
Abstract
The genus Henipavirus contains two members—Hendra virus (HeV) and Nipah virus (NiV)—and each can cause fatal disease in humans and animals. HeV and Niv are currently classified as biosafety level 4, and NiV is classified as a category C priority pathogen. The aim of this article is to discuss the pathology of laboratory animal models of henipavirus infection and to assess their suitability as animal models for the development and testing of human therapeutics and vaccines. There has been considerable progress in the development of animal models for henipavirus disease. Suitable animal models include the golden hamster, ferrets, cats, and pigs, which develop disease resembling that observed in humans. Guinea pigs are a less reliable model for henipavirus disease, but they do develop henipavirus-induced encephalitis. Because human efficacy studies with henipaviruses are not permitted, animal studies are critical for the development of antiviral therapeutics and vaccines. Current research indicates that passive immunotherapy using monoclonal antibodies is protective of ferrets against NiV infection and that passive immunotherapy using NiV antibodies protects hamsters from HeV. Recombinant vaccines have been used to protect cats and pigs against NiV infection. Ribavirin and 6-aza-uridine were able to delay but not prevent NiV-induced mortality in hamsters. Further research is needed to develop a model and therapy for late-onset henipavirus encephalitis.
The genus Henipavirus, family Paramyxoviridae, has two relatively recently discovered members: Hendra virus (HeV) and Nipah virus (NiV). Both viruses can cause human and animal disease, and both continue to pose a threat with sporadic disease outbreaks. Henipaviruses are single-stranded negative-sense RNA viruses related to morbilliviruses and respiroviruses. 42,81 Their genomes are among the largest of the paramyxoviruses with the HeV genome comprising 18,234 nucleotides and the NiV, 18,246 nucleotides. 33 This increased genome size is due to the long untranslated regions at the 3′ end of most transcription units. 27,75 The amino acid sequence identity between the N, P, and L proteins of HeV and NiV is 92%, 71%, and 87%, respectively. 27 Molecular characterization of NiV from Malaysia, Cambodia, India, and Bangladesh indicates that there are strain differences in NiV. 32,35,43
HeV and NiV can infect various animal species, and they each have a wide tissue tropism. Henipaviruses grow in vitro in a range of mammalian, avian, reptile, amphibian, and fish cells. 27 The pathogenicity of henipaviruses is related to the ability of the virus to circumvent the host interferon response. 16,72 However, henipaviruses are sensitive to pH, temperature, and desiccation, indicating a need for close contact between hosts for transmission to occur. 19 Bossart recently reviewed an assessment of henipavirus therapeutics and vaccines, 5 and human henipavirus vaccines and passive immunotherapy will likely be available in the future. 6,23 The aim of this article is to discuss the pathology of laboratory animal models of henipavirus infection and to assess their suitability as animal models for the development and testing of therapeutics and vaccines.
Hendra Virus
Since 1994, there have been 7 human HeV infections. Four humans have died of HeV infection, 1 of respiratory disease and 3 with encephalitis. Three other humans, previously infected with HeV, remained healthy without evidence of further disease. 2,57 All human HeV infections have been associated with horses at necropsy or during the care of infected horses. HeV is classified as biosafety level 4 (BSL-4), and guidelines for handling infected horses have been developed by the Queensland Government of Australia. 2
The first known human case of HeV infection was associated with an outbreak of fatal respiratory disease in horses in the Brisbane suburb of Hendra, Queensland, Australia, in 1994. 53 A horse trainer died with severe respiratory disease, and a stable hand became infected, seroconverted to HeV, but recovered. 66 A second fatal human case occurred in Mackay, Queensland, with HeV exposure from 2 horses. 57,63 His infection could be traced to exposure in August 1994 from 2 fatal horses. 57,63 Thirteen months later, this farmer developed neurological symptoms and a severe nonsuppurative meningoencephalitis. In November 2004, a veterinarian conducting a necropsy on a horse developed a dry cough, a sore throat, cervical lymphadenopathy, and fever, but he recovered and remained in good health. 31 In June 2008, there was fatal HeV disease in 5 horses at a Queensland veterinary clinic. Two staff members became ill with HeV disease, one of whom died (a veterinarian). 60 In August 2009, a veterinarian who treated 2 horses became infected with the virus and died 3 weeks later. 61
