
Abstract
Prenatal exposure to endogenous or exogenous androgens alters the development of the female reproductive tract. Although lesions in ovaries and external genitalia of androgenized female sheep have been reported, lesions of the tubular genitalia have not. Testosterone propionate (TP) or dihydrotestosterone (DHT) was administered by intramuscular injection twice weekly to 32 ewes from 30 to 90 days of pregnancy. The ewes lambed normally. The reproductive tracts from 24 treated and 13 control postpubertal female offspring were examined at 10 months of age. The ovaries, oviducts, and uteri were grossly and histologically normal in both TP- and DHT-exposed sheep. However, in the DHT-treated sheep, the uterus connected to a misshapen, saccular vagina that opened into the urethra; in the TP-treated sheep, it ended in a blind sac. In both TP- and DHT-treated sheep, the urethra was approximately 5 times longer than that of control sheep, and it resembled a male urethra with bilateral male accessory genital glands. The urethra terminated in a fully developed penis in both TP- and DHT-treated sheep, and a scrotal sac was present (without testes). These results show that prenatal exposure of female sheep to exogenous androgens results in masculinization of the tubular and external genitalia.
Keywords
In sheep, as in other mammals, gonadal differentiation is determined by the interaction of several genes, particularly the Sry gene on the Y chromosome. 17,24,26 The Sry gene encodes the transcription factor known as the testis-determining factor that upregulates the expression of other genes, including Sox9 in Sertoli cell precursors. 15 In turn, Sox9 upregulates the expression of growth factors that induce testicular differentiation and development, such as fibroblast growth factor 9. 15 The expression of certain hormones, after gonadal sex has been genetically determined, can result in masculinization of the ovary. For example, anti-Müllerian hormone can induce the formation of seminiferous cords within the ovary, and 5α-dihydrotestosterone (DHT) increases ovarian androgen receptor. 20,24
Sox9 transcription promotes production of anti-Müllerian hormone by Sertoli cells in the fetal testes, which causes regression of the female ductal system (paramesonephric or Müllerian). 26 In male fetuses, testosterone produced by testicular interstitial (Leydig) cells and expression of transcription factors, such as Pax2, Pax8, Lim1, and Emx2, promote development of the mesonephric (Wolffian) ductal system into the epididymis, vas deferens, and accessory genital glands. 10,26 5α-Reductase produced by the epididymis and the accessory genital glands converts testosterone to DHT, which in turn induces stabilization of the ductal system and development of male external genitalia. 10 In the absence of these stimuli, the urogenital ridge develops into the vagina, which connects to the uterus as it develops from the paramesonephric ductal system.
Abnormal sex determination can manifest as an ovotesticular disorder of sexual development (DSD), in which the animal has both male and female gonadal tissue, a condition previously termed true hermaphrodism or intersex. 13,26 Freemartinism in cattle is an example of an ovotesticular DSD in which a female fetus shares placental circulation with a male twin and becomes an XX/XY blood chimera. 22,26 Freemartin calves have variable masculinization of the reproductive tract; many have ovotestes, partial accessory genital gland development, abnormal vesicular glands, and hypoplastic female tubular structures. Although about 90% of female calves with male twins are freemartins, 16,22 freemartinism in sheep is much less common, affecting only 1% of females that are twins to males. 1,22 The rate of freemartinism in sheep has increased with the more numerous lambs/litter following the introduction of high-fecundity genes. 2,22 As in cattle, freemartin sheep often have gonads that appear as cordlike thickenings or resemble testes, as well as variable female tubular development. 22,28,29
In male mammals, prenatal exposure to androgens is responsible for the normal development of gender-specific tubular genitalia, 26 whereas exposure of female fetuses to elevated concentrations of androgens can lead to abnormal development of tubular genitalia. 26 Androgenized female fetuses have female gonads, but variable development of both male and female tubular and external genitalia, a condition termed XX DSD (previously termed female pseudohermaphrodism). 13,26 Severe maldevelopment results in infertility. 26
The effect of environmental steroids (“endocrine disruptors”) that can bind and activate receptors on reproductive organs has received global attention. 12 The increased incidence of human reproductive abnormalities, including testicular dysgenesis and polycystic ovary syndromes, with associated infertility 6,19,32 has been attributed to prenatal exposure to these endocrine disrupters. 3,27 In sheep, abnormalities of the reproductive, metabolic, and growth axes can be induced by exposure to abnormal steroid concentrations or environmental endocrine disrupters during vulnerable stages of in utero development. 11,18,19,21,23,34
Prenatal exposure to excess androgens between days 30 and 90 of gestation has a virilizing effect on the external genitalia of female lambs, and affected animals have a penis and a scrotal sac without testes. 4,14 The ovaries are in the correct anatomical position but have gross abnormalities soon after birth. 5,30,35–37 The degree of masculinization of the external genitalia is proportional to the duration and timing of androgen exposure, and it depends on the type of androgen. 25,36 The purpose of this study is to describe the gross and histologic changes of tubular genitalia after in utero androgen exposure of female sheep.
