
Abstract
Background
Chronic restraint stress (CRS) is widely used to recapitulate depression phenotypes in rodents but is frequently criticized for a perceived lack of efficacy. The aim of this study was to evaluate anhedonic-like behavior in the CRS model in rodents by performing a meta-analysis of studies that included sucrose preference tests.
Methods
This meta-analysis was conducted following the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) recommendations. We comprehensively searched for eligible studies published before June 2021 in the PubMed, Embase, Medline, and Web of Science databases. We chose sucrose preference ratio as the indicative measure of anhedonia because it is a core symptom of depression in humans.
Results
Our pooled analysis included 34 articles with 57 studies and seven rodent species/strains and demonstrated decreased sucrose preference in the stress group compared with controls. The duration of CRS differentially affected the validity of anhedonic-like behavior in the models. Rats exhibited greater susceptibility to restraint stress than mice, demonstrating inter-species variability.
Conclusions
Our meta-analysis of studies that used the CRS paradigm to evaluate anhedonic-like behavior in rodents was focused on a core symptom of depression (anhedonia) as the main endpoint of the model and identified species-dependent susceptibility to restraint stress.
Introduction
Depression is currently among the top five leading causes of the global disease burden, affecting 20% of the world’s population.1,2 According to the World Health Organization, over 300 million people suffer from major depressive disorder (MDD) worldwide. 3 Depression is a mood disorder characterized by a depressed mood, social isolation, anhedonia, and feelings of worthlessness that negatively influence overall quality of life, sometimes even causing patients to endanger their lives through recurrent suicidal thoughts.4,5 Depression represents a chronic and recurrent psychiatric condition with varying symptoms among patients. Patients with chronic diseases have a higher risk of depression, which in turn reduces recovery from chronic diseases and treatment compliance. Depression not only imposes a large healthcare and economic challenge on society but also presents considerable social impacts. MDD is now the main risk factor for suicide-related deaths and the second leading cause of disability worldwide. 5 Unfortunately, 30% to 50% of patients suffering from depression do not respond to current antidepressant treatments. 6 Stress, or psychological stress, is a reaction mode. When the human body is stimulated by external adverse factors it will trigger stress reactions (anxiety, depression, fear, and other adverse emotions). Chronic stress, also called long-term stress, means that the stress process and event that cause stress will last longer. 7 It has been recognized that physiological responses to chronic stress are potent modulators of immune, endocrine, and metabolic pathways. 8 Chronic stress is a significant risk factor for the development of depression, which leads to synaptic changes and depressive-like behaviors in rodents. Currently, chronic stress models are the most widely used animal models of depression. 9
It is difficult to determine what the underlying mechanisms of MDD might be in human studies. In contrast, animal studies allow the experimental induction of depression-relevant behaviors, which permits deeper investigations into molecular pathways. Thus, modeling depression in animals is vital for uncovering mechanisms underlying the human condition. Great progress has been made over the past 50 years in elucidating the pathophysiology of depression, much of which is attributable to the implementation of numerous animal models of depression.2,10,11 Most of the current knowledge about the mechanisms underlying depression has come from animal models, although no animal model can be entirely congruent with the human condition. Chronic psychosocial stressors are risk factors for the development of depression in humans.12,13 Chronic stressors are detrimental because they disrupt the normal stress response of the brain, eventually contributing to the development of depression.14–16 Additionally, chronic stressors enhance levels of stress-related hormones by disrupting the hypothalamic-pituitary-adrenal (HPA) axis and suppress the production of new neurons in the hippocampus.16–18
Several chronic stress models including chronic social defeat stress (CSDS), chronic restraint stress (CRS), and chronic unpredictable mild stress (CUMS) have been shown to recapitulate depression-like behaviors in rodents, and thus have been used to model depression and investigate its underlying mechanisms. Depression-like behaviors induced by the animal models have been examined including by the sucrose preference test (SPT; indicative of anhedonia) and forced swim and tail suspension test (indicative of despair). Changes in the performance of model animals in these tests can often be reversed by chronic antidepressant treatments. 19 However, it is noteworthy that stress designs in the model contribute to stress susceptibility. Anhedonia is a decreased ability to experience pleasure that is recognized as a core symptom of human depression. SPT is widely applied as a behavioral measure of anhedonia. 20 Experimental animals are given a free choice between drinking water or a weak sucrose solution (1%–2% [weight/volume] sucrose) 8 and exhibit a preference for the latter, reflecting the hedonic state of rodents.
The CRS model is a convenient, inexpensive, and stable rodent model of chronic stress because of its relative simplicity and easy workflow; therefore, it is widely used to establish depression rodent models. 2 Previous publications have used many strains of rats and mice to establish the CRS model. Additionally, the restraint duration, intensity, and other conditions have been varied across different studies. Some studies have reported that exposure to CRS induced anhedonia in rodents on the basis of decreased sucrose preference, a core symptom of human depression.21,22 In contrast, a conflicting study reported that CRS failed to induce anhedonia-like behaviors. 23 Thus, it remains unclear whether CRS can be used as a valid animal model of depression that recapitulates anhedonia-like behavior in different rodent species and strains.