In horses, there have been 13 episodes of HeV infection, all in Australia; 2 12 of these occurred in the state of Queensland and 1 in northern New South Wales. Infection in horses is typically associated with close proximity of Pteropus spp. HeV infection is usually sporadic, affecting a single horse in a paddock, although multiple fatalities have occurred where horses have been in close proximity in stables. 53,60 HeV disease in horses is of low morbidity but high mortality, with a case fatality rate of approximately 75% in natural infections. Horses diagnosed with HeV infection develop clinical disease between 5 and 16 days postexposure. 17,36,53,63,84 Symptoms include increased respiratory rate, dyspnea, fever, increased heart rate, lethargy, neurological signs, muscle trembling, hemorrhagic nasal discharge, and swelling of the face, lips, neck, and supraorbital fossa. 36,53,63
Experimental work by Middleton indicates that infected horses have a potential to excrete virus through nasal secretions from 2 days following exposure to HeV. 2 The most significant gross lesions in horses are diffuse pulmonary edema and congestion, with marked dilation of the pulmonary lymphatics. Less frequently, there is hydrothorax and hydropericardium, edema of the mesentery, and congestion of lymph nodes. 36,82 In field cases but not experimental cases, there is thick froth in the trachea, which exudes from the nares. 53,84,83 Microscopically, there is severe diffuse necrotizing interstitial pneumonia with fibrinoid necrosis and thrombosis of blood vessels, pulmonary edema, and increased alveolar macrophages. 36,82 There is vascular and endothelial necrosis in many tissues, and endothelial syncytial cells are a unique histological feature of HeV infection in all species. 36,53,83
With an overall HeV antibody prevalence of 42% in wild-caught flying foxes, 46 flying foxes are regarded as the reservoir host of HeV. 28,91 HeV has been isolated from the uterine fluids of a grey-headed flying fox (Pteropus poliocephalus). 29,30,85 P poliocephalus has been shown to be susceptible to experimental HeV infection and subclinical disease. 84,85 There is a need for a greater understanding of the interaction between the HeV and flying foxes. HeV antibodies have not been identified in people caring for wildlife, 30 and there is no known human HeV infections contracted from flying foxes. There have been no HeV infections in cats or other domestic nonequine animals in Australia or elsewhere.
Nipah Virus
NiV was first identified in peninsular Malaysia in September 1999, and it caused 265 human cases of encephalitis, with a mortality rate of 40%. Pigs are the amplifying host. 11,10 Oral ingestion or aerosol inhalation is thought to be responsible for pig-to-human transmission of NiV. 58 The disease in pigs was highly contagious and characterized by acute fever with respiratory and nervous signs. 10,41 Since 1999, human NiV cases have occurred in Bangladesh in 2001–2005, 2007, and 2008, as well as in India in 2001 and 2007. 8,11,40,43,44,47 Some of the recent human NiV cases in Bangladesh were primarily respiratory rather than encephalitic. 5,34,47,49,50 The National Institute of Allergy and Infectious Biodefense Research Agenda classified NiV as a BSL-4 and category C priority pathogen. 7 Risk factors for NiV transmission are human-to-human contact, contact with infected animals, food-borne transmission, and nosocomial transmission. 24,25,38,39,44 In Bangladesh, human-to-human transmission accounts for approximately 51% of the recognized cases. 44 One report indicated that human infection with NiV was a result of consuming palm date juice contaminated with NiV; as such, it may be that domestic animals are infected through ingestion of masticated fruit pulp from flying foxes. 18
In humans, NiV causes a severe, rapidly progressing febrile encephalitis 11,21 or a severe respiratory disease. 13,39,59,88 Human NiV disease results in widespread vasculitis and thrombosis, particularly in small arteries, arterioles, venules, and capillaries; there are also NiV-induced endothelial syncytial cells. 15,88 Viral inclusions can be observed in cases of encephalitis. 15 A relapsing, late-onset neurologic disease occurs months after the initial acute NiV disease episode has been reported. 54,65,67,89
The natural reservoir host of NiV is the Pteropus bats. 11,12,56,62,74,90 Unlike notable viral agents of biodefense concern, such as smallpox or Ebola virus, NiV can be isolated from natural hosts 11,73 and readily grown in cell culture to high titers. 14 Experimentally, NiV infections in P poliocephalus do not develop overt clinical disease. 48 NiV is intermittently shed in the urine, 12 which may be sufficient to maintain the virus in the bat population.
