
Abstract
Studies focusing on the molecular basis of canine mammary tumors (CMT) have long been hampered by limited numbers of molecular tools specific to the canine species. The lack of molecular information for CMT has impeded the identification of clinically relevant tumor markers beyond histopathology and the introduction of new therapeutic concepts. Additionally, the potential use for the dog as a model for human breast cancer is debatable until questions are answered regarding cellular origin, mechanisms, and cellular pathways. During the past years, increasing numbers of canine molecular tools have been developed on the genomic, RNA, and protein levels, and an increasing number of studies have shed light on specific aspects of canine carcinogenesis, particularly of the mammary gland. This review summarizes current knowledge on the molecular carcinogenesis of CMT, including the role of specific oncogenes, tumor suppressors, regulators of apoptosis and DNA repair, proliferation indices, adhesion molecules, circulating tumor cells, and mediators of angiogenesis in CMT progression and clinical behavior. Whereas the data available are far from complete, knowledge of molecular pathways has a significant potential to complement and refine the current diagnostic and therapeutic approach to this tumor type. Furthermore, current data show that significant similarities and differences exist between canine and human mammary tumors at the molecular level. Clearly, this is only the beginning of an understanding of the molecular mechanisms of CMT and their application in clinical patient management.
Research on the origin and cellular and molecular mechanisms of neoplastic diseases has identified a large number of individual molecules and signaling pathways, particularly in humans and laboratory mice. Despite the widely accepted concept of cancer resulting from defective genes—primarily, oncogenes and tumor suppressor genes—new principles, pathways, and molecules are still being discovered as essential factors for the neoplastic transformation of cells and the malignant progression and lethal outcome of metastatic disease. In contrast to humans, little is known of the mechanisms and molecules that contribute to canine tumors. This deficiency is further augmented by limited or unknown specificity of the molecular tools used in studies on canine tumors, by small group sizes, and by the lack of patient follow-up information required for the assessment of the clinical significance of individual molecular data.
With a prevalence of 0.2%, canine mammary tumors (CMT) are the most common neoplasms in the female dog. Histologically, approximately 50% of these neoplasms are diagnosed as malignant, and distant metastases are common causes of death in these patients. 2,68,85 Because of the concept that CMT may serve as a relevant model for human breast cancer, 134 similarities and differences in their molecular alterations, when compared to human disease, have become of increasing interest. This review article is intended to summarize the current knowledge on molecules, pathways, and molecular diagnostic tools that may add to the understanding, diagnosis, prognostic assessment, and clinical management of CMT. Specific functional groups of molecules will be addressed separately, including mediators of apoptosis and DNA repair, oncogenes and tumor suppressors, adhesion molecules, mediators of angiogenesis, and markers for circulating tumor cells that are involved in different steps of tumor progression and metastasis. In addition, the prognostic value of common markers of cellular proliferation and angiogenesis are reviewed.
Tumor Growth: Proto-oncogenes and Oncogenes
Cell proliferation in nonneoplastic epithelium is strictly regulated by several molecular pathways, including regulators of cell cycle checkpoints. 125 Neoplastic mammary tissue is characterized by increased cell proliferation, when compared to nonneoplastic mammary epithelium. CMT cells must have increased expression levels or hyperactive growth-promoting gene products (proto-oncogenes) or decreased expression levels or functional activity of growth-inhibiting gene products (tumor suppressors) involved in growth promotion. However, the initiating impulses of enhanced cell division in CMT are unclear.
Growth-promoting proto-oncogenes are either directly involved in cell cycle progression, like members of the cell cycle checkpoints (cyclins, cyclin-dependent kinases [CDK], retinoblastoma protein), or they are members of molecular networks that indirectly induce cell cycle progression, like growth factor receptor pathways. Of the numerous potential proto-oncogenes directly involved in cell cycle progression, only expression levels of cyclin A and cyclin D1 have been analyzed in detail for CMT. Both proteins have increased expression levels in malignant tumors. 93 However, the prognostic significance of this protein expression is unclear because precancerous mammary dysplasia also expresses increased cyclin D1. 129 SATB1, a global chromatin organizer and transcription factor, has emerged as a possible key factor that promotes the growth and metastasis of human breast cancer cells, and its overexpression is associated with a poor prognosis in humans. On the transcriptional level, the opposite is true for CMT. Adenomas and metastatic CMT had decreased levels of SATB1 when compared to the nonneoplastic glands of the same dog. 58 SATB1 is an example of one of the differences in the molecular mechanism of CMT when compared to human breast cancer, although they are similar in many other aspects.
Growth factor signaling may cause both growth-stimulating and growth-inhibiting effects on CMT. The consequences of growth factor signaling are highly dependent on the pathway, the affected cell type, and the activity status of pathway modulators. Protein and mRNA expression levels of growth factors and their receptors have been extensively studied in CMT. Because of its relevance in human breast cancer, research has focused on the expression of epidermal growth factor 2 (ERBB2) as a growth-promoting signaling pathway in CMT. ERBB2 is overexpressed in 25% of human breast cancers, and this overexpression is associated with a poor prognosis. 25 Importantly, it identifies those patients who will benefit from therapy with trastuzumab (Herceptin), a monoclonal antibody that binds and blocks the HER-2 receptor.
A number of studies have attempted to identify a similar prognostic relevance for ERBB2 expression in CMT. So far, loss of ERBB2 expression has been associated with a poor prognosis when linked with estrogen receptor negativity and expression of one of 3 basal cell markers (P-cadherin, p63, cytokeratin 5). 27 A tendency toward prolonged survival of dogs with ERBB2-positive tumors was observed in another study, although the association between ERBB2 overexpression and prolonged survival time was not statistically signifcant. 47 Expression of epidermal growth factor 1 (EGFR1) was not associated with any clinicopathologic parameters in dogs with CMT. 28 Despite an unaltered expression level, EGFR may also cause increased growth stimulation through the loss of the ERBB receptor feedback inhibitor 1 (MIG6). MIG6 is downregulated in various human cancers and physiologically attenuates the mitogenic epidermal growth factor signaling by inhibition of EGFR autophosphorylation and reduction of mitogen-activated protein kinase activity. A similar downregulation was observed in canine metastatic mammary carcinomas but not in adenomas. 58 The intensity of EGFR signaling is therefore not unequivocally reflected in its protein expression levels but may be modulated by regulatory proteins in its downstream signaling cascade.
In addition to EGFR, metastatic canine mammary carcinomas have decreased expression of several transmembrane growth factor receptors when compared to nonmetastatic carcinomas or normal glands. These include members of the transforming growth factor β receptors (TGFBR), the fibroblast growth factor receptor (FGFR), and the growth hormone receptor (GHR). 58-61,136 In contrast, expression of other growth factor receptors, such as the insulin-like growth factor 1 and 2 receptors (IGF1R, IGF2R) and the insulin receptor (INSR), are unaltered in metastatic carcinomas (Figs. 1, 2 ). 56 There has been no associating link identified between the expression patterns of these growth factor receptors and clinical prognosis at this point.
