
Abstract
In both man and animals, inflammatory changes in the pancreas often occur with disturbances in lipid metabolism, including hypertriglyceridemia and an excess of free fatty acids. Hyperlipoproteinemia type I is a human condition caused by a deficiency of lipoprotein lipase. A similar metabolic disturbance that occurs in mink is of considerable comparative interest, as it is also followed by pancreatitis. Pancreatic lesions in hyperlipoproteinemic mink included overt variably sized nodules with hemorrhage and necrosis. These lesions began as intralobular necrosis of exocrine cells and progressed to total lobular destruction, with eventual involvement of interlobular tissue. Remnants of epithelial cells and lipid-filled macrophages were seen in necrotic areas, along with other types of inflammatory cells scattered in a lipid-rich exudate. Granulation tissue developed rapidly in necrotic areas. Additional observations included ductal proliferation, replacement of epithelial cells with fat, and mural arterial thickening, most conspicuously with vacuolated cells and endothelial proliferation. Extravasation of lipid-rich plasma is thought to be a major intensifier of the inflammatory response.
Inflammatory lesions of the exocrine pancreas are not as common in domestic animals as in humans. Nevertheless, pancreatitis has occasionally been reported in a number of animal species, although information on causal factors is sparse and incompletely understood. 19 The majority of reported animal cases occur in dogs and cats. 2,4,14,19 Obesity has been considered a predisposing factor in dogs, and pancreatitis has been reported in dogs fed a high-fat, low-protein diet. 14,17 Recent observations indicate that pancreatitis is more common in cats than previously suggested, although many cases are subclinical. 5
Most human cases of pancreatitis are associated with alcohol abuse or gallstones. 18,24 Gallstones are infrequent in domestic animals, except in cattle, which rarely show evidence of pancreatitis. The authors are unaware of any articles discussing pancreatitis in mink other than the previous reports involving hyperlipemic mink. 3,21
In both man and animals, inflammatory changes in the pancreas often occur in association with disturbances in lipid metabolism and are frequently linked to hypertriglyceridemia. The association between hypertriglyceridemia and pancreatitis has been recognized for many years. 11 It is thought that an excess of free fatty acids plays an important role, 3,21 but the exact physiological mechanisms are not well explained. 9 Hyperlipoproteinemia type I is a human condition associated with a deficiency of lipoprotein lipase (LPL), which leads to severe hyperlipoproteinemia and pancreatitis. 3 Interestingly, a similar condition has also been observed in mink; this condition depends on a mutation in the LPL gene. 16 The anomaly occurs in both sexes and is almost always followed by pancreatitis. A related condition has been described in cats, but it does not show pancreatic involvement. 13 Therefore, mink appear to be the only known animal model for pancreatitis resulting from hyperlipoproteinemia in man. 3
The early changes in exocrine pancreas cells of mink with hyperlipoproteinemia have been reported and include mitochondrial swelling and vacuolation in kits from 55–59 days old, 21 there is also a very brief report of later changes observed in mink 4–21 months old. 3 This report addresses the exocrine pancreatic lesions at the intermediate phase and the progression from the earlier lesions with necrosis to later granulomatous lesions in 4- to 19-week-old mink.
Materials and Methods
Sixteen standard mink, 7 males and 9 females, were examined. The LPL gene was genetically analyzed, as previously reported. 16 Twelve mink were homozygous for Pro 214 mutation in the LPL gene. Two heterozygotes and 2 normal standard mink of corresponding ages were used as controls. To ensure approximately the same duration of the pathological lesions, the investigation was limited to animals between 14 and 19 weeks of age. The animals were born and raised at the Research Farm of the Norwegian School of Veterinary Science, Heggedal. They were fed a standard commercial wet mink feed for fur-bearing animals, as described in a previous article. 21 Fifteen animals were euthanized at the end of the study, and one of the homozygous mink died suddenly. For euthanasia, animals were anesthetized by intramuscular injections of Ketalar 10 mg/kg (Parke-Davis, Barcelona, Spain), and Domitor vet 2 mg/kg (Farmos, Åbo, Finland). They were then exsanguinated by cardiac puncture and autopsied immediately after death.