In the 1999 Malaysian outbreak, many pigs became infected with NiV, but a large number remained asymptomatic. Pigs were probably infected by fruits contaminated by bat secretions and body fluids. 34 Pigs younger than 6 months developed respiratory disease, whereas older pigs developed neurological signs. 76 Approximately 95% of NiV-diseased pigs recovered. 55 In pigs, there is evidence of pig-to-pig spread by oronasal secretions. 34 Respiratory lesions include tracheitis, bronchitis, and interstitial pneumonia characterized by the presence of syncytial cells with intracytoplasmic inclusion bodies. 5,70 All pigs with central nervous system lesions had meningitis, but encephalitis was less common. 34 NiV antigen using standard immunohistochemistry 68 can be found in a variety of tissues. 34,68
Only one field case of NiV in cats has been confirmed by necropsy and immunochemistry. 34 One horse with NiV had a nonsuppurative meningitis. 34 A serological survey showed that 46 to 55% of dogs from infected pig farms had antibodies to NiV virus but that only 2 had NiV disease. 34
Pathology of Laboratory Animals
Hendra Virus
Golden hamsters
The golden hamster is highly susceptible to HeV infection. 23 Hamsters inoculated with as little as 10 plaque-forming unit (PFU) by intraperitoneal (IP) injection developed paralysis, trembling limbs, dyspnea, and prostration and died 14 and 27 days postinoculation (PI). The median lethal dose (LD50) was 12 PFU: Hamsters inoculated with 1,000 LD50 died within 5 days PI, and hamsters inoculated with 100 LD50 died from 5 days to 12 weeks PI. Lesions were first observed 2 days after inoculation, and antigen was detected with standard immunohistochemistry methods. 32 Increasing amounts of HeV antigen can be found from days 3 to 5, with nodular lesions and necrotizing vasculitis in the lung, liver, kidney, spleen, and endothelial syncytial cells. By 4 days, there was positive immunoreactivity in the brain, lung, kidney, spleen, liver, and heart. By 5 days, lesions and positive immunoreactivity were observed in the brain, lungs, kidney, heart, liver, and spleen. Lesions in the brain were in the cerebellum and cerebral cortex, with positive immunoreactivity in the neurons, meninges, blood vessels, and ependyma. Positive immunoreactivity was observed in the glomeruli, tubules, and blood vessels of the kidney. In the liver, immunoreactivity was restricted to the vasculature and was not identified in the parenchyma.