Materials and Methods
Animals
Poll Dorset ewes were born from April 9 to May 10, 2008, and maintained at the University of Glasgow farm (55°55′N). Studies on the reproductive, growth, and metabolic axes were approved by the university’s Welfare and Ethics Committee and carried out under Home Office License PPL 60/3485.
Treatments
Fetuses were exposed to testosterone propionate (TP) or the nonaromatizable androgen DHT, as reported. 31 Briefly, the reproductive cycles in 56 ewes were synchronized using an intravaginal CIDR device (Inter Ag, New Zealand), and the ewes were mated to a Poll Dorset ram. Mating was monitored by fitting the ram with a marking harness. Pregnancy was assessed by ultrasound at about 60 days postmating, at which point 48 of 56 ewes were confirmed to be pregnant. Before confirmation of pregnancy, ewes were randomly allocated to treatment groups to receive TP, DHT, or no injections as follows.
Of the 48 pregnant ewes, 32 were given twice-weekly intramuscular injections of 100 mg of TP (n = 17) or DHT (n = 15) in vegetable oil. Injections began at day 30 after mating and continued until day 90 of pregnancy (term, 147 days). 34 Sixteen pregnant ewes served as controls and did not receive any injections. Data from a similar study in sheep have shown that this treatment regime raises androgen concentrations in maternal blood to values like those in adult males (about 8 ng/mL). 34 Umbilical arterial testosterone concentration at days 65 and 90 of gestation of TP-treated female fetuses was about 0.5 ng/mL, comparable to that in male fetuses. 34 Control dams were not treated, because we and others have found no differences between vehicle-treated and nontreated controls. 31 The 32 treated ewes delivered a total of 73 live lambs: 29 male and 44 female. Four female lambs (n = 1 TP- and 3 DHT-treated animals) of low birth weight (< 2 kg) died soon after birth. Two TP- and 1 DHT-treated lamb died of unknown cause before completion of the study. The reproductive tracts from these 7 lambs were not examined. The remaining 37 female lambs were euthanized (Somulose, 1 mL/kg body weight, Dechra Veterinary Products, Shrewsbury, United Kingdom) at approximately 10 months of age, following puberty (at approximately 7 months of age) and during the anestrous season. The reproductive tracts from all 37 animals were examined grossly.
The ovaries of 15 TP-treated, 9 DHT-treated, and 13 control lambs were removed, immersion fixed in 10% neutral buffered formalin, and examined histologically for evidence of male gonadal structures. 8,9 The entire tubular and external genitalia of 2 TP-treated, 6 DHT-treated, and 2 control lambs were similarly fixed. After fixation, the urethras were measured from the external orifice to the connection with the urinary bladder. Representative sections of each segment of the reproductive tract were routinely processed for histologic examination. Paraffin-embedded, 4- to 5-μm-thick sections were stained with hematoxylin and eosin.
