
Abstract
Anencephaly results from defects in neural tube closure early in gestation and, to the authors' knowledge, has not been reported in dogs. In this case, the canine fetus was stillborn at the 62nd day of gestation and had a hypoplastic calvarium, with flattened base of the skull and shallow orbits, causing protrusion of the eyes. Macroscopically, the brain was completely missing. Histologically, well-differentiated nerve fibers, fragments of cerebellar folia, and ganglia with large neurons and glial cells were detected in a loose stroma in sections through the cranial bone and adjacent soft tissue in the rudimentary cranial cavity. Immunohistochemically, single cells within the stroma expressed NeuN, consistent with mature neurons, whereas intracranial ganglion cells and nerves had mild expression of doublecortin. The presence of many immature, and only a few mature, neurons in the rudimentary nerve tissue in this case indicates a failure of physiological brain development and differentiation.
Anencephaly is a malformation that belongs to the group of neural tube defects (NTDs). The neural tube develops from the neural plate. In higher vertebrates, the process of primary neurulation involves folding of the neuroepithelium with fusion in the midline to generate the tube. 6 Currently, there is evidence that neural tube closure is initiated at multiple sites along the craniocaudal axis. 5,13 Anencephaly develops as a consequence of cranial NTDs in which the neural folds remain open and the neuroepithelium protrudes from the developing brain, a condition known as exencephaly. Because the cranial vault does not form over the open region, neuroepithelium comes into contact with amniotic fluid leading to degeneration of neural tissue and the typical appearance of anencephaly in late gestation. 8,19
In humans, NTDs are common malformations. However, this condition seems to be rare in veterinary medicine. Even though existence of anencephaly is mentioned in veterinary textbooks, 10 anencephaly has not been reported in dogs. The aim of this article is twofold: first, to describe the macroscopic and histologic findings of anencephaly in a dog; second, to gauge the maturity of neurons and glial cells in cranial ganglia with immunohistochemistry for doublecortin and NeuN as markers for the migratory and postmitotic stages of neurogenesis, respectively. 17
History and Case Presentation
The anencephalic German Shepherd Dog was delivered dead by caesarean section at the 62nd day of gestation. It was 1 of 6 littermates, of which 5 were macerated at term. Because of the advanced maceration, it was not possible to detect any abnormality in these littermates. The bitch was 2 years old and originated from a large breeding colony. The kennel has maintained dogs for more than 4 generations with no inbreeding during this time. There was no history of previous NTDs or other congenital malformations. All bitches were regularly vaccinated against canine herpes virus and canine parvovirus, and there was no history of any infectious disease or medical treatment of the dam during early gestation.
Pathologic Findings
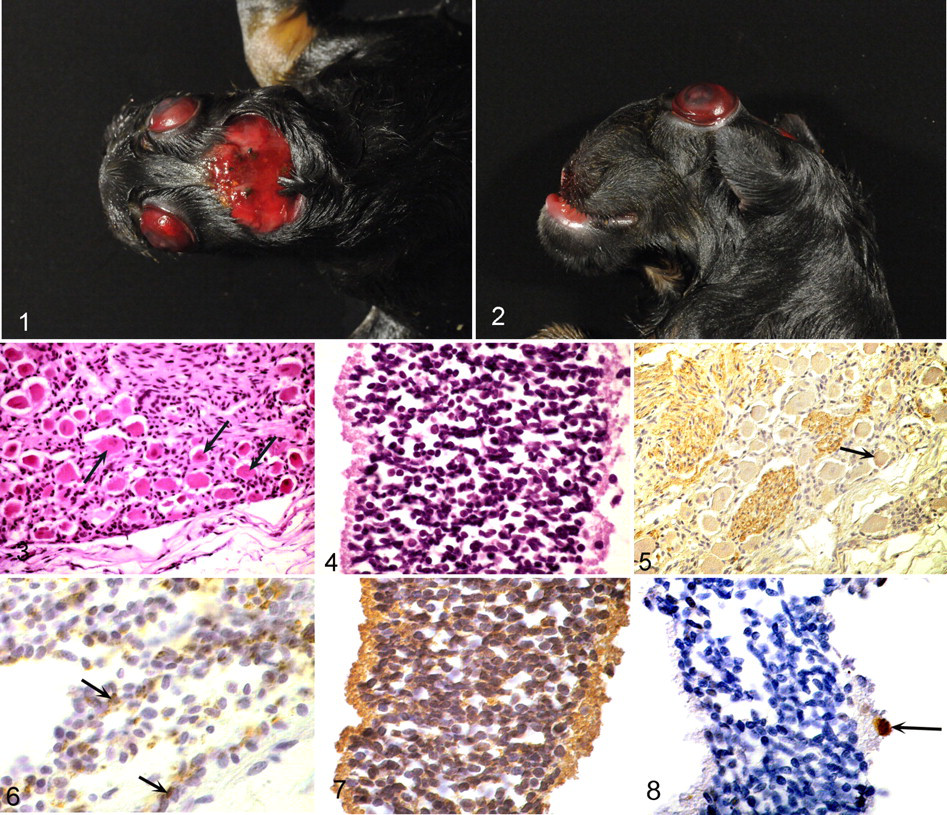