Cats
Cats are susceptible to HeV infection and are a good model of disease, with its similarity to HeV infection in humans and horses. Cats inoculated with 5,000 and 50,000 TCID50 of a low-passage, plaque-purified HeV by subcutaneous injection, oral, or intranasal (IN) administration develop severe dyspnea, open-mouth breathing, and reluctance to move within 4 to 10 days PI. 80,84 HeV can be isolated from the lung, spleen, brain, kidney, and urine of experimentally infected cats. 80 Furthermore, HeV has been transmitted from a subcutaneously inoculated cat to an uninoculated cat housed in the same cage. 79 The gross lesions of HeV in cats are hydrothorax; heavy, wet, congested lungs; and edematous lymph nodes (Figs. 1, 2). Cats develop severe pulmonary lesions with pulmonary edema, necrosis of alveolar walls, and vascular lesions. There is fibrinoid necrosis and positive immunoreactivity in many small blood vessels in the lung, kidney, spleen, gastrointestinal tract, and lymph nodes (Fig. 3). HeV antigen can be demonstrated by standard immunohistochemistry methods in the vascular walls of a variety of tissues. 37,84
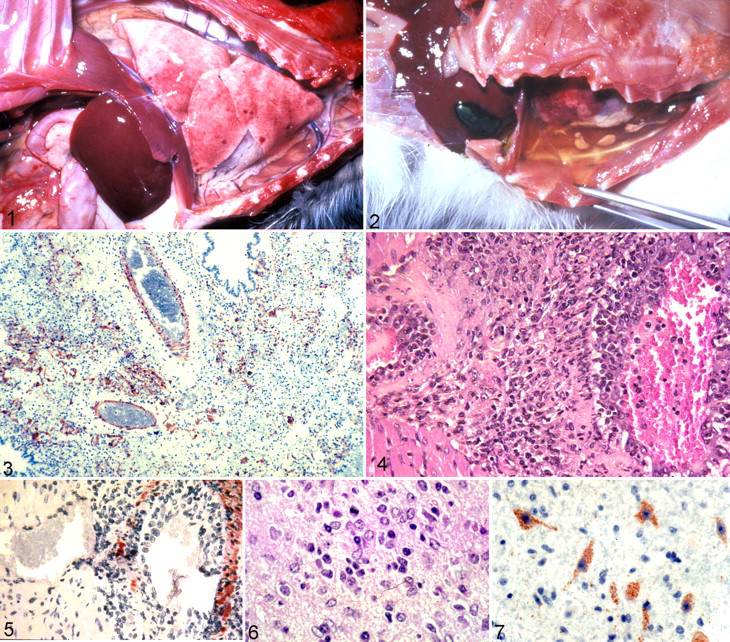
Guinea pigs
Guinea pigs inoculated with HeV are a useful model of encephalitis. However, they demonstrate little or no pulmonary disease while developing generalized vascular disease, and their experimental infections are not typical of the HeV respiratory infection in humans or horses. Overall, the disease in guinea pigs is qualitatively different from that in cats, and guinea pigs take longer to die. Guinea pigs inoculated with 5,000, 30,000, or 50,000 TCID50 HeV by subcutaneous injection develop severe clinical disease between 7 and 15 days PI. 80,85 Guinea pigs inoculated IN or intradermal do not develop severe systemic disease. 86 Gross lesions include cyanosis, with congestion and edema in the gastrointestinal tract. Histologically, vascular lesions are in the small to medium-sized arterioles and veins in the lung, kidney, spleen, lymph nodes, gastrointestinal tract, and skeletal and intercostal muscles. 33 Vascular lesions are characterized by large numbers of perivascular lymphocytes and macrophages and vascular fibrinoid necrosis (Fig. 4). Endothelial syncytial cells can be found in the large and small arteries and veins of the kidney, bladder, and lung. 37 In guinea pigs, there is extensive strong immunoreactivity for viral antigen 85 in the endothelial cells, tunica intima, and tunica media (Fig. 5) and in the conjunctival and bladder epithelium. 37,86 HeV can be isolated from the kidney and urine of guinea pigs. 79
Encephalitis is a feature of HeV disease in guinea pigs, making this a suitable model system to study HeV encephalitis. The source of inoculum does not appear to be important in the production of encephalitis, 86 as characterized by lymphocytic perivascular cuffing, gliosis, neuronal necrosis, and severe fibrinoid degeneration and necrosis of blood vessels in the medulla, cerebellum, and thalamus (Fig. 6). Eosinophilic intracytoplasmic inclusions are observed in neurons. The accompanying degeneration of blood vessels in HeV infection suggests that encephalitis follows a primary necrotizing vasculitis. There is positive immunoreactivity in the cytoplasm of the neurons, neuropil (Fig. 7), choroid plexus, ependyma, and blood vessels, indicating that there could be circulation of viral antigen in the cerebrospinal fluid.
HeV crosses the placenta of guinea pigs between 31 and 41 days of gestation and is associated with severe placentitis and abortion. 85 HeV can be isolated from fetuses with positive immunoreactivity, but fetal lesions have not been observed. 85 Furthermore, virus titers from tissues are generally higher in pregnant guinea pigs versus nonpregnant guinea pigs. 85 Guinea pigs would be a suitable model for the study of transplacental transmission of HeV.