Systematic reviews and meta-analyses, as standard practices in clinical research, have been increasingly performed to validate preclinical studies of disease etiology, diagnosis, and prognosis. In terms of animal experiments, it has been estimated that approximately 50% of published results are not reproducible, which has been described as a “replication crisis”. 2 However, few pooled analyses have been conducted within basic life science research to evaluate the reliability of results. The aim of this study was to evaluate the anhedonic-like behavior induced by the CRS model in rodents by performing a meta-analysis of studies that reported SPT results.
Methods and materials
Search strategy
The meta-analysis was conducted following the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) recommendations. We comprehensively searched for eligible studies published before June 2021 in the PubMed, Embase, Medline, and Web of Science databases. We searched for the following keywords and corresponding terms in titles and/or abstracts: “chronic restraint stress” OR “chronic psychological stress” AND “animal model”.
Eligibility criteria and study selection
All studies enrolled in this meta-analysis satisfied the following criteria: (1) published in English; (2) reported as original research; (3) reported the implementation of CRS protocols in rodents (mice or rats) for at least 1 week; (4) examined depressive-like behaviors including SPT (calculated according to the following formula: % sucrose preference = [sucrose intake/total fluid intake] × 100); (5) provided SPT outcomes (%) in the text, figures, and/or graphs; and (6) used normal (wild-type) experimental animals that were housed in a suitable environment. Studies were excluded from the meta-analysis if they did not meet all of the above criteria. The selection of included studies was conducted independently by two authors (YM and YX). Discrepancies between the two authors were solved in face-to-face conferences with the third author (XY).
Data extraction
Two authors (YM and YX) independently extracted data from the included studies and any disagreements were settled in face-to-face consultations with the third author (XY). The authors summarized the main characteristics of the studies and collected all information regarding CRS design and SPT protocols. The following information was directly extracted from the selected studies: name of first author, publication year, model animal features (sex, strains), CRS model design (restraint stress duration, period/day), examined depression-like behaviors, measurement of water and food consumption, measurement of body weight, determination of corticosterone and catecholamine, details of SPT (test onset time, training protocols, water and food deprivation period, sucrose concentrations, testing period), and sample sizes (n) of the experimental and control groups. For the pooled analysis, SPT outcomes, including mean and standard error (SE), standard deviation (SD), or standard error of mean (SEM), were directly extracted from graphs or figures using Engauge Digitizer software.
Statistical analysis
We evaluated the efficacy and stability of the CRS protocol in modeling depressive-like behavior according on SPT results in model animals. Standardized mean differences (SMDs) with 95% confidence intervals were defined as the indicator of efficacy, and the meta-analysis was performed by pooling mean sucrose preference (%) results, SD/SEM/SE of the mean, and sample size 2 using Stata software version 11.1 (STATA Corporation, College Station, TX, USA). SMD is a measure of effect size that reflects the degree of outcomes in the experimental (stressed) group differing from that of the controls (calculated according to the following formula: SMD = (M1–M2) ÷ SD, where M1–M2 is the difference in the means of the two groups, and SD is the pooled and weighted standard deviation). 2 A fixed-effect model was adopted in the pooled analysis. Results of the meta-analysis are displayed as forest plots.
The Higgins I2 statistic was used to estimate the heterogeneity among the enrolled studies. This statistic represents the percentage of variation between studies ranging from 0% to 100%. A P value ≤0.1 or I2 ≥50% indicates substantial statistical heterogeneity between studies. Publication bias was assessed using a funnel plot (a visual aid for detecting bias). The effect measure (log|SMD|) versus its precision (SE of log|SMD|) was plotted in the funnel plot. In cases of absence of publication bias, the data are expected to be distributed in a funnel-shaped area in the plot.
Results
Literature search and study selection
The flowchart for identifying eligible articles for the meta-analysis is shown in Figure 1. The initial literature search in the PubMed, Embase, Medline, and Web of Science databases yielded a total of 2217 distinct articles. Subsequently, 1761 articles were excluded on the basis of their abstracts, and the full-texts of the remaining 456 articles were reviewed. Ultimately, 33 articles were selected. One article was identified by manually checking reference lists, and therefore a total of 34 articles20–22,24 –54 were enrolled in this meta-analysis.