Samples of pancreatic tissue, liver, kidney, heart, and lung were fixed in 4% phosphate-buffered formaldehyde, pH 7.4, for at least 48 hours; embedded in paraffin wax; and cut into 5-μm-thick sections. Paraffin sections were stained with hematoxylin and eosin (HE) and elastin van Gieson. Immunolabeling was performed on paraffin sections using the EnVision+ HRP system (Dako, Glostrup, Denmark) for polyclonal antibodies against von Willebrand factor (vWF) (Dako) diluted 1:800 and lysozyme (NeoMarkers, Fremont, California, USA) diluted 1:400, as well as a monoclonal antibody against human cytokeratin (Dako) diluted 1:400. Microwave heat antigen retrieval in 0.01 M citrate buffer, pH 6.0, was performed for vWF and lysozyme immunolabeling, whereas sections for cytokeratin immunolabeling were pretreated with trypsin.
Primary antibodies were incubated for 1 hour at room temperature, and EnVison+ anti-mouse or anti-rabbit secondary antibodies (for monoclonal or polyclonal antibodies, respectively) were incubated at room temperature for 30 minutes. Sections of normal sheep kidney were used as a positive control for vWF and cytokeratin, and sections of normal sheep tonsil were used as a positive control for lysozyme. Since it was difficult to make high-quality frozen sections from unfixed, lipid-rich pancreatic tissue, frozen sections were also prepared from fixed material, and both were stained with Sudan III and Oil red O.
The study was carried out on premises approved by the Norwegian Animal Research Authority, and the procedures were approved under the Norwegian Regulation on Animal Experimentation of 1996.
Gross Pathology
All animals were in relatively good condition. All homozygous animals showed macroscopic pancreatic lesions. In 3 cases, the pancreas adhered to the stomach or other adjacent tissues (Fig. 1). In 4 animals, the pancreas was pale and smaller than normal, whereas diffuse enlargement was observed in one animal. In 7 animals, randomly distributed lesions consisted of either multifocal, scarcely visible white spots, or larger, raised nodules measuring up to 1 cm in diameter. Smaller nodules had a light grey/white surface and were dry, or occasionally moist, and white on cut section. Larger lesions sometimes contained necrotic and hemorrhagic foci, with occasional liquefactive areas. The most constant and striking extrapancreatic changes were marked fatty change in the liver and considerable enlargement of the spleen. On gross examination, blood was light pink, with a creamy consistency. All organs in control animals, including pancreas, appeared macroscopically normal.
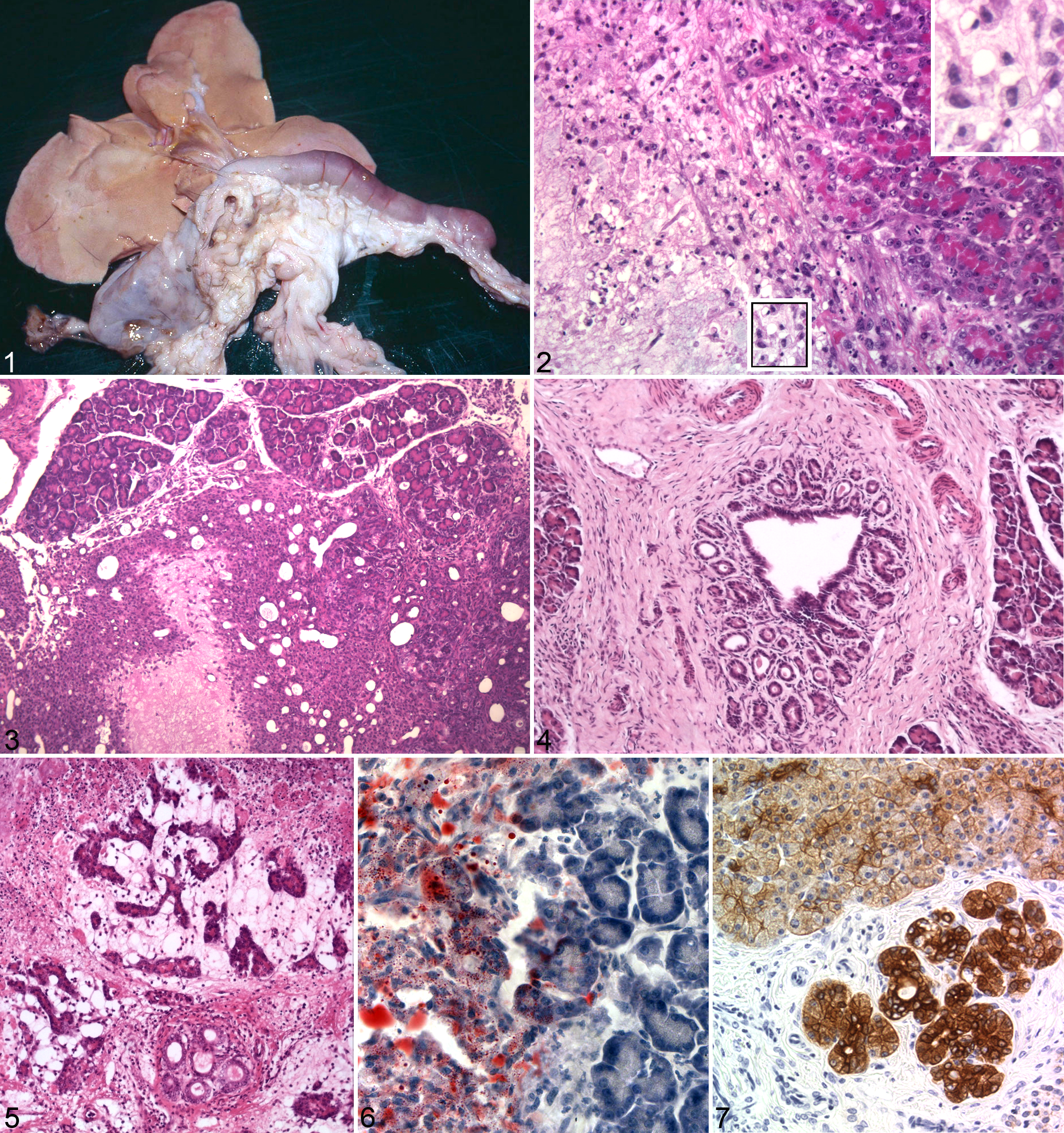
Pancreas from a 14-week-old hyperlipemic mink. There is nodular enlargement of the organ; both branches seem equally affected. The liver is pale and enlarged, with fatty change.
Microscopic Changes