At postmortem examination, the fetus had a flattened skull with a hypoplastic calvarium that had failed to close, leading to an irregular reddish membrane covering the rudimentary cranial cavity (Fig. 1). The base of the skull was thickened and flattened. The orbits were shallow, causing protrusion of the eyes; the jaw was affected by brachygnathia superior (Fig. 2 ). Macroscopically, no brain tissue was detected. Additionally, the left hind limb had an inverse flexion of the tarsus. All other organs were well developed without abnormalities. No bacterial pathogens were cultured from internal organs or placental sites.
The eviscerated fetus was fixed in 10% neutral buffered formalin, and sections of the cranium, vertebral column, and internal organs were routinely processed for histologic examination and immunohistochemistry. Bony tissue was decalcified in an EDTA-containing solution. Histologic sections were stained with hematoxylin and eosin (HE). Immunohistochemistry was performed with primary antisera against neurofilament, neuronal nuclear antigen NeuN, doublecortin (a protein that promotes microtubule polymerization), glial fibrillary acidic protein, neuron cell surface antigen A2B5, S-100 protein (a multigene family of low-molecular-weight calcium-binding proteins), vimentin, and factor VIII–related antigen (Table 1 ). The positive immunohistochemical controls consisted of tissues that express these markers. Consecutive sections were used as negative controls in which a nonreacting mouse monoclonal antibody against chicken lymphocytes (monoclonal antibodies) or normal rabbit or goat serum (polyclonal antibodies) replaced primary antibodies.
Antibodies and Detection Systems Used for Immunohistochemistry a
a NF, neurofilament; mAb, monoclonal antibody; ABC, avidin–biotin complex method (Vector Laboratories, Burlingame, CA); NeuN, neuronal nuclear antigen; pAb, polyclonal antibody; GFAP, glial fibrillary acidic protein; PAP, peroxidase–antiperoxidase method (DakoCytomation, Hamburg, Germany); A2B5, neuron cell surface antigen A2B5; FVIII, FVIII–related antigen.
b Dako / DakoCytomation, Hamburg, Germany; Santa Cruz Biotechnology, Inc, Santa Cruz, California; Chemicon / Millipore, Jaffrey, New Hampshire.
c Sigma, Deisenhofen, Germany.
Histologically, the cranial bone and soft tissue that filled the rudimentary cranial cavity consisted of a loosely arranged stroma with a few cross sections and tangential sections of well-differentiated peripheral nerves, which were distinguished from cranial nerves by their fibrous perineurium. Also observed were a group of small granule neurons overlying a rudimentary molecular layer (interpreted as fragments of cerebellar folia) and large neurons surrounded by glial cells (consistent with cranial ganglia; Fig. 3). Purkinje cells were not found. Both eyes were well formed with no abnormality of the anterior portion; however, the optic nerves were small and could not be traced in their entirety to the optic chiasm. The retina was completely detached and consisted of a uniform layer of small round-to-oval cells that could not be assigned to a specific retinal layer (Fig. 4). Retinal ganglion cells were scarce; the nerve fiber layer was absent. The pituitary gland was not found. The spinal cord consisted of disordered tissue without clear separation of white and gray matter in cervical segments. Lesions within the lumbar segment resembled syringomyelia, characterized by a cleftlike cavity in the inner portion of the cord without ependymal lining. Spinal ganglia were regular in number and well differentiated in all locations.