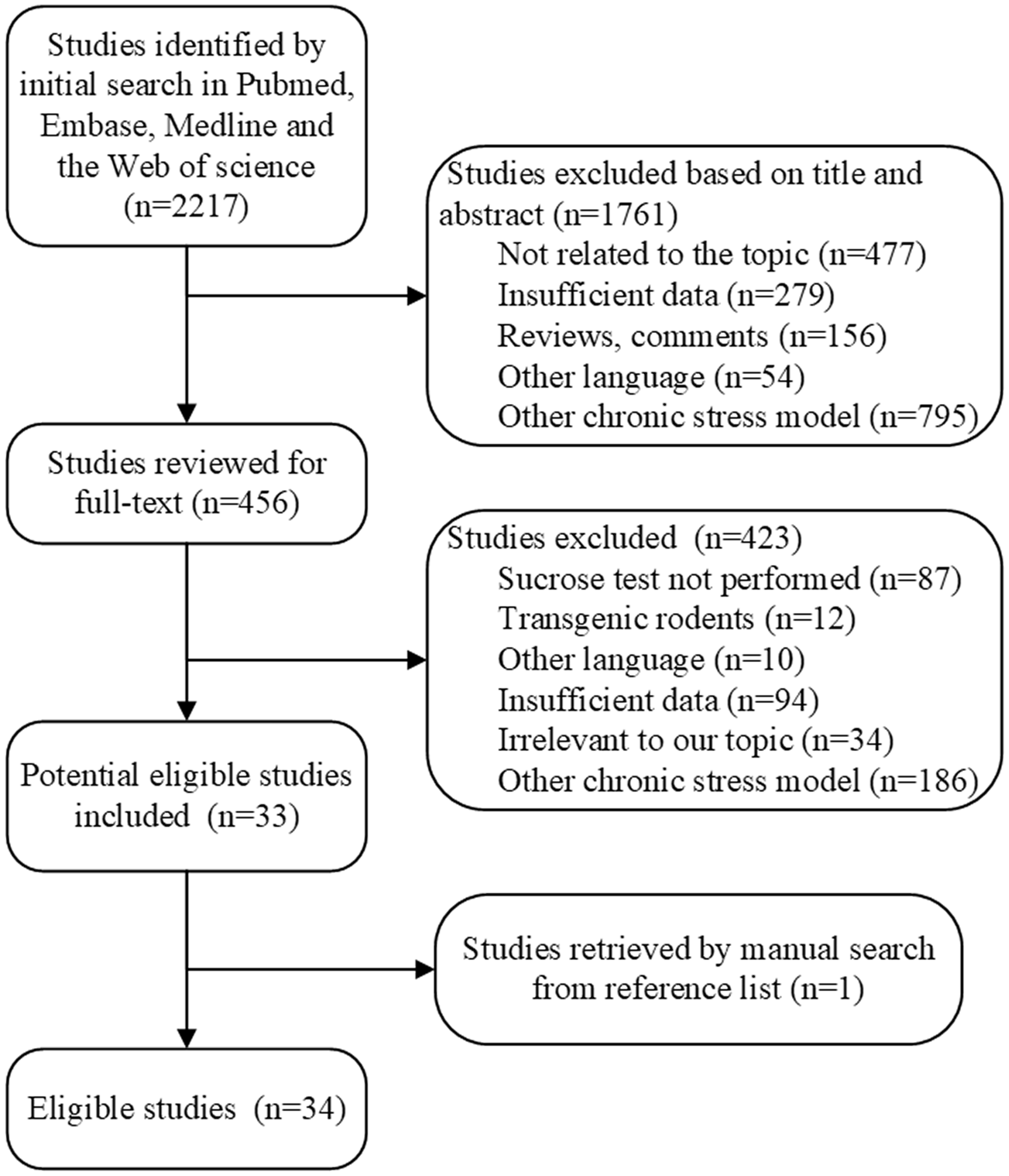
Flow chart of selection process for eligible studies.
Study characteristics
The pooled analysis involved 57 studies in the 34 enrolled publications according to different CRS model designs and included seven rodent species/strains (i.e., Sprague–Dawley (SD) and Wistar rats and Kunming, C57BL/6J, ICR, athymic nude, and BALB/c mice). An overwhelming majority of the studies established the CRS-induced depression model in male rodents, while only 3.5% of the studies (2/57) selected female rodents as the research subjects. Almost all studies successfully modeled depression by CRS on the basis of SPT results; however, different CRS designs (e.g., duration and intensity) and SPT protocols (e.g., test onset time, training protocols, water and food deprivation period, sucrose concentrations, and testing period) were used in the included studies. Rodent characteristics and details of CRS designs are summarized in Table 1; details of SPT protocols are summarized in Table 2.
Primary characteristics of the included studies.