Early pancreatic changes included scattered areas of vacuolated acinar cells. These foci rapidly progressed to necrosis involving both acinar cells and intercalated ducts. Individual lesions frequently coalesced, creating larger necrotic processes that sometimes accounted for the greater part of lobules. Inflammation was observed in affected areas, with infiltration of numerous neutrophils and macrophages, fewer lymphocytes, and rare eosinophils. Neutrophilic or macrophagic dominance was variable between animals, as well as between individual nodules in the same animal. Proliferating vessels and fibroblasts were prominent within lesions. There were numerous lipid macrophages in necrotic areas, either singly or in aggregates (Fig. 2).
Occasionally, a peripheral rim of intact parenchyma was observed (Fig. 3). In later developmental stages, damaged acinar cells were seen in irregular patterns, with shrinkage, loss of zymogen granules, flattening of the cytoplasm, and enlargement of the intercellular spaces. In many areas, irregular cellular architecture was observed, which frequently seemed to represent transitional stages between acinar and intercalated ductular cells (Fig. 3). In most sections, proliferative changes were seen in the interlobular ducts, which were tortuous and contained proteinaceous precipitations (Fig. 4). The proliferative duct changes were present even in areas with unaltered parenchyma.
Collections of tightly packed, intact lipid macrophages occasionally occurred in fibrotic tissue between the acini in early lesions. In later stages, fat replacement and fibrosis were common findings. Areas of acinar regeneration were occasionally observed, sometimes with a tubular pattern (Fig. 5).
Changes in interlobular vessels were present in most histological sections, in both arteries and veins. In arteries, the changes consisted of endothelial proliferation, mural thickening with vacuolated cells, and narrowing of lumina. Obvious tortuosity was also commonly observed. Venous dilations and thrombotic occlusions were occasionally seen. In the liver, there was heavy lipid deposition in hepatocytes. No histological lesions were detected in myocardium, lung, or kidney.
Oil red O and Sudan III-stained, unfixed frozen sections revealed heavy amounts of positive material in necrotic and inflamed areas, which frequently obscured the cellular architecture. Examination of similarly stained, fixed frozen sections showed that the positive material was localized within macrophages, whereas acinar cells were free of fat (Fig. 6).
vWF immunoreactivity was observed in areas with the previously described blood vessel changes and ranged from a continuous lining of endothelium to only remnants of endothelial cells. Lipid macrophages exhibited weak to moderate lysozyme immunoreactivity. Acinar and ductal cells had varying cytokeratin immunoreactivity, and the most intense reactions were observed in proliferating areas (Fig. 7).
Control animals did not show any significant changes apart from moderate proliferation of some interlobular pancreatic ducts.