Other laboratory animal infections
When inoculated with 5,000 TCID50 of HeV, mice, rats, and dogs do not develop clinical disease or seroconvert. 80 Rabbits also do not develop clinical signs of disease when inoculated, but they do seroconvert. 80 HeV can be lethal if administered intracranially into suckling mice. 50
Nipah virus
Golden hamsters
The golden hamster has been used to successfully replicate acute NiV infection in humans, developing severe respiratory disease, encephalitis, and multisystemic disease. 87 The time interval between infection and onset of clinical signs is shorter with IP inoculations that with IN inoculation. Hamsters inoculated IP die between 5 and 9 days PI and less than 24 hours after clinical signs of tremor and limb paralysis develop. Hamsters infected IN with 6 × 105 PFU of NiV die between 9 and 29 days PI. 87 Hamsters that survive following a lower viral dose seroconvert but do not develop clinical signs. Regardless of the route of inoculation, there is no difference in the pathology observed in hamsters.
In golden hamsters, NiV produces gross and histological lesions similar to those of HeV. Hamsters develop encephalitis and severe lesions in multiple organs, including the lungs. Vasculitis and thrombosis are prominent, whereas endothelial syncytia are rare. NiV antigen and RNA have been localized in blood vessels and parenchymal tissue in the brain, lung, kidney, and spleen by immunohistochemistry and in situ hybridization. Parenchymal fibrinoid necrosis in the lungs, as observed in the human patient, is rare in hamsters. 87 Histopathologic findings in the lungs of infected hamsters consist of small multifocal or coalescing areas of macrophage, neutrophil, and lymphocyte infiltration often associated with vasculitis. Syncytial cells are not observed in bronchial epithelium. In the terminal stages of the infection, virus and viral RNA have been recovered from most solid organs and urine. 87 NiV-infected hamsters can develop myocardial infarcts similar to those seen in humans. 87
The brain is a major target of NiV infection in hamsters. NiV infection causes vasculitis, ischemia, infarcts, and edema in the brain. 22,87 A mild meningitis and choroiditis has been reported; however, viral antigen was not detected in the choroid plexus with immunohistochemistry.
Cats
Experimental NiV infection in cats is similar to that seen in cats experimentally inoculated with HeV, except that with NiV there is greater involvement of the upper and lower respiratory tracts. With NiV, cats inoculated either parenterally or oronasally become febrile 6 to 8 days PI. Gross lesions include increased pulmonary edema, hydrothorax, edematous pulmonary lymph nodes, petechial hemorrhages, serosanguineous pleural fluid, mediastinal edema, froth in the bronchi, and dense red areas of consolidation in the lung. 34,47,49,50 Less frequent gross lesions include edema of the bladder wall and mild jaundice. 49
Histologically, vasculitis affects multiple sites, with endothelial syncytial cell formation. Vasculitis can be associated with infarcts in multiple organs, fibrinoid necrosis, and numerous endothelial syncytia. The respiratory tract is severely affected, with ulcerative tracheobronchitis and alveolitis and with multiple foci of infarction and widely distributed syncytial cells. 34 There is diffuse severe meningitis and vasculitis with endothelial syncytia, as well as degeneration of the trigeminal ganglion with vasculitis. Encephalitis has not been observed in cats infected with NiV, a finding similar to that of HeV-infected cats. The inflammatory process can extend into the optic tracts. 34,49 Syncytial cells are found in the renal glomeruli, renal interstitium, and tonsillar crypts.