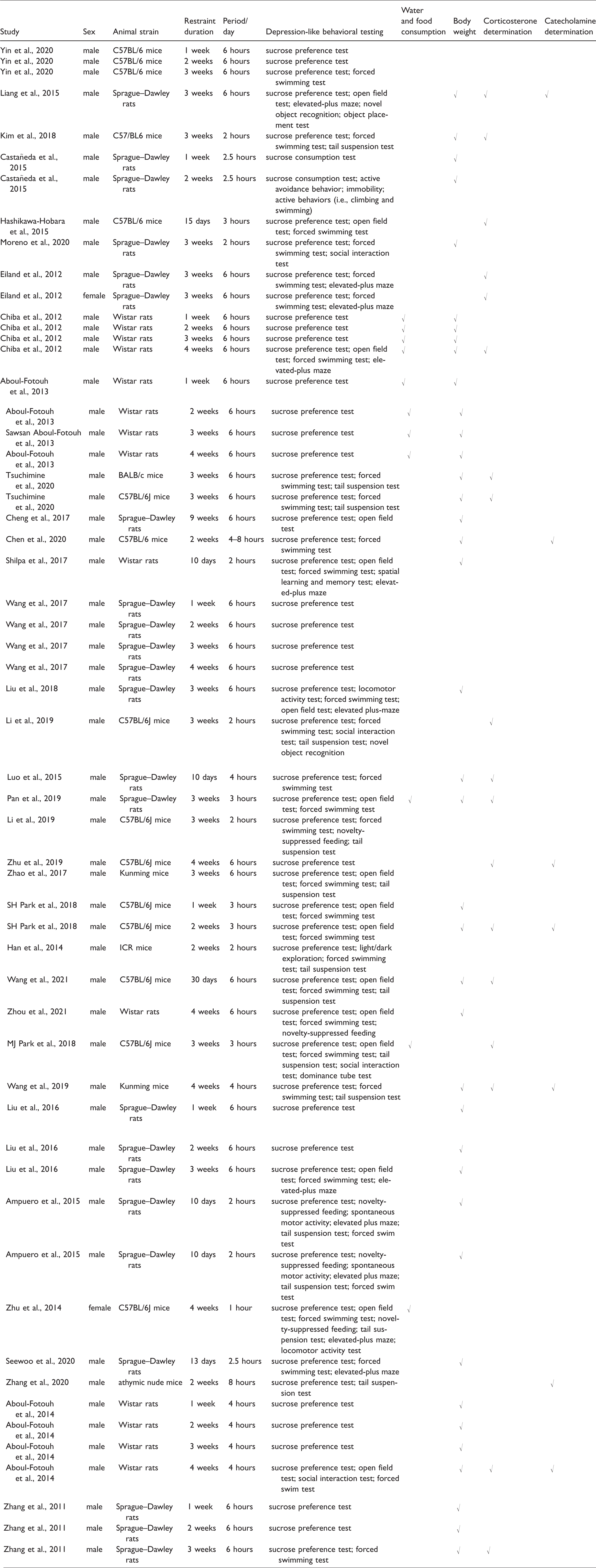
Summary of the sucrose preference test protocols used in included studies.

NA: not available in the article.
The validity of using CRS to model depression
Pooled analyses were performed based on the availability of mean, SE, SD, or SEM, and sample size (n) data for each stress and control group. SPT results were directly extracted from graphs or figures using a digitizing software and are shown in Table 3.
Summary of the sucrose preference test results from studies included in the pooled analysis

*Mean indicates the results of sucrose preference tests, which were calculated according to the following formula: % sucrose preference = [sucrose intake ÷ total fluid intake] × 100.
**SD: standard deviation; SE: standard error; SEM: standard error of mean; in the included studies, the usage of SD, SE, and SEM was not consistent, and they were used in different studies.
#Only the indicated article provided the direct results of sucrose preference tests (%); the results of other studies were extracted directly from graphs or figures using Engauge Digitizer software. Regarding the number of control and stressed animals, narrow ranges of values were provided in several articles, and in these cases, the numbers were defined as the medians of the ranges for the pooled analysis.