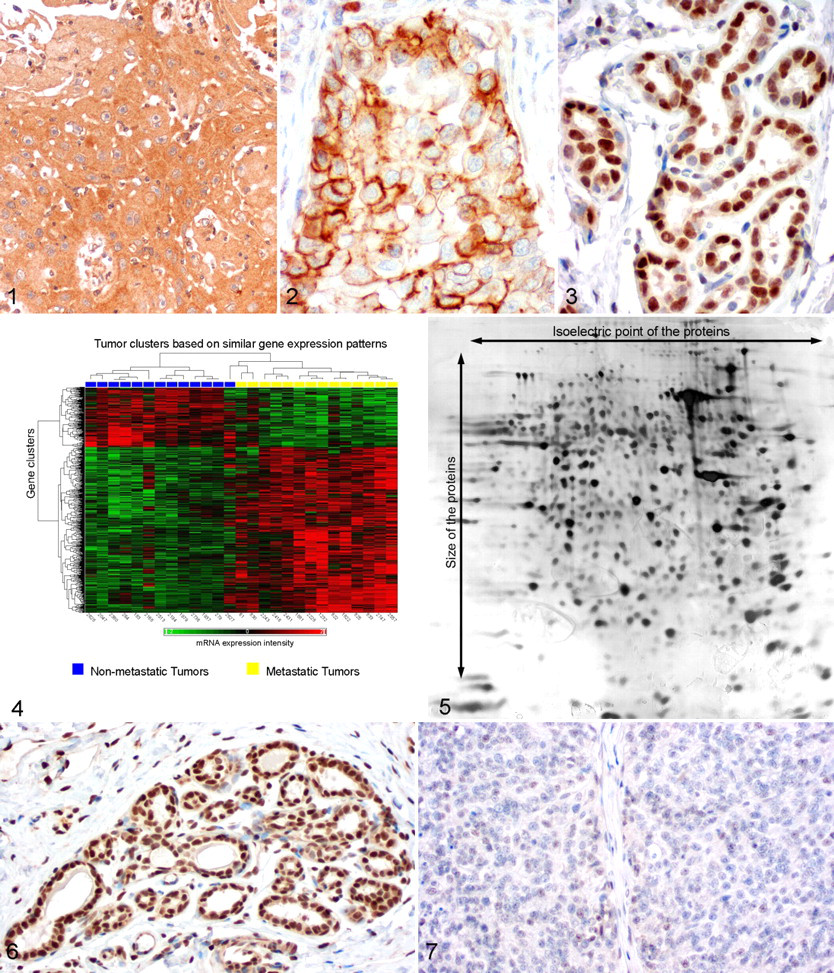
Insulin-like growth factors 1 and 2 (IGF1 and IGF2) have a major impact on the carcinogenesis of human breast cancer, 139 and a similar impact is expected for CMT. 89 This belief is based on the fact that progestins induce secretion of growth hormone (GH) in the mammary gland. 88 GH may in turn have a direct local growth stimulatory effect on the mammary epithelium but also induce synthesis of IGF1. 89 Lysates of malignant mammary tumors contain increased levels of IGF1 when compared to benign tumors. 110 However, its cellular source in CMT mammary tumors is currently unclear because IGF1 and IGF2 mRNA expression levels are decreased in malignant tumors and protein expression levels have not yet been analyzed immunohistochemically. 56
Estrogen receptor activity is another major stimulator of mammary epithelial cell proliferation, and its expression is physiologically closely associated with progesterone receptor expression. 87 Presumably, either stimulation of estrogen receptors or differences in expression levels may contribute to the growth and clinical behavior of CMT. The general assumption of steroid receptor influence on mammary carcinogenesis is based on the observation that dogs spayed at a young age have a smaller prevalence of mammary tumors. 127,130 Several studies attempted to identify prognostic estrogen receptor protein expression levels at different grades of malignancy. Although there are conflicts in the literature, most of the studies found a decrease in estrogen receptor expression with increasing grades of malignancy when compared to normal glands (Fig. 3). 9,15,80 Therefore, CMT cells seem to become increasingly resistant to the stimulatory proliferative effects as malignancy progresses. However, there is controversy as to the prognostic value of steroid receptors. Although most authors found that steroid receptors were not independent prognostic factors for overall survival, 15,75,80 others found that decreased estrogen receptor, but increased progesterone receptor, expression was associated with poor clinical behavior in canine carcinomas. 9
Microarray studies on CMT revealed a vast body of information focused on altered mRNA expression of numerous growth-promoting genes (Fig. 4). 59,60,112,133 The impact of these gene products on mammary tumor development needs further confirmation by other methods, including protein levels. A proteome analysis that compared metastatic and nonmetastatic simple mammary carcinomas identified altered expression patterns of several growth-promoting and proliferation-associated genes (Fig. 5). 60 These included the Ran/TC4-binding protein (RANBP1) involved in spindle cell assembly, the elongation factor 1 δ (EEF1D) protein that is possibly involved in neoplastic transformation and carcinogenesis, and the proliferating cell nuclear antigen (PCNA) that is a prerequisite for DNA replication. 60
The available reports indicate that our knowledge of the impact of proto-oncogenes on the pathogenesis of CMT is limited.
Loss of Growth Inhibition: Tumor Suppressors
Dys-regulated cellular proliferation in nonneoplastic cells is strictly regulated by several molecular pathways, including cell cycle checkpoints and different tumor suppressors. Malignant CMT have an increased fraction of cells with active cell cycle progression, as shown by Ki67 and PCNA immunohistochemistry. An increased number of actively cycling cells appears to be associated with the clinical prognosis. 125
One of best-characterized tumor suppressors and growth inhibitors is p53. This protein is activated by several triggers, including oxidative stress and DNA damage, and it leads to irreversible cell cycle arrest (senescence). 138 Furthermore, p53-induced cell cycle arrest is mediated by a transcriptional increase of p21, an inhibitor of cyclin E/Cdk2 at the G1 phase–M phase transition checkpoint of the cell cycle. The impact of p53 protein expression levels on the functional activity of CMT has been intensely studied, but its prognostic relevance is still questionable.
The p21 protein is overexpressed in canine metastatic carcinomas when compared to adenomas and lymph node metastases. 54 Increased expression of p21 in metastatic mammary tumors may be a p53-induced attempt to inhibit cell cycle progression that fails in the majority of tumor cells.
The cyclin-dependent kinase inhibitor p27 is regulated by extracellular stimuli, such as transforming growth factor β but not p53. Similar to p21, p27 regulates cell cycle progression by interacting with cyclin E/Cdk2 or cyclin D/Cdk4. In addition, p27 expression is reduced in metastatic carcinomas and their metastases, as well as in adenomas, thereby indicating that loss of p27 expression occurs early in the malignant transformation of mammary epithelium (Figs. 6, 7 ). 54,62 According to observations of a CMT cell line, mutations in the p16 gene, a cell cycle inhibitor at the G1 phase–S phase transition, may be associated with carcinogenesis. 19 To date, the presence and clinical effect of p16 mutations have not been confirmed using tissue samples of CMT.
The phosphatase and tensin homolog (PTEN) appears to be another interesting tumor suppressor with prognostic relevance for CMT. PTEN reduces cell proliferation but is involved in apoptosis induction and cell adhesion. In CMT, loss of PTEN expression was associated with a significant shorter overall survival in 76% of malignant tumors. 116 A similar association between loss of PTEN expression and survival rates is true for human breast cancer. 21 Interestingly, loss of expression failed to be of prognostic relevance in feline mammary tumors in the same study. 114 PTEN seems to be a promising candidate to complement histology-based prognostic evaluation of CMT.
Global mRNA expression analysis of CMT has revealed downregulation of several growth-inhibiting molecules. The most significantly affected pathways included negative modulators of the WNT receptor signaling pathway and the mitogen-activated protein kinase (MAPK) pathway. 59,60,112,133 Correlation between mRNA expression and protein expression levels is sometimes poor, and further studies on protein expression are needed to confirm the relevance of these mRNA patterns. Proteome analyses identified several differentially expressed growth-inhibiting proteins when metastatic simple carcinomas were compared with nonmetastatic carcinomas. 60 These included downregulation of tumor suppressors in metastatic tumors, such as Rho GTPase activating protein, which is involved in proliferation and metastasis inhibition.
In conclusion, several tumor suppressors have been identified that are likely involved in the carcinogenesis and progression of CMT. However, a prognostic relevance has been shown only for PTEN. Information is lacking on the mutational status and functional activity of other candidates.
Cell Proliferation and Prognosis: PCNA, Ki67, and AgNOR
Although the mechanisms of uncontrolled cellular proliferation in CMT are not fully understood, increased cell proliferation is a clear feature of malignant CMT. Three histologic methods to estimate proliferation activity have been successfully and repeatedly applied in several studies on CMT: PCNA, Ki67, and AgNOR.
PCNA and Ki67
PCNA is located in the nucleus, and it acts as a cofactor of DNA polymerase δ, thereby increasing DNA replication. 90 In addition, PCNA plays a role in RAD6-dependent DNA repair and inhibition of apoptosis via negative regulation of the tyrosine kinase c-abl stability. 44 PCNA concentration peaks during the G1 and S phases of the cell cycle, but because of its long half-life time of 8 to 20 hours, it is also found in cells that have recently concluded the M phase. 90 Due to its involvement in DNA repair, PCNA can be detected in noncycling cells.
Ki67, a heterodimer protein with a peak expression in the M phase of the cell cycle, is commonly visualized with the MIB1 antibody. 34 A half-life of less then 1 hour allows rare detection of Ki67 in noncycling cells, malignant cells with an abnormal or prolonged expression during G0, and cells with an increased time to traverse G1 or G2. 34,142 The results of both staining methods are reported as PCNA or Ki67 labeling indices, which describe the ratio of the number of positive cells to the number of positive and negative cells.