With immunohistochemistry, neurons in all locations were neurofilament positive. Cranial and spinal nerves had marked doublecortin expression, whereas this marker was only mildly expressed in perikarya of cranial and spinal ganglia (Fig. 5). Single cells within the stroma were also positive for doublecortin. NeuN expression could be found only in individual neurons of rudimentary cerebellar folia (Fig. 6) and spinal cord. Glial cells had diffuse positivity for glial fibrillary acidic protein but were all negative for S-100, vimentin, and A2B5. The stroma that filled the remaining cranial cavity was composed of loosely arranged spindloid cells that expressed vimentin and numerous spaces lined by cells positive for factor VIII–related antigen. Within the retina, the small round-to-oval cells described above all had diffuse cytoplasmic doublecortin expression (Fig. 7). A few retinal ganglion cells faintly expressed NeuN (Fig. 8).
Discussion
Macroscopic absence of the brain, hypoplastic calvarium, thickened base of the skull, 11 and recognizable cranial nerves and fragments of cerebellar folia 2 are characteristic of anencephaly in humans and were found in the canine fetus reported here. During embryogenesis, the eyes evolve from evaginations of the forebrain. 4 Because anencephaly is thought to be secondary to degeneration of previously developed neural tissue from contact with amniotic fluid, 8,19 eye development should have been initiated before the onset of brain degeneration. Consequently, the reduction of retinal ganglion cells, which is also described in anencephalic humans, 1,15 should be interpreted as a sequel of secondary retrograde degeneration. This could be the consequence of a lack of synaptic contact with the dorsal lateral geniculate nucleus, 9 in which case the optic nerves could be interpreted as abiotrophic.
The expression of doublecortin by all neuronal processes and retinal cells, in combination with absence of NeuN expression, indicates neuronal immaturity. 14,16 In contrast, remaining ganglion cells of the retina expressed NeuN—which denotes maturity of these neurons, as described for the adult human retina 18 —and would thus support the assumption of secondary retinal ganglion cell degeneration. The immunohistochemical reactivity of glial cells—which were positive for only glial fibrillary acidic protein and failed to express S-100 protein, A2B5, or vimentin—matches the expression profile of mature astrocytes of adult animals. 3 Faint doublecortin expression was detected in the perikarya of cranial and spinal ganglia with strong immunohistochemical signal in neuronal processes. Because doublecortin is usually detected equally in neuronal perikarya and processes, 7 the doublecortin expression pattern in this case might reflect aberrant neuronal maturation.
The rudimentary cranial cavity was filled with a loosely arranged connective tissue intermingled with vascular spaces, as demonstrated by immunohistochemistry for factor VIII–related antigen. These structures could be interpreted as an equivalent of the area cerebrovasculosa, which is described in many cases of human anencephaly. 2
The development of the neural tube is a multistep process controlled by various genes and environmental factors. Human NTDs are familial disorders without a strict Mendelian pattern of inheritance. Environmental factors that influence neural tube closure comprise chemical toxins, such as organic solvents and pesticides, and hyperthermia. 8,13 Studies of human populations with a high prevalence of NTDs demonstrated a significant reduction of NTDs following periconceptional supplementation of folate. Despite ongoing investigations of the genes involved in folate metabolism, no definitive role of these genes in the pathogenesis of NTDs has been found. 12 Because anencephaly has not been reported in dogs and no NTDs had been documented in the kennel until this case, we speculate that the NTD leading to anencephaly in this dog represents an incidental environmental influence during early gestation.
Footnotes
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Financial Disclosure/Funding
The authors declared that they received no financial support for their research and/or authorship of this article.