The pooled analysis of SPT results from the included studies indicated a significant induction of anhedonic-like behavior in CRS model groups of C57BL/6J mice (Figure 2), SD rats (Figure 3), Wistar rats (Figure 4), Kunming mice, ICR mice, athymic nude mice, and BALB/c mice (Figure 5). Further analysis indicated substantial statistical heterogeneity between studies. These results are summarized in Table 4.

Forest plots of standardized mean difference (SMD) of sucrose preference (%) in C57BL/6J mice following exposure to chronic restraint stress. The effect size was determined by calculating the SMD combined with their 95% confidence intervals. Diamonds indicate SMD values, and the horizontal lines represent 95% confidence intervals.
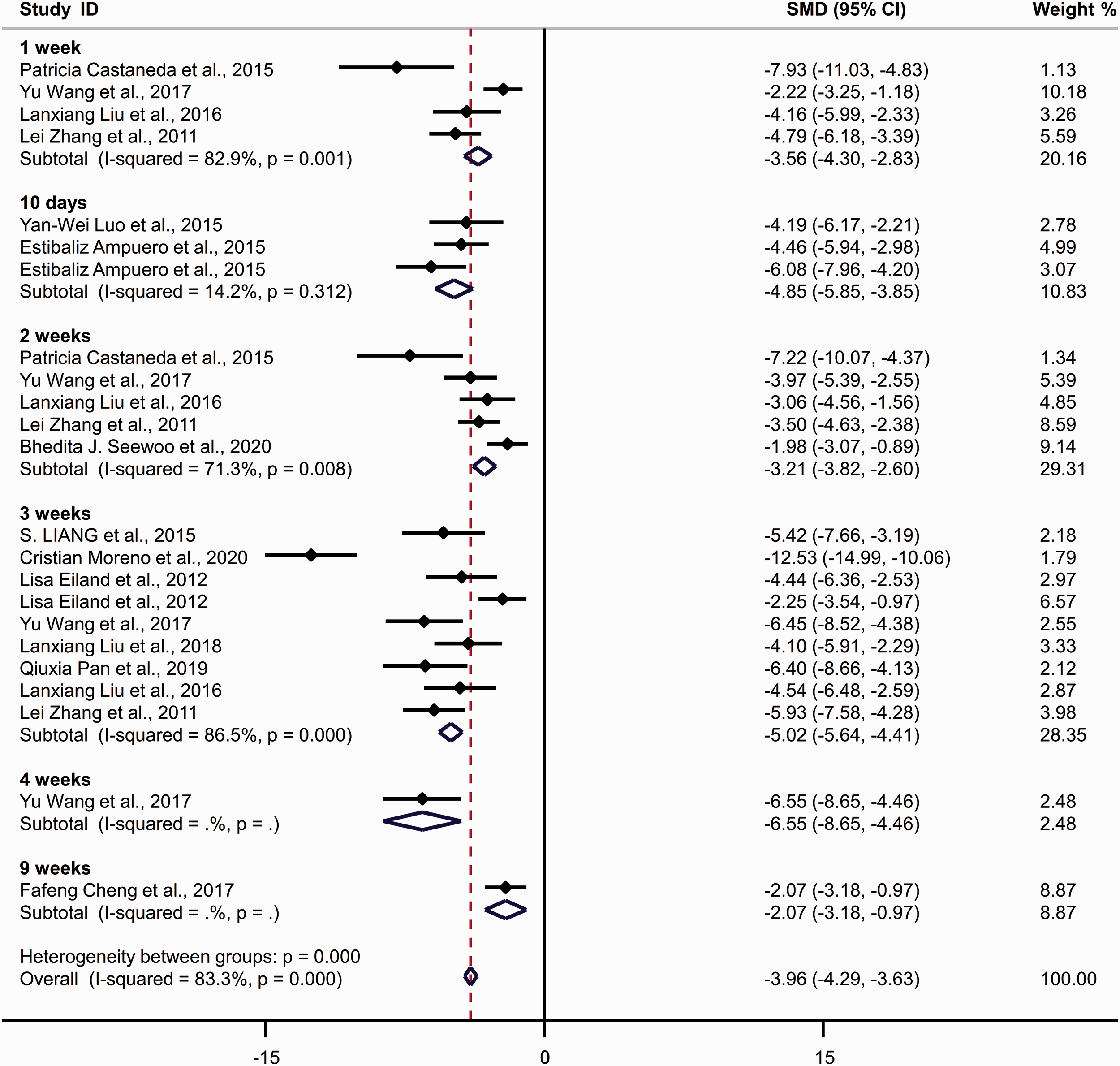
Forest plots of standardized mean difference (SMD) of sucrose preference (%) in Sprague–Dawley rats following exposure to chronic restraint stress. The effect size was determined by calculating the SMD combined with their 95% confidence intervals. Diamonds indicate SMD values, and the horizontal lines represent 95% confidence intervals.
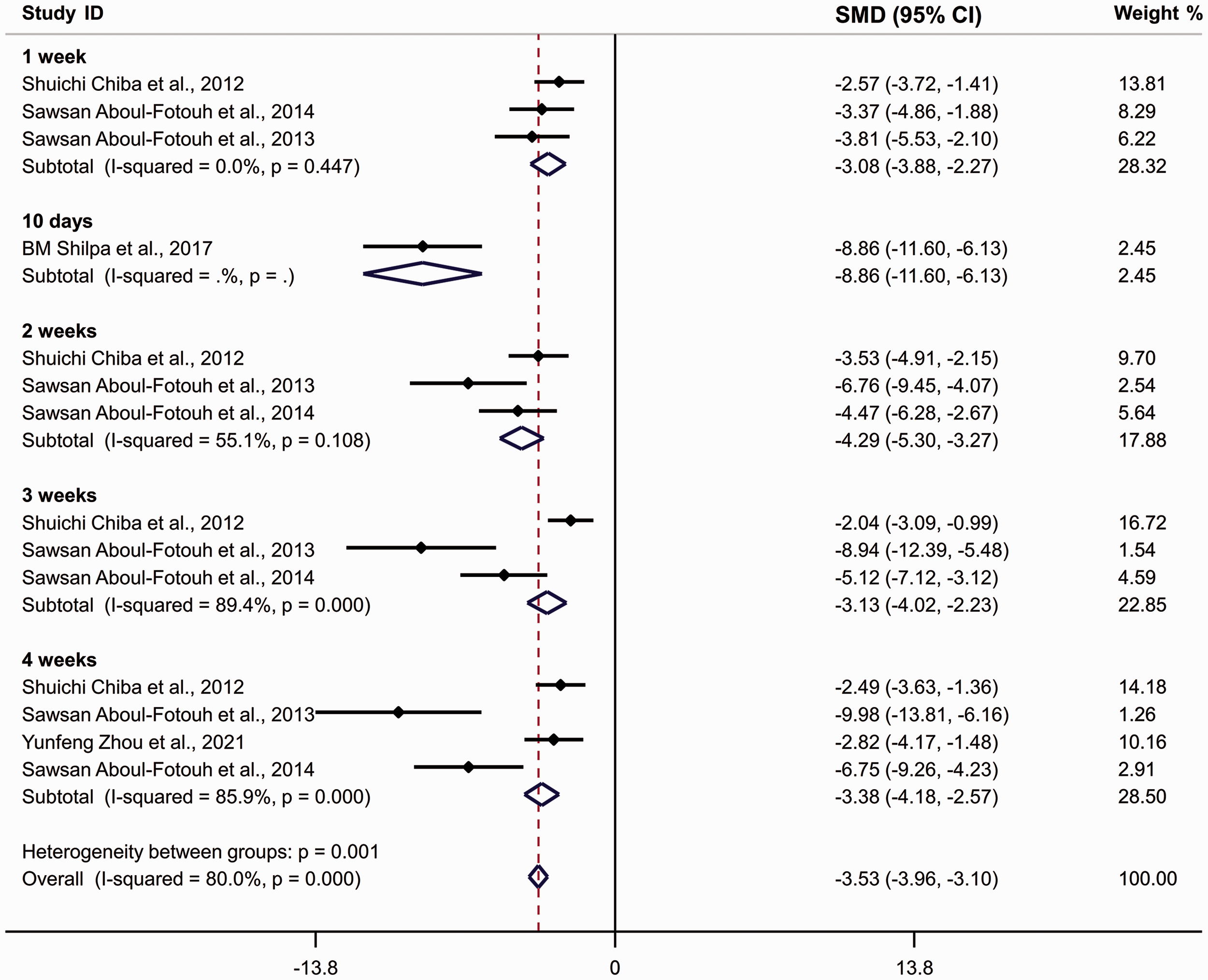
Forest plots of standardized mean difference (SMD) of sucrose preference (%) in Wistar rats following exposure to chronic restraint stress. The effect size was determined by calculating the SMD combined with their 95% confidence intervals. Diamonds indicate SMD values, and the horizontal lines represent 95% confidence intervals.