Results
The distal tubular and external genitalia of the 24 TP- or DHT-treated female lambs were strikingly abnormal, resembling those of a ram. Each lamb had male external genitalia with a fully formed penis and an empty scrotal sac (Fig. 1). The urethra was elongated with 2 distinct curves, resembling the sigmoid flexure of a ram. Urethral length was 27 to 33 cm (29.17 ± 2.14) in treated sheep, compared to 4 to 5 cm in the control female lambs. Histologically, the proximal portion of the urethra was surrounded by dense layers of muscle and had a lumen lined by transitional epithelium.
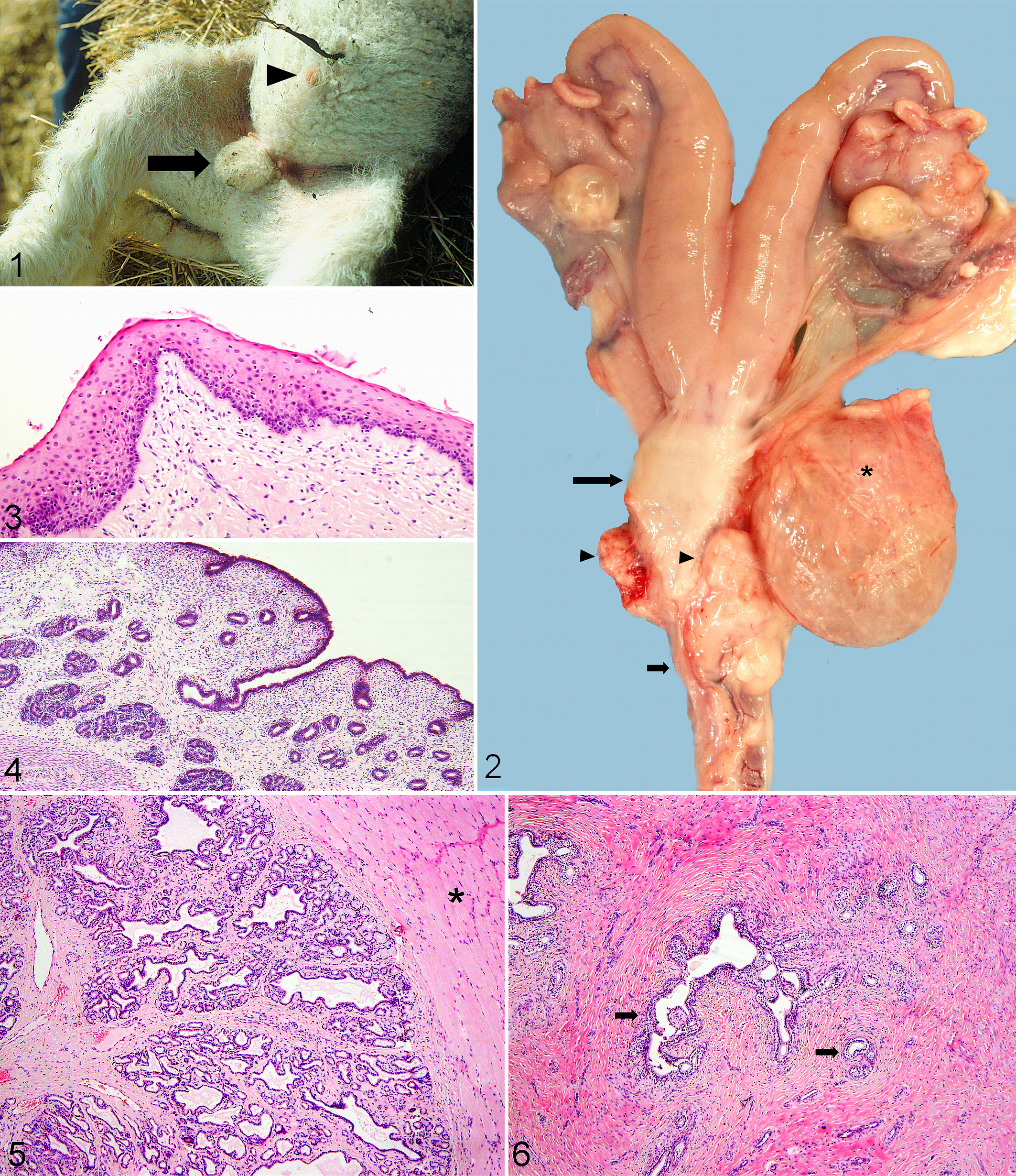
External genitalia; female lamb treated in utero with dihydrotestosterone (DHT). Note preputial orifice (arrowhead) and scrotal sac (arrow).