Due to the expression differences in their target proteins, both labeling indices are often applied synchronously and as mutual controls. However, there is controversy regarding the correlation between PCNA and Ki67 indices in CMT. One study found a significant correlation with malignancy, 71 whereas a second study found a correlation of both indices in benign tumors and dysplasias but not malignant tumors. 102 The correlation of Ki67 and PCNA with other proliferation indexes varies among studies and by mammary tumor type. The PCNA index showed a good correlation with mitotic index, 124 whereas PCNA and Ki67 labeling indexes did not correlate with the S phase fraction in mammary tumors. 143 In addition, the PCNA and Ki67 indices are inversely correlated with estrogen receptor α expression. 33
The prognostic relevance of PCNA indices were evaluated in 3 studies on CMT, 71,102,108 which found a reduced PCNA index in benign tumors when compared to carcinomas (Table 1) and in well-differentiated carcinomas when compared to less-differentiated solid and anaplastic carcinomas. 71,102,108 There was also an association between the PCNA index and the nuclear grade. More important, univariate and multivariate analysis found an association between increased PCNA indices and postoperative survival. 71
Indexes of Canine Mammary Tumors to Assess Prognostic Relevance a
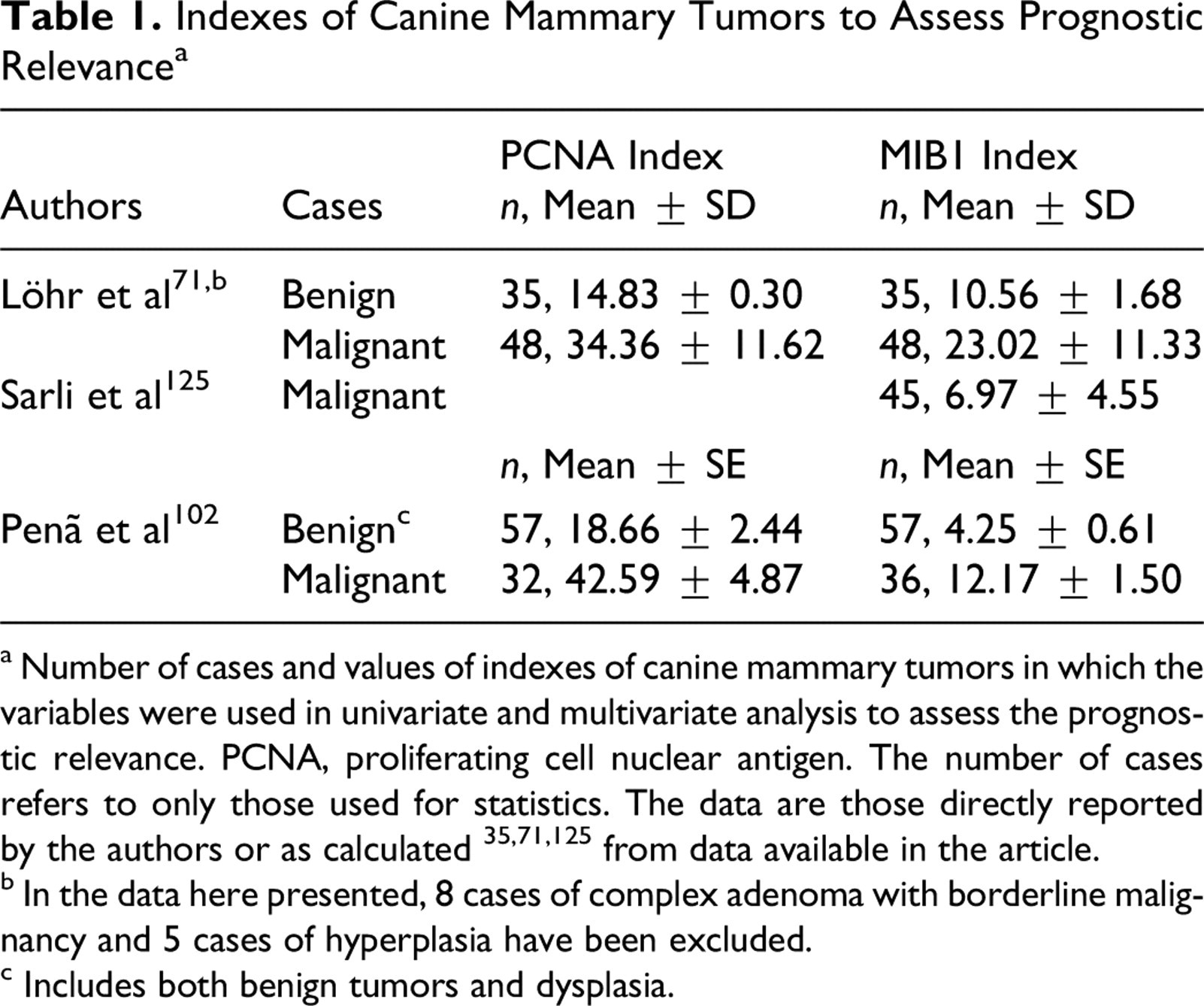
a Number of cases and values of indexes of canine mammary tumors in which the variables were used in univariate and multivariate analysis to assess the prognostic relevance. PCNA, proliferating cell nuclear antigen. The number of cases refers to only those used for statistics. The data are those directly reported by the authors or as calculated 35,71,125 from data available in the article.
b In the data here presented, 8 cases of complex adenoma with borderline malignancy and 5 cases of hyperplasia have been excluded.
c Includes both benign tumors and dysplasia.
As with the PCNA index, the Ki67 index was significantly different between benign tumors/dysplasia and malignant tumors, with higher indices found in solid carcinomas when compared to tubular carcinomas (Table 1). 71,102,125 In a series of 60 mammary intraepithelial lesions, increasing Ki67 indices were observed with increasing tumor grade, with the highest indices in high-grade carcinoma in situ. High Ki67 index values were also associated with several other clinical and histopathologic variables, including size 24 and lymph node involvement and metastasis. 102,125 Consequently, survival time was significantly decreased for dogs baring high Ki67 index tumors. 125 However, PCNA and Ki67 indexes did not correlate with breed, size of the dog, some reproductive variables, rate of growth, ulceration, or adhesion to underlying tissues. 102
Despite their proven prognostic value, PCNA and Ki67 are still not routinely used in CMT diagnosis. The most important problems in assessing proliferative activity using PCNA and MIB1 involve the difficulty in standardizing both the staining procedure and the laborious quantification procedure. For example, PCNA expression varies within the tumor and shows variability in expression intensity, so both strongly and weakly stained nuclei are identified. 71,102,108,125
AgNOR
Nuclear organizer regions (NOR) are chromosomal segments involved in the biogenesis of ribosomes. They are composed of argyrophilic proteins—hence, AgNOR—that colocalize with ribosomal DNA loops of the nucleolus. The rearrangement of these loops is directly related to the proliferative activity of the cell. 71 Decondensation of the ribosomal DNA loops during higher transcription activity leads to segregation of the associated proteins. The subsequent increase in the number and area of silver-stainable intranuclear and intranucleolar AgNOR allows the interphase of cell division to be visualized in paraffin sections and routine cytologic preparations. 71 Therefore, AgNOR give an indication of the cell duplication rate but have a more complicated and elaborate quantification process than other proliferation indices, such as mitoses, PCNA, and Ki67.
