
Abstract
Over the period of a year, colitis was observed in 44 mice raised in a conventional nonspecific pathogen–free colony, 41 of these having concomitant retrovirus–induced murine acquired immunodeficiency syndrome (MAIDS). The lesions varied from bacterial colonization to hyperplasia of colonic mucosa to severe, often fatal, ulceration. Citrobacter rodentium was isolated from the colon and/or liver of 2 mice with colitis. When C57BL/6 mice with or without MAIDS were given graded doses of the bacterium, only those with MAIDS developed colitis, and C rodentium was reisolated from their livers. Thus, mice with MAIDS can develop severe disease following opportunistic infection with an environmental contaminant of the colony that is nonpathogenic for normal adult mice.
Murine acquired immunodeficiency syndrome (MAIDS) is a disease of lymphoproliferation and severe immunodeficiency induced in sensitive strains of mice by the replication–defective component (BM5 def) of the retrovirus mixture designated LP-BM5 murine leukemia virus (MuLV). 14 Whereas MAIDS has some strong resemblances to human AIDS, 21 a feature lacking is morbidity associated with infection by adventitious infectious agents (eg, Pneumocystis jirovecii), which complicates the human disease. Thus, most mice with MAIDS, although profoundly compromised immunologically, appear to die from indirect effects of lymphadenopathy (possibly airway obstruction by enlarged thoracic lymph nodes) or from extensive lymphoid infiltration and associated dysfunction of liver or kidney. Mice with MAIDS do show enhanced susceptibility to parenterally administered pathogens to which normal mice are resistant, including ectromelia, 7 Cryptosporidia, 9 Toxoplasma gondii, 12 Listeria monocytogenes, 16 Leishmania major, 10,11 and mouse cytomegalovirus. 24 Reasons for the lack of opportunistic infection may be (1) a different spectrum of susceptibility to infectious agents in MAIDS as compared with AIDS, (2) a failure to detect diseases caused by such agents, or (3) absence of such agents in the environment. Recently, however, we observed disease and death from ulcerative colitis in mice with overt MAIDS.
A naturally occurring disease of mice, transmissible murine colonic hyperplasia (TMCH), is caused by Citrobacter rodentium, formerly known as Citrobacter freundii biotype 4280. 2,15,25 Features of TMCH include rectal prolapse, diarrhea, and thickened, dilated colon, as well as a histologic picture of epithelial cell hyperplasia and crypt elongation. The severity of lesions varies with mouse strain and age, C57BL/6 being relatively resistant, and most infections resolve spontaneously. 15,19 This report describes the colonic lesions seen in MAIDS mice and their etiologic association with infection by C rodentium.
Materials and Methods
Mice, Viruses, and Bacterial Infections
C57BL/6J (B6), CXBK/ByJ (CXBK), P/J (P), and C57L/J (C57L) mice purchased from the Jackson Laboratory (Bar Harbor, Maine) were maintained in a conventional animal facility. Serologic screenings of mice in the colony identified rare positive tests for GDVII and mouse hepatitis virus (MHV). These serologic findings were not associated with clinical signs of neurologic disease or diarrhea, poor growth, and death of infant mice characteristic of virulent GDVII or MHV infections, respectively. Examination results for parasites including pinworms were routinely negative.
B6, CXBK, and P mice were infected at 6 to 8 weeks of age with the LP-BM5 mixture of MuLV that causes MAIDS as described previously. 14 MAIDS was induced in 6–week–old C57L mice by infection with the BM5 defective virus pseudotyped by amphotropic MuLV. 20 Mice were monitored several times per week for lymphadenopathy, splenomegaly, labored breathing, or other signs of morbidity. Moribund mice were necropsied, and selected tissues were fixed in neutral buffered formalin for later preparation of paraffin–embedded sections to be stained with hematoxylin and eosin. For bacterial infections, mice were administered 1 ml of saline containing high or low numbers of viruses by gavage using an intubation needle. All procedures were carried out in compliance with protocols approved by the Institutional Animal Care and Use Committee of the National Institute of Allergy and Infectious Diseases.
Results
Spontaneous Colitis in Mice With MAIDS
A total of 44 histopathologically confirmed cases of colitis appeared sporadically during a 1–year period in mice housed in a conventional animal facility. In all but three cases, the mice had been inoculated with BM5def MuLVs 5 to 16 weeks earlier and had developed lymphadenopathy and splenomegaly typical of moderate to advanced MAIDS. The exceptions were otherwise normal uninoculated mice, 2 C57L and 1 B6. Approximately one third of the cases were severe, the mice being thin, with ruffled fur, some degree of diarrhea, and, often, blood in the perianal region. These signs were not detected (1) in a number of other strains maintained in the same colony that were genetically resistant to MAIDS and did not develop disease after infection or (2) in strains of various genetic backgrounds that were MAIDS susceptible and developed typical disease after infection.