Just proximal to the urethral junction with the urinary bladder, a dilated sac, interpreted as a malformed vagina (Fig. 2), was filled with variable amounts of white to tan, thick, mucoid material. In DHT-treated sheep, the sac communicated with the urethra through a narrow opening, but a distinct vaginal vestibule was not apparent. In TP-treated sheep, the sac was blind-ended, with no communication with the urethra. In 8 DHT- or TP-treated animals examined histologically, this sac resembled normal vagina with stratified squamous epithelium surrounded by a thick wall of smooth muscle (Fig. 3).
The vaginal sac communicated with a cervix. Proximal to the cervix, the tubular genitalia of the TP- and DHT-treated lambs were grossly (n = 24; Fig. 2) and histologically (n = 8) normal, resembling those of the control female lambs. Oviducts and a well-developed uterus were in the correct anatomical location. Histologically, the uterus was lined by cuboidal to columnar epithelium with endometrial glands in the lamina propria (Fig. 4). Retained proximal mesonephric (Wolffian) ducts were found in the uterine broad ligament of all 24 androgenized lambs examined grossly but none of the 13 control lambs.
The 15 TP- and 9 DHT-treated lambs had ovaries that were of normal gross appearance and anatomic location. The lack of testicular structures or histologic features of male gonadal tissue in TP- or DHT-treated sheep makes an ovotesticular DSD (such as freemartinism) unlikely. The presence of this change in female offspring with all female siblings is further proof that these changes are not related to freemartinism.
Along the tubular genitalia, 2 firm, bilaterally symmetrical glandular structures were found in all 24 treated sheep examined grossly. Approximately midway along the urethra, just proximal to the sigmoid flexure, slender glands, interpreted as accessory genital glands, tightly straddled the urethra. Histologically, acini were lined by cuboidal epithelium, and ducts, by stratified squamous epithelium (Fig. 5). Scanty proteinaceous secretion was in glandular lumens. The glands were in thin fibrous stroma; peripherally, there was abundant smooth muscle. The more proximal set of glands straddled the distal portion of the vaginal sac just cranial to its junction with the urethra (Fig. 2). These glands were tortuous, lined by vacuolated columnar epithelium and supported by abundant fibrous stroma (Fig. 6).
Discussion
The presence of male tubular structures (including persistent mesonephric ducts) and male external genitalia in the TP- and DHT-treated sheep indicates extensive masculinization of the female reproductive tract. In the TP- and DHT-treated sheep, the development of the paramesonephric (Müllerian) ducts was not inhibited, allowing full formation of the oviducts and uterus. This is in contrast to freemartinism in sheep in which 68% of XX/XY blood chimeras lack development of the paramesonephric ductal system and have testicular tissue. 29
Interestingly, although TP and DHT function at different times in the development and differentiation of the reproductive organs, the distal tubular genitalia, accessory genital glands, and external genitalia were similar in both treatment groups. The only difference between these 2 groups was that the TP-treated sheep had vaginal structures that ended in a blind sac with no communication with the urethra. This lack of communication could represent a failure of fusion of the paramesonephric ductal system and urogenital ridge.
