancer Inst.
1981;66(6):1175–1181.
214.
TatarMBartkeAAntebiA. The endocrine regulation of aging by insulin-like
signals. Science.
2003;299(5611):1346–1351.
215.
TatarM. Can we develop genetically tractable models to assess
healthspan (rather than life span) in animal models?J Gerontol A Biol Sci Med Sci.
2009;64(2):161–163.
216.
ThigpenJELocklearJRominesC. A standard procedure for measuring pellet hardness of
rodent diets. Lab Anim Sci.
1993;43(5):488–491.
217.
ThigpenJELocklearJHasemanJK. Effects of the dietary phytoestrogens daidzein and
genistein on the incidence of vulvar carcinomas in 129/J mice.
Cancer Detect Prev.
2001;25(6):527–532.
218.
ThigpenJESetchellKDSaundersHE. Selecting the appropriate rodent diet for endocrine
disruptor research and testing studies. ILAR J.
2004;45(4):401–416.
ThoolenBMaronpotRRHaradaT. Proliferative and nonproliferative lesions of the rat and
mouse hepatobiliary system. Toxicol Pathol.
2010;38(7)(suppl):5S–81S.
221.
ThreadgillDWMatinAYeeD. SSLPs to map genetic differences between the 129 inbred
strains and closed-colony, random-bred CD-1 mice. Mamm
Genome.
1997;8(6):441–442.
222.
ThreadgillDWPardo-ManueldeVillenaFMillerDR. The collaborative cross: a recombinant inbred mouse
population for the systems genetic era. ILAR J.
2011;52(1):24–31.
223.
TillmannTKaminoKDasenbrockC. Subcutaneous soft tissue tumours at the site of implanted
microchips in mice. Exp Toxicol Pathol.
1997;49(3–4):197–200.
224.
TreutingPCliffordCBSellersR. Of mice and microflora: considerations for genetically
engineered mice. Vet Pathol. In
press.
225.
TreutingPMLinfordNJKnoblaughSE. Reduction of age-associated pathology in old mice by
overexpression of catalase in mitochondria. J Gerontol A Biol
Sci Med Sci.
2008;63(8):813–822.
226.
TsaiPPPachowskyUStelzerHD. Impact of environmental enrichment in mice, 1: effect of
housing conditions on body weight, organ weights and haematology in different
strains. Lab Anim.
2002;36(4):411–419.
227.
TwaddleNCChurchwellMIMcDanielLP. Autoclave sterilization produces acrylamide in rodent
diets: implications for toxicity testing. J Agric Food
Chem.
2004;52(13):4344–4349.
228.
Ullman-CullereMHFoltzCJ. Body condition scoring: a rapid and accurate method for
assessing health status in mice. Lab Anim Sci.
1999;49(3):319–323.
229.
VahleJBradleyAHaradaT. The international nomenclature project: an
update. Toxicol Pathol.
2009;37(5):694–697.
230.
Van LooPLVan de WeerdHAVan ZutphenLF. Preference for social contact versus environmental
enrichment in male laboratory mice. Lab Anim.
2004;38(2):178–188.
231.
VetvickaVHolubMHoustekJ. Effect of ambient temperature on immune functions of
athymic nude mice. Thymus.
1993;21(1):11–23.
232.
VlahakisG. Possible carcinogenic effects of cedar shavings in bedding
of C3H-Avy fB mice. J Natl Cancer Inst.
1977;58(1):149–150.
233.
VolkMJPughTDKimM. Dietary restriction from middle age attenuates
age-associated lymphoma development and interleukin 6 dysregulation in C57BL/6
mice. Cancer Res.
1994;54(11):3054–3061.
234.
WakefieldLMThordarsonGNietoAI. Spontaneous pituitary abnormalities and mammary
hyperplasia in FVB/NCr mice: implications for mouse modeling.
Comp Med.
2003;53(4):424–432.
WardJMAnverMRMahlerJF. Pathology of mice commonly used in genetic engineering
(C57BL/6; 129; B6;129; FVB). In: WardJMMaronpotRRSundbergJP, eds. Pathology of Genetically Engineered Mice.
Ames: Iowa State University
Press;
2000:161–179.
237.
WardJM. Lymphomas and leukemias in mice.
Exp Toxicol Pathol.
2006;57(5–6):377.
238.
WardJMThoolenB. Grading of lesions. Toxicol
Pathol.
2011;39(4):745–746.
239.
WaterstonRHLindblad-TohKBirneyE. Initial sequencing and comparative analysis of the mouse
genome. Nature.
2002;420(6915):520–562.
240.
WebsterLTPritchettIW. Microbic virulence and host susceptibility in
paratyphoid-enteritidis infection of white mice, V: the effect of diet on host
resistance. J Exp Med.
1924;40(3):397–404.
241.
WeindruchRWalfordRL. Dietary restriction in mice beginning at 1 year of age:
effect on life-span and spontaneous cancer incidence.
Science.
1982;215(4538):1415–1418.
242.
WeindruchR. The retardation of aging by caloric restriction: studies
in rodents and primates. Toxicol Pathol.
1996;24(6):742–745.
243.
WolfDCMannPC. Confounders in interpreting pathology for safety and risk
assessment. Toxicol Appl Pharmacol.
2005;202(3):302–308.
244.
WolfNSGiddensWEMartinGM. Life table analysis and pathologic observations in male
mice of a long-lived hybrid strain (Af × C57BL/6)F1. J
Gerontol.
1988;43(3):B71–B78.
245.
WolffCSchottCPorschewskiP. Successful protein extraction from over-fixed and
long-term stored formalin-fixed tissues. PLoS One.
2011;6(1):e16353.
246.
WongAABrownRE. Visual detection, pattern discrimination and visual acuity
in 14 strains of mice. Genes Brain Behav.
2006;5(5):389–403.
247.
WotjakCT. C57BLack/BOX? The importance of exact mouse strain
nomenclature. Trends Genet.
2003;19(4):183–184.
248.
WurstWde AngelisMH. Systematic phenotyping of mouse mutants.
Nat Biotechnol.
2010;28(7):684–685.
249.
YamamotoHFujiiKHayakawaT. Inhibitory effect of cold stress on lung tumours induced
by 7,12-dimethylbenz[a]anthracene in mice. J Cancer Res Clin
Oncol.
1995;121(7):393–396.
250.
YamamotoHFujiiKHayakawaT. Inhibitory effect of cold stress against
acetaminophen-induced hepatic injury in B6C3F1 and ICR mice.
Toxicol Lett.
1995;81(2–3):125–130.
251.
YoungJKHallRLO’BrienP. Best practices for clinical pathology testing in
carcinogenicity studies. Toxicol Pathol.
2011;39(2):429–434.
252.
YuanRTsaihS-WPetkovaSB. Aging in inbred strains of mice: study design and interim
report on median lifespans and circulating IGF1 levels. Aging
Cell.
2009;8(3):277–287.
253.
YuanRTsaihSWPetkovaSB. Aging in inbred strains of mice: study design and interim
report on median lifespans and circulating IGF1 levels. Aging
Cell.
2009;8(3):277–287.
254.
YunJPBehanJWHeisterkampN. Diet-induced obesity accelerates acute lymphoblastic
leukemia progression in two murine models. Cancer Prev Res
(Phila).
2010;3(10):1259–1264.
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.