Histologically, disease was restricted to the colon except in 3 cases of severe colitis that additionally had segmental hepatic necrosis apparently associated with thrombosis. The degree of colonic disease varied, the earliest stage amounting only to colonization with what appeared to be a single type of bacterium adherent to the membrane of surface epithelial cells. More severely affected mice showed extensive bacterial growth associated with mucosal hyperplasia, the most advanced cases having ulceration and extensive inflammation. Except for the early stage of surface bacterial colonization, the range of lesions seen in spontaneous cases was duplicated in the experimental study described below.
Isolation of C rodentium
Isolations on blood and McConkey agar were made from the colons and livers of 2 mice with advanced MAIDS and colitis. Both mice had severe colitis that was grossly discernible, the first case also having segmental liver necrosis. Pure colonies developed within 24 hours, and both isolations were the same atypical strain of Citrobacter, referred to as isolate 4280 by Barthold 3 and now designated C rodentium, as determined by nonmotility and established fermentation characteristics, 2 particularly the inability to utilize citrate. One of the liver isolates was used for transmission studies.
Experimental Induction of Colitis by Infection With C rodentrium in Mice With MAIDS
Adult B6 mice were either uninoculated or infected 13 weeks previously with LP-BM5 MuLV and had developed MAIDS-related splenomegaly and lymphadenopathy. These mice were divided into groups of 2 or 3 for administration of a high or low dose of bacteria by gavage. As shown in Table 1, only mice with MAIDS developed colitis, detected 11 to 15 days postinfection or at the termination of the experiment at 17 days. No gross or microscopic evidence of colonic disease was found in control B6 mice inoculated with C rodentium that were not previously infected with the retrovirus mixture, although cultures of liver from 2 mice yielded 1 or 2 colonies. Among mice with MAIDS, 1 that had received 2.5 × 106 organisms died at 11 days postinfection with severe colitis, and large numbers of bacteria were isolated from the liver. Two additional mice in this group developed severe illness and were found to have ulcerative colitis on necropsy. Mice that received the lower dose survived until termination of the experiment at day 17 with definite but considerably less severe colitis.
Induction of Colitis in C57BL/6 Mice With Murine Acquired Immunodeficiency Syndrome by Administration of Citrobacter rodentium
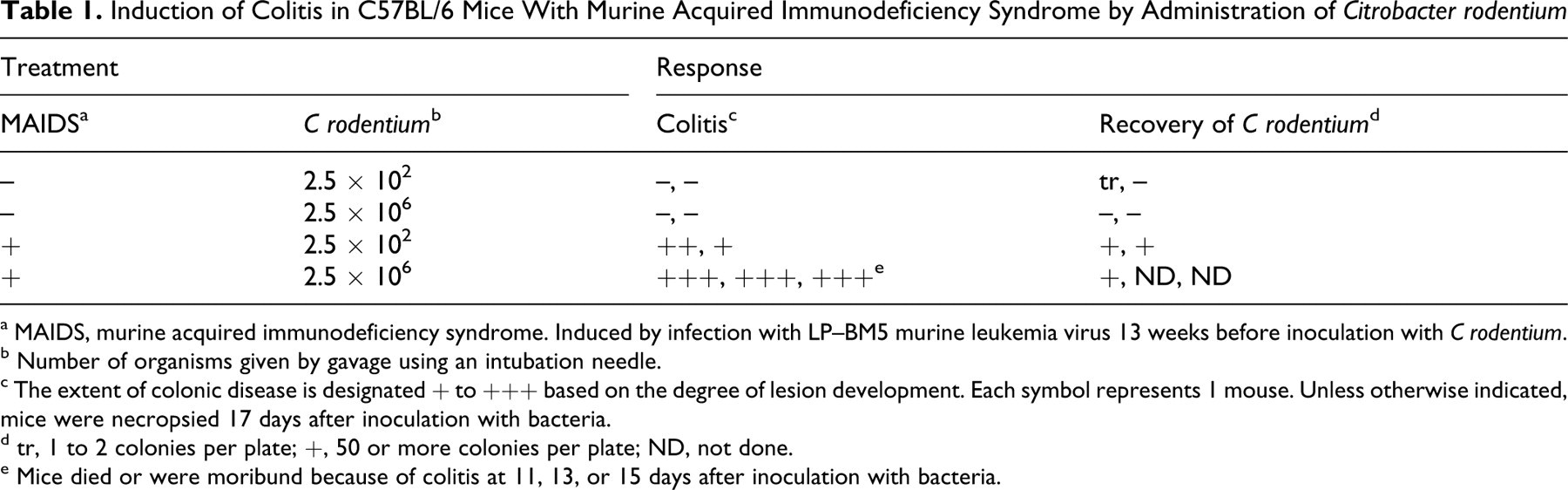
a MAIDS, murine acquired immunodeficiency syndrome. Induced by infection with LP–BM5 murine leukemia virus 13 weeks before inoculation with C rodentium.