In contrast to the regard of PCNA and Ki67 staining, an international committee for AgNOR quantification proposed guidelines for the standardization of the stain shortly after the introduction of this technology. Most investigations on CMT have followed these recommendations and are therefore comparable. 8,71,125 AgNOR can be quantified in several ways, but the most common methods include the number of AgNOR per cell, the AgNOR area per cell, and the ratio between the AgNOR area and the nucleus area using lymphocytes or endothelial cells as an internal standard. In canine mammary samples, a comparison of sensitivity and specificity between the AgNOR counts and the AgNOR area indicated that AgNOR counts are more precise than estimation of the area. For instance, AgNOR counts were able to correctly classify 89% of given cases into a malignant or nonmalignant category using 2.10 AgNOR counts per cell as a cutoff point (sensitivity, 0.90; specificity, 0.81; accuracy, 0.89). 8 Both AgNOR number per cell and AgNOR area per cell were significantly higher in malignant versus benign tumors 21,71 or hyperplastic mammary gland epithelia. 8 However, AgNOR count was unable to discriminate between nonmetastatic and metastatic mammary carcinomas 5 and was not associated with invasive features of mammary carcinoma. 125
For estimating clinical behavior, univariate analyses found that higher AgNOR areas or numbers per cell were associated with a cancer-free interval and survival. 125 However, in multivariate analyses, AgNOR was predictive for 1-year survival, 73 but only the histopathologic diagnosis and tumor invasiveness were predictive for 2-year survival. 71,125
Apoptosis: Loss of the Emergency Switch
In nonneoplastic cells, any genetic and metabolic imbalance that may lead to malignant transformation is prevented by several, mostly redundant, molecular mechanisms that eventually lead to apoptosis, or programmed cell death. Under physiologic conditions, severe DNA damage, incorrect DNA replication, dysregulation of cell cycle progression, hypoxic stress, or accumulation of misfolded proteins are typical events that trigger the activation of proapoptotic pathways or the suppression of antiapoptotic pathways and molecules. 6,144
Malignant CMT cells experience all the triggers for apoptosis but somehow avoid the induction of apoptosis. It has been hypothesized that defects in cellular surveillance systems (eg, the DNA damage response, overexpression, or permanent activation of apoptosis inhibitors) or functional defects of proapoptotic molecules are responsible for this phenotype. Several studies have addressed the hypothesis of dysregulated apoptotic pathways in CMT and have analyzed the expression patterns or functions of genes associated with apoptosis.
One of the core regulators in the early phase of apoptosis induction is the tumor suppressor p53. In stressed cells and, notably, in cases of severe DNA damage, activated p53 accumulates in the cell and leads to cell cycle arrest, which either activates the DNA damage response or induces apoptosis if DNA damage is irreparable via activation of the proapoptotic protein bax. 18,31 Activation of bax ultimately leads to the activation of apoptosis-related initiator cysteine peptidase (caspase), including caspases 8 and 9, and the effector caspase 3 that degrades a variety of cellular proteins.
The expression pattern and mutational status of p53 has been studied intensely in CMT. Several p53 mutations have been identified, but the functional relevance of these mutations is unclear, and none of the mutations are unequivocally associated with malignant clinical behavior in CMT. 10,70,94 Furthermore, p53 mRNA and protein expression data for malignant tumors vary in reports, between normal to increased expression, when compared to benign tumors or the normal gland. 54,67,122 Only one study found an association between increased p53 protein expression and decreased survival time. 70
Proteins involved in the downstream signaling of p53 and other triggers of apoptosis have been intensely analyzed. In these studies, CMT had increased or unaltered expression of antiapoptotic proteins (BCL2, BCLX, SFRP2), whereas proapoptotic proteins (BAX, caspase 8, caspase 9) had decreased expression. 67,141 Again, the lack of information on the functionality of the overexpressed proteins and the clinical and prognostic relevance of these expression patterns limit their use as prognostic factors.
The rapid growth of malignant tumors causes cytotoxic stress, including hypoxia and nutrient deprivation. Consequently, tumor cells accumulate increased amounts of misfolded proteins in the endoplasmic reticulum, which ultimately reduce cell survival by inducing apoptosis. 65 In response to this stress, both nonneoplastic cells and tumor cells express various proteins of the protective unfolded protein response. 66 These proteins include heat shock proteins (HSP) and Derlin-1 (Derl1), a newly discovered member of the unfolded protein response. 66 It is therefore not surprising that metastatic CMT and their metastases have increased expression of HSP70, HSP90, and Derl1 (Fig. 8). 53,64 Targeted inhibition of these stress proteins may be an interesting therapeutic strategy to decrease or prevent metastatic spread.
Microarray studies on complex malignancy-associated gene expression patterns in CMT cells revealed a downregulation or upregulation of several apoptosis inhibitors and apoptosis inducers when compared to normal and hyperplastic mammary gland or benign tumors. 59,60,112,133 However, metastatic carcinomas clearly had increased proapoptotic and antiapoptotic gene transcription when compared to nonmetastatic carcinomas. 60 These findings were supported by comparative proteome analyses of metastatic versus nonmetastatic CMT that detected downregulation of proapoptotic proteins, including tropomyosin 1 and peroxiredoxin 6, whereas the antiapoptotic scavenger protein, thioredoxin domain-containing 5, was upregulated in metastatic tumors. 60
In summary, it is hypothesized that malignant tumor cells have a new equilibrium of a broad spectrum of proapoptotic and antiapoptotic proteins. Therefore, diagnostically and prognostically relevant information may be obtained only by evaluating the entire complex group of members involved in the intricate apoptotic network rather than by focusing on a single protein.
DNA Repair: Failure of the DNA Quality Management
Genomic instability is a fundamental feature of cancer cells, 49 but the causes and mechanism of this instability appear extremely variable. Carcinogens may initially induce DNA damage, 46 whereas later stages of cancer progression show chromosomal instability when telomeres in the tumor become critically short and prone to chromosomal fusions. 77 Aberrant cell proliferation caused by mutationally activated proto-oncogenes or inactivation of tumor suppressors continuously induces DNA replication stress. 41 Furthermore, chronic hypoxia and/or cycles of hypoxia and reoxygenation contribute to genomic instability. 6 This genomic DNA damage is not restricted to tumor cells but is a rather common event in any nonneoplastic cell in the body at any time. 76 In nonneoplastic cells, the integrity of the genome and, hence, the survival of the cell in its environment are ensured by an intriguing network of DNA damage sensors and DNA repair enzymes. 49 Due to the wide variety of DNA damage forms, the mammalian cell possesses several DNA repair tools, including nonhomologous end joining, homologous recombination, base excision repair, nucleotide excision repair, mismatch repair, and translesion synthesis. 46 Mutational or epigenetic inactivation of the DNA repair genes accumulates during tumor development and causes alterations in DNA repair mechanisms, which accelerates malignant progression by increasing the mutation rate. 49
The hypothesis that defects in the DNA repair system are involved in the carcinogenesis of CMT was initially based on observations in human breast cancer. Between 5% and 10% of human breast cancer cases are considered hereditary breast cancers associated with germline mutations in the breast cancer susceptibility genes 1 and 2 (BRCA1 and BRCA2). 79 Both proteins belong to the group of tumor suppressor genes and play critical roles in the preservation of genomic stability by acting as DNA damage sensors and inducers of the DNA repair response. 137
Due to the relevance of BRCA1 and BRCA2 mutations in the prognosis of human breast cancer, numerous studies tried to identify similar mutations in CMT. In contrast to studies in humans, none of the canine studies revealed any association between genomic polymorphisms of BRCA1 and BRCA2 and the development or presence of CMT. 55,119,131
However, several single-nucleotide polymorphisms in the genomic sequence of BRCA1 and BRCA2, as well as a slightly different BRCA1 mRNA splicing pattern, were identified in CMT when compared to the normal gland. The association of these genomic variations with an increased incidence of CMT has not yet been confirmed in clinical studies. 48,119 Immunohistochemically, malignant CMT have reduced nuclear, and increased cytoplasmic, expression of BRCA1 protein, although changes in location and expression are not significantly associated with clinicopathologic variables like disease-free interval and overall survival. 98 A similar malignancy-associated shift from nuclear to cytoplasmic protein expression can be observed for DNA topoisomerase 2-binding protein 1 (TOPBP1) in human breast cancer. 35 TOPBP1 colocalizes with BRCA1 during the S phase of the cell cycle, and it is involved in initiating DNA repair similar to BRCA1 in feline mammary tumors. 91 The clinical and prognostic relevance of TOPBP1 is so far unproven.
BRCA-mediated DNA repair is accomplished through the interaction and activation of BRCA1 and BRCA2 with DNA repair proteins. 128 Several genes are involved in BRCA downstream signaling, but only RAD51 has been analyzed in greater detail for CMT. Increased RAD51 and BRCA2 expression levels were found in metastatic CMT and their lymph node metastases when compared to normal glands or adenomas, which indicates that malignant CMT may have genetic instability due to an unknown cause (Fig. 9). 55,60,63 Another major protein involved in the identification and repair of DNA damage is the tumor suppressor p53, which is activated after the identification of DNA damage and which induces cell cycle arrest (see growth inhibiting molecules) to allow either DNA repair or apoptosis, if DNA damage is too severe (see section on apoptosis).