With immunohistochemistry, positive immunoreactivity can be found along the brush border of the tracheal epithelium and in tracheal submucosa, bronchial and bronchiolar epithelium, pulmonary alveolar endothelium, epithelium syncytial cells, and connective tissue alveolar debris. In cats, NiV infection of the alveoli precedes vascular infection, suggesting an increased tropism for alveolar epithelium compared to vascular tissue. 5 There is NiV antigen in the syncytial cells, connective tissue, bladder wall, and lymphoid tissue. NiV genome can be detected in the blood 6 to 9 days PI and in oral swabs and urine at necropsy. 50,51 NiV can cross the placenta of a pregnant cat, 50 resulting in placentitis and pulmonary vasculitis in the fetus with high levels of virus in the placental fluids. 51
Guinea pigs
Guinea pigs infected with NiV have variable clinical expression of disease, possibly because of the NiV isolate used for inoculation and the route of inoculation. 31 Guinea pigs inoculated with 107 PFU by IP injection showed transient fever and weight loss after 5 to 7 days but recovered. 87 Only 3 of the 8 guinea pigs developed clinical signs when inoculated IP with 50,000 TCID50 of a nonplaque-purified third passage in Vero cells from the central nervous system of a fatal infection from a human. 48 In a study where guinea pigs were inoculated by IP injection with 6 × 104 PFU, 71 all but 2 animals died by day 8 PI.
NiV produces gross and histological lesions similar to those of HeV. Gross lesions include edema in the mesentery, broad ligament, and retroperitoneal tissues. Histologically, there is systemic vasculitis, and the most severely affected organs are the spleen, lymph nodes, urinary bladder, ovary, uterus, and brain. 45,71 Endothelial and epithelial syncytia are prominent, particularly in the urothelium and endometrium. In the lymphoid organs, there is lymphoid depletion and multifocal necrosis, particularly in the periarteriolar lymphoid sheath of the spleen. Viral antigen can be detected as early as 2 days PI in the lymphoid tissue and is subsequently widespread in the vascular endothelial, tunica media, and syncytial cells in heart, kidney, lymph node, bladder, spleen, pleura, uterus, and ovaries. 45,71
Guinea pigs develop severe meningoencephalitis characterized by nonsuppurative inflammation, with intracytoplasmic and intranuclear inclusions in neurons and microglia, especially in the brainstem. 71 NiV antigen is present in the meninges, neurons, microglial cells, and scattered ependymal cells. Guinea pigs with meningoencephalitis show subtle histopathological findings in the lung, kidney, and gastrointestinal tract. 71
Ferrets
NiV-infected ferrets develop severe respiratory and neurological disease resembling the infection in humans and are a good model of NiV disease. 6 Ferrets infected with 500, 5,000, and 50,000 TCID50 NiV by the oronasal route develop NiV disease. NiV-infected ferrets initially develop fever, then inappetence, as followed by severe depression, coughing, nasal discharge, dyspnea, and in some individuals, signs of central nervous dysfunction. 5,6 At necropsy, there is subcutaneous edema of the head, hemorrhagic lymphadenopathy of submandibular lymph nodes, and petechial hemorrhages in the lung and kidney. Histologically, there is vasculitis, pulmonary alveolitis, glomerular necrosis, splenic necrosis, cystitis, salpingitis, adrenal necrosis, thyroiditis, and less frequently, nonsuppurative meningitis. 6 Syncytial cells are frequently observed in the lesions. 5,6 NiV antigen has been detected by immunohistochemistry in blood vessels and syncytial cells in many tissues. 6 NiV genome has been detected in the adrenal gland, kidney, lung, bronchial lymph node, and spleen. Low levels have been detected in the bladder, liver, ovary, testes, and brain. In the brain, the highest levels of NiV genome were present in the olfactory bulbs.