b Number of organisms given by gavage using an intubation needle.
c The extent of colonic disease is designated + to +++ based on the degree of lesion development. Each symbol represents 1 mouse. Unless otherwise indicated, mice were necropsied 17 days after inoculation with bacteria.
d tr, 1 to 2 colonies per plate; +, 50 or more colonies per plate; ND, not done.
e Mice died or were moribund because of colitis at 11, 13, or 15 days after inoculation with bacteria.
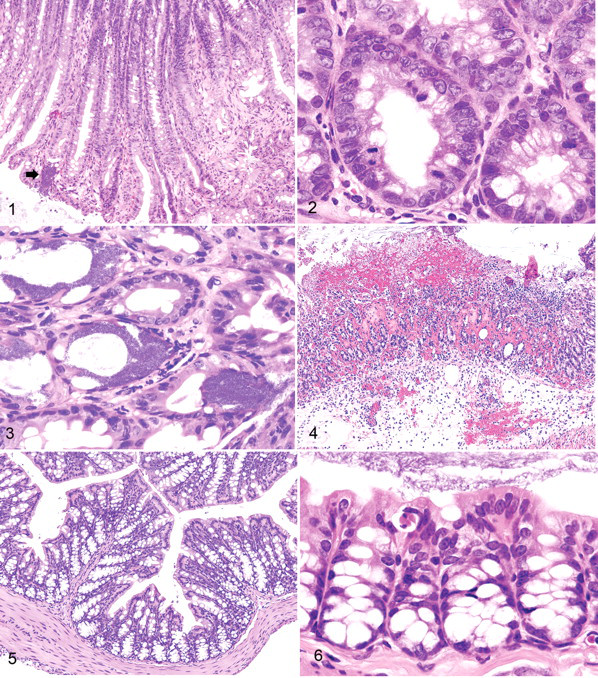
Three degrees of severity of colonic lesions were observed in mice inoculated with C rodentium. The least severe type was associated with mucosal hyperplasia with increased length of colonic crypts (Fig. 1 ) and hyperplasia of crypt epithelium (Fig. 2). Bacterial colonies were numerous but had not penetrated into the pits of the colonic crypts, and inflammation was minimal (Fig. 3). The next stage of severity was marked by the development of small circular ulcers characterized by the presence of large numbers of bacteria, hemorrhage, and focal destruction of colonic mucosa. The most severe lesion was ulceration over much of the surface of the colon accompanied by extensive edema and neutrophilic infiltration (Fig. 4). Mice without MAIDS showed no bacterial growth or colitis (Figs. 5, 6).
Mice developing disease following naturally acquired infection were successfully treated by adding tetracycline hydrochloride to the drinking water as reported for C rodentium infection. 1,26
Discussion
Descriptions of spontaneous severe C rodentium–associated colitis with significant mortality in adult mice are relatively rare, the majority of reports being of self–limited hyperproliferative responses of the colonic epithelium (reviewed in Holcombe and Schauer 15 ) that have been associated with activation of NF-κB and alterations in CKIϵ that influence β-catenin signaling. 29,32 Genetic background affects severity and outcome of natural and experimental infection, with C3H/HeJ mice being the most sensitive adult strain examined. 4,5,30 The B6 mouse appears to be relatively resistant to any effect of C rodentium infection. 4,6 The sensitivity of C57L, CXBK, and P mice that developed severe colitis in association with MAIDS was not assessed. As shown by experimental infection, concomitant MAIDS clearly reduces the resistance of adult B6 mice to C rodentium, allowing development of sometimes fatal ulcerative colitis. In line with previous findings, 3 severe colitis was associated with septicemia, as indicated by isolation of bacteria from livers of 3 mice exhibiting segmental hepatic necrosis; however, with these exceptions, no lesions were seen in organs other than the colon. In spontaneous cases, we observed the same range of colonic response previously described for C rodentium infection in suckling mice, 1,3 from moderate growth of bacterial colonies associated with mucosal hyperplasia to ulceration of the colonic mucosa. In experimentally induced disease, we also noted a gradation in response that appeared to be dose related (Table 1).