Activated DNA repair systems in CMT have been observed in several microarray studies focused on the CMT transcriptome. These studies identified mRNA expression patterns of activated DNA repair systems, including increased expression of genes such as GADD45, RAD17, RAD21, RPA1, SMC1A, and TOPBP1 in metastatic simple carcinomas when compared to nonmetastatic carcinomas and normal mammary glands of the same dogs. 59,60,112,133 However, comparing the proteome of metastatic versus nonmetastatic carcinomas failed to identify any differences in protein expression levels associated with DNA repair. 60
In conclusion, CMT—especially, metastatic CMT—are characterized by a gene expression pattern of activated DNA repair genes that show genetic instability. Causes for genetic instability are unknown, but it is likely that DNA replication stress due to aberrant tumor cell division, hypoxia, and accumulation of mutations and epigenetic modifications to DNA repair genes contributes to this phenomenon. It may be that, in the near future, complete sequencing of the CMT genome from affected dogs may identify mutations in DNA repair genes in some CMT that act as initiators of carcinogenesis, similar to that in human breast cancer.
Cell Adhesion Molecules: Departure and Arrival
The development of multicellular organisms requires a dynamic and well-regulated intercellular adhesion system. During various physiologic and pathologic processes, such as tissue development, cell growth, differentiation, embryogenesis, immune response, and tumor development and progression, cellular junctions are reorganized and regulated by a network of protein complexes. During the carcinogenic process, this complex network of cell–cell contacts is modified and results in a barrier breakdown that promotes cancer progression and metastasis. 74 In fact, changes in the expression or function of adhesion molecules in tight junctions, adherens junctions, and gap junctions (Fig. 10) have been implicated at all steps of tumor progression, including detachment of tumor cells from the primary site, invasion of vessels, extravasation into distant target organs, and formation of metastases. 74
Cadherins and Catenins
Cadherins are a superfamily of calcium-binding transmembrane glycoproteins with an extracellular domain responsible for cell–cell interactions, a transmembrane domain, and a cytoplasmic domain that is frequently linked to the cytoskeleton. 140
Most studies implicating cadherins in tumorigenesis have focused on E-cadherin because it is the major cadherin expressed by epithelial cells. 7,16,30,78,113 It is localized at the basolateral membrane in adherens junctions, defining apicobasal polarity (Fig. 11). 103 The mature E-cadherin molecule contains an ectodomain that is essential for calcium-dependent cell–cell adhesion. 4 The clustering into junctional structures is facilitated and mediated by interaction between E-cadherin cytoplasmic domain and β-catenin and plakoglobin. Both interact with α-catenin, which links cadherins to the cytoskeleton. 42
In the normal mammary gland, the polarized epithelial cells lining the ducts and alveoli express E-cadherin, which holds them together. In contrast, myoepithelial cells express P-cadherin but not E-cadherin. 29,101 Several mechanisms have been described that associate E-cadherin downregulation or inactivation with tumor development and progression. 4 These mechanisms include mutation of the human E-cadherin gene CDH1, epigenetic silencing, transcriptional silencing, microRNA, and posttranslational glycosylation (N-linked) that modulates and regulates E-cadherin functionality. 3,40,72
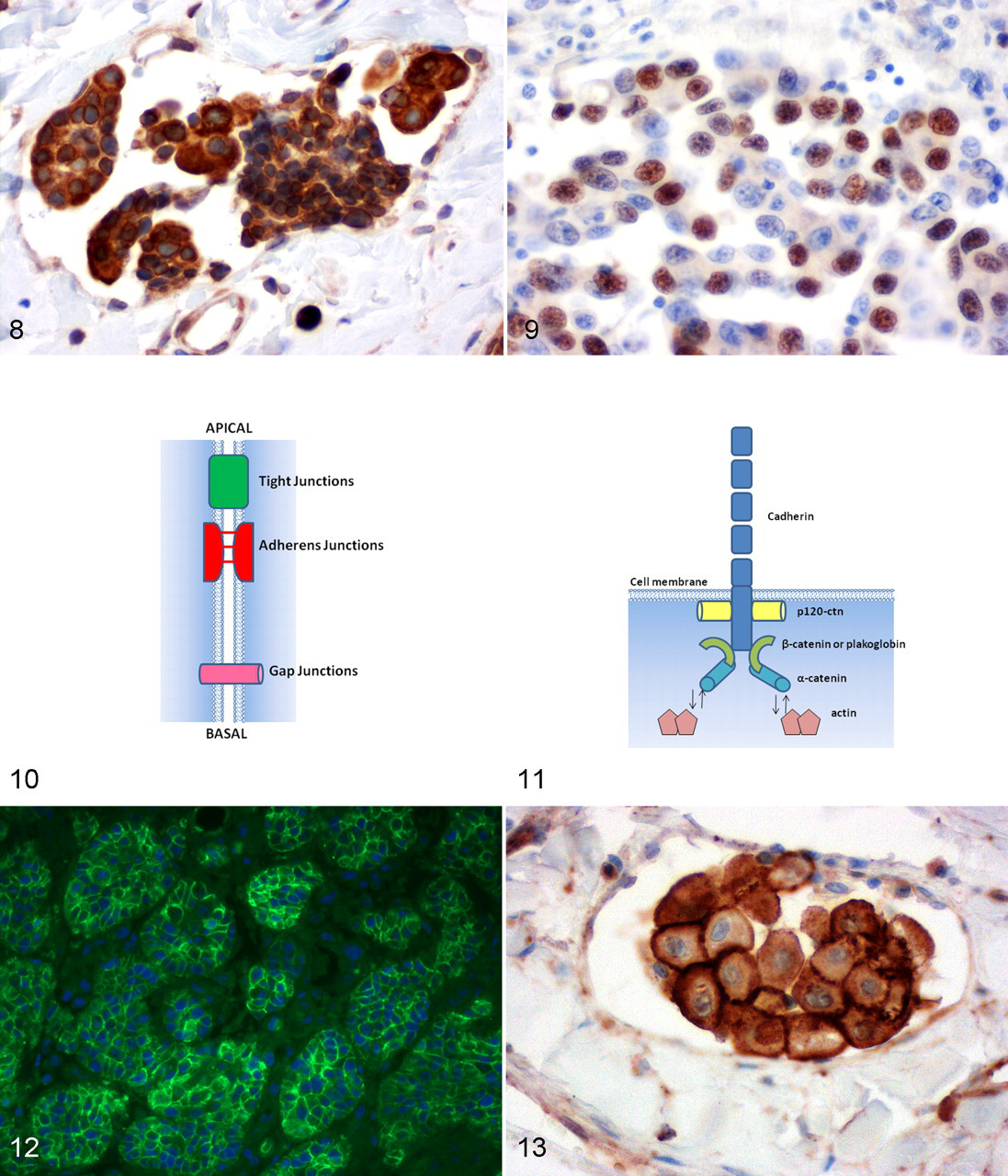
Poorly differentiated, invasive, and metastatic canine mammary carcinomas show a loss of E-cadherin expression in some tumor cell subpopulations when compared to well-differentiated carcinomas, suggesting that altered E-cadherin expression may be an important process in malignant transformation (Fig. 12). 7,16,30,78,113 This hypothesis is supported by the observation that the loss of E-cadherin expression correlates with a shorter overall survival and disease-free period. 30,78
The molecular mechanisms behind E-cadherin downregulation in CMT have not yet been demonstrated in vitro or in vivo. However, a recent study indicated that posttranslational modification of E-cadherin by N-glycosylation could represent an alternative regulatory mechanism of E-cadherin in CMT. 104 Using mass spectrometry, a deleterious β1,6 GlcNAc–branched N-glycan structure was identified in the extracellular domain of canine E-cadherin. 105
P-cadherin is the cadherin expressed by myoepithelial cells in mammary tissue. The mature P-cadherin is a glycoprotein with a structure similar to that of the classic aforementioned cadherins. 101 In normal adult nonlactating mammary tissue, spatially selective expression of E-cadherin and P-cadherin appears to be important for mammary gland differentiation. E-cadherin is expressed by luminal epithelial cells, whereas P-cadherin is restricted to myoepithelial cells. 101 However, during late pregnancy and lactation in humans and dogs, P-cadherin is not found at cell–cell borders, as expected for an adhesion molecule, but rather appears to be secreted by the epithelial cells. 101
In human breast cancer, P-cadherin is frequently overexpressed in high-grade invasive carcinomas, and it is a well-established indicator of poor prognosis. 101 In canine mammary neoplasms, there is a significant correlation between P-cadherin expression and higher tumor grade, suggesting that aberrant P-cadherin expression is a marker of biological aggressiveness of these tumors. 29 Furthermore, aberrant P-cadherin expression in these tumors is associated with an infiltrative tumor growth pattern. 30
The interaction of β-catenin with cadherin on the cell membrane and the cytoplasmic α-catenin is crucial for effective cell adhesion. For instance, tyrosine phosphorylation of β-catenin disrupts its binding to cadherin, leading to decreased cell–cell adhesion. 121 In addition to its function as a component of the cadherin complex, β-catenin is an integral component of the WNT signaling pathway that controls cell proliferation and differentiation. Both functions of β-catenin are deregulated in cancer, thereby leading to the loss of cell–cell adhesion and to the increased transcription of WNT target genes. 96
In CMT, Brunetti et al demonstrated that reduced β-catenin expression is correlated with progression from noninfiltrating to highly infiltrating malignant mammary tumors and therefore with increased invasiveness of these tumors. 7 Moreover, reduced β-catenin expression, which is accompanied by reduced E-cadherin expression, is significantly associated with a shorter overall survival and disease-free period. 30 Nevertheless, Restucci et al did not confirm these findings. They observed a switch of β-catenin membrane expression toward cytoplasmic accumulation in CMT that often coincided with disrupted APC protein expression in the WNT pathway. 117
Sialylated Antigens
Sialylation plays a role in a variety of biological processes, such as cell–cell communication, cell–matrix interaction, adhesion, and protein targeting during the process of tumor development, differentiation, and progression. 126 Sialylation is defined as the transfer of sialic acid to an oligosaccharide. Sialic acids are typically linked to cell surface glycoproteins and glycolipids via α2,6 or α2,3 linkage to galactosamine or to N-acetylgalactosamine via α2,6-linkage. 126