Pigs
Several investigators have studied NiV infection in pigs. 1,34,49,76,77 Pigs develop severe respiratory disease and encephalitis following NiV subcutaneous inoculation, resembling field infections. There are no reports of field or experimental HeV infections in pigs. Pigs inoculated orally with NiV do not develop clinical signs of NiV infection. 49 At necropsy, NiV-diseased pigs have red consolidated lungs, petechiation of the pleura, prominent interlobular septa, and rarely, herniation of the cerebellar vermis. Consistent histological findings include severe fibrinous pleuritis and lobar pneumonia, with edema, necrosis, subpleural hemorrhage, thrombosis, mild focal alveolitis, increased alveolar macrophages, fibrin, and cellular debris in airways. 34 There is widespread vasculitis, with endothelial and bronchiolar syncytial cells. In the kidney, there is renal tubular degeneration, interstitial nephritis, and syncytial cells. 49 There is also histological evidence of meningitis. 34,49 By immunohistochemistry, the virus is present in airway epithelium, endothelial cells, and subjacent vascular smooth muscle in multiple organs, renal glomeruli, meninges, astrocytes, and some cranial nerves, including the trigeminal ganglion. 49 NiV can be isolated from NiV-infected pigs from nasal swabs 2 days PI. 49 Subclinical and clinically infected pigs excrete virus from oropharynx as early as day 4. 49,78 NiV can invade the central nervous system by cranial nerves or across the blood brain barrier. 76,78 Concurrent with spread through the nervous system, there is replication in the respiratory tract. 55 Nasal and oropharyngeal shedding of NiV can be detected 2 to 17 days. 1,78 NiV can be spread from pig to pig by nasal secretions. 49 Virus can be isolated from the serum at 24 days, indicating a slower clearance of virus in some NiV-infected pigs. 1
Mice
Mice are not a suitable model of NiV disease. Swiss mice inoculated IN or IP with NiV do not develop clinical signs, but NiV antibodies can be produced after repeated infection. 87 However, as with many viruses, NiV can be lethal if administered intracranially into suckling mice. 50
Chicken embryos
NiV is highly pathogenic to chicken embryos. 69 Whereas allantoic inoculation of NiV results in considerable variation and only partial mortality, yolk sac inoculation results in generalized fatal disease of chicken embryos, with gross lesions of petechial to ecchymotic hemorrhages and congestion in the kidneys. Histologically, there is severe vasculitis with hemorrhage and meningitis, myocarditis, splenitis, and proventriculitis. Immunohistochemistry is used to detect NiV antigen in the heart, arteries, kidney, spleen, brain, and feather epithelium. There is no other field or experimental evidence of NiV infection in poultry.
Suitability of Animal Models and Their Use Under the Animal Rule
Because HeV and NiV are BSL-4 pathogens, human efficacy studies are not permitted during vaccine development. Therefore, animal studies are critical for undertaking all the preliminary, intermediate, and final work in vaccine development for these two emergent pathogens. Through the animal efficacy rule, the US Food and Drug Administration heavily relies on evidence derived from animal studies for the evaluation of therapeutic products. As such, an understanding of the pathogenesis of these viral diseases is necessary to assist in the design of control mechanisms aimed at decreasing viral shedding and interrupting transmission, as well as developing vaccines and therapeutics to control and lessen disease. For consideration of potential human vaccines, it is essential that the therapeutic action be demonstrated in more than one animal species. For vaccine trials, performing all work at BSL-4 becomes problematic because of the cost and difficulty in managing animals at BSL-4. An additional problem is that animals are not often allowed to progress to serious clinical disease for welfare reasons, which means that a full understanding of the disease is not always available. 20,50
There are several suitable models of acute henipavirus infection that resemble the lethal disease observed in humans. Golden hamsters develop systemic vasculitis, pulmonary disease, and encephalitis. Ferrets develop severe respiratory and neurological disease resembling the infection in humans. 5 NiV is similar to HeV infection in cats except there is more involvement of the upper and lower respiratory tract. Cats may be a suitable model for the respiratory aspects of NiV, but they are not useful for studying the encephalitic form. Guinea pigs are a useful model of HeV-induced encephalitis, but while developing systemic vasculitis, they do not develop significant pulmonary disease. Guinea pigs appear to be less susceptible to HeV inoculation by the IN/oral route when compared to cats, which succumb to IN and oral inoculation following 5,000 TCID50. 79 These species are suitable models to develop new immunotherapeutic approaches using antiviral drug testing and vaccine development against acute NiV infection. 87 Pigs have been successfully used to replicate experimental infections of NiV, but housing and management require specialized BSL-4 facilities. 78 Chicken embryos could be a useful animal model for studying NiV and the effects on the vascular endothelium or neurons.