Discussion
Noninfectious inflammation in parenchymal organs usually follows a restricted morphological pattern, which is more or less specific for each organ. Causal factors may influence the pathomorphological picture to varying degrees. Although ductal proliferation, often with protein plugs, has been observed in pancreatitis in a number of animal species related to several etiological factors, the accumulation of lipid macrophages in the present study was most likely a direct result of the extreme hyperlipemia.
Previous studies have shown that mitochondrial swelling and derangement of christae are the earliest detectable morphological changes in the pancreas of LPL-deficient mink kits. 21 The mechanisms believed to be involved include accumulation of chylomicrons and development of toxic amounts of free fatty acids. In the presence of an extreme accumulation of chylomicrons, the intact particles are internalized in the acinar cells. Previous studies have shown that intact chylomicrons can be taken up in the cells of most organs in LPL-deficient mink. 22 Since acinar cells appear to have a low capacity for mitochondrial fatty acid oxidation, 21 free fatty acids liberated by hydrolysis of the internalized chylomicrons may accumulate in toxic concentrations. It has been proposed that pancreatic lipase can contribute to the liberation of free fatty acids, although it is not known whether chylomicrons can serve as substrates for pancreatic lipase. 21
Free fatty acids can be intercalated in the phospholipid bilayer of intracellular particles. 23 A deranged intracellular membrane structure can be particularly deleterious for mitochondrial function, since even in intact cells, the mitochondrion electron transport chain is the main source of free radical formation. 8,26
Normally, the intracellular defense mechanisms preventing oxidative stress, including glutathione peroxidase, vitamin E, and superoxide dismutase, efficiently protect against the deleterious effects of free radicals. 26 In structurally deranged mitochondria, these defense mechanisms may be insufficient to neutralize free radical production. 7,8,26 Interestingly, antioxidants, including vitamin E, have been used in the treatment of LPL deficiency in humans and to prevent recurrent pancreatitis. 12,20
The pathological changes in the present study appear to show considerable similarities to the human condition, which is not surprising, since the etiology is probably the same. 3 The pancreas may be particularly vulnerable to the deleterious effects of extreme hypertriglyceridemia seen in LPL-deficient animals, since this organ shows high metabolic activity. The pancreas synthesizes more proteins per gram of tissue than any other organ and has a correspondingly high consumption of precursor substrates. 2,10 Enzyme proteins account for 20% of its dry weight. The pancreas is able to produce 3 times the amount of protein as the combined capacity of the liver and reticuloendothelial system. 10 This exacting task relies on optimal metabolic function, especially in animals on a growth-promoting, high-energy diet, and oxidative stress may occur under certain circumstances. Our hypothesis is that increased activity of the mitochondrial electron chain for ATP production, in support of the increased synthesis of enzyme protein, may make the mitochondria more vulnerable to the toxic effects of an excess of free fatty acids and thus increase the production of free radicals in these animals.
The pancreas does not have a stable cellular morphology, but it is instead partially influenced by diet. Transitional stages between acinar and ductular structures are known to occur. 1,2,15 Hyperplasia of acinar cells, with observable enlargement of the organ, occurs in response to diets rich in protein and fat. 1,2 To optimize growth and fur quality, the mink on this farm received a high-energy feed, with a fat content of up to 49 energy %.
Pancreatic tissue possesses considerable potential for regeneration and differentiation after damage. 2 In the absence of further stress, healing usually occurs, either by total restitution or by scarring. The progression from an acute attack of pancreatitis to a chronic pancreatic condition is most likely explained by the persistence of a detrimental factor. In these mink, it was most likely the prolonged presence of lipid-rich plasma within the damaged tissue that caused persistent inflammation. In this study, pathomorphologic progression was easily observed because of the young age of the animals and the rapid development of pathologic changes, which prevented complications in examination caused by recurrent lesions.
In conclusion, the pathomorphological pancreatic lesions observed in hyperlipemic mink show some similarities to pancreatic changes observed in humans with hyperlipidemia as well as pancreatitis caused by other factors. Ductal proliferation, in particular, is a common finding in pancreatitis from a variety of causes. The most striking feature in this study was the accumulation of lipid macrophages, a phenomenon thought to be a direct result of extreme hyperlipemia. The liberation of toxic concentrations of free fatty acids seems to play a key role in the pathogenesis of pancreatitis associated with hypertriglyceridemia. 6,25
Footnotes
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The authors received no financial support for the research, authorship, and/or publication of this article.