MAIDS is a disease of profound immunodeficiency, with impairment in function and phenotype of both T and B cells, including responsiveness to mitogens and antigen, decreased responsiveness of B cells to T helper cell function, and progressive activation of B cells followed by anergy. Although mice with MAIDS show increased susceptibility to experimental infection by a number of bacterial, viral, and parasitic agents, 7,11,21 the syndrome has not been identified as a predisposing factor for opportunistic infections causing overt disease, possibly because laboratory mice live in an environment comparatively free of continual challenge from potentially pathogenic microorganisms. Our observations regarding C rodentium indicate that when such an organism does occur in the environment of mice with MAIDS, it can induce fatal disease, even though such infection would be inconsequential in normal adult mice.
The mechanisms responsible for this phenotype remain to be determined, but several published observations on mice with MAIDS are likely to be quite relevant. First, mice infected for 1 week were shown to express high levels of IL-2, IL-4, IL-6, and IL-10 as well as IFN-γ, the cytokine profile of activated Th0 CD4+ T cells, but only IL-4 and IL-10 continued to be expressed at high levels after week 2, indicating a switch to a dominant Th2 population. 22 Both cytokines act to restrict the development of Th1 CD4 T cells. Thus, Th1 cytokines shown to contribute to the control of C rodentium—IL-2, IL-6, and IFN-γ—were expressed only transiently, whereas IL-10, a repressor of Th1 cells as well as more recently described Th17 cells that contribute to the control of C rodentium, 8,28 was expressed throughout the course of disease. Second, CD4+ T cells in mice with MAIDS become progressively more anergic during the course of disease 17 and thus increasingly impaired in their ability to generate adaptive immune responses to the pathogen. Although the etiologic significance of bacterial burden in sensitivity to disease manifestations in different mouse strains is not wholly clear 6,13,23 and may relate in susceptible hosts to the degree of colonic crypt infection, 30 replication of colonic C rodentium in mice with MAIDS was markedly enhanced (Table 1). Nonetheless, why this unique combination of immunologic abnormalities is manifested as susceptibility to lethal infection with C rodentium remains to be fully understood.
One additional point for consideration is whether the abnormal lymphoid populations of mice with MAIDS might contribute directly to the pathology that we found associated with infection by C rodentium. This possibility is suggested by the finding that adoptive transfer of spleen cells from mice with MAIDS to congenitally athymic “nude” mice resulted in death within 6 weeks owing to cachexia with diarrhea and anal bleeding. 27 Histologic studies revealed lymphocytic infiltration and swelling associated with erosions of the colonic mucosa and bleeding. The mice were maintained specific pathogen free, and obvious infection was not detected, so contributions from C rodentium would be most unlikely. This colonic phenotype is not unlike that of SCID (severe combined immunodeficiency) recipients of naive CD4+ T cells, which could be reversed by the addition of CD4+CD25+– subset of regulatory T cells, 31 termed Tregs, which are capable of suppressing autoimmune and inflammatory conditions. 18 It may be that the pathology seen in mice infected with C rodentium reflects the direct effects of the organism in an environment characterized by immunodeficiency associated with the presence of autoreactive T cells unconstrained by Tregs.
Footnotes
Acknowledgements
Dr Thomas C. Moore, previously of the Laboratory Sciences Section, Veterinary Resources Program, National Institutes of Health, identified the bacterial isolates and provided the culture used for inoculation. We thank Dr Jerrold Ward for assistance with photography and National Institute of Allergy and Infectious Diseases intramural editor Brenda Rae Marshall for assistance.
Because the authors are government employees and this is a government work, the work is in the public domain in the United States. Notwithstanding any other agreements, the National Institutes of Health reserves the right to provide the work to PubMedCentral for display and use by the public, and PubMedCentral may tag or modify the work consistent with its customary practices. You can establish rights outside of the United States subject to a government use license.
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
This work was supported by the Intramural Research Program of the Division of Intramural Research, National Institute of Allergy and Infectious Diseases, National Institutes of Health.