In NiV and HeV infections in humans, there can be late-onset henipavirus encephalitis. However, there is no animal model for this debilitating human condition. Further investigation of viral pathogenesis and enhanced comparative studies may be helpful in delineating why this problem occurs. Yet, it is difficult to conduct extended animal experiments in BSL-4 because of difficulties of husbandry and maintenance of animals under these conditions. In the future, the potential availability of henipavirus infectious clones that retain virus virulence and pathological properties in animal models may enable studies to further define viral pathogenesis. Furthermore, reverse genetic studies on various negative-stranded RNA viruses have provided a basis for the prevention of infection by these viruses and for the development of novel therapies, and these may aid further understanding of henipavirus biology. 73
Many experimental studies have focused on the end stage of henipavirus infections, but it is also important to study the early events. The key to understanding and perhaps preventing the early stages of NiV infection could be the discovery of the ephrin b2 receptor, which is involved in NiV binding and cell entry. 3 This receptor is widely expressed on endothelial cells, smooth muscle cells of blood vessels, bronchial epithelium, and neurons, which may explain the wide systemic distribution of the virus in experimentally infected animals. 26 Further work on potential receptor-blocking strategies could prove to be an effective preventive strategy.
Antiviral Therapeutics and Vaccines
Successful human vaccines often rely on inducing neutralizing antibodies that cross-react with the relevant virus, producing protective immunity. 5 The limitation to this is that for some infectious agents, effective immunity is achieved by cell-mediated immunity. However, for paramyxoviruses, the envelope glycoproteins result in the production of neutralizing antibody. 43 Several candidate henipavirus vaccines have been evaluated using different approaches, and all include the henipavirus envelope glycoproteins. 5 Work by Mungall 50 and Bossart 5 indicates that a HeV-derived vaccine would protect against HeV and NiV. 47 A canarypox NiV vaccine was protective in pigs, and it restricted virus replication and nasal and pharyngeal shedding, thereby limiting the chance for spread of the virus to uninfected animals. 77 Furthermore, one report suggested that NiV F and G proteins may be involved in inducing CD8+ cytotoxic T-cell response to NiV. 76 Hamsters have been immunized with NiV G and F glycoproteins expressed in vaccinia virus recombinants that induced an immune response and prevented fatal infection. 22 NiV and HeV sG is highly immunogenic in cats, mice, and rabbits, yielding homologous serum-neutralizing titers of greater than 1:20,000 in cats. 50 Complete protection from fatal NiV disease has been observed in 4 cats immunized three times with a 100-μg dose of recombinant HeV sG or with NiV sG. 50 With a ferret model, a neutralizing human monoclonal antibody, m102.4, targeting the henipavirus G glycoprotein fully protected NiV-infected ferrets when given 10 hours postchallenge and protected one ferret when given before the challenge. 6 Thus, m102.4 may be a useful therapeutic to treat NiV disease in humans. Passive immunization with monoclonal antibodies to either NiV G and F viral glycoproteins protects hamsters from fatal NiV infection. 22
Efficient henipavirus treatment can be achieved by the combination of drug therapy and immunotherapy. Ribavirin has been shown to be effective in the treatment of viral diseases, decreasing viral load. 64 Humans with acute NiV encephalitis who were treated with ribavirin had partial recovery from the disease. 11 Ribavirin and 6-aza-uridine have been shown to delay but not prevent NiV-induced mortality in hamsters. 20 Hamsters given the interferon inducer poly(I)-poly(C12U) daily from the day of infection to 10 days PI prevented NiV mortality in 5 of 6 hamsters. 9 RNA interference inhibits henipavirus replication in vitro, which could provide an effective future therapy. 52
The changing ecological pressures on flying foxes owing to deforestation, urban development, altered foraging, and behavioral patterns all play a role in the continuing reemergence of henipavirus infections. Since 1994, significant advances have been made in henipavirus research and the way that henipavirus infections are combated and treated. Use of appropriate laboratory animal models can provide the opportunity to study and further develop antiviral therapeutics and vaccines with which to combat henipavirus infections.
Footnotes
Acknowledgements
Hendra virus photographs are from Dr M. Williamson’s University of Melbourne doctoral thesis and studies undertaken at Australian Animal Health Laboratory–Commonwealth Scientific and Industrial Research Organisation, Geelong, Australia.
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
The authors declared that they received no financial support for their research and/or authorship of this article.