The most widely reported modifications of sialylated antigens associated with cancer are Sialyl Tn, Sialyl Lewis a, and Sialyl Lewis x (sLex). 13 A recent study in CMT reported that each of the studied antigens—Sialyl Lewis a, T, and Tn—was an tumor-associated antigen in CMT. 99 However, only the T and Tn antigens seemed to be associated with malignant transformation of mammary gland cells and were of potential value as diagnostic markers. 99
sLex is involved in the attachment of leukocytes to cytokine-activated endothelial cells through its interaction with the endothelial adhesion molecule E-selectin, leading to leukocyte extravasation and migration into the tissues during inflammation. 92 The involvement of sLex antigen in tumor malignancy and progression resembles leukocyte–endothelium interactions in the inflammatory response. Thus, as in inflammation, sLex expressed on tumor cells binds to E-selectin expressed on cytokine-activated endothelial cells, culminating in the extravasation of tumor cells to form new tumor foci. Therefore, sLex is closely associated with human breast cancer hematogenic metastasis and shorter survival of human breast cancer patients. 13,107
In canine mammary carcinomas, sLex is overexpressed compared to normal mammary gland and benign proliferative lesions, both negative for sLex expression. 104 Therefore, de novo expression of the sLex determinant may be considered a marker of the malignant phenotype in canine mammary carcinomas. There is also a significant association between levels of sLex and the presence of lymph node metastases, which suggests that sLex is important for local invasion and metastasis of CMT. 104 Moreover, Nakagawa et al recently suggested that sLex expression might be involved in the hematogenic spread of malignant CMT. 95 The role of sLex in the metastatic process of CMT and in the cooperative interactions among adhesion molecules in CMT progression is supported by another study. 104 Indeed, an inverse relationship between sLex and E-cadherin expression was observed in canine mammary carcinomas and in a model of inflammatory breast cancer in women. 106 The results suggest a biologically coordinated mechanism of E-cadherin and sLex expression, termed molecular plasticity, which could be essential for tumor establishment and metastatic progression.
Mucins are another important family of glycoproteins that undergo changes in glycosylation upon malignant transformation. Mucins are a family of heavily glycosylated proteins of high molecular weight, expressed by epithelial tissues. In the mammary gland, mucin 1 (MUC1) glycosylation has been studied most extensively, and MUC1 overexpression has been associated with a poor prognosis in human breast cancer. 135 In canine mammary neoplasms, there is a significant association between MUC1 overexpression and the presence of distant metastases. 17 However, additional studies are required to further evaluate the specific role of MUC1 in CMT.
Claudins
Epithelial cell–cell contacts contain another major adhesive structure: tight junctions (Fig. 10). Claudins are one of the 3 main types of transmembrane proteins of the tight junctions, together with occludins and immunoglobulin G–like family of junctional adhesion molecules. 97 The major role of claudins is to form paired strands through homophilic and heterophilic interactions with claudins on neighboring cells. 26 Claudins 1, 2, 3, 4, 5, and 7 are commonly expressed on the normal canine mammary gland, indicating that loss or reduction of expression of claudins 1, 2, 5, and 7 may lead to cellular detachment and invasion of CMT. 51
Connexins
Gap junctions represent another important type of cellular junction found in vertebrate epithelial tissues (Fig. 10). Gap junctions are the only communicating junction in animal tissues that directly connect the cytoplasm of 2 cells. 99 Connexins, or gap junction proteins, are a family of structurally related transmembrane proteins that assemble into groups of 6 to form hemichannels, with 2 hemichannels combining to form a gap junction. 32 Several studies have suggested that decreased connexin expression and, therefore, a reduction in gap junction intercellular communication play a role in human caner development. 12 In canine mammary carcinomas, there is a decrease in the expression of connexins 26 and 43, and their expression and distribution are inversely correlated with cell proliferation. 132 Furthermore, connexin 26 mRNA expression is lost in malignant mammary tumors. 38
CD44
The CD44 antigen is a cell surface glycoprotein involved in cell–cell interactions, cell adhesion, migration, and lymphocyte homing and activation. 36 CD44 is an important receptor of hyaluronic acid, a major component of the extracellular matrix; therefore, it is involved in cell–extracellular matrix interactions. 36 In human breast cancer, CD44 has been implicated in the progression of breast tumor cells by promoting vascular and tissue invasion, and it is considered a useful prognostic factor. 36 In CMT, little is known about the clinical and pathologic relevance of CD44. Decreased CD44 expression was associated with increased tumor size, infiltrative tumor growth, and vascular invasion of some histologic types of canine mammary carcinomas. 73 However, other studies found CD44 to be upregulated in benign CMT when compared to carcinomas, or they found no differences among normal mammary gland, adenomas, carcinomas, and lymph node metastases. 58,100
HEPACAM1 and HEPACAM2
Expression levels of the p53-regulated hepatocyte cell adhesion molecules 1 and 2 (HEPACAM1 and HEPACAM2) have recently been studied in benign and malignant CMT compared to normal glands of the same dog. 58 An approximately 0.5-fold decrease in HEPACAM 2 mRNA expression levels was found in 12 of 13 carcinomas and in 13 of 13 lymph node metastases but not for HEPACAM 1 protein, which was expressed in normal gland, carcinomas, and lymph node metastases (Fig. 13). 57 However, no HEPACAM1 protein expression was observed in adenomas, which showed a loss of HEPACAM1 compared with normal and malignant tumor cells. 57 HEPACAM2 might therefore be involved in maintaining physiologic mammary cell adhesion in normal glands and benign neoplasms, whereas it is reduced or even lost during malignant progression. In contrast, HEPACAM1 protein expression does not function as a tumor suppressor at late stages of malignant transformation, but it might be involved in physiologic mammary cell adhesion and CMT metastasis. 57
Angiogenesis: Supply Unit and Exit Strategy for Tumor Cells
Angiogenesis is the formation of new capillaries from preexisting blood vessels in the adult. It is induced in some physiologic and pathologic (wound-healing) nonneoplastic conditions, and it has a great impact in the biology of tumor growth and metastasis. 37 There are 3 main functions of these newly built vessels in the tumor: supply of the rapidly growing tumor mass with nutrients and oxygen, removal of metabolites, and an easily accessible exit for metastatic tumor cells that will enter the circulation of the animal. 14,37 Angiogenic activity of a tumor is the result of the action of proangiogenic factors or antiangiogenic inhibitors on endothelial or stromal cells that subsequently form capillary sprouts. Another form of angiogenesis was recently described in inflammatory CMT: vasculogenic mimicry. 11 In this process, microvascular channels are produced by tumor cells, called endothelial-like cells, without endothelial cell participation. These endothelial-like cells express cytokeratins AE1/AE3 and 14 and are mostly negative for endothelial markers. 11 However, further studies on noninflammatory carcinomas will reveal whether this form of angiogenesis can be observed in other histologic types of canine mammary carcinomas.