Forest plots of standardized mean difference (SMD) of sucrose preference (%) in mice of other strains following exposure to chronic restraint stress. The effect size was determined by calculating the SMD combined with their 95% confidence intervals. Diamonds indicate SMD values, and the horizontal lines represent 95% confidence intervals.
Meta-analyses of sucrose preference tests and heterogeneity assessments.

SMD: standardized mean difference.
The pooled analysis of SPT results demonstrated a stronger effect of restraint stress on rats than mice. Notably, the pooled analysis showed that SD rats (SMD = −3.956 [−4.286, −3.626], p < 0.001, I2 = 83.8%) and Wistar rats (SMD = −3.531 [−3.960, −3.102], p < 0.001, I2 = 80.0%) exhibited greater susceptibility to restraint stress than C57BL/6J mice (SMD = −2.80 [−3.221, −2.380], p < 0.001, I2 = 90.4%). Furthermore, the total effect size in SD rats was higher than in Wistar rats.
Additionally, the meta-analysis demonstrated differential sensitivity to restraint stress of varied durations. In C57BL/6J mice, the total effect size indicated the instability and invalidity in the induction of anhedonic-like behavior after 1 week of CRS exposure (SMD = −0.954 [−2.037, 0.128], p = 0.084, I2 = 97.1%). A longer CRS exposure protocol resulted in a sufficient effect size, with a higher SMD value found after 3 weeks (SMD = −3.389 [−4.122, −2.655], p < 0.001, I2 = 88.80%) than after 2 weeks (SMD = −2.396 [−3.196, −1.597], p < 0.001, I2 = 91.7%). Four weeks of CRS exposure (SMD =−3.613 [−4.467, −2.759], p < 0.001, I2 = 84.5%) resulted in a stronger behavioral effect than 3 weeks of CRS exposure. In SD and Wistar rats, only 1 week of exposure successfully recapitulated the anhedonic-like behavior according to the SPT results. Interestingly, a 10-day CRS protocol resulted in stronger behavioral effects than a 2-week protocol in SD rats. Similarly, the effect of 2-week CRS exposure was stronger than 3-week exposure in Wistar rats.
Regarding heterogeneity tests, the single group heterogeneity was low for SD rats after 10-day CRS exposure (I2 = 14.20%). However, high heterogeneity was observed in the other groups. In SD rats, heterogeneity in the single group with 3-week exposure (I2 = 86.5%) was higher than that of the group with 2-week exposure (I2 = 71.3%). For C57BL/6J mice, longer exposure protocols resulted in decreased heterogeneity (1-week: I2 = 97.10%; 2-week: I2 = 91.70%; 3-week: I2 = 88.80%; and 4-week: I2 = 84.50%).
Discussion
The CRS model is widely used to recapitulate depression features due to its relative simplicity. However, it is frequently criticized for its perceived lack of efficacy. We performed a meta-analysis of studies that used CRS protocols to evaluate anhedonic-like behavior in rodents. As the primary endpoint of this study, we attempted to identify strain-dependent susceptibilities to CRS on the basis of a core symptom of depression, anhedonia.
CRS is one of the most extensively used stress paradigms in laboratory animals to model human psychological stress. CRS protocols are simple and require less time, cost, and labor than CUMS. This study evaluated the validity of CRS in rodent models by analyzing effects on anhedonic-like behavior. After undergoing CRS for at least 1 week, there was decreased responsiveness to sucrose consumption analogous to anhedonia, the core symptom of MDD. However, there were methodological differences between the CRS protocols, including in restraint conditions, duration, and handling.
A comparative study demonstrated that increasingly severe movement restrictions led to greater behavioral stress responses. 49 Our pooled analysis of SPT results confirmed that duration of CRS exposure contributed to anhedonia-like behavioral responses, especially in C57BL/6J mice. Other differences in experimental procedures, including light/dark phase, water and food deprivation, presence of a foreign object, and novel noises and odors in the housing may simultaneity function as the stimuli, thereby potentially altering endocrine physiology and the development of depressive-like behaviors.