As in normal fetal male development, the presence of circulating testosterone during gestation of the TP-treated sheep resulted in partial formation of the male tubular genitalia, including retained mesonephric (Wolffian) ducts and formation of a long, masculine urethra complete with a sigmoid flexure and poorly developed accessory genital glands. The external genitalia were male in these sheep, with a fully formed penis and scrotal sac. The masculinization of the external genitalia in TP-treated sheep suggests a response to exogenous DHT or that the accessory glands had produced α-reductase and converted testosterone to DHT.
The presence of rudimentary accessory genital glands in the TP- and DHT-treated sheep resembles that of freemartin sheep. 29 Rams typically have 4 accessory genital glands: the ampullae, seminal vesicles, prostate, and bulbourethral glands. The ampullae and seminal vesicles are tightly associated and straddle the urethra at its junction with the urinary bladder and ductus deferens. The paired ampullae are slender and ovoid, whereas seminal vesicles are more rounded and situated slightly caudal to the ampullae. The prostate, situated caudal to the seminal vesicles, is flattened and dispersed. The paired bulbourethral glands are more concentrated and rounded, forming discrete protruding structures.
In the TP- and DHT-treated female lambs, one set of male accessory genital glands was proximal to the junction of the urinary bladder with the reproductive tract where it joined the urethra. Based on location, this set of glands probably represents either rudimentary ampullae or seminal vesicles. Based on their large and bulbous nature as well as the ventral position, the glands more closely resemble seminal vesicles. The ampullae and seminal vesicles of normal male sheep are predominantly epithelial, with little connective tissue, and are difficult to distinguish histologically. The glands in the treated female sheep were rudimentary with abundant collagen, more like those of castrated male sheep, which have extensive atrophy of the ampullae and seminal vesicles. 33 The lack of testosterone after birth may have prevented further development or resulted in atrophy of these glands.
The second set of glandular structures was just proximal to the curvature of the urethra and, based on location, could represent either the prostate or the bulbourethral glands. The prostate of the ram is more diffuse, whereas the bulbourethral gland is more discrete and rounded. In the TP- and DHT-treated lambs, the glandular structures were slender, straddling the urethra. The prostate of the ram has serous and secretory cells, whereas the bulbourethral gland has predominantly secretory cells. Because the glands in the treated sheep were rudimentary, definitive identification of the gland was not possible.
Normal development of the male accessory genital glands is thought to be driven by testosterone, with the role of DHT limited to the development of the external genitalia. 10 The results of this study suggest that in utero exposure to testosterone is not solely responsible for the development of the accessory genital glands. The ovaries of DHT-treated lambs may have produced higher levels of testosterone, which would have promoted the development of the accessory genital glands. 4 It is also possible that nonovarian sources of testosterone, such as the adrenal gland, influenced development of accessory genital glands in DHT-treated female sheep. 7 Upregulation of androgen receptors may also have participated in the formation of accessory genital glands in the DHT-treated sheep. 4 Increased testosterone concentrations or expression of androgen receptors may be related to altered concentrations of other hormones associated with the reproductive-growth axis, including gonadotropin-releasing hormone, luteinizing hormone, follicle stimulating hormone, inhibin, and activin. 4 Another possibility is that hormones other than testosterone participate in the formation of accessory genital glands and that the continued release of testosterone is required for maturation and growth of these glands rather than their formation.
Based on the results of this study, in utero exposure to TP or DHT results in malformations in the tubular and external genitalia of female lambs. These findings underscore the importance of monitoring exposure to environmental steroids, many of which are known to bind androgen receptors, to ascertain the effects on human and animal reproduction.
Footnotes
Acknowledgements
The project was funded by the Biotechnology and Biological Sciences Research Council. We thank Richard Irvine for invaluable assistance in the postmortem room and photography.
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The authors received no financial support for the research, authorship, and/or publication of this article.