One of the most important and most studied stimulators of angiogenesis in CMT is the vascular endothelial growth factor (VEGF) protein family. Expression analysis of ligands and receptors of the VEGF family is an indirect way to quantify the angiogenic potential and mitogenic stimulus that tumors exert on endothelial cells. In CMT, both VEGF and the receptor VEGFR2 (flk-1/KDR) have been investigated immunohistochemically. 81,84,115,118 VEGF protein is synthesized and localized in the cytoplasmic granules of neoplastic epithelial cells, endothelial cells, and stromal cells, indicating that both autocrine and paracrine signaling induces proliferation of endothelial sprouts. 84,123
However, the cytoplasmic expression pattern differs between benign and malignant neoplastic mammary cells. In benign tumors, a small number of cells contain few, small, weakly VEGF-positive granules, localized to the luminal aspect of the cell. In contrast, a large number of cells in carcinomas often contain large, strongly VEGF-positive granules diffusely dispersed in the cytoplasm. 118 In addition, tumor-infiltrating macrophages show a strong VEGF cytoplasmic expression and may therefore contribute to the angiogenesis in the tumors. 84,123 The quantification of VEGF ligand and receptors has been performed quantitatively (ie, the percentage of positive cells) and semiquantitatively (ie, cases considered positive [> 10% of cells] or negative [< 10% of cells]), or slides are scored and assigned to one of 4 groups: < 25%, 25–50%, 50–75%, > 75%. 81,84,115,123
Based on these quantification methods, contradictory results have been obtained regarding malignancy-associated VEGF expression. Whereas significantly higher VEGF expression was reported in noninflammatory and inflammatory carcinomas, 81 other studies found no differences between benign and malignant tumors. 84,123 When related to histopathologic or clinicopathologic features, VEGF expression was associated with tumor grade in one study 118 but not with any clinicopathologic parameter, such as size, lymphatic invasion, and survival in another study. 84 Significantly higher VEGF expression has, however, been found in inflammatory carcinomas when compared to noninflammatory carcinomas. 81 The latter study suggested that VEGF expression occurs in benign and malignant tumors and is independent of histologic type, proliferation, tissue invasion, or local metastatic capacity. 81 VEGFR2 expression in endothelial cells was higher in malignant than in benign tumors. 115
Angiopoietin (ANG) 1 and 2 are the only other angiogenic factors beside VEGF that have been analyzed immunohistochemically in CMT. Both have been identified in carcinomas, but their expression level, distribution, or association with other clinicopathologic factors has not been analyzed. 52
Comparison of the transcriptome of metastastic versus nonmetastastic simple canine carcinomas revealed a surprising downregulation of 7 genes involved in angiogenesis, including NRP1, JMJD6, CYR61, FGFR1, in the metastatic tumors (Fig. 4). 60 Similarly, when compared to the normal gland of the same dog, the transcriptome of metastatic carcinomas had an increase in several angiogenic genes (CEACAM1, CXCR4, GJC1, ITGA4, SERPINE1, VEGFA) but a concomitant downregulation of other, well-described proangiogenic genes (ANGPT2, ANGPTL1, ANGPTL2, ANGPTL4, FGFR1, FIGF, TIE1). 59 These results have yet to be confirmed on the protein level. However, the mRNA data indicate that the simple view—that high expression of proangiogenic genes equals malignancy—may not be true for each gene or pathway analyzed.
Blood supply, as estimated by the microvessel density (MVD) in a tumor, has a major impact on the tumor metabolism and behavior. Therefore, the prognostic relevance of MVD has been intensely analyzed in CMT. Microvessels are defined as those vessels with a transverse diameter less than 50 μm, 39 but in some studies small venules were included in the count. 84,116 The MVD is best visualized by immunohistochemical staining of endothelial cell with markers such as FVIIIRA, 39 PECAM/CD31, 84,116 and claudin 5. 50 Clusters of cells positive for endothelial markers without a clear lumen or single positive cells are also counted as microvessel entities. The fields scored are those of highest microvessel concentration (hot spots), generally found at the perimeter of the tumors, or randomly selected fields. 39,84,116 Interestingly, MVD values are almost similar when obtained from “hot spot” areas or from randomly selected fields (Table 2). 83,116 Microvessels in areas of inflammation, ulceration, and granulation tissue are excluded. Finally, the vessel density count is reported as the mean MVD—that is, the mean number of microvessels per square millimeter (Table 2).
Investigations Into the Microvessel Density of Canine Mammary Tumors a

a Number of cases and values of investigations into the microvessel density of canine mammary tumors. The data are those directly reported by the authors or as calculated from data available in the articles.
b Values expressed as mean 200× magnification and corresponding to an area of 0.27 mm2.
c Values expressed as mean 200× magnification.
d Values expressed as mean 200× magnification and corresponding to an area of 0.304 mm2.
In CMT, the MVD count is significantly lower in benign mammary tumors than in malignant. 39,116 In addition, MVD in carcinomas is correlated with histologic type and tumor grade but not with lymphatic invasion and tumor size. 84,116 Clinicopathologically, the mean survival times of dogs with values lower than the median MVD were longer (23.7 months) than those of dogs with higher values (17.5 months), with a significant association between shorter survival and higher MVD. 84,116
In addition to the MVD count, the area and perimeter of the vessels differ in tumors at different stages of malignancy. In one study, an association was observed between tumor grade and the width and shape of microvessels. Benign neoplasms and grade I carcinomas had wide and regularly shaped microvessels with erythrocytes in their lumina, whereas grade II and III carcinomas contained small and irregular microvessels, often without a distinct lumen, or they appeared as isolated endothelial cells. 116
Intravascular or circulating tumor cells (CTC) are neoplastic cells circulating in the peripheral blood or lymph vessels of patients, as shed from the primary tumor or its metastases. 1 In human breast cancer, CTC are rather rare events, with 1 CTC in 10 6 to 10 7 peripheral blood leukocytes. 120 Despite this rarity, several studies on human cancer showed that CTC are detectable in the peripheral blood long before metastases are recognized clinically and are an independent prognostic factor for overall survival. 43 Recently, a first study showed that known human CTC marker genes, including cytokeratin 19, ERBB2, epidermal growth factor receptor, and claudin 7, are also potential markers for CMT CTC. 14 In addition, microarray technology identified several new canine-specific mRNA types that may be used for detection of CMT CTC in the peripheral blood of dogs with mammary tumors. 14 However, further studies are needed to evaluate the predictive potential of CTC for the disease outcome in dogs.
Taken together, both MVD and VEGF signaling contribute to the biology and clinical behavior of canine mammary carcinomas. However, at least for VEGF expression levels, their influence on postoperative survival seems minor.