Rodents naturally avidly consume sweet foods and selectively drink sweet liquids when given a free choice of two bottles with separate access to sucrose solution and regular water.55,56 Sucrose preference is a valid read-out of hedonic behavior, and a reduced sucrose preference ratio in stressed animals relative to controls is indicative of anhedonia. 56 Some studies have measured absolute sucrose consumption as a measure of anhedonia; 57 however, it is unclear how this measure affects reliability. First, the intake volume of sucrose solution can fluctuate considerably in rodents due to weight differences in experimental animals. Second, in some cases the rodents consume a decreased volume of liquid including sucrose solution and regular water. Occasionally, they consume large amounts of both liquids. 56 Thus, in our meta-analysis, we chose to use the sucrose preference ratio rather than the absolute sucrose consumption as the indicative measure of SPT. Sucrose preference ratio is a widely accepted parameter for anhedonia-like behavior in depressive rodents.
The test designs differed between the included studies, including test onset time, training protocols, water and food deprivation period, sucrose concentrations, and testing period. According to the recommendations of previous studies in the field of depression, a 1% to 2% (weight/volume) sucrose solution is the optimal concentration to elicit a preference over water. Some of the included studies ignored habituation to sucrose solution and two-bottle conditioning and did not conduct baseline measurements. Food and water deprivation prior to SPT can act as an additional stressor that affects anhedonic behavioral response. It is notable that the time chosen for SPTs is also important because circadian rhythms influence drinking behavior. Accordingly, it is appropriate to adopt a standard protocol for SPT to estimate anhedonic phenotypes in depression models.
Although there was decreased sucrose preference in the stressed groups compared with controls, the duration of CRS can differentially affect anhedonic-like behavior in model animals. Experimental animals present different degrees of decreased sucrose preference (%) depending on CRS duration. For example, sucrose intake tended to decrease in C57BL/6J mice over exposure durations from 1 to 4 weeks. Publication bias was assessed in different rodent species/strains by funnel plot (Figure 6), which indicated marginal effects of publication bias that were mostly attributable to small sample sizes and insufficient reporting of negative data. The trim and fill method allows estimates of an adjusted meta-analysis in the presence of publication bias; 58 thus, we performed a trim and fill analysis of the included studies. The results indicated that the presence of publication bias did not greatly affect the pooled analysis of effect size.
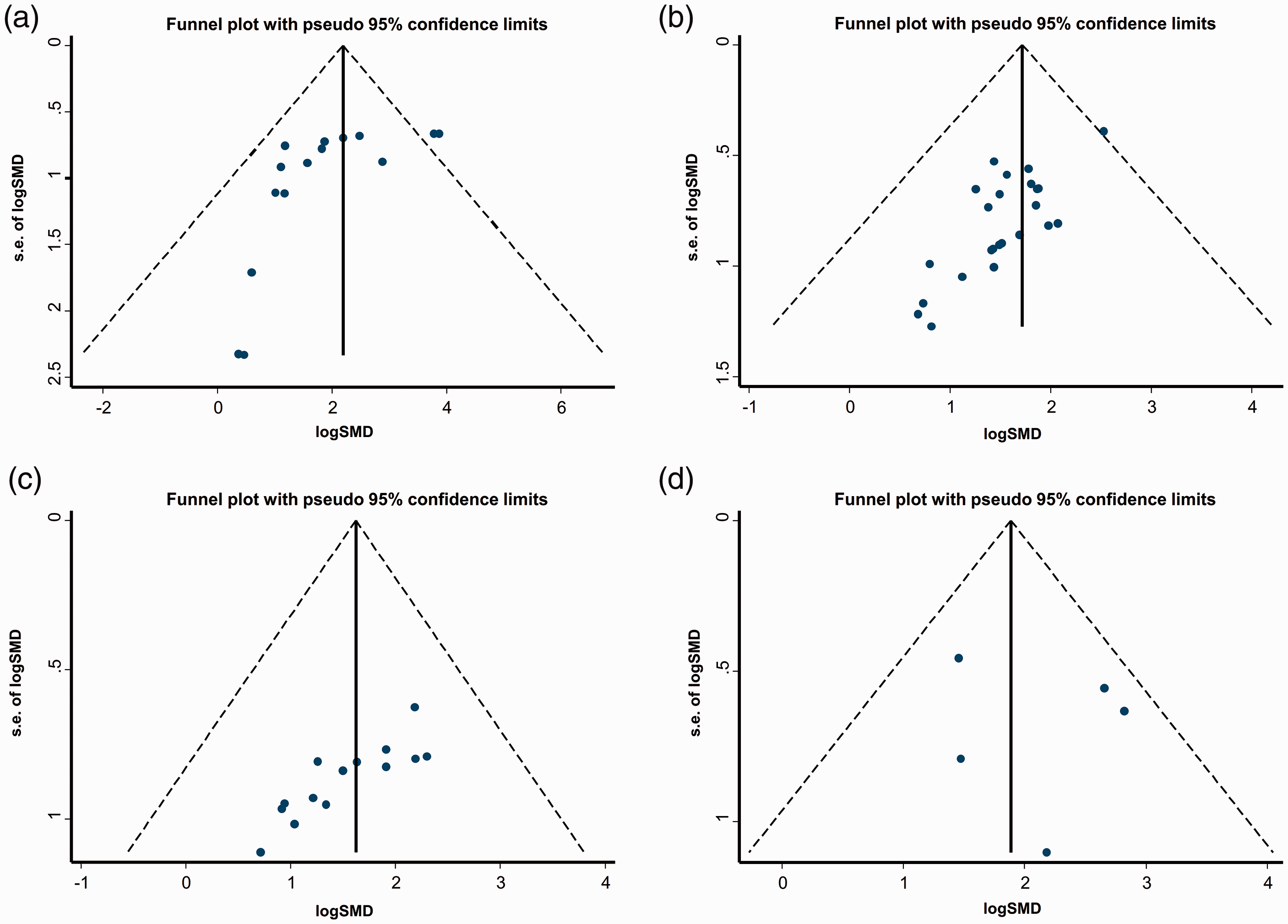
Funnel plots of the effect measure (log|SMD|) versus its precision (standard error [SE] of log|SMD|) of sucrose preference (%) for C57BL/6J mice (a), Sprague–Dawley rats (b), Wistar rats (c) and mice of other strains (d).