COX2: A Potential Therapeutic Target in CMT
Cyclooxygenase (COX) catalyzes the formation of prostaglandins from arachidonic acid. Two isoforms of the enzyme have been identified: COX1 (constitutive) and COX2 (inducible). COX2 upregulation can be induced by hypoxia, growth factors, inflammatory stimuli, and several oncogenes. In tumors, prostaglandins promote resistance to apoptosis, proliferation, enhanced angiogenesis, invasiveness, and immunosuppression—typical features of malignant tumors. 20 In canine mammary samples, both COX1 and COX2 have been investigated, 109,111 with more studies focusing on COX2 expression. 22,69,82 Most of these studies used different primary antibodies but a similar, semiquantitative scoring system. Briefly, positivity, distribution, and intensity of COX immunoreactivity are evaluated at 200× magnification fields. First, the percentage of positive cells is scored as 0 to 4—respectively, 0%, < 10%, 10–50%, 51–80%, > 80% positive cells. Second, the staining intensity is scored as 0 to 3—negative, weak, moderate, strong. Finally, the scores for percentage of positive cells and the staining intensity are multiplied and thus result in a COX score between 0 and 12. 24,109,111 Slight modifications of the scoring system were used in other investigations. 45,69,82
Use of this scoring system with different antibodies and different antigen retrieval methods lead to contradictory results on COX2 expression in normal canine mammary gland (Table 3). No expression was found using a goat anti-COX2 antibody. 82 In contrast, weak cytoplasmic expression was found using the primary antibody COX2 AB 82 or clone 33 109 in lobules of normal gland surrounding benign or malignant lesions. Cytoplasmic expression was weaker or similar to the tumor when the primary antibody was clone 33, 109 MF243, 23 or SP21. 22 Application of antibody clone SP21 found COX2 expressed in normal mammary tissue with a membrane on the apical luminal epithelium during dioestrus and weak diffuse cytoplasmic labeling of the terminal end bud epithelial cells. 22
Primary COX2 Antibodies and Antigen Retrieval Information a
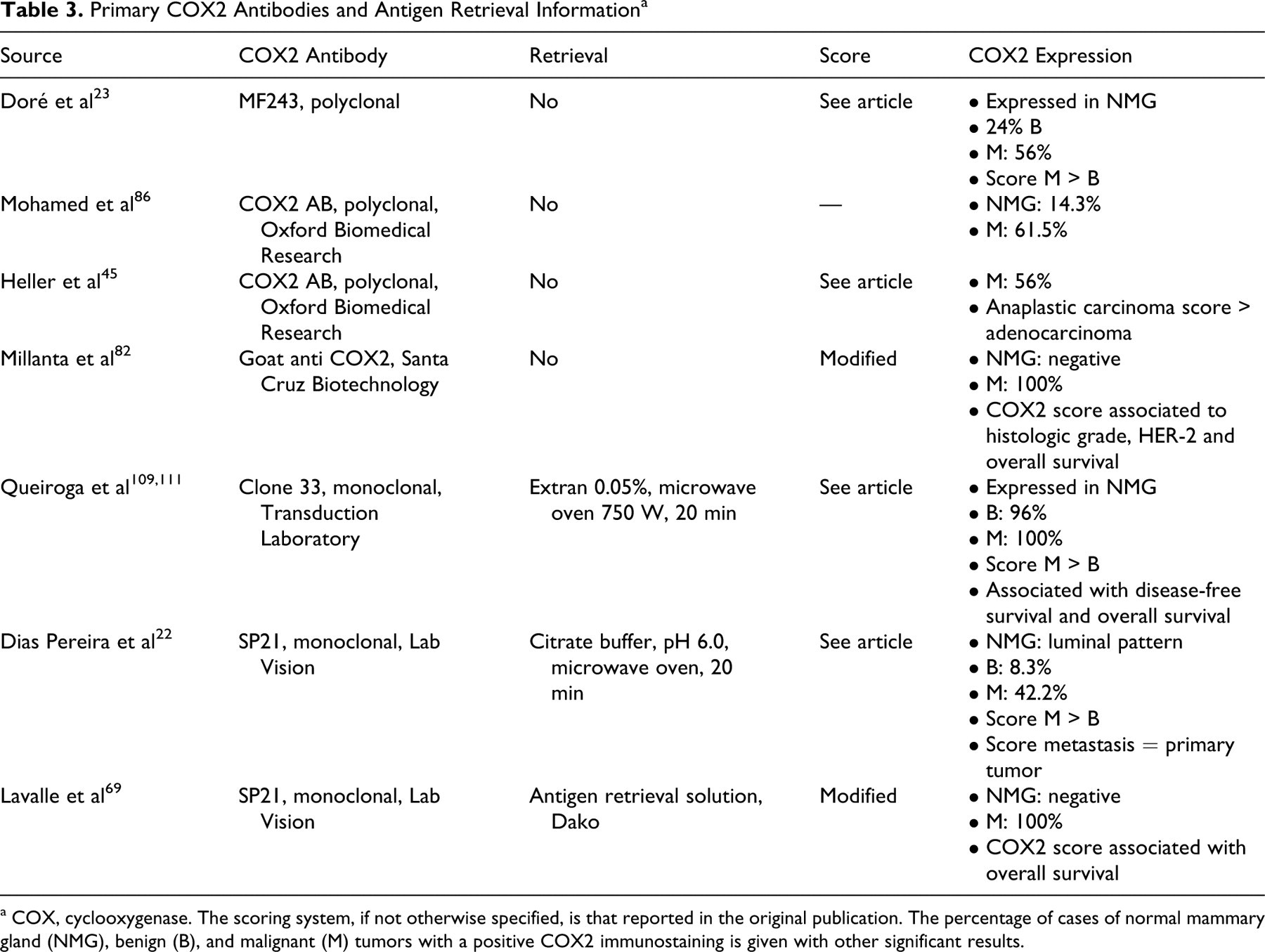
a COX, cyclooxygenase. The scoring system, if not otherwise specified, is that reported in the original publication. The percentage of cases of normal mammary gland (NMG), benign (B), and malignant (M) tumors with a positive COX2 immunostaining is given with other significant results.
In benign tumors, COX2 immunohistochemistry was variable in distribution (from 10% to 75%), and the intensity ranged from weak to moderate. Furthermore, COX2 expression was prevalent in luminal cells of benign tumors and, to a lesser extent, in chondroblasts of mixed tumors and in myoepithelial cells. 22 In malignant tumors, the expression was moderate to intense in the luminal neoplastic cells and occasionally positive in fusiform stromal cells, but in carcinomas there was also perinuclear and nuclear staining. 23,109 When different histologic subtypes of mammary carcinomas were compared, anaplastic tumors (versus better-differentiated carcinomas) had a significantly higher number of cells that stained positive with increased staining intensity, thereby indicating an increase in COX2 expression with increasing malignancy of the tumors. 38 In addition, COX2 levels were higher in lymph node–positive carcinomas than in lymph node–negative carcinomas and similar or higher in distant metastases and the primary tumors. 22,111 However, COX2 score was not associated with tumor size or histologic type, necrosis, vascular endothelial growth factor expression, vessel invasion, hormone receptor expression, and Ki67 index. 82,111
Clinically, high COX2 scores were significantly associated with the development of postoperative distant metastasis, disease-free survival, and shortened overall survival, 69,82,111 whereas in multivariate survival analysis, COX2 did not retain its significance as an independent prognostic factor for either disease-free interval or overall survival. 111
In summary, increased COX2 expression seems to be associated with increasing malignancy of CMT, whereas COX1 expression is not significantly associated with any clinicopathologic feature of CMT. The high expression of COX2 in aggressive tumors, however, indicates that COX2-selective inhibitors may be a useful alternative option for the treatment of mammary carcinomas. COX2 therapy has been demonstrated as not being useful in human cancer treatment. Nevertheless, veterinary clinical studies on the usefulness of a preoperative or postoperative treatment of CMT patients are needed to evaluate this hypothesis.
Conclusions
Although CMT are among the most common tumors of the dog, their molecular carcinogenesis is far from being understood, and therapeutic options are scarce. Recent advances in molecular tools for dogs, including complete coverage of the canine genome, cDNA microarrays, and proteomic analyses, have drastically increased the understanding of this group of tumors. Aberrant expression levels of several functional classes of genes—including mediators of apoptosis, DNA repair systems, proto-oncogenes, tumor suppressors, cell adhesion molecules, COX2, and angiogenesis—are clear hints toward the potential molecular mechanisms behind the development of CMT, and these genes and the proteins they encode may represent new potential therapeutic targets. However, veterinary oncology is still in its infancy when it comes to defining driver genes of carcinogenesis with initially relevant mutations and mere passenger genes whose expression levels are only reactively changed. Nevertheless, a fundamental lack of known driver genes is true for human breast cancer research, despite much higher financial and structural resources in that field.
A major focus in human cancer research and a future “hot spot” for veterinary oncologists is therefore the increasingly available high-throughput sequencing technology for complete cancer genomes. First and mostly preliminary results from human cancer genome studies indicate that the redundancy in gene functions is reflected in individual mutation patterns in each tumor despite their similar histologic and clinical appearances. It will therefore be fascinating to see whether the increasing knowledge on the molecular basis of CMT will lead to new prognostic and clinical classification schemes and initiation of an individualized patient-specific or even tumor-specific veterinary oncologic chemotherapy.
Footnotes
Acknowledgment
This study represents an initiative of the American College of Veterinary Pathologists' Oncology Committee. We would like to thank the college for its support and guidance.
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Research by R.K. and A.D.G. is funded by the German Research Council (DFG KL 2240/1). H.V. is funded by the European Union Seventh Framework Programme for Research and Technological Development of the European Commission, LUPA (GA-201370), and AGRIA Pet Insurance Research Foundation. Research by S.S.P. and F.G. was supported by grant No. PTDC/CVT/65537/2006 from the Portuguese Foundation for Science and Technology; S.S.P. also acknowledges foundation grant No. SFRH/BPD/63094/2009. The Institute of Molecular Pathology and Immunology of the University of Porto is an associate laboratory of the Portuguese Ministry of Science, Technology and Higher Education and is partially supported by the foundation.