There was high heterogeneity among studies in the single-group analysis, which suggested difficulties in achieving reproducible effects of the CRS protocol by different research groups. Numerous factors can bring heterogeneity into the pooled results, including the animal strains, animal sex, CRS protocol (e.g., duration, intensity, and housing and restraint conditions), and SPT protocols (e.g., test onset time, training protocols, water and food deprivation period, sucrose concentrations, and testing period), which should be considered when designing CRS protocols for modeling anhedonic-like behavior. Additionally, circadian rhythm and restraint placement are important factors in CRS protocols that should not be overlooked. The restraint placement and time periods used in the included studies are summarized in Table 5. Most of the included studies performed CRS over a fixed daily time period to avoid circadian rhythm fluctuations. Experimental animals were periodically constrained from movement by placing them in tubes of suitable volumes depending on the animal species/strain.
The restraint placements and time periods used in the included studies.

NA: not available in the article.
The effectiveness of CRS is not confined to a particular strain/species of animal. Our pooled analysis demonstrated inter-species variability, with rats exhibiting greater susceptibility to restraint stress compared with mice. In terms of murine CRS-induced depression models, BALB/c mice were not commonly used. In 2020, Tsuchimine et al. conducted a comparison of the physiological and behavioral responses to CRS between C57BL/6J and BALB/c mice. 30 The results showed that BALB/c, but not C57BL/6J, mice presented anhedonia-like behavior after CRS according to SRT results, indicating a greater behavior stress response in BALB/c than in C57BL/6J mice.
Chronic stress results in a higher magnitude of corticosterone responses, and it has been reported that chronic administration of corticosterone to mice induces anhedonia-like behavior. 59 Consistently, human studies have shown that anhedonia symptoms are associated with higher corticosterone levels in patients with depression. 60 Inter-strain variability in the development of anhedonia-like behavior could be explained by differences in the functionality of the HPA axis. Another explanation for inter-strain variability is differences in the type of immune responses involved including Th1 and Th2 immunity, which may contribute to CRS susceptibility. 61
An overwhelming majority of the studies established CRS-induced depression models in male rodents, with only 3.5% of the studies (2/57) selecting female rodents as the research subjects. In 2012, Eiland et al. found a significant effect of sex in CRS-induced depression-like behavior, with females exhibiting greater locomotion than males under restraint stress. 29 A similar finding was reported following CRS, in that CRS did not induce distinguishable anhedonia-like behavior in female C57BL/6J mice, while other studies using male C57BL/6J mice reported a positive effect. 50
A growing literature suggests sexual dimorphisms in the endocrine and immune systems and in stress resilience. 2 These sex differences are likely attributable to steroid hormones, such as estrogens and androgens, which can modulate the effects of stress on dendritic remodeling and regulate susceptibility to stressful events.2,62 It was reported that in rats with heart failure induced by myocardial infarction, in contrast to males, females do not develop depression-like behavior or an increase in prefrontal cortex cytokines, and this discrepancy was attributed to the role of estrogens. 63 Thus, the sex of model animals should be considered when designing experiments.
In conclusion, this meta-analysis indicated that the CRS protocol is a reliable and effective rodent model of anhedonic-like behavior. However, there was high heterogeneity in the single subgroup analysis, which may be attributable to the duration and intensity of CRS and to SPF protocols. This work may provide a reference stress duration and intensity for CRS models in specific animal species/strains.
Footnotes
Author contributions
XY conceived and designed the analysis, solved disagreements during study selection and data extraction, and reviewed the manuscript; YM and YX selected the included studies and extracted data; YM analyzed the data, edited the figures and tables, and wrote the manuscript.
Declaration of conflicting interest
The authors declare that there is no conflict of interest.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the Youth Project of Science and Technology Research Project of Jiangxi Education Department (No. GJJ200251) and the Hunan Cancer Hospital Climb Plan (No. 